
Clinical and Molecular Aspects Associated with Defects in the Transcription Factor POU3F4: A Review


Abstract
1. Introduction
2. X-Linked Hearing Loss
3. The Transcription Factor POU3F4
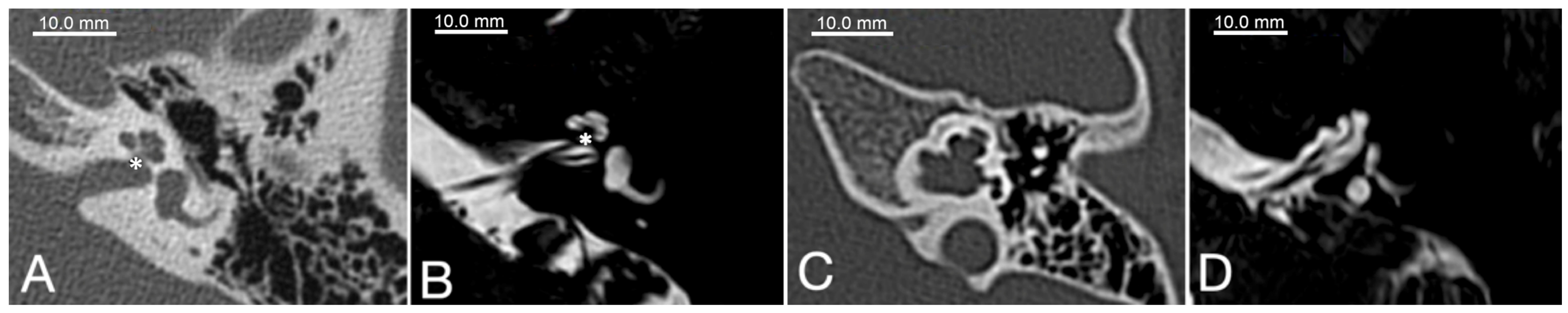
4. Clinical and Anatomical Features of POU3F4-Linked Hearing Loss
5. Structural Features of the POU Family of Transcription Factors
6. Transcriptional Targets of POU3F4
7. Variants of POU3F4 Identified in Hearing Loss Patients and Genotype-Phenotype Correlation
8. Hearing Loss Phenotypes Described in Female Carriers
9. Further Symptoms Described in POU3F4 Patients
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koffler, T.; Ushakov, K.; Avraham, K.B. Genetics of Hearing Loss: Syndromic. Otolaryngol. Clin. N. Am. 2015, 48, 1041–1061. [Google Scholar] [CrossRef]
- Pandya, A. Genetic hearing loss: The journey of discovery to destination—How close are we to therapy? Mol. Genet. Genom. Med. 2016, 4, 583–587. [Google Scholar] [CrossRef]
- Corvino, V.; Apisa, P.; Malesci, R.; Laria, C.; Auletta, G.; Franze, A. X-Linked Sensorineural Hearing Loss: A Literature Review. Curr. Genom. 2018, 19, 327–338. [Google Scholar] [CrossRef]
- Del Castillo, I.; Morín, M.; Domínguez-Ruiz, M.; Moreno-Pelayo, M.A. Genetic etiology of non-syndromic hearing loss in Europe. Hum. Genet. 2022, 141, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.F.; Hostikka, S.L.; Zhou, J.; Chow, L.T.; Oliphant, A.R.; Gerken, S.C.; Gregory, M.C.; Skolnick, M.H.; Atkin, C.L.; Tryggvason, K. Identification of Mutations in the COL4A5 Collagen Gene in Alport Syndrome. Science 1990, 248, 1224–1227. [Google Scholar] [CrossRef] [PubMed]
- Poretti, A.; Palla, A.; Tarnutzer, A.A.; Petersen, J.A.; Weber, K.P.; Straumann, D.; Jung, H.H. Vestibular impairment in patients with Charcot-Marie-Tooth disease. Neurology 2013, 80, 2099–2105. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Han, D.; Li, J.; Han, B.; Ouyang, X.; Cheng, J.; Li, X.; Jin, Z.; Wang, Y.; Bitner-Glindzicz, M.; et al. Loss-of-Function Mutations in the PRPS1 Gene Cause a Type of Nonsyndromic X-linked Sensorineural Deafness, DFN2. Am. J. Hum. Genet. 2010, 86, 65–71. [Google Scholar] [CrossRef]
- Huebner, A.K.; Gandia, M.; Frommolt, P.; Maak, A.; Wicklein, E.M.; Thiele, H.; Altmüller, J.; Wagner, F.; Viñuela, A.; Aguirre, L.A.; et al. Nonsense Mutations in SMPX, Encoding a Protein Responsive to Physical Force, Result in X-Chromosomal Hearing Loss. Am. J. Hum. Genet. 2011, 88, 621–627. [Google Scholar] [CrossRef]
- Wang, Q.J.; Li, Q.Z.; Rao, S.Q.; Lee, K.; Huang, X.S.; Yang, W.Y.; Zhai, S.Q.; Guo, W.W.; Guo, Y.F.; Yu, N.; et al. AUNX1, a novel locus responsible for X linked recessive auditory and peripheral neuropathy, maps to Xq23-27.3. J. Med. Genet. 2006, 43, e33. [Google Scholar] [CrossRef] [PubMed]
- Rost, S.; Bach, E.; Neuner, C.; Nanda, I.; Dysek, S.; Bittner, R.E.; Keller, A.; Bartsch, O.; Mlynski, R.; Haaf, T.; et al. Novel form of X-linked nonsyndromic hearing loss with cochlear malformation caused by a mutation in the type IV collagen gene COL4A6. Eur. J. Hum. Genet. 2014, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Cremers, C.W.; Snik, A.; Huygen, P.; Joosten, F.; Cremers, F. X-Linked Mixed Deafness Syndrome with Congenital Fixation of the Stapedial Footplate and Perilymphatic Gusher (DFN3). Adv. Otorhinolaryngol. 2002, 61, 161–167. [Google Scholar] [CrossRef] [PubMed]
- De Kok, Y.J.; van der Maarel, S.M.; Bitner-Glindzicz, M.; Huber, I.; Monaco, A.P.; Malcolm, S.; Pembrey, M.E.; Ropers, H.H.; Cremers, F.P. Association between X-linked mixed deafness and mutations in the POU domain gene POU3F4. Science 1995, 267, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.-X.; Gong, R.-Z.; Zhao, B. HRCT and MRI findings in X-linked non-syndromic deafness patients with a POU3F4 mutation. Int. J. Pediatr. Otorhinolaryngol. 2014, 78, 1756–1762. [Google Scholar] [CrossRef] [PubMed]
- Stankovic, K.M.; Hennessey, A.M.; Herrmann, B.; Mankarious, L.A. Cochlear Implantation in Children with Congenital X-Linked Deafness Due to Novel Mutations in POU3F4 Gene. Ann. Otol. Rhinol. Laryngol. 2010, 119, 815–822. [Google Scholar] [CrossRef]
- Sennaroğlu, L.; Bajin, M.D. Incomplete partition type III: A rare and difficult cochlear implant surgical indication. Auris Nasus Larynx 2018, 45, 26–32. [Google Scholar] [CrossRef]
- Samadi, D.S.; Saunders, J.C.; Crenshaw, E.B., 3rd. Mutation of the POU-domain gene Brn4/Pou3f4 affects middle-ear sound conduction in the mouse. Hear. Res. 2005, 199, 11–21. [Google Scholar] [CrossRef]
- Brooks, P.M.; Rose, K.P.; MacRae, M.; Rangoussis, K.M.; Gurjar, M.; Hertzano, R.; Coate, T.M. Pou3f4-expressing otic mesenchyme cells promote spiral ganglion neuron survival in the postnatal mouse cochlea. J. Comp. Neurol. 2020, 528, 1967–1985. [Google Scholar] [CrossRef]
- Kidokoro, Y.; Karasawa, K.; Minowa, O.; Sugitani, Y.; Noda, T.; Ikeda, K.; Kamiya, K. Deficiency of Transcription Factor Brn4 Disrupts Cochlear Gap Junction Plaques in a Model of DFN3 Non-Syndromic Deafness. PLoS ONE 2014, 9, e108216. [Google Scholar] [CrossRef]
- Barashkov, N.A.; Klarov, L.A.; Teryutin, F.M.; Solovyev, A.V.; Pshennikova, V.G.; Konnikova, E.E.; Romanov, G.P.; Tobokhov, A.V.; Morozov, I.V.; Bondar, A.A.; et al. A novel pathogenic variant c.975G>A (p.Trp325*) in the POU3F4 gene in Yakut family (Eastern Siberia, Russia) with the X-linked deafness-2 (DFNX2). Int. J. Pediatr. Otorhinolaryngol. 2018, 104, 94–97. [Google Scholar] [CrossRef]
- Marlin, S.; Moizard, M.; David, A.; Chaissang, N.; Raynaud, M.; Jonard, L.; Feldmann, D.; Loundon, N.; Denoyelle, F.; Toutain, A. Phenotype and genotype in females with POU3F4 mutations. Clin. Genet. 2009, 76, 558–563. [Google Scholar] [CrossRef]
- Sennaroglu, L.; Ozbal Batuk, M.; Bajin, M.D. Incomplete Partition Type III. In Inner Ear Malformations: Classification, Evaluation and Treatment; Sennaroglu, L., Ed.; Springer: International Publishing: Cham, Switzerland, 2022; pp. 271–282. [Google Scholar]
- Phelps, P.D.; Reardon, W.; Pembrey, M.; Bellman, S.; Luxom, L. X-linked deafness, stapes gushers and a distinctive defect of the inner ear. Neuroradiology 1991, 33, 326–330. [Google Scholar] [CrossRef]
- Sennaroğlu, L.; Bajin, M.D. Classification and Current Management of Inner Ear Malformations. Balk. Med. J. 2017, 34, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Roesch, S.; Rasp, G.; Sarikas, A.; Dossena, S. Genetic Determinants of Non-Syndromic Enlarged Vestibular Aqueduct: A Review. Audiol. Res. 2021, 11, 423–442. [Google Scholar] [CrossRef]
- Bernardinelli, E.; Roesch, S.; Simoni, E.; Marino, A.; Rasp, G.; Astolfi, L.; Sarikas, A.; Dossena, S. Novel POU3F4 variants identified in patients with inner ear malformations exhibit aberrant cellular distribution and lack of SLC6A20 transcriptional upregulation. Front. Mol. Neurosci. 2022, 15, 999833. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.-Q.; Zeng, J.-L.; Yuan, Y.-Y.; Dai, P. A novel mutation in POU3F4 in a Chinese family with X-linked non-syndromic hearing loss. J. Otol. 2015, 10, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Shearer, A.E.; Zhou, G.W.; Kenna, M.; Poe, D.; Licameli, G.R.; Brodsky, J.R. Peripheral Vestibular Dysfunction Is a Common Occurrence in Children with Non-syndromic and Syndromic Genetic Hearing Loss. Front. Neurol. 2021, 12, 714543. [Google Scholar] [CrossRef]
- Herr, W.; Sturm, R.A.; Clerc, R.G.; Corcoran, L.M.; Baltimore, D.; Sharp, P.A.; Ingraham, H.A.; Rosenfeld, M.G.; Finney, M.; Ruvkun, G. The POU domain: A large conserved region in the mammalian pit-1, oct-1, oct-2, and Caenorhabditis elegans unc-86 gene products. Genes Dev. 1988, 2, 1513–1516. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Wang, Y.; Zhao, P.; Wei, S.; Li, Z.; Chang, H.; He, H. Biochemical characterization and functional analysis of the POU transcription factor POU-M2 of Bombyx mori. Int. J. Biol. Macromol. 2016, 86, 701–708. [Google Scholar] [CrossRef]
- McGinnis, W.; Garber, R.L.; Wirz, J.; Kuroiwa, A.; Gehring, W.J. A homologous protein-coding sequence in drosophila homeotic genes and its conservation in other metazoans. Cell 1984, 37, 403–408. [Google Scholar] [CrossRef]
- Phillips, K.; Luisia, B. The virtuoso of versatility: POU proteins that flex to fit. J. Mol. Biol. 2000, 302, 1023–1039. [Google Scholar] [CrossRef]
- Otting, G.; Qian, Y.Q.; Billeter, M.; Müller, M.; Affolter, M.; Gehring, W.J.; Wüthrich, K. Protein-DNA contacts in the structure of a homeodomain-DNA complex determined by nuclear magnetic resonance spectroscopy in solution. EMBO J. 1990, 9, 3085–3092. [Google Scholar] [CrossRef]
- Assa-Munt, N.; Mortishire-Smith, R.J.; Aurora, R.; Herr, W.; Wright, P.E. The solution structure of the Oct-1 POU-specific domain reveals a striking similarity to the bacteriophage lambda repressor DNA-binding domain. Cell 1993, 73, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Engström, Y. Regulation of immune and tissue homeostasis by Drosophila POU factors. Insect Biochem. Mol. Biol. 2019, 109, 24–30. [Google Scholar] [CrossRef]
- Chang, Y.K.; Srivastava, Y.; Hu, C.; Joyce, A.; Yang, X.; Zuo, Z.; Havranek, J.J.; Stormo, G.D.; Jauch, R. Quantitative profiling of selective Sox/POU pairing on hundreds of sequences in parallel by Coop-seq. Nucleic Acids Res. 2016, 45, 832–845. [Google Scholar] [CrossRef]
- Malik, V.; Zimmer, D.; Jauch, R. Diversity among POU transcription factors in chromatin recognition and cell fate reprogramming. Cell. Mol. Life Sci. 2018, 75, 1587–1612. [Google Scholar] [CrossRef] [PubMed]
- Sturm, R.A.; Das, G.; Herr, W. The ubiquitous octamer-binding protein Oct-1 contains a POU domain with a homeo box subdomain. Genes Dev. 1988, 2, 1582–1599. [Google Scholar] [CrossRef] [PubMed]
- Ingraham, H.A.; Chen, R.; Mangalam, H.J.; Elsholtz, H.P.; Flynn, S.E.; Lin, C.R.; Simmons, D.M.; Swanson, L.; Rosenfeld, M.G. A tissue-specific transcription factor containing a homeodomain specifies a pituitary phenotype. Cell 1988, 55, 519–529. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Treacy, M.N.; Simmons, D.M.; Ingraham, H.A.; Swanson, L.W.; Rosenfeld, M.G. Expression of a large family of POU-domain regulatory genes in mammalian brain development. Nature 1989, 340, 35–42. [Google Scholar] [CrossRef]
- Mathis, J.M.; Simmons, D.; He, X.; Swanson, L.; Rosenfeld, M. Brain 4: A novel mammalian POU domain transcription factor exhibiting restricted brain-specific expression. EMBO J. 1992, 11, 2551–2561. [Google Scholar] [CrossRef]
- Scheidereit, C.; Cromlish, J.A.; Gerster, T.; Kawakami, K.; Balmaceda, C.-G.; Currie, R.A.; Roeder, R.G. A human lymphoid- specific transcription factor that activates immunoglobulin genes is a homoeobox protein. Nature 1988, 336, 551–557. [Google Scholar] [CrossRef]
- Andersen, B.; Weinberg, W.C.; Rennekampff, O.; McEvilly, R.J.; Bermingham, J.R.; Hooshmand, F.; Vasilyev, V.; Hansbrough, J.F.; Pittelkow, M.R.; Yuspa, S.H.; et al. Functions of the POU domain genes Skn-1a/i and Tst-1/Oct-6/SCIP in epidermal differentiation. Genes Dev. 1997, 11, 1873–1884. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Yoshioka, T.; Nathans, J. Retina-derived POU-domain factor-1: A complex POU-domain gene implicated in the development of retinal ganglion and amacrine cells. J. Neurosci. 1996, 16, 2261–2274. [Google Scholar] [CrossRef] [PubMed]
- Phippard, D.; Heydemann, A.; Lechner, M.; Lu, L.; Lee, D.; Kyin, T.; Crenshaw, E. Changes in the subcellular localization of the Brn4 gene product precede mesenchymal remodeling of the otic capsule. Hear. Res. 1998, 120, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Ye, M.; Feng, R.; Graf, T. Stepwise Reprogramming of B Cells into Macrophages. Cell 2004, 117, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Bar-Nur, O.; Verheul, C.; Sommer, A.G.; Brumbaugh, J.; Schwarz, B.A.; Lipchina, I.; Huebner, A.J.; Mostoslavsky, G.; Hochedlinger, K. Lineage conversion induced by pluripotency factors involves transient passage through an iPSC stage. Nat. Biotechnol. 2015, 33, 761–768. [Google Scholar] [CrossRef]
- Choi, B.Y.; Kim, D.-H.; Chung, T.; Chang, M.; Kim, E.-H.; Kim, A.R.; Seok, J.; Chang, S.O.; Bok, J.; Kim, D.; et al. Destabilization and Mislocalization of POU3F4 by C-Terminal Frameshift Truncation and Extension Mutation. Hum. Mutat. 2012, 34, 309–316. [Google Scholar] [CrossRef]
- Lee, H.K.; Song, M.H.; Kang, M.; Lee, J.T.; Kong, K.-A.; Choi, S.-J.; Lee, K.Y.; Venselaar, H.; Vriend, G.; Lee, W.-S.; et al. Clinical and molecular characterizations of novel POU3F4 mutations reveal that DFN3 is due to null function of POU3F4 protein. Physiol. Genom. 2009, 39, 195–201. [Google Scholar] [CrossRef]
- Parzefall, T.; Shivatzki, S.; Lenz, D.R.; Rathkolb, B.; Ushakov, K.; Karfunkel, D.; Shapira, Y.; Wolf, M.; Mohr, M.; Wolf, E.; et al. Cytoplasmic Mislocalization of POU3F4 Due to Novel Mutations Leads to Deafness in Humans and Mice. Hum. Mutat. 2013, 34, 1102–1110. [Google Scholar] [CrossRef]
- Okazawa, H.; Imafuku, I.; Minowa, M.T.; Kanazawa, I.; Hamada, H.; Mouradian, M.M. Regulation of striatal D1A dopamine receptor gene transcription by Brn-4. Proc. Natl. Acad. Sci. USA 1996, 93, 11933–11938. [Google Scholar] [CrossRef] [PubMed]
- Coate, T.M.; Raft, S.; Zhao, X.; Ryan, A.K.; Crenshaw, E.B.; Kelley, M.W. Otic Mesenchyme Cells Regulate Spiral Ganglion Axon Fasciculation through a Pou3f4/EphA4 Signaling Pathway. Neuron 2012, 73, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Minowa, O.; Ikeda, K.; Sugitani, Y.; Oshima, T.; Nakai, S.; Katori, Y.; Suzuki, M.; Furukawa, M.; Kawase, T.; Zheng, Y.; et al. Altered Cochlear Fibrocytes in a Mouse Model of DFN3 Nonsyndromic Deafness. Science 1999, 285, 1408–1411. [Google Scholar] [CrossRef]
- Gao, J.; Chen, Y.; Yang, Y.; Liang, J.; Xie, J.; Liu, J.; Li, S.; Zheng, G.; Xie, L.; Zhang, R. The transcription factor Pf-POU3F4 regulates expression of the matrix protein genes Aspein and Prismalin-14 in pearl oyster (Pinctada fucata). FEBS J. 2016, 283, 1962–1978. [Google Scholar] [CrossRef] [PubMed]
- Raft, S.; Coate, T.M.; Kelley, M.W.; Crenshaw, E.B.; Wu, D.K. Pou3f4-Mediated Regulation of Ephrin-B2 Controls Temporal Bone Development in the Mouse. PLoS ONE 2014, 9, e109043. [Google Scholar] [CrossRef]
- Inoue, F.; Kurokawa, D.; Takahashi, M.; Aizawa, S. Gbx2 Directly Restricts Otx2 Expression to Forebrain and Midbrain, Competing with Class III POU Factors. Mol. Cell. Biol. 2012, 32, 2618–2627. [Google Scholar] [CrossRef]
- Malik, K.F.; Kim, J.; Hartman, A.; Kim, P.; Young, W.S. Binding preferences of the POU domain protein Brain-4: Implications for autoregulation. Mol. Brain Res. 1996, 38, 209–221. [Google Scholar] [CrossRef]
- De Kok, Y.J.; Vossenaar, E.R.; Cremers, C.W.R.J.; Dahl, N.; Laporte, J.; Hu, L.J.; Lacombe, D.; Fischel-Ghodsian, N.; Friedman, R.A.; Parnes, L.S.; et al. Identification of a hot spot for microdeletions in patients with X- linked deafness type 3 (DFN3) 900 kb proximal to the DFN3 gene POU3F4. Hum. Mol. Genet. 1996, 5, 1229–1235. [Google Scholar] [CrossRef]
- Bitner-Glindzicz, M.; Turnpenny, P.; Höglund, P.; Kääriäinen, H.; Sankila, E.-M.; van der Maarel, S.M.; de Kok, Y.J.; Ropers, H.-H.; Cremers, F.P.; Pembrey, M.; et al. Further mutations in Brain 4 (POU3F4) clarify the phenotype in the X-linked deafness, DFN3. Hum. Mol. Genet. 1995, 4, 1467–1469. [Google Scholar] [CrossRef][Green Version]
- Oza, A.M.; DiStefano, M.T.; Hemphill, S.E.; Cushman, B.J.; Grant, A.R.; Siegert, R.K.; Shen, J.; Chapin, A.; Boczek, N.J.; Schimmenti, L.A.; et al. Expert specification of the ACMG/AMP variant interpretation guidelines for genetic hearing loss. Hum. Mutat. 2018, 39, 1593–1613. [Google Scholar] [CrossRef]
- Wu, J.; Cao, Z.; Su, Y.; Wang, Y.; Cai, R.; Chen, J.; Gao, B.; Han, M.; Li, X.; Zhang, D.; et al. Molecular diagnose of a large hearing loss population from China by targeted genome sequencing. J. Hum. Genet. 2022, 67, 643–649. [Google Scholar] [CrossRef]
- Wu, H.-M.; Jie, H.-Q.; Wang, H.; Wu, Y.-Q.; Chen, Z.-N.; Xing, Y.-Z.; Wang, J.-P.; Shi, H.-B.; Yin, S.-K. A novel POU domain class 3 transcription factor 4 mutation causes X-linked non-syndromic hearing loss in a Chinese family. Chin. Med. J. 2019, 132, 2251–2253. [Google Scholar] [CrossRef] [PubMed]
- Pollak, A.; Lechowicz, U.; Kędra, A.; Stawiński, P.; Rydzanicz, M.; Furmanek, M.; Brzozowska, M.; Mrówka, M.; Skarżyński, H.; Skarżyński, P.H.; et al. Novel and De Novo Mutations Extend Association of POU3F4 with Distinct Clinical and Radiological Phenotype of Hearing Loss. PLoS ONE 2016, 11, e0166618. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Liu, S.; Wang, H.; Ding, Y.; Chen, Y.; Liu, H. Genetic etiology study of four Chinese families with two nonsyndromic deaf children in succession by targeted next-generation sequencing. Mol. Genet. Genom. Med. 2021, 9, e1634. [Google Scholar] [CrossRef]
- Chen, Y.; Qiu, J.; Wu, Y.; Jia, H.; Jiang, Y.; Jiang, M.; Wang, Z.; Sheng, H.-B.; Hu, L.; Zhang, Z.; et al. Genetic findings of Sanger and nanopore single-molecule sequencing in patients with X-linked hearing loss and incomplete partition type III. Orphanet J. Rare Dis. 2022, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Kanno, A.; Mutai, H.; Namba, K.; Morita, N.; Nakano, A.; Ogahara, N.; Sugiuchi, T.; Ogawa, K.; Matsunaga, T. Frequency and specific characteristics of the incomplete partition type III anomaly in children. Laryngoscope 2016, 127, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- Waryah, A.M.; Ahmed, Z.M.; Binder, M.A.; Choo, D.I.; Sisk, R.A.; Shahzad, M.; Khan, S.N.; Friedman, T.B.; Riazuddin, S.; Riazuddin, S. Molecular and clinical studies of X-linked deafness among Pakistani families. J. Hum. Genet. 2011, 56, 534–540. [Google Scholar] [CrossRef]
- Jiang, Y.; Wu, L.; Huang, S.; Li, P.; Gao, B.; Yuan, Y.; Zhang, S.; Yu, G.; Gao, Y.; Wu, H.; et al. Study of complex structural variations of X-linked deafness-2 based on single-molecule sequencing. Biosci. Rep. 2021, 41. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.; Lee, K.; Lim, E.; Choi, S.; Park, R.; Kim, U. Novel POU3F4 mutations and clinical features of DFN3 patients with cochlear implants. Clin. Genet. 2009, 75, 572–575. [Google Scholar] [CrossRef]
- Chao, X.; Xiao, Y.; Zhang, F.; Luo, J.; Wang, R.; Liu, W.; Wang, H.; Xu, L. Cochlear Implantation in a Patient with a Novel POU3F4 Mutation and Incomplete Partition Type-III Malformation. Neural Plast. 2020, 2020, 8829587. [Google Scholar] [CrossRef]
- Jo, H.D.; Han, J.H.; Lee, S.M.; Choi, D.H.; Lee, S.-Y.; Choi, B.Y. Genetic Load of Alternations of Transcription Factor Genes in Non-Syndromic Deafness and the Associated Clinical Phenotypes: Experience from Two Tertiary Referral Centers. Biomedicines 2022, 10, 2125. [Google Scholar] [CrossRef]
- Hagiwara, H.; Tamagawa, Y.; Kitamura, K.; Kodera, K. A new mutation in the POU3F4 gene in a japanese family with x-linked mixed deafness (DFN3). Laryngoscope 1998, 108, 1544–1547. [Google Scholar] [CrossRef]
- Han, J.J.; Nguyen, P.D.; Oh, D.-Y.; Han, J.H.; Kim, A.-R.; Kim, M.Y.; Park, H.-R.; Tran, L.H.; Dung, N.H.; Koo, J.-W.; et al. Elucidation of the unique mutation spectrum of severe hearing loss in a Vietnamese pediatric population. Sci. Rep. 2019, 9, 1604. [Google Scholar] [CrossRef] [PubMed]
- Vore, A.P.; Chang, E.H.; Hoppe, J.E.; Butler, M.G.; Forrester, S.; Schneider, M.C.; Smith, L.L.H.; Burke, D.W.; Campbell, C.A.; Smith, R.J.H. Deletion of and Novel Missense Mutation in POU3F4 in 2 Families Segregating X-Linked Nonsyndromic Deafness. Arch. Otolaryngol. Head Neck Surg. 2005, 131, 1057–1063. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, J.; Cheng, J.; Lu, Y.; Lu, Y.; Chen, A.; Sun, Y.; Kang, D.; Zhang, X.; Dai, P.; Han, D.; et al. Identification of a novel mutation in POU3F4 for prenatal diagnosis in a Chinese family with X-linked nonsyndromic hearing loss. J. Genet. Genom. 2010, 37, 787–793. [Google Scholar] [CrossRef]
- Su, Y.; Gao, X.; Huang, S.-S.; Mao, J.-N.; Huang, B.-Q.; Zhao, J.-D.; Kang, D.-Y.; Zhang, X.; Dai, P. Clinical and molecular characterization of POU3F4 mutations in multiple DFNX2 Chinese families. BMC Med. Genet. 2018, 19, 157. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, R.; Diñeiro, M.; Cifuentes, G.A.; Castillo, D.; Pruneda, P.C.; Álvarez, R.; Sánchez-Durán, N.; Capín, R.; Plasencia, A.; Viejo-Díaz, M.; et al. Comprehensive genomic diagnosis of non-syndromic and syndromic hereditary hearing loss in Spanish patients. BMC Med. Genom. 2018, 11, 58. [Google Scholar] [CrossRef]
- Wu, D.; Huang, W.; Xu, Z.; Li, S.; Zhang, J.; Chen, X.; Tang, Y.; Qiu, J.; Wang, Z.; Duan, X.; et al. Clinical and genetic study of 12 Chinese Han families with nonsyndromic deafness. Mol. Genet. Genom. Med. 2020, 8, e1177. [Google Scholar] [CrossRef]
- Mei, X.; Zhou, Y.; Amjad, M.; Yang, W.; Zhu, R.; Asif, M.; Hussain, H.M.J.; Yang, T.; Iqbal, F.; Hu, H. Next-Generation Sequencing Identifies Pathogenic Variants in HGF, POU3F4, TECTA, and MYO7A in Consanguineous Pakistani Deaf Families. Neural Plast. 2021, 2021, 5528434. [Google Scholar] [CrossRef]
- Bademci, G.; Lasisi, A.O.; Yariz, K.O.; Montenegro, P.; Menendez, I.; Vinueza, R.; Paredes, R.; Moreta, G.; Subasioglu, A.; Blanton, S.; et al. Novel domain-specific POU3F4 mutations are associated with X-linked deafness: Examples from different populations. BMC Med. Genet. 2015, 16, 9. [Google Scholar] [CrossRef]
- Moteki, H.; Shearer, A.E.; Izumi, S.; Kubota, Y.; Azaiez, H.; Booth, K.T.; Sloan, C.M.; Kolbe, D.L.; Smith, R.J.H.; Usami, S.-I. De Novo Mutation in X-Linked Hearing Loss–Associated POU3F4 in a Sporadic Case of Congenital Hearing Loss. Ann. Otol. Rhinol. Laryngol. 2015, 124 (Suppl. S1), 169S–176S. [Google Scholar] [CrossRef]
- Giannantonio, S.; Agolini, E.; Scorpecci, A.; Anzivino, R.; Bellacchio, E.; Cocciadiferro, D.; Novelli, A.; Digilio, M.C.; Marsella, P. Genetic identification and molecular modeling characterization of a novel POU3F4 variant in two Italian deaf brothers. Int. J. Pediatr. Otorhinolaryngol. 2020, 129, 109790. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Huang, S.; An, L.; Zhang, C.; Dai, P.; Gao, H.; Ma, X. Variant analysis of 92 Chinese Han families with hearing loss. BMC Med. Genom. 2022, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-J.; Li, Q.-Z.; Rao, S.-Q.; Zhao, Y.-L.; Yuan, H.; Yang, W.-Y.; Han, D.-Y.; Shen, Y. A Novel Mutation of POU3F4 Causes Congenital Profound Sensorineural Hearing Loss in a Large Chinese Family. Laryngoscope 2006, 116, 944–950. [Google Scholar] [CrossRef] [PubMed]
- Tekin, A.M.; Matulic, M.; Wuyts, W.; Assadi, M.Z.; Mertens, G.; van Rompaey, V.; Li, Y.; van de Heyning, P.; Topsakal, V. A New Pathogenic Variant in POU3F4 Causing Deafness Due to an Incomplete Partition of the Cochlea Paved the Way for Innovative Surgery. Genes 2021, 12, 613. [Google Scholar] [CrossRef]
- Wu, C.-C.; Lin, Y.-H.; Liu, T.-C.; Lin, K.-N.; Yang, W.-S.; Hsu, C.-J.; Chen, P.-L.; Wu, C.-M. Identifying Children with Poor Cochlear Implantation Outcomes Using Massively Parallel Sequencing. Medicine 2015, 94, e1073. [Google Scholar] [CrossRef]
- De Kok, Y.J.; Cremers, C.W.; Ropers, H.H.; Cremers, F.P. The molecular basis of X-linked deafness type 3 (DFN3) in two sporadic cases: Identification of a somatic mosaicism for a POU3F4 missense mutation. Hum. Mutat. 1997, 10, 207–211. [Google Scholar] [CrossRef]
- Schild, C.; Prera, E.; Lüblinghoff, N.; Arndt, S.; Aschendorff, A.; Birkenhäger, R. Novel Mutation in the Homeobox Domain of Transcription Factor POU3F4 Associated with Profound Sensorineural Hearing Loss. Otol. Neurotol. 2011, 32, 690–694. [Google Scholar] [CrossRef]
- Roesch, S.; Bernardinelli, E.; Nofziger, C.; Tóth, M.; Patsch, W.; Rasp, G.; Paulmichl, M.; Dossena, S. Functional Testing of SLC26A4 Variants—Clinical and Molecular Analysis of a Cohort with Enlarged Vestibular Aqueduct from Austria. Int. J. Mol. Sci. 2018, 19, 209. [Google Scholar] [CrossRef]
- Vijayasekaran, S.; Halsted, M.; Boston, M.; Meinzen-Derr, J.; Bardo, D.; Greinwald, J.; Benton, C. When Is the Vestibular Aqueduct Enlarged? A Statistical Analysis of the Normative Distribution of Vestibular Aqueduct Size. Am. J. Neuroradiol. 2007, 28, 1133–1138. [Google Scholar] [CrossRef]
- Song, M.H.; Lee, H.K.; Choi, J.Y.; Kim, S.; Bok, J.; Kim, U.-K. Clinical evaluation of DFN3 patients with deletions in the POU3F4 locus and detection of carrier female using MLPA. Clin. Genet. 2010, 78, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Luo, J.; Yu, H.; Rattner, A.; Mo, A.; Wang, Y.; Smallwood, P.M.; Erlanger, B.; Wheelan, S.J.; Nathans, J. Cellular Resolution Maps of X Chromosome Inactivation: Implications for Neural Development, Function, and Disease. Neuron 2014, 81, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Smeds, H.; Wales, J.; Karltorp, E.; Anderlid, B.-M.; Henricson, C.; Asp, F.; Anmyr, L.; Lagerstedt-Robinson, K.; Löfkvist, U. X-linked Malformation Deafness: Neurodevelopmental Symptoms Are Common in Children with IP3 Malformation and Mutation in POU3F4. Ear Hear. 2021, 43, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Green, T.; Shrestha, S.B.; Chromik, L.C.; Rutledge, K.; Pennington, B.F.; Hong, D.S.; Reiss, A.L. Elucidating X chromosome influences on Attention Deficit Hyperactivity Disorder and executive function. J. Psychiatr. Res. 2015, 68, 217–225. [Google Scholar] [CrossRef]
- Tian, H.; Wang, L.; Gao, F.; Liang, W.; Peng, K.A. Cochlear implantation using a custom guide catheter in 14 patients with incomplete partition type III. Clin. Otolaryngol. 2018, 43, 1379–1383. [Google Scholar] [CrossRef]
- Smeds, H.; Wales, J.; Asp, F.; Löfkvist, U.; Falahat, B.; Anderlid, B.-M.; Anmyr, L.; Karltorp, E. X-linked Malformation and Cochlear Implantation. Otol. Neurotol. 2017, 38, 38–46. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sequence Information | Clinical Information | Pathogenicity | Ref. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Amino Acid Change | Nucleotide Change | Sequencing Technique | Domain | Ethnicity | Hearing Loss | Malformations | Other Findings | ClinVar | |||
| Severity | Type | ||||||||||
| Ser22Cysfs*19 | 66del | NGS Panel | N-term* | Chinese | NA | NA | NA | - | 1185600 | P PVS1+PS2+PM2 | [62] C |
| Gln26∗ | 76C > T | WES | N-term* | Chinese | Severe | NA | Hypoplastic inferior gyrus of cochlea, absence of spiral plate of inferior gyrus | - | P PVS1+PP1+PP4 | [63] S | |
| Gln27* | 79C > T | WES | N-term* | NA | Profound | SNHL | IP-III | - | P PVS1+PP1+PP4 | [64] C | |
| Val46= | 138T > C | Sanger | N-term | Chinese | Profound | SNHL | EVA, Mondini | - | LB BP7 | [65] S | |
| Trp57* | 171G > A | Sanger | N-term* | Chinese | Profound | NA | IP-III | Developmental retardation | P PVS1+PS2+PP4 | [66] C | |
| Ser74Alafs*8 | c.220delA | Sanger | N-term* | NA | Severe | Mixed | EVA, IP-III | - | 1701827 | P PVS1+PS3+PP1 | [25] S |
| Gln79* | 235C > T | NGS Panel | N-term* | Israeli | Profound | SNHL | IP-II (Mondini) | - | 236062 | LP PVS1+PS3 | [51] S |
| Arg85Alafs*17 | 249delC | Sanger | N-term* | Japanese | Profound | Mixed | EVA, IP-III | Mental issues, hemiparalysis | LP PVS1+PP1+PP4 | [67] C | |
| Ser98* | NA | Sanger | N-term* | NA | Mild | Mixed | Bilateral dilatation of internal auditory canals | - | LP PVS1+PP1 | [20] C | |
| Trp114* | 341G > A | Sanger | N-term* | Pakistani | Severe to profound | Mixed | IP-III# | - | 43346 | P PVS1+PM2+PP1+PP4 | [68] S |
| Ala116fs* | 346delG | Sanger | N-term* | Korean | Severe | NA | IP-III# | - | P PVS1+PP1+PP4 | [50] S | |
| Ala116Glyfs*77 | 346dup | NGS Panel | N-term* | Chinese | NA | NA | IP-III | - | LP PVS1+PM2+PP4 | [69] C | |
| Ser117Argfs*26 | 346_350dup | NGS Panel | N-term* | Chinese | NA | SNHL | IP-III | - | LP PVS1+PM2+PP4 | [69] C | |
| Gly128fs* | 383delG | Sanger | N-term* | Korean | Profound | NA | IP-III# | - | P PVS1+PP1+PP4 | [70] S | |
| Gln136* | 406C > T | Sanger | N-term* | Pakistani | Severe to profound | Mixed | Mid and apical turns not clearly partitioned wide modiolus | - | P PVS1+PM2+PP1+PP4 | [68] S | |
| Gln136Leufs*58 | 400_401insACTC | WES | N-term* | Chinese | Profound | SNHL | IP-III | - | P PVS1+PP1+PP4 | [71] S | |
| Gln136Leufs*58 | 401_404dup | NGS Panel | N-term* | Chinese | NA | SNHL | IP-III | - | LP PVS1+PM2+PP4 | [69] C | |
| Val141* | 421_422delinsTA | NGS Panel | N-term* | Chinese | NA | Mixed | IP-III | - | LP PVS1+PM2+PP4 | [69] C | |
| His147Glnfs*94 | 441del | NGS Panel | N-term* | Chinese | NA | SNHL | IP-III | - | LP PVS1+PM2+PP4 | [69] C | |
| Pro153Leufs*88 | c.458delC | WES | N-term* | Korean | Profound | Mixed | IP-III | - | LP PVS1+PM2+PP3+PP4 | [72] C | |
| Pro164Argfs*77 | 491delC | Sanger | N-term* | Japanese | Moderate | Mixed | EVA, IP-III | - | LP PVS1+PP1+PP4 | [67] C | |
| Arg167* | 499C > T | Sanger | N-term* | NA | Severe to profound | Mixed | IP-III#, dysplasia of vestibule and semicircular canals | Attention disorders, learning delays, motor delays | 43347 | LP PVS1+PP1+PP4 | [14] S |
| Ser177* | 530C > A | Sanger | N-term* | Chinese | Profound | SNHL | IP-III, vestibule dysplasia | - | 988337 | LP PVS1+PP1+PP4 | [26] S |
| Gln181* | 541C > T | Sanger | N-term* | Chinese | Severe | NA | IP-III | - | LP PVS1+PP1+PP4 | [66] C | |
| Glu187* | 559G > T | WES | N-term* | Caucasian | Severe | SNHL | IP-III | - | P PVS1+PM2+PP4 | [64] C | |
| Phe201/Lys202del | 601–606del TTCAAA | Sanger | POUS NLS | Japanese | Severe | Mixed | IP-III# | - | LP PP1+PM1+PP4 | [73] S | |
| Lys202Glu | 604A > G | WES | POUS NLS | Vietnam | NA | NA | NA | - | VUS PM1+PM2 | [74] C | |
| Gln203fs* | 607_610delCAAA | Sanger | POUS* NLS | NA | NA | SNHL | NA | Vestibular dysfunction | 228391 | P PVS1+PM1 | [27] C |
| Arg204Lysfs*21 | 610del | NGS Panel | POUS NLS | NA | Severe | NA | NA | - | 1185660 | P PVS1+PM1+PM2 | [62] C |
| Arg205del | 614_616delGAA | Sanger | POUS NLS | Chinese | Severe | NA | IP-III | - | LP PM1+PM2+PP4 | [66] C | |
| Leu208* | 623T > A | Sanger | POUS* NLS | Chinese | Profound | IP-III# | Mild mental retardation | P PVS1+PP1+PP4 | [70] S | ||
| Ser228Leu | NA | Sanger | POUS | Korean | Profound | SNHL | IP-III# | Delayed motor development | LP PM1+PP1+PP4 | [75] S | |
| Gln229Arg | c.626A>G | WES | POUS | NA | Profound | Mixed | IP-III | - | LP PM1+PP1+PP4 | [72] C | |
| Thr211Met | 632C > T | Sanger | POUS | Korean | Moderate | NA | IP-III | - | LP PM3+PS3+PP4 | [49] S | |
| Val215Gly | 644T > G | Sanger | POUS | Chinese | Severe | NA | IP-III | - | LP PM1+PM2+PP1+PP4 | [66] C | |
| Gly216Glu | 647G > A | Sanger | POUS | Chinese | Profound (males)/mild to moderate (females) | Mixed (males)/SNHL (females) | IP-III# | - | 422492 | LP PM1+PM2+PP1+PP4 | [76] S |
| Leu217Valfs*9 | 648dupG | Sanger | POUS* | Chinese | Profound | NA | IP-III | - | P PVS1+PM1+PP4 | [66] C | |
| Leu217* | 650T > A | WES | POUS* | Caucasian | Profound | SNHL | IP-III | - | P PVS1+PM1+PM2+PP4 | [64] C | |
| Tyr223* | 669 T > A | Sanger | POUS* | Chinese | Profound | SNHL | IP-III | - | 1185618 | P PVS1+PM1+PM2+PP4 | [77] C |
| Gln229Arg | 686A > G | Sanger | POUS | Korean | Profound | NA | IP-III | - | 1879006 | LP PM1+PS3+PP1+PP4 | [49] S |
| Thr231Ile | 692C > T | NGS Panel | POUS | Caucasian | NA | SNHL | NA | - | LP PM1+PM2+PP3 | [78] C | |
| Cys233* | 699C > A | WES | POUS* | Chinese | Severe to Profound | SNHL | IP-III | - | P PVS1+PM1+PP1+PP4 | [79] C | |
| Glu236Asp | NA | Sanger | POUS | NA | Normal | NA | Normal | - | NA PM1+PP1 | [20] C | |
| Glu236Lys | 706G > A | NGS Panel | POUS | Pakistani | Profound | NA | - | - | LP PM1+PM2+PP1 | [80] S | |
| Glu236Ala | 707A > C | Sanger | POUS | Mixed | Profound | SNHL | IP-III# | - | P PM1+PM2+PP1+PP4 | [81] S | |
| Asn244Lysfs*26 | 727_728insA | NGS Panel | POUS* | Japanese | Severe | SNHL | IP-III | - | P PVS1+PM1+PP4 | [82] C | |
| Glu258Argfs*30 | 772delG | Sanger | POUS* | Mixed | Moderate to severe | SNHL | IP-III | - | P PVS1+PM1+PM2+PP1+PP4 | [81] S | |
| Gln275* | 823C > T | WES | Linker* | Caucasian | Severe | SNHL | IP-III | - | P PVS1+PM1+PP1+PP4 | [64] C | |
| Arg282Gln | 845G > A | Sanger | POUH NLS | Chinese | Profound | NA | IP-III | Cataract | LP PM1+PM2+PP1+PP4 | [66] C | |
| Ile285Asn | NA | Sanger | POUH | NA | Normal | NA | NA | - | NA PM1+PP1 | [20] C | |
| Ile285Argfs*43 | 853_854del | NGS Panel | POUH* | Israeli | Profound (male)/mild (female) | NA | IP-II (Mondini) | - | 43352 | P PVS1+PM1+PP1 | [51] S |
| Ser288Cysfs*40 | NA | Sanger | POUH* | NA | Normal | NA | NA | - | NA PVS1+PM1+PP1 | [20] C | |
| Val289fs | 862del4 | Sanger | POUH* | Caucasian | Profound | Mixed | NA | - | 11681 | P PVS1+PM1+PP1 | [60] S |
| Lys290Asn | 870G > T | WES | POUH | Caucasian | Severe | Mixed | IP-III#, enlarged semicircular canals | Concentration difficulties, developmental retardation | LP PM1+PM2+PP1+PP4 | [83] S | |
| Leu293Val | 877C > G | NGS Panel | POUH | Chinese | Severe | NA | NA | - | 417622 | LP PM1+PM2 | [62] C |
| Glu294Gly | 881A > G | NGS Panel | POUH | Chinese | NA | NA | NA | - | LP PM1+PM2+PP1+PP3 | [84] C | |
| Pro301Leu | 902C > T | Sanger | POUH | Mixed | Severe | Mixed | NA | - | P PM1+PM2+PP1 | [81] S | |
| Lys302Alafs*25 | 903_912 delins TGCCA | Sanger | POUH* | Chinese | Moderate | NA | IP-III | - | P PVS1+PM1+PP1+PP4 | [66] C | |
| Gln306* | 916C > T | WES | POUH | Caucasian | Severe | SNHL | IP-III | - | 1799532 | P PVS1+PM1+PM2+PP4 | [64] C |
| Ile308Asn | NA | Sanger | POUH | NA | Moderate | Mixed | Normal | - | NA PM1+PP1 | [20] C | |
| Ile308Ilefs*28 | NA | Sanger | POUH* | NA | Normal | - | Normal | - | NA PVS1+PM1+PP1 | [20] C | |
| Ser309Pro | 925T > C | Sanger | POUH | Chinese | Profound | Mixed | IP-III# | Mild sinus arrhythmia | LP PM1+PM2+PP4 | [85] S | |
| Ser310del | 927-929delCTC | Sanger | POUH | Korean | Severe | Mixed | IP-III | - | 43353 | P PM1+PS3+PP1+PP4 | [50] S |
| Ala312Pro | 934G > C | NGS Panel | POUH | Caucasian | Profound | Mixed | IP-III | - | LP PM1+PM2+PP1+PP4 | [86] S | |
| Ala312Val | 935C > T | Sanger | POUH | Caucasian | Profound | SNHL | IP-III# | Learning difficulties | 11682 | LP PM1+PP1+PP4 | [60] S |
| Gln316* | 946C > T | Sanger | POUH* | Chinese | Severe | NA | IP-III | - | P PVS1+PM1+PM2+PP1+PP4 | [66] C | |
| Leu317* | 950 T > A | NGS Panel | POUH* | Chinese | NA | NA | NA | - | P PVS1+PM1+PM2 | [87] C | |
| Leu317Phefs*12 | 950dupT | Sanger | POUH* | Korean | Severe | NA | IP-III | - | 41457 | LP PVS1+PM1+PM2+PS3+PP1+PP4 | [49] S |
| Glu320* | c.958G>T | WES | POUH* | Chinese | Moderate to severe | Mixed | IP-III | - | LP PVS1+PM2+PP4 | [72] C | |
| Val321Gly | 962T > G | WES | POUH | Chinese | Severe | SNHL | IP-III | - | LP PS1+PM1+PM2+PP1+ PP4 | [66,69] C | |
| Arg323Gly | 967C > G | PCR/SSC | POUH | Caucasian | NA | Mixed | dilation of semicircular canals and IAM | - | 11684 | P PM1+PP4 | [88] S |
| Val324Asp | 971T > A | WES | POUH | Caucasian | Severe | Mixed | IP-III | - | P PM1+PM2+PP1+PP4 | [64] C | |
| Trp325Glyfs*12 | 973delT | Sanger | POUH* | Chinese | NA | Mixed | IP-III | - | P PVS1+PM1+PP1+PP4 | [77] C | |
| Trp325Arg | 973 T > A | Sanger | POUH | Caucasian | Profound | SNHL | dysplastic cochlea on both sides, flat angle of the facial nerve | - | LP PP1+PM1+PP3 | [89] S | |
| Trp325* | 975G > A | Sanger | POUH* | Siberia | NA | Mixed | IP-III#, enlarged semicircular canals | Vestibular dysfunction, dystaxia | 599350 | P PVS1+PM1+PM2+PP1+PP4 | [19] S |
| Cys327* | c.979T > A | Sanger | POUH* | Caucasian | Profound to Severe | Mixed | EVA, IP-III | Cognitive deficit | 1701826 | P PVS1+PM1+PS3+PP1+ PP4 | [25] S |
| Arg329Gly | 985C > G | NGS Panel | POUH NLS | Chinese | NA | NA | NA | - | 1297060 | LP PM1+PM2 | [62] C |
| Arg329Pro | 986 G > C | Sanger | POUH NLS | Korean | Severe | Mixed | IP-III | - | P PM1+PS3+PP4+PP1 | [50] S | |
| Ile308Thr | 987T > C | Sanger | POUH NLS | Mixed | Severe | Mixed | NA | - | P PM1+PM2+PP1 | [81] S | |
| Arg330Lys | c.989G>A | WES | POUH NLS | Chinese | Severe | Mixed | IP-III | - | LP PS2+PM2+PP3+PP4 | [72] C | |
| Arg330Ser | 990A > T | PCR/SSC | POUH NLS | Caucasian | NA | NA | Enlarged semicircular canals IAM | Growth retardation, subclinical hypothyroidism, diarrhea | 11683 | P PM1 | [88] S |
| Gln331Pro | 992A > C | Sanger | POUH NLS | Japan | Left: normal/Right: moderate | Mixed | EVA, IP-III | - | LP PM1+PM2+PP1+PP4 | [67] C | |
| Thr354Glnfs*115 | 1069delA | Sanger | C-term ext. | Korean | Profound | NA | IP-III | - | LP PM2+PS3+PP1+PP4 | [49] S | |
| *362Argext*113 | 1084T > C | Sanger | C-term ext. | Korean | Profound | NA | IP-III | - | LP PM2+PS3+PP4 | [49] S | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardinelli, E.; Huber, F.; Roesch, S.; Dossena, S. Clinical and Molecular Aspects Associated with Defects in the Transcription Factor POU3F4: A Review. Biomedicines 2023, 11, 1695. https://doi.org/10.3390/biomedicines11061695
Bernardinelli E, Huber F, Roesch S, Dossena S. Clinical and Molecular Aspects Associated with Defects in the Transcription Factor POU3F4: A Review. Biomedicines. 2023; 11(6):1695. https://doi.org/10.3390/biomedicines11061695
Chicago/Turabian StyleBernardinelli, Emanuele, Florian Huber, Sebastian Roesch, and Silvia Dossena. 2023. "Clinical and Molecular Aspects Associated with Defects in the Transcription Factor POU3F4: A Review" Biomedicines 11, no. 6: 1695. https://doi.org/10.3390/biomedicines11061695
APA StyleBernardinelli, E., Huber, F., Roesch, S., & Dossena, S. (2023). Clinical and Molecular Aspects Associated with Defects in the Transcription Factor POU3F4: A Review. Biomedicines, 11(6), 1695. https://doi.org/10.3390/biomedicines11061695