
Direct Effects of Mifepristone on Mice Embryogenesis: An In Vitro Evaluation by Single-Embryo RNA Sequencing Analysis

 , , , and
, , , and
Abstract
1. Introduction
2. Material and Methods
2.1. Collection of Mouse Blastocysts
2.2. Standard Blastocyst Culture
2.3. Blastocyst Culture and RU-486 Treatment
2.4. Single Embryo RNA Isolation, cDNA Synthesis, and RNA Library Preparation
2.5. Sequencing and Analysis
2.6. Gene Sets Enrichment Analysis
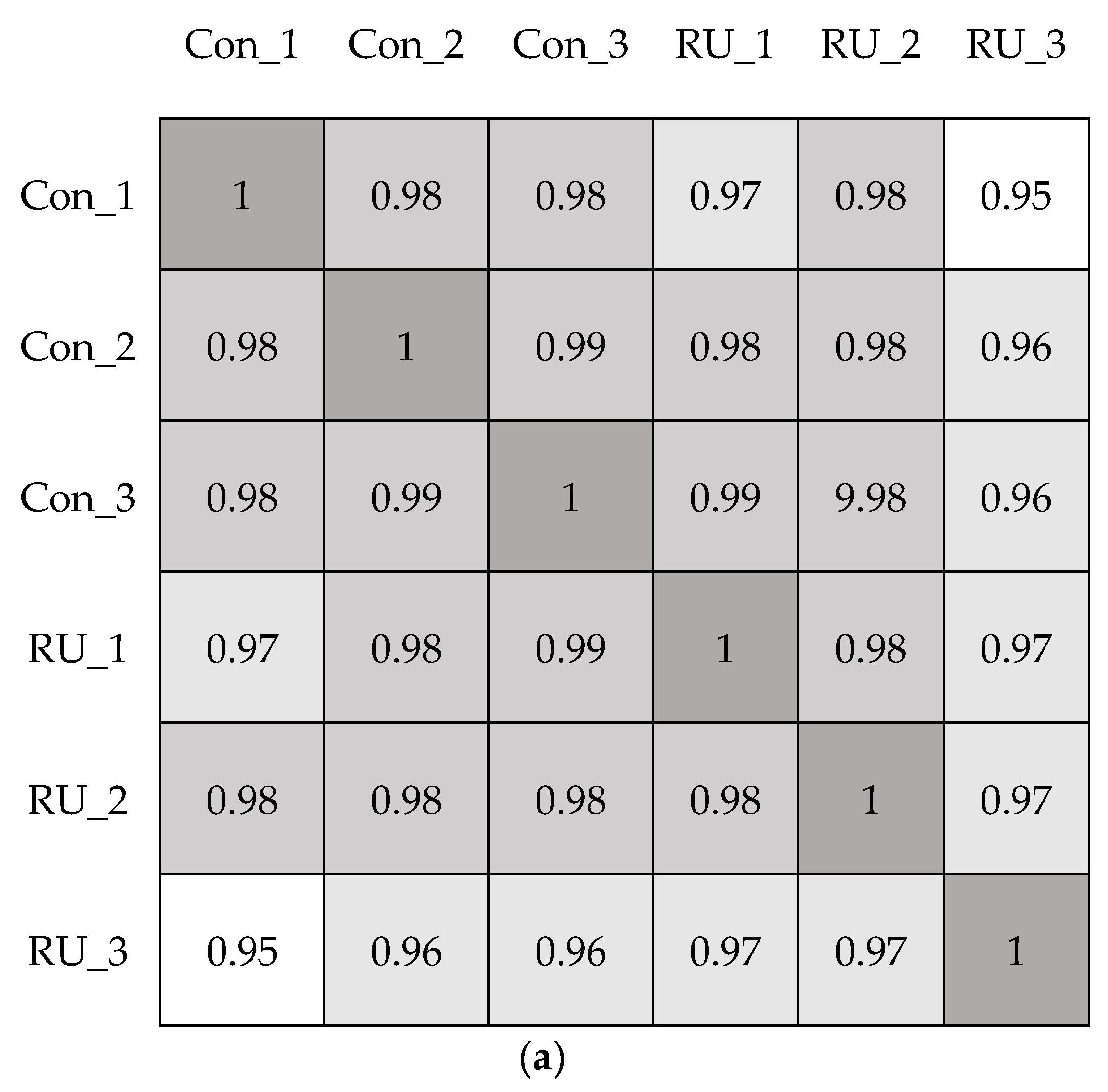
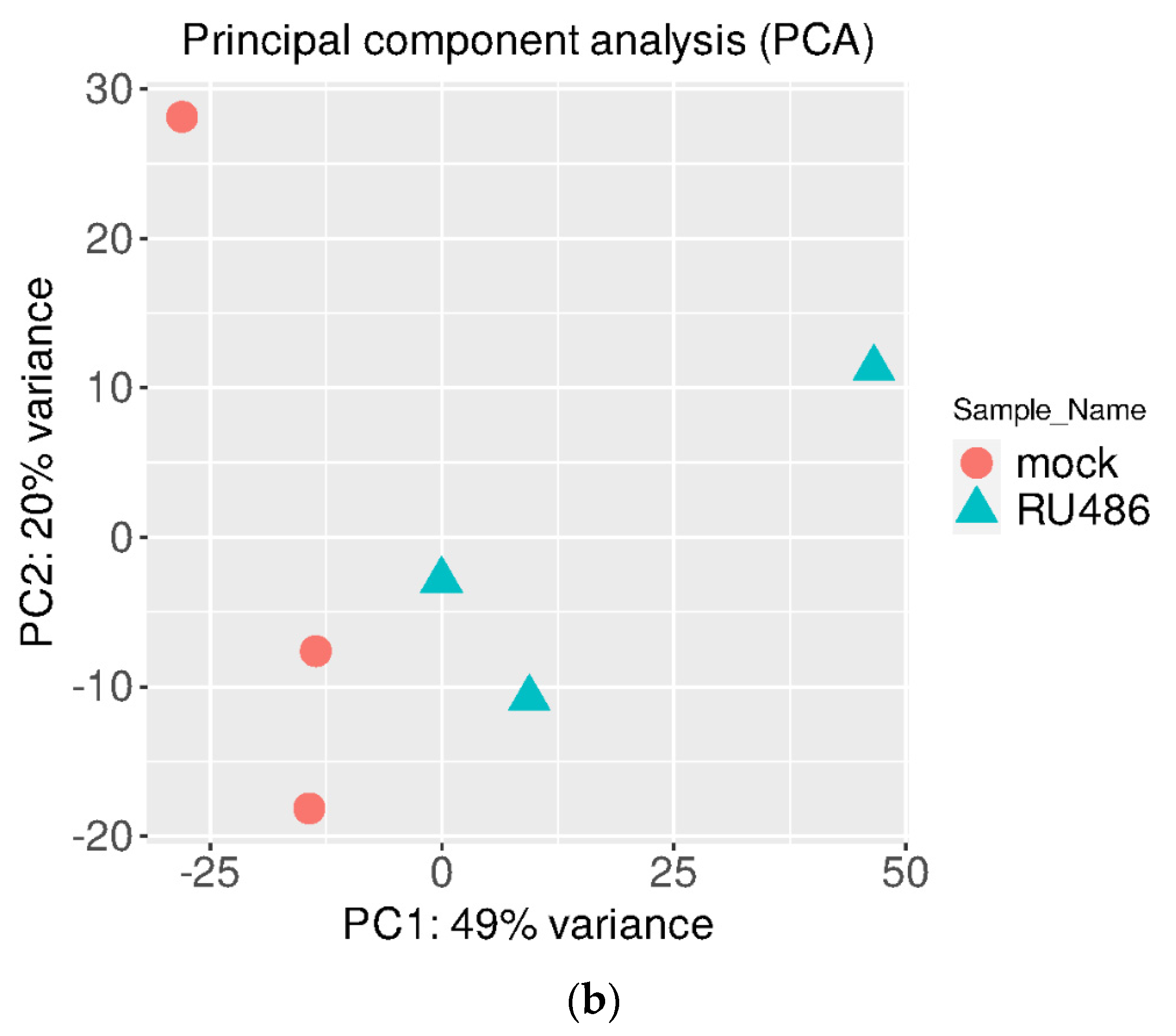
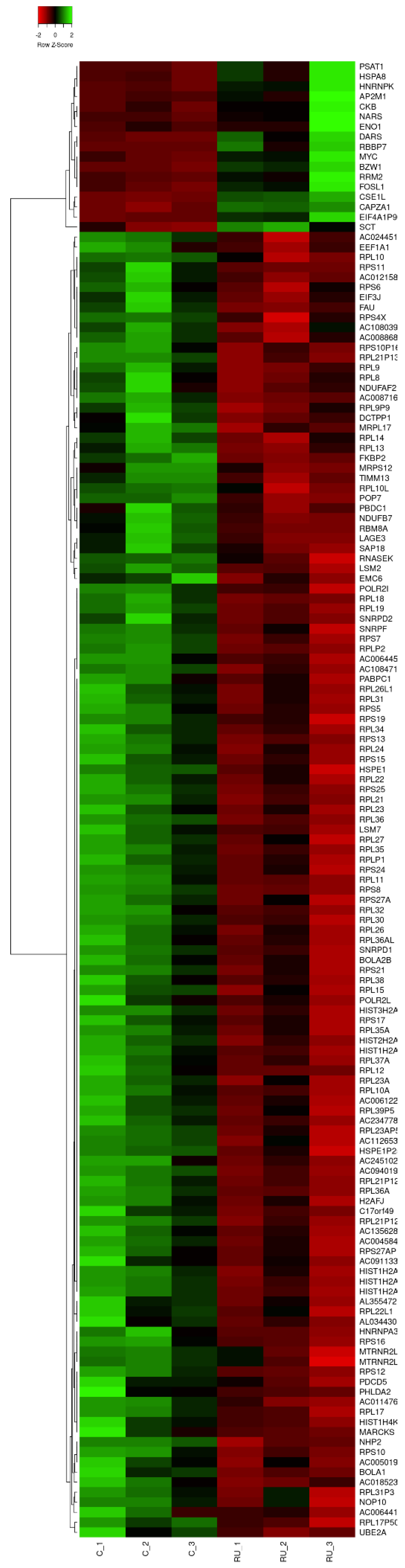
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, M.J.; Creinin, M.D. Mifepristone With Buccal Misoprostol for Medical Abortion: A Systematic Review. Obstet. Gynecol. 2015, 126, 12–21. [Google Scholar] [CrossRef]
- Murji, A.; Whitaker, L.; Chow, T.L.; Sobel, M.L. Selective progesterone receptor modulators (SPRMs) for uterine fibroids. Cochrane Database Syst. Rev. 2017, 4, CD010770. [Google Scholar] [CrossRef]
- Ngoc, N.T.; Shochet, T.; Raghavan, S.; Blum, J.; Nga, N.T.; Minh, N.T.; Phan, V.Q.; Winikoff, B. Mifepristone and misoprostol compared with misoprostol alone for second-trimester abortion: A randomized controlled trial. Obstet. Gynecol. 2011, 118, 601–608. [Google Scholar] [CrossRef]
- Gallagher, P.; Young, A.H. Mifepristone (RU-486) treatment for depression and psychosis: A review of the therapeutic implications. Neuropsychiatr. Dis. Treat. 2006, 2, 33–42. [Google Scholar]
- Spitz, I.M.; Bardin, C.W.; Benton, L.; Robbins, A. Early pregnancy termination with mifepristone and misoprostol in the United States. N. Engl. J. Med. 1998, 338, 1241–1247. [Google Scholar] [CrossRef]
- Hsia, J.K.; Lohr, P.A.; Taylor, J.; Creinin, M.D. Medical abortion with mifepristone and vaginal misoprostol between 64 and 70 days’ gestation. Contraception 2019, 100, 178–181. [Google Scholar] [CrossRef]
- Delgado, G.; Davenport, M.L. Progesterone use to reverse the effects of mifepristone. Ann. Pharmacother. 2012, 46, e36. [Google Scholar] [CrossRef] [PubMed]
- Garratt, D.; Turner, J.V. Progesterone for preventing pregnancy termination after initiation of medical abortion with mifepristone. Eur. J. Contracept. Reprod. Health Care 2017, 22, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Delgado, G.; Condly, S.J.; Davenport, M.; Tinnakornsrisuphap, T.; Mack, J.; Khauv, V.; Zhou, P.S. A case series detailing the successful reversal of the effects of mifepristone using progesterone. Issues Law Med. 2019, 33, 21–31. [Google Scholar]
- Bhatti, K.Z.; Nguyen, A.T.; Stuart, G.S. Medical abortion reversal: Science and politics meet. Am. J. Obstet. Gynecol. 2018, 218, 315.e311–315.e316. [Google Scholar] [CrossRef]
- Grossman, D.; White, K. Abortion “Reversal”—Legislating without Evidence. N. Engl. J. Med. 2018, 379, 1491–1493. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.J.; Lan, K.C.; Kang, H.Y.; Liu, Y.C.; Hsuuw, Y.D.; Chan, W.H.; Huang, K.E. Effect of curcumin on in vitro early post-implantation stages of mouse embryo development. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 166, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Poli, M.; Ori, A.; Child, T.; Jaroudi, S.; Spath, K.; Beck, M.; Wells, D. Characterization and quantification of proteins secreted by single human embryos prior to implantation. EMBO Mol. Med. 2015, 7, 1465–1479. [Google Scholar] [CrossRef]
- Wu, S.J.; Khoriaty, R.; Kim, S.H.; O’Shea, K.S.; Zhu, G.; Hoenerhoff, M.; Zajac, C.; Oravecz-Wilson, K.; Toubai, T.; Sun, Y.; et al. SNARE protein SEC22B regulates early embryonic development. Sci. Rep. 2019, 9, 11434. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Sun-Wada, G.H.; Kawamura, N.; Yasukawa, J. Membrane dynamics in mammalian embryogenesis: Implication in signal regulation. Birth Defects Res. C Embryo Today 2016, 108, 33–44. [Google Scholar] [CrossRef]
- Zuniga, A.; Hodar, C.; Hanna, P.; Ibanez, F.; Moreno, P.; Pulgar, R.; Pastenes, L.; Gonzalez, M.; Cambiazo, V. Genes encoding novel secreted and transmembrane proteins are temporally and spatially regulated during Drosophila melanogaster embryogenesis. BMC Biol. 2009, 7, 61. [Google Scholar] [CrossRef]
- Rion, N.; Castets, P.; Lin, S.; Enderle, L.; Reinhard, J.R.; Eickhorst, C.; Ruegg, M.A. mTOR controls embryonic and adult myogenesis via mTORC1. Development 2019, 146, dev.172460. [Google Scholar] [CrossRef]
- Murakami, M.; Ichisaka, T.; Maeda, M.; Oshiro, N.; Hara, K.; Edenhofer, F.; Kiyama, H.; Yonezawa, K.; Yamanaka, S. mTOR is essential for growth and proliferation in early mouse embryos and embryonic stem cells. Mol. Cell. Biol. 2004, 24, 6710–6718. [Google Scholar] [CrossRef]
- Land, S.C.; Scott, C.L.; Walker, D. mTOR signalling, embryogenesis and the control of lung development. Semin. Cell Dev. Biol. 2014, 36, 68–78. [Google Scholar] [CrossRef]
- Correia, B.; Sousa, M.I.; Ramalho-Santos, J. The mTOR pathway in reproduction: From gonadal function to developmental coordination. Reproduction 2020, 159, R173–R188. [Google Scholar] [CrossRef]
- Sciarretta, S.; Forte, M.; Frati, G.; Sadoshima, J. New Insights Into the Role of mTOR Signaling in the Cardiovascular System. Circ. Res. 2018, 122, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Bloch, W.; Iden, S.; Ruegg, M.A.; Hall, M.N.; Leptin, M.; Partridge, L.; Eming, S.A. mTORC1 and mTORC2 regulate skin morphogenesis and epidermal barrier formation. Nat. Commun. 2016, 7, 13226. [Google Scholar] [CrossRef]
- Chirala, S.S.; Chang, H.; Matzuk, M.; Abu-Elheiga, L.; Mao, J.; Mahon, K.; Finegold, M.; Wakil, S.J. Fatty acid synthesis is essential in embryonic development: Fatty acid synthase null mutants and most of the heterozygotes die in utero. Proc. Natl. Acad. Sci. USA 2003, 100, 6358–6363. [Google Scholar] [CrossRef]
- Haggarty, P.; Wood, M.; Ferguson, E.; Hoad, G.; Srikantharajah, A.; Milne, E.; Hamilton, M.; Bhattacharya, S. Fatty acid metabolism in human preimplantation embryos. Hum. Reprod. 2006, 21, 766–773. [Google Scholar] [CrossRef]
- McKeegan, P.J.; Sturmey, R.G. The role of fatty acids in oocyte and early embryo development. Reprod. Fertil. Dev. 2011, 24, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Dubeibe Marin, D.F.; da Costa, N.N.; di Paula Bessa Santana, P.; de Souza, E.B.; Ohashi, O.M. Importance of lipid metabolism on oocyte maturation and early embryo development: Can we apply what we know to buffalo? Anim. Reprod. Sci. 2019, 211, 106220. [Google Scholar] [CrossRef]
- Esteban-Vives, R.; Ziembicki, J.; Sun Choi, M.; Thompson, R.L.; Schmelzer, E.; Gerlach, J.C. Isolation and Characterization of a Human Fetal Mesenchymal Stem Cell Population: Exploring the Potential for Cell Banking in Wound Healing Therapies. Cell Transplant. 2019, 28, 1404–1419. [Google Scholar] [CrossRef]
- Billon, N.; Kolde, R.; Reimand, J.; Monteiro, M.C.; Kull, M.; Peterson, H.; Tretyakov, K.; Adler, P.; Wdziekonski, B.; Vilo, J.; et al. Comprehensive transcriptome analysis of mouse embryonic stem cell adipogenesis unravels new processes of adipocyte development. Genome Biol. 2010, 11, R80. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef]
- Dunwoodie, S.L. The role of hypoxia in development of the Mammalian embryo. Dev. Cell 2009, 17, 755–773. [Google Scholar] [CrossRef]
- Scully, D.; Keane, E.; Batt, E.; Karunakaran, P.; Higgins, D.F.; Itasaki, N. Hypoxia promotes production of neural crest cells in the embryonic head. Development 2016, 143, 1742–1752. [Google Scholar] [CrossRef] [PubMed]
- Cowden Dahl, K.D.; Fryer, B.H.; Mack, F.A.; Compernolle, V.; Maltepe, E.; Adelman, D.M.; Carmeliet, P.; Simon, M.C. Hypoxia-inducible factors 1alpha and 2alpha regulate trophoblast differentiation. Mol. Cell. Biol. 2005, 25, 10479–10491. [Google Scholar] [CrossRef]
- Schipani, E.; Ryan, H.E.; Didrickson, S.; Kobayashi, T.; Knight, M.; Johnson, R.S. Hypoxia in cartilage: HIF-1alpha is essential for chondrocyte growth arrest and survival. Genes Dev. 2001, 15, 2865–2876. [Google Scholar] [CrossRef]
- Amarilio, R.; Viukov, S.V.; Sharir, A.; Eshkar-Oren, I.; Johnson, R.S.; Zelzer, E. HIF1alpha regulation of Sox9 is necessary to maintain differentiation of hypoxic prechondrogenic cells during early skeletogenesis. Development 2007, 134, 3917–3928. [Google Scholar] [CrossRef]
- Farr, R.L.; Lismont, C.; Terlecky, S.R.; Fransen, M. Peroxisome biogenesis in mammalian cells: The impact of genes and environment. Biochim. Biophys. Acta 2016, 1863, 1049–1060. [Google Scholar] [CrossRef]
- Nardacci, R.; Falciatori, I.; Moreno, S.; Stefanini, S. Immunohistochemical localization of peroxisomal enzymes during rat embryonic development. J. Histochem. Cytochem. 2004, 52, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, H.; Aulehla, A. Revisiting the role of metabolism during development. Development 2018, 145, dev.131110. [Google Scholar] [CrossRef]
- Oginuma, M.; Moncuquet, P.; Xiong, F.; Karoly, E.; Chal, J.; Guevorkian, K.; Pourquie, O. A Gradient of Glycolytic Activity Coordinates FGF and Wnt Signaling during Elongation of the Body Axis in Amniote Embryos. Dev. Cell 2017, 40, 342–353.e10. [Google Scholar] [CrossRef] [PubMed]
- Lunt, S.Y.; Vander Heiden, M.G. Aerobic glycolysis: Meeting the metabolic requirements of cell proliferation. Annu. Rev. Cell Dev. Biol. 2011, 27, 441–464. [Google Scholar] [CrossRef]
- Tixier, V.; Bataille, L.; Etard, C.; Jagla, T.; Weger, M.; Daponte, J.P.; Strahle, U.; Dickmeis, T.; Jagla, K. Glycolysis supports embryonic muscle growth by promoting myoblast fusion. Proc. Natl. Acad. Sci. USA 2013, 110, 18982–18987. [Google Scholar] [CrossRef] [PubMed]
- Julian, L.M.; Liu, Y.; Pakenham, C.A.; Dugal-Tessier, D.; Ruzhynsky, V.; Bae, S.; Tsai, S.Y.; Leone, G.; Slack, R.S.; Blais, A. Tissue-specific targeting of cell fate regulatory genes by E2f factors. Cell Death Differ. 2016, 23, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Hemmati-Brivanlou, A. Xenopus embryonic E2F is required for the formation of ventral and posterior cell fates during early embryogenesis. Mol. Cell 2000, 5, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.D.; Funk, L.B.; FitzGerald, G.A.; Samuelsson, B. Characterization of human 12-lipoxygenase genes. Proc. Natl. Acad. Sci. USA 1992, 89, 3962–3966. [Google Scholar] [CrossRef]
- Duronio, R.J.; O’Farrell, P.H.; Xie, J.E.; Brook, A.; Dyson, N. The transcription factor E2F is required for S phase during Drosophila embryogenesis. Genes Dev. 1995, 9, 1445–1455. [Google Scholar] [CrossRef]
- King, J.C.; Moskowitz, I.P.; Burgon, P.G.; Ahmad, F.; Stone, J.R.; Seidman, J.G.; Lees, J.A. E2F3 plays an essential role in cardiac development and function. Cell Cycle 2008, 7, 3775–3780. [Google Scholar] [CrossRef]
- Heim, M.H. The Jak-STAT pathway: Cytokine signalling from the receptor to the nucleus. J. Recept. Signal Transduct. Res. 1999, 19, 75–120. [Google Scholar] [CrossRef]
- Akira, S. Functional roles of STAT family proteins: Lessons from knockout mice. Stem Cells 1999, 17, 138–146. [Google Scholar] [CrossRef]
- Nemetz, C.; Hocke, G.M. Transcription factor Stat5 is an early marker of differentiation of murine embryonic stem cells. Differentiation 1998, 62, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.S.; Noor, S.M.; Fraser, F.W.; Sertori, R.; Liongue, C.; Ward, A.C. Regulation of embryonic hematopoiesis by a cytokine-inducible SH2 domain homolog in zebrafish. J. Immunol. 2014, 192, 5739–5748. [Google Scholar] [CrossRef]
- Chiabrando, D.; Vinchi, F.; Fiorito, V.; Mercurio, S.; Tolosano, E. Heme in pathophysiology: A matter of scavenging, metabolism and trafficking across cell membranes. Front. Pharm. 2014, 5, 61. [Google Scholar] [CrossRef]
- Shi, J.; Mei, W.; Yang, J. Heme metabolism enzymes are dynamically expressed during Xenopus embryonic development. Biocell 2008, 32, 259–263. [Google Scholar] [CrossRef]
- Ponka, P. Cell biology of heme. Am. J. Med. Sci. 1999, 318, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Sun, J.; Taketani, S.; Nakajima, O.; Nishitani, C.; Sassa, S.; Hayashi, N.; Yamamoto, M.; Shibahara, S.; Fujita, H.; et al. Heme mediates derepression of Maf recognition element through direct binding to transcription repressor Bach1. EMBO J. 2001, 20, 2835–2843. [Google Scholar] [CrossRef] [PubMed]
- Marro, S.; Chiabrando, D.; Messana, E.; Stolte, J.; Turco, E.; Tolosano, E.; Muckenthaler, M.U. Heme controls ferroportin1 (FPN1) transcription involving Bach1, Nrf2 and a MARE/ARE sequence motif at position −7007 of the FPN1 promoter. Haematologica 2010, 95, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, H.; Ji, Y.L.; Zhang, Y.; Yu, T.; Ning, H.; Zhang, C.; Zhao, X.F.; Wang, Q.; Liu, P.; et al. Maternal fenvalerate exposure during pregnancy persistently impairs testicular development and spermatogenesis in male offspring. Food Chem. Toxicol. 2010, 48, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Maksoud, F.M.; Ali, F.A.Z.; Akingbemi, B.T. Prenatal exposures to bisphenol A and di (2-ethylhexyl) phthalate disrupted seminiferous tubular development in growing male rats. Reprod. Toxicol. 2019, 88, 85–90. [Google Scholar] [CrossRef]
- Tincello, D.G.; Saunders, P.T.; Hodgins, M.B.; Simpson, N.B.; Edwards, C.R.; Hargreaves, T.B.; Wu, F.C. Correlation of clinical, endocrine and molecular abnormalities with in vivo responses to high-dose testosterone in patients with partial androgen insensitivity syndrome. Clin. Endocrinol. 1997, 46, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Pihlajamaa, P.; Sahu, B.; Janne, O.A. Determinants of Receptor- and Tissue-Specific Actions in Androgen Signaling. Endocr. Rev. 2015, 36, 357–384. [Google Scholar] [CrossRef] [PubMed]
- Crocoll, A.; Zhu, C.C.; Cato, A.C.; Blum, M. Expression of androgen receptor mRNA during mouse embryogenesis. Mech. Dev. 1998, 72, 175–178. [Google Scholar] [CrossRef]
- Patterson, M.N.; McPhaul, M.J.; Hughes, I.A. Androgen insensitivity syndrome. Baillieres. Clin. Endocrinol. Metab. 1994, 8, 379–404. [Google Scholar] [CrossRef]
- Chang, C.Y.; Hsuuw, Y.D.; Huang, F.J.; Shyr, C.R.; Chang, S.Y.; Huang, C.K.; Kang, H.Y.; Huang, K.E. Androgenic and antiandrogenic effects and expression of androgen receptor in mouse embryonic stem cells. Fertil. Steril. 2006, 85 (Suppl. 1), 1195–1203. [Google Scholar] [CrossRef]
- Bernard, N.; Elefant, E.; Carlier, P.; Tebacher, M.; Barjhoux, C.E.; Bos-Thompson, M.A.; Amar, E.; Descotes, J.; Vial, T. Continuation of pregnancy after first-trimester exposure to mifepristone: An observational prospective study. Int. J. Obstet. Gynaecol. 2013, 120, 568–574. [Google Scholar] [CrossRef]
- da Silva Dal Pizzol, T.; Knop, F.P.; Mengue, S.S. Prenatal exposure to misoprostol and congenital anomalies: Systematic review and meta-analysis. Reprod. Toxicol. 2006, 22, 666–671. [Google Scholar] [CrossRef] [PubMed]
- da Silva Dal Pizzol, T.; Tierling, V.L.; Schuler-Faccin, L.; Sanseverino, M.T.; Mengue, S.S. Reproductive results associated with misoprostol and other substances utilized for interruption of pregnancy. Eur. J. Clin. Pharmacol. 2005, 61, 71–72. [Google Scholar] [CrossRef] [PubMed]
- Auffret, M.; Bernard-Phalippon, N.; Dekemp, J.; Carlier, P.; Gervoise Boyer, M.; Vial, T.; Gautier, S. Misoprostol exposure during the first trimester of pregnancy: Is the malformation risk varying depending on the indication? Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 207, 188–192. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
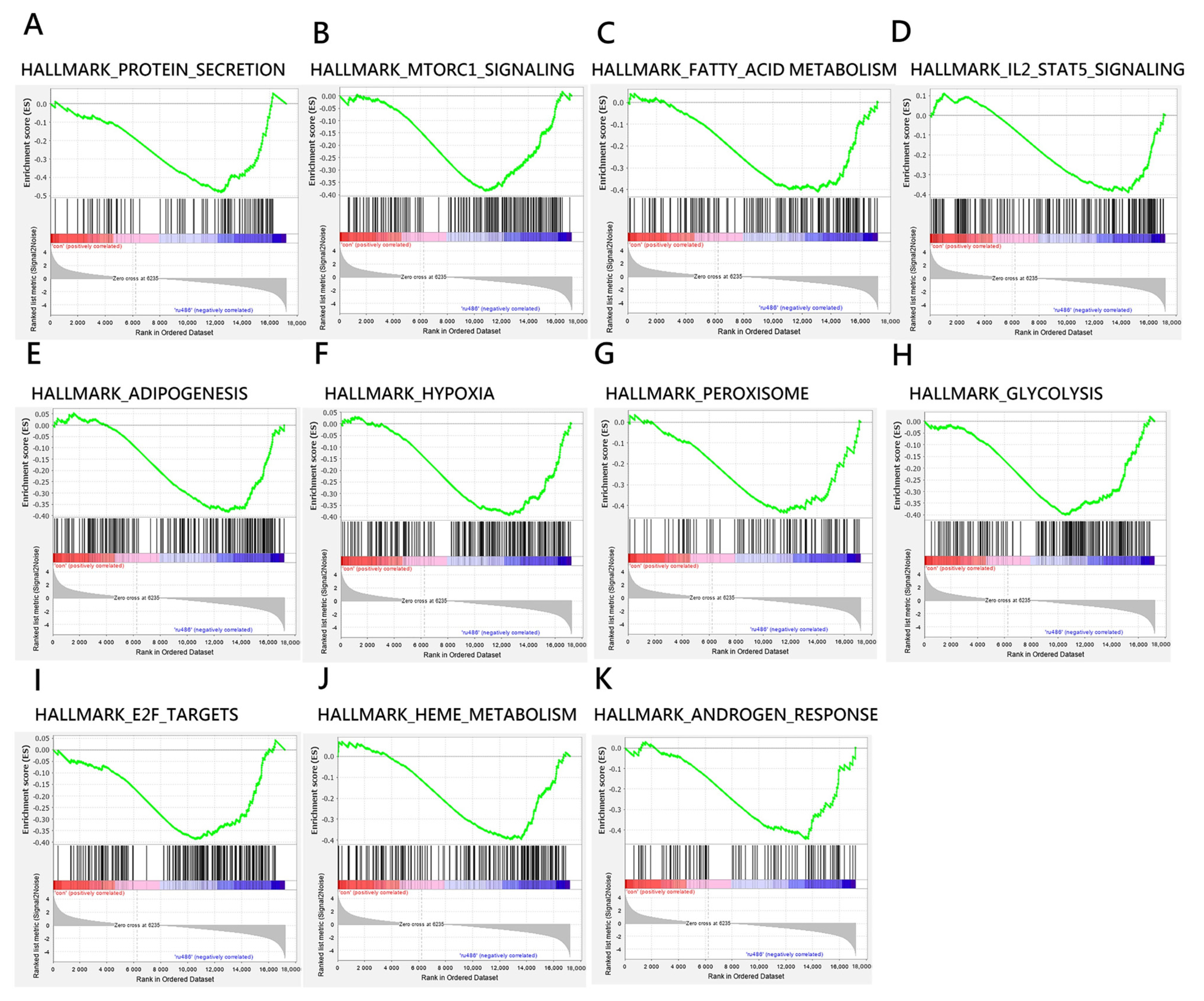
| NAME | SIZE | ES | NES | NOM p-Value (p < 0.05) | FDR q-Value (q < 0.25) |
|---|---|---|---|---|---|
| HALLMARK_PROTEIN_SECRETION | 94 | −0.481 | −1.564 | 0.004 | 0.087 |
| HALLMARK_MTORC1_SIGNALING | 187 | −0.384 | −1.344 | 0.020 | 0.125 |
| HALLMARK_FATTY_ACID_METABOLISM | 155 | −0.409 | −1.409 | 0.008 | 0.126 |
| HALLMARK_IL2_STAT5_SIGNALING | 192 | −0.389 | −1.371 | 0.012 | 0.128 |
| HALLMARK_ADIPOGENESIS | 194 | −0.384 | −1.346 | 0.018 | 0.134 |
| HALLMARK_HYPOXIA | 196 | −0.393 | −1.376 | 0.003 | 0.138 |
| HALLMARK_PEROXISOME | 102 | −0.435 | −1.445 | 0.013 | 0.140 |
| HALLMARK_GLYCOLYSIS | 193 | −0.400 | −1.418 | 0.003 | 0.141 |
| HALLMARK_E2F_TARGETS | 192 | −0.387 | −1.348 | 0.016 | 0.146 |
| HALLMARK_HEME_METABOLISM | 184 | −0.396 | −1.381 | 0.011 | 0.150 |
| HALLMARK_ANDROGEN_RESPONSE | 95 | −0.444 | −1.455 | 0.014 | 0.179 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.-T.; Chen, J.-S.; Lan, K.-C.; Lee, Y.-K.; Chu, T.-H.; Ho, Y.-C.; Wu, C.-C.; Huang, F.-J. Direct Effects of Mifepristone on Mice Embryogenesis: An In Vitro Evaluation by Single-Embryo RNA Sequencing Analysis. Biomedicines 2023, 11, 907. https://doi.org/10.3390/biomedicines11030907
Su Y-T, Chen J-S, Lan K-C, Lee Y-K, Chu T-H, Ho Y-C, Wu C-C, Huang F-J. Direct Effects of Mifepristone on Mice Embryogenesis: An In Vitro Evaluation by Single-Embryo RNA Sequencing Analysis. Biomedicines. 2023; 11(3):907. https://doi.org/10.3390/biomedicines11030907
Chicago/Turabian StyleSu, Yu-Ting, Jia-Shing Chen, Kuo-Chung Lan, Yung-Kuo Lee, Tian-Huei Chu, Yu-Cheng Ho, Cheng-Chun Wu, and Fu-Jen Huang. 2023. "Direct Effects of Mifepristone on Mice Embryogenesis: An In Vitro Evaluation by Single-Embryo RNA Sequencing Analysis" Biomedicines 11, no. 3: 907. https://doi.org/10.3390/biomedicines11030907
APA StyleSu, Y.-T., Chen, J.-S., Lan, K.-C., Lee, Y.-K., Chu, T.-H., Ho, Y.-C., Wu, C.-C., & Huang, F.-J. (2023). Direct Effects of Mifepristone on Mice Embryogenesis: An In Vitro Evaluation by Single-Embryo RNA Sequencing Analysis. Biomedicines, 11(3), 907. https://doi.org/10.3390/biomedicines11030907