
Integration of Ultrastructural and Computational Approaches Reveals the Protective Effect of Astaxanthin against BPA-Induced Nephrotoxicity

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Design
2.2. Light Microscopy
2.3. Transmission Electron Microscopy (TEM)
2.4. In Silico Analysis of the Potential Effect of ATX on P450 CYP2C9
2.4.1. System Preparation
2.4.2. Molecular Dynamic (MD) Simulations
2.4.3. Post-MD Analysis
2.5. Statistical Analysis
3. Results
3.1. Light Microscope
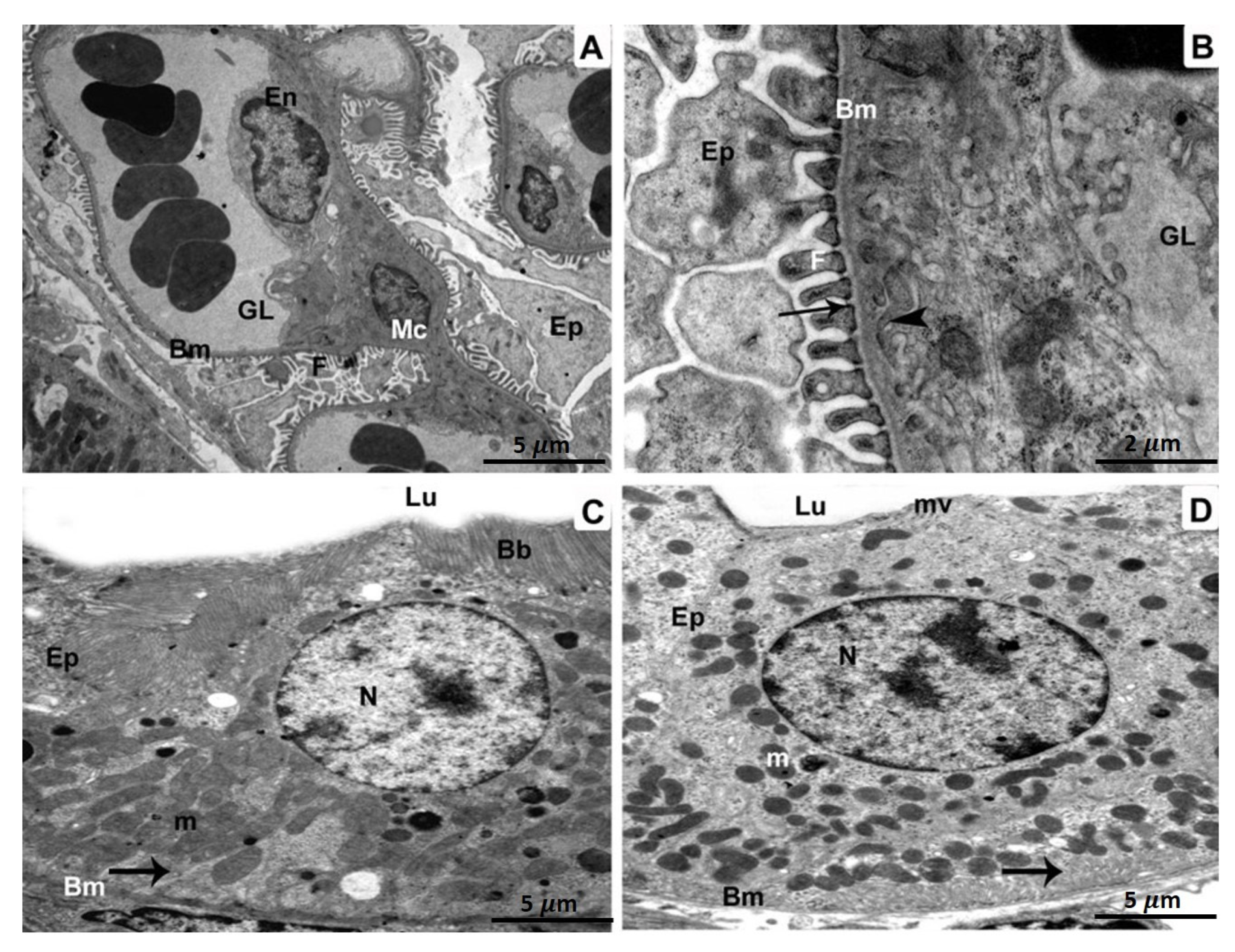
3.2. Transmission Electron Microscope (TEM)
3.3. In Silico Drug Discovery
3.3.1. Molecular Dynamic and System Stability
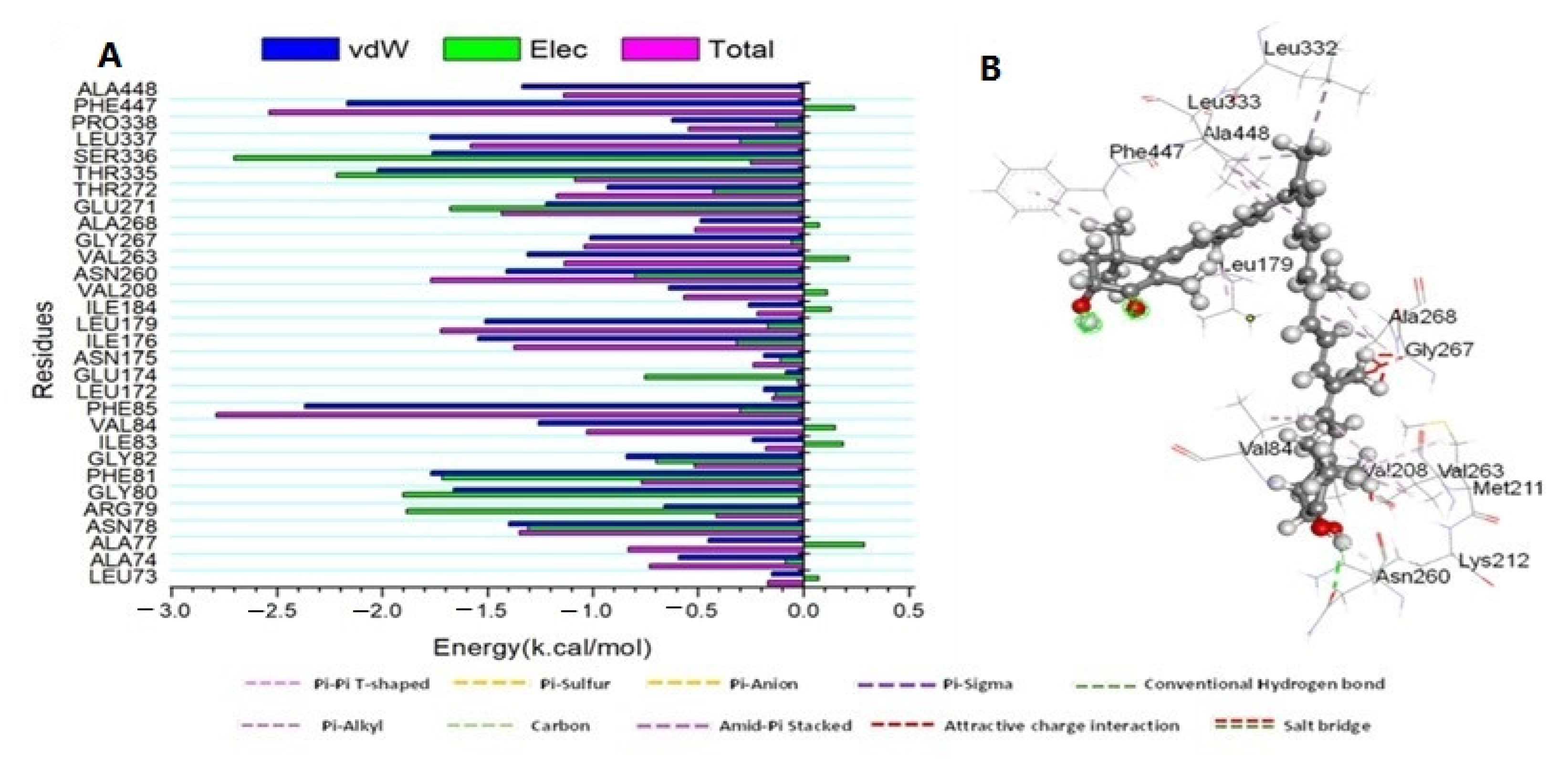
3.3.2. Binding Interaction Mechanism Based on Binding Free Energy Calculation
3.3.3. Determination of the Essential Amino Acid Residues Implicated in Ligand Binding and Complex Formation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Staniszewska, M.; Graca, B.; Nehring, I. The fate of bisphenol A, 4-tert-octylphenol and 4-nonylphenol leached from plastic debris into marine water—Experimental studies on biodegradation and sorption on suspended particulate matter and nano-TiO2. Chemosphere 2016, 145, 535–542. [Google Scholar] [CrossRef]
- Liguori, F.; Moreno-Marrodan, C.; Barbaro, P. Biomass-derived chemical substitutes for bisphenol A: Recent advancements in catalytic synthesis. Chem. Soc. Rev. 2020, 49, 6329–6363. [Google Scholar] [CrossRef]
- Ejaredar, M.; Lee, Y.; Roberts, D.J.; Sauve, R.; Dewey, D. Bisphenol A exposure and children’s behavior: A systematic review. J. Expo. Sci. Environ. Epidemiol. 2017, 27, 175–183. [Google Scholar] [CrossRef] [PubMed]
- lo Turco, V.; Potortì, A.G.; ben Mansour, H.; Dugo, G.; di Bella, G. Plasticizers and BPA in spices and aromatic herbs of Mediterranean areas. Nat. Prod. Res. 2020, 34, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Calafat, A.M.; Ye, X.; Wong, L.Y.; Reidy, J.A.; Needham, L.L. Exposure of the U.S. population to Bisphenol A and 4-tertiary-octylphenol: 2003–2004. Environ. Health Perspect. 2008, 116, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Mouneimne, Y.; Nasrallah, M.; Khoueiry-Zgheib, N.; Nasreddine, L.; Nakhoul, N.; Ismail, H.; Abiad, M.; Koleilat, L.; Tamim, H. Bisphenol A urinary level, its correlates, and association with cardiometabolic risks in Lebanese urban adults. Environ. Monit. Assess. 2017, 189, 517. [Google Scholar] [CrossRef] [PubMed]
- Fenichel, P.; Dechaux, H.; Harthe, C.; Gal, J.; Ferrari, P.; Pacini, P.; Wagner-Mahler, K.; Pugeat, M.; Brucker-Davis, F. Unconjugated bisphenol A cord blood levels in boys with descended or undescended testes. Hum. Reprod. 2012, 27, 983–990. [Google Scholar] [CrossRef]
- Kabuto, H.; Hasuike, S.; Minagawa, N.; Shishibori, T. Effects of bisphenol A on the metabolisms of active oxygen species in mouse tissues. Environ. Res. 2003, 93, 31–35. [Google Scholar] [CrossRef]
- Moghaddam, H.S.; Samarghandian, S.; Farkhondeh, T. Effect of bisphenol A on blood glucose, lipid profile and oxidative stress indices in adult male mice. Toxicol. Mech. Methods 2015, 25, 507–513. [Google Scholar] [CrossRef]
- Tiwari, D.; Kamble, J.; Chilgunde, S.; Patil, P.; Maru, G.; Kawle, D.; Bhartiya, U.; Joseph, L.; Vanage, G. Clastogenic and mutagenic effects of bisphenol A: An endocrine disruptor. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2012, 743, 83–90. [Google Scholar] [CrossRef]
- Xia, W.; Jiang, Y.; Li, Y.; Wan, Y.; Liu, J.; Ma, Y.; Mao, Z.; Chang, H.; Li, G.; Xu, B.; et al. Early-life exposure to bisphenol a induces liver injury in rats involvement of mitochondria-mediated apoptosis. PLoS ONE 2014, 9, e90443. [Google Scholar] [CrossRef]
- Knizatova, N.; Tokárová, K.; Greifová, H.; Jambor, T.; Massányi, P.; Lukáč, N. Bisphenol A Analogues: A Brief Review of their Occurrence in Food, Biological Samples and Endocrine Effects. Arch. Ecotoxicol. 2020, 2, 89–94. [Google Scholar] [CrossRef]
- Tao, S.; Luo, Y.; He, B.; Liu, J.; Qian, X.; Ni, Y.; Zhao, R. Paraoxonase 2 modulates a proapoptotic function in LS174T cells in response to quorum sensing molecule N-[3-oxododecanoyl]-L-homoserine lactone. Sci. Rep. 2016, 6, 28778. [Google Scholar] [CrossRef]
- Li, J.; Li, N.; Yan, S.; Lu, Y.; Miao, X.; Gu, Z.; Shao, Y. Melatonin attenuates renal fibrosis in diabetic mice by activating the AMPK/PGC1α signaling pathway and rescuing mitochondrial function. Mol. Med. Rep. 2019, 19, 1318–1330. [Google Scholar] [CrossRef]
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2017, 39, 73–82. [Google Scholar] [CrossRef]
- Uzunhisarcikli, M.; Aslanturk, A. Hepatoprotective effects of curcumin and taurine against bisphenol A-induced liver injury in rats. Environ. Sci. Pollut. Res. 2019, 26, 37242–37253. [Google Scholar] [CrossRef]
- Zangar, R.C.; Davydov, D.R.; Verma, S. Mechanisms that regulate production of reactive oxygen species by cytochrome P450. Toxicol. Appl. Pharmacol. 2004, 199, 316–331. [Google Scholar] [CrossRef]
- Gonzalez, F.J. Role of cytochromes P450 in chemical toxicity and oxidative stress: Studies with CYP2E1. Mutat. Res. Mol. Mech. Mutagen. 2005, 569, 101–110. [Google Scholar] [CrossRef]
- Acharya, P.; Liao, M.; Engel, J.C.; Correia, M.A. Liver Cytochrome P450 3A endoplasmic reticulum-associated degradation: A major role for the P97 AAA ATPase in cytochrome P450 3A extraction into the cytosol. J. Biol. Chem. 2011, 286, 3815–3828. [Google Scholar] [CrossRef]
- Li, J.; Wang, F.; Xia, Y.; Dai, W.; Chen, K.; Li, S.; Liu, T.; Zheng, Y.; Wang, J.; Lu, W.; et al. Astaxanthin Pretreatment Attenuates Hepatic Ischemia Reperfusion-Induced Apoptosis and Autophagy via the ROS/MAPK Pathway in Mice. Mar. Drugs 2015, 13, 3368–3387. [Google Scholar] [CrossRef]
- Kohandel, Z.; Farkhondeh, T.; Aschner, M.; Pourbagher-Shahri, A.M.; Samarghandian, S. Anti-inflammatory action of astaxanthin and its use in the treatment of various diseases. Biomed. Pharmacother. 2022, 145, 112179. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Dohl, J.; Chen, Y.; Gasier, H.G.; Deuster, P.A. Astaxanthin but not quercetin preserves mitochondrial integrity and function, ameliorates oxidative stress, and reduces heat-induced skeletal muscle injury. J. Cell. Physiol. 2019, 234, 13292–13302. [Google Scholar] [CrossRef] [PubMed]
- van Westing, A.C.; Küpers, L.K.; Geleijnse, J.M. Diet and Kidney Function: A Literature Review. Curr. Hypertens. Rep. 2020, 22, 14. [Google Scholar] [CrossRef]
- Hines, C.J.; Christianson, A.L.; Jackson, M.V.; Ye, X.; Pretty, J.R.; Arnold, J.E.; Calafat, A.M. An Evaluation of the Relationship among Urine, Air, and Hand Measures of Exposure to Bisphenol A [BPA] in US Manufacturing Workers. Ann. Work. Expo. Health 2018, 62, 840–851. [Google Scholar] [CrossRef]
- Quiroga, B. Strategies to Protect Dialysis Patients against Bisphenol A. Biomolecules 2021, 11, 1375. [Google Scholar] [CrossRef]
- Jiang, W.; Zhao, H.; Zhang, L.; Wu, B.; Zha, Z. Maintenance of mitochondrial function by astaxanthin protects against bisphenol A-induced kidney toxicity in rats. Biomed. Pharmacother. 2020, 121, 109629. [Google Scholar] [CrossRef]
- Janardhan, K.S.; Jensen, H.; Clayton, N.P.; Herbert, R.A. Immunohistochemistry in Investigative and Toxicologic Pathology. Toxicol. Pathol. 2018, 46, 488–510. [Google Scholar] [CrossRef]
- Eid, R.A.; Bin-Meferij, M.M.; El-Kott, A.F.; Eleawa, S.M.; Zaki, M.S.A.; Al-Shraim, M.; El-Sayed, F.; Eldeen, M.A.; Alkhateeb, M.A.; Alharbi, S.A.; et al. Exendin-4 Protects Against Myocardial Ischemia-Reperfusion Injury by Upregulation of SIRT1 and SIRT3 and Activation of AMPK. J. Cardiovasc. Transl. Res. 2020, 14, 619–635. [Google Scholar] [CrossRef]
- Katoh, H.; Negishi, M. RhoG activates Rac1 by direct interaction with the Dock180-binding protein Elmo. Nature 2003, 424, 461–464. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Li, H.; Robertson, A.D.; Jensen, J.H. Very fast empirical prediction and rationalization of protein pKa values. Proteins Struct. Funct. Genet. 2005, 61, 704–721. [Google Scholar] [CrossRef]
- Evans, D.A. History of the Harvard ChemDraw Project. Angew. Chem.-Int. Ed. 2014, 53, 11140–11145. [Google Scholar] [CrossRef]
- Hospital, A.; Goñi, J.R.; Orozco, M.; Gelpí, J.L. Molecular dynamics simulations: Advances and applications. Adv. Appl. Bioinform. Chem. 2015, 8, 37–47. [Google Scholar]
- Lee, T.-S.; Cerutti, D.S.; Mermelstein, D.; Lin, C.; LeGrand, S.; Giese, T.J.; Roitberg, A.; Case, D.A.; Walker, R.C.; York, D.M. GPU-Accelerated Molecular Dynamics and Free Energy Methods in Amber18: Performance Enhancements and New Features. J. Chem. Inf. Model. 2018, 58, 2043–2050. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E., III. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef]
- Seifert, E. OriginPro 9.1: Scientific Data Analysis and Graphing Software—Software Review. J. Chem. Inf. Model. 2014, 54, 1552. [Google Scholar] [CrossRef]
- Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Huang, C.C.; Ferrin, T.E. Tools for integrated sequence-structure analysis with UCSF Chimera. BMC Bioinform. 2006, 7, 339. [Google Scholar] [CrossRef]
- Hayes, J.M.; Archontis, G. MM-GB[PB]SA Calculations of Protein-Ligand Binding Free Energies. In Molecular Dynamics—Studies of Synthetic and Biological Macromolecules; InTech: London, UK, 2012. [Google Scholar]
- Ylilauri, M.; Pentikäinen, O.T. MMGBSA As a Tool To Understand the Binding Affinities of Filamin–Peptide Interactions. J. Chem. Inf. Model. 2013, 53, 2626–2633. [Google Scholar] [CrossRef]
- Kollman, P.A.; Massova, I.; Reyes, C.; Kuhn, B.; Huo, S.; Chong, L.; Lee, M.; Lee, T.; Duan, Y.; Wang, W.; et al. Calculating Structures and Free Energies of Complex Molecules: Combining Molecular Mechanics and Continuum Models. Accounts Chem. Res. 2000, 33, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Wang, J.; Li, Y.; Wang, W. Assessing the Performance of the MM/PBSA and MM/GBSA Methods. 1. The Accuracy of Binding Free Energy Calculations Based on Molecular Dynamics Simulations. J. Chem. Inf. Model. 2011, 51, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Sitkoff, D.; Sharp, K.A.; Honig, B. Accurate Calculation of Hydration Free Energies Using Macroscopic Solvent Models. J. Phys. Chem. 1994, 98, 1978–1988. [Google Scholar] [CrossRef]
- Greenidge, P.A.; Kramer, C.; Mozziconacci, J.-C.; Wolf, R.M. MM/GBSA Binding Energy Prediction on the PDBbind Data Set: Successes, Failures, and Directions for Further Improvement. J. Chem. Inf. Model. 2013, 53, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, S.; Eisvand, F.; Hadizadeh, F.; Mosaffa, F.; Ghasemi, A.; Ghodsi, R. Design, synthesis and biological evaluation of novel 5,6,7-trimethoxy-N-aryl-2-styrylquinolin-4-amines as potential anticancer agents and tubulin polymerization inhibitors. Bioorganic Chem. 2020, 98, 103711. [Google Scholar] [CrossRef]
- Machaba, K.E.; Mhlongo, N.N.; Soliman, M.E.S. Induced Mutation Proves a Potential Target for TB Therapy: A Molecular Dynamics Study on LprG. Cell Biochem. Biophys. 2018, 76, 345–356. [Google Scholar] [CrossRef]
- Cournia, Z.; Allen, B.; Sherman, W. Relative Binding Free Energy Calculations in Drug Discovery: Recent Advances and Practical Considerations. J. Chem. Inf. Model. 2017, 57, 2911–2937. [Google Scholar] [CrossRef]
- Franzin, R.; Stasi, A.; Fiorentino, M.; Stallone, G.; Cantaluppi, V.; Gesualdo, L.; Castellano, G. Inflammaging and Complement System: A Link Between Acute Kidney Injury and Chronic Graft Damage. Front. Immunol. 2020, 11, 734. [Google Scholar] [CrossRef]
- Mitrakou, A. Kidney: Its impact on glucose homeostasis and hormonal regulation. Diabetes Res. Clin. Pract. 2011, 93, S66–S72. [Google Scholar] [CrossRef]
- Anet, A.; Olakkaran, S.; Purayil, A.K.; Puttaswamygowda, G.H. Bisphenol A induced oxidative stress mediated genotoxicity in Drosophila melanogaster. J. Hazard. Mater. 2019, 370, 42–53. [Google Scholar] [CrossRef]
- Beltifa, A.; Feriani, A.; Macherki, M.; Ghorbel, A.; Ghazouani, L.; Di Bella, G.; Sire, O.; Van Loco, J.; Reyns, T.; Ben Mansour, H. Persistent plasticizers and bisphenol in the cheese of Tunisian markets induced biochemical and histopathological alterations in male BALB/c mice. Environ. Sci. Pollut. Res. 2018, 25, 6545–6557. [Google Scholar] [CrossRef]
- Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and other Nutrients from Haematococcus pluvialis—Multifunctional Applications. Mar. Drugs 2020, 18, 459. [Google Scholar] [CrossRef]
- Alugoju, P.; Swamy, V.K.D.K.; Anthikapalli, N.V.A.; Tencomnao, T. Health benefits of astaxanthin against age-related diseases of multiple organs: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2022, 1–66. [Google Scholar] [CrossRef]
- Arslan, E.; Turk, H.; Caglayan, M.; Turkmenoglu, T.T.; Gonel, A.; Tayman, C. Protective Effects of Astaxanthin on Nephrotoxicity in Rats with Induced Renovascular Occlusion. Comb. Chem. High Throughput Screen. 2021, 24, 1236–1242. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Y.; Zou, M.; Qi, X.; Xu, S. Bisphenol A aggravates renal apoptosis and necroptosis in selenium-deficient chickens via oxidative stress and PI3K/AKT pathway. J. Cell Physiol. 2022, 237, 3292–3304. [Google Scholar] [CrossRef]
- Li, Y.; Dong, Z.; Liu, S.; Gao, F.; Zhang, J.; Peng, Z.; Wang, L.; Pan, X. Astaxanthin improves the development of the follicles and oocytes through alleviating oxidative stress induced by BPA in cultured follicles. Sci. Rep. 2022, 12, 7853. [Google Scholar] [CrossRef]
- Lim, S.R.; Kim, D.W.; Sung, J.; Kim, T.H.; Choi, C.H.; Lee, S.J. Astaxanthin inhibits autophagic cell death induced by bisphenol a in human dermal fibroblasts. Antioxidants 2021, 10, 1273. [Google Scholar] [CrossRef]
- Li, L.; Chen, Y.; Jiao, D.; Yang, S.; Li, L.; Li, P. Protective effect of astaxanthin on ochratoxin A-induced kidney injury to mice by regulating oxidative stress-related Nrf2/Keap1 pathway. Molecules 2020, 25, 1386. [Google Scholar] [CrossRef]
- Barbonetti, A.; Castellini, C.; di Giammarco, N.; Santilli, G.; Francavilla, S.; Francavilla, F. In vitro exposure of human spermatozoa to bisphenol A induces pro-oxidative/apoptotic mitochondrial dysfunction. Reprod. Toxicol. 2016, 66, 61–67. [Google Scholar] [CrossRef]
- Chang, M.X.; Xiong, F. Astaxanthin and its Effects in Inflammatory Responses and Inflammation-Associated Diseases: Recent Advances and Future Directions. Molecules 2020, 25, 5342. [Google Scholar] [CrossRef]
- Ratliff, B.B.; Abdulmahdi, W.; Pawar, R.; Wolin, M.S. Oxidant mechanisms in renal injury and disease. Antioxid. Redox Signal. 2016, 25, 119–146. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Yi, H.; Huang, L.; An, Q.; Wang, H. Long-term arsenite exposure induces testicular toxicity by redox imbalance, G2/M cell arrest and apoptosis in mice. Toxicology 2019, 411, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Qu, W.; Wang, K.; Chen, S.; Zhang, L.; Wu, D.; Chen, Z. Bisphenol A inhibits mucin 2 secretion in intestinal goblet cells through mitochondrial dysfunction and oxidative stress. Biomed. Pharmacother. 2019, 111, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.; Taivassalo, T.; Gouspillou, G.; Hepple, R.T. Mitochondria: Isolation, structure and function. J. Physiol. 2011, 589, 4413–4421. [Google Scholar] [CrossRef]
- Blokhina, O.; Fagerstedt, K.V. Oxidative metabolism, ROS and NO under oxygen deprivation. Plant Physiol. Biochem. 2010, 48, 359–373. [Google Scholar] [CrossRef]
- Morgan, E.T. Regulation of cytochrome p450 by inflammatory mediators: Why and how? Drug Metab. Dispos. 2001, 29, 207–212. [Google Scholar]
- Guengerich, F.P. Common and Uncommon Cytochrome P450 Reactions Related to Metabolism and Chemical Toxicity. Chem. Res. Toxicol. 2001, 14, 611–650. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Energy Components (kcal/mol) | |||||
|---|---|---|---|---|---|
| Complex | ΔEvdW | ΔEelec | ΔGgas | ΔGsolv | ΔGbind |
| ATX | −81.27 ± 0.30 | −42.58 ± 0.43 | −123.85 ± 0.51 | 44.21 ± 0.27 | −79.63 ± 0.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
A. Eid, R.; Alaa Edeen, M.; Soltan, M.A.; Al-Shraim, M.; Samir A. Zaki, M.; M. Al-Qahtani, S.; Fayad, E.; T. Salem, E.; K. Abdulsahib, W.; Emam, H.; et al. Integration of Ultrastructural and Computational Approaches Reveals the Protective Effect of Astaxanthin against BPA-Induced Nephrotoxicity. Biomedicines 2023, 11, 421. https://doi.org/10.3390/biomedicines11020421
A. Eid R, Alaa Edeen M, Soltan MA, Al-Shraim M, Samir A. Zaki M, M. Al-Qahtani S, Fayad E, T. Salem E, K. Abdulsahib W, Emam H, et al. Integration of Ultrastructural and Computational Approaches Reveals the Protective Effect of Astaxanthin against BPA-Induced Nephrotoxicity. Biomedicines. 2023; 11(2):421. https://doi.org/10.3390/biomedicines11020421
Chicago/Turabian StyleA. Eid, Refaat, Muhammad Alaa Edeen, Mohamed A. Soltan, Mubarak Al-Shraim, Mohamed Samir A. Zaki, Saleh M. Al-Qahtani, Eman Fayad, Eman T. Salem, Waleed K. Abdulsahib, Hebatallah Emam, and et al. 2023. "Integration of Ultrastructural and Computational Approaches Reveals the Protective Effect of Astaxanthin against BPA-Induced Nephrotoxicity" Biomedicines 11, no. 2: 421. https://doi.org/10.3390/biomedicines11020421
APA StyleA. Eid, R., Alaa Edeen, M., Soltan, M. A., Al-Shraim, M., Samir A. Zaki, M., M. Al-Qahtani, S., Fayad, E., T. Salem, E., K. Abdulsahib, W., Emam, H., & M. Hassan, H. (2023). Integration of Ultrastructural and Computational Approaches Reveals the Protective Effect of Astaxanthin against BPA-Induced Nephrotoxicity. Biomedicines, 11(2), 421. https://doi.org/10.3390/biomedicines11020421