
TMPRSS2 Impacts Cytokine Expression in Murine Dendritic Cells
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Isolation of Monocytes from Mouse Bone Marrow, Differentiation to Dendritic Cells and Treatment
2.3. Verification of Differentiation
2.4. Analysis of Secreted Cytokines
2.5. RNA Isolation, RT-PCR Analysis and qRT-PCR
2.6. Statistics
3. Results
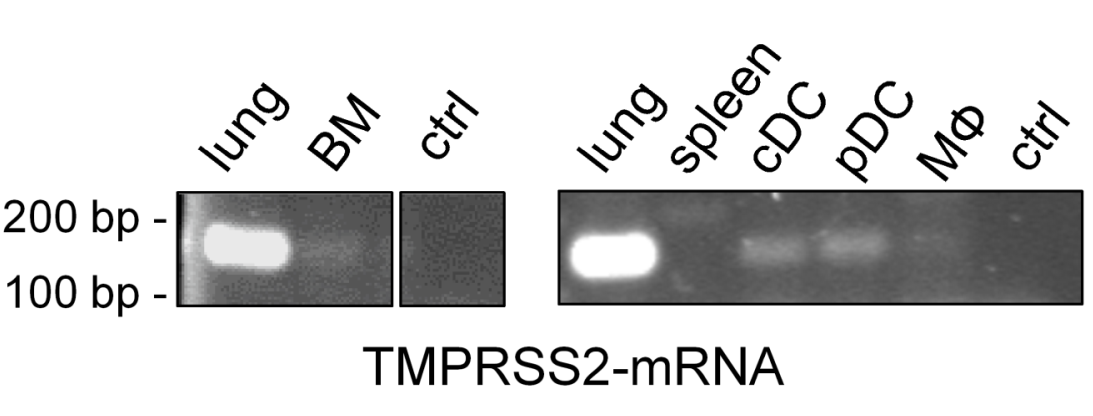
3.1. TMPRSS2 Is Expressed in Murine Dendritic Cells
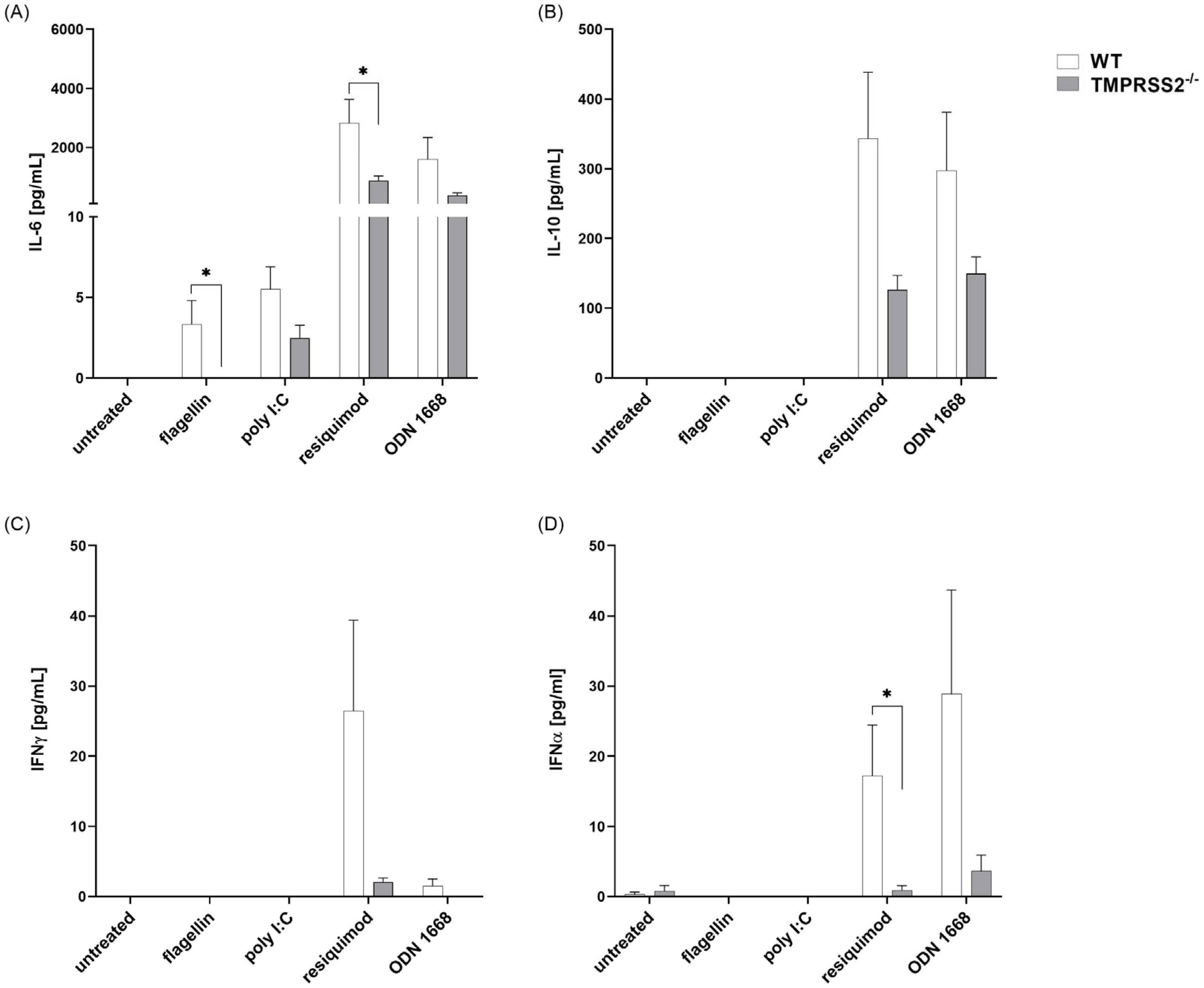
3.2. TMPRSS2 Regulates the Secretion of Proinflammatory Cytokines
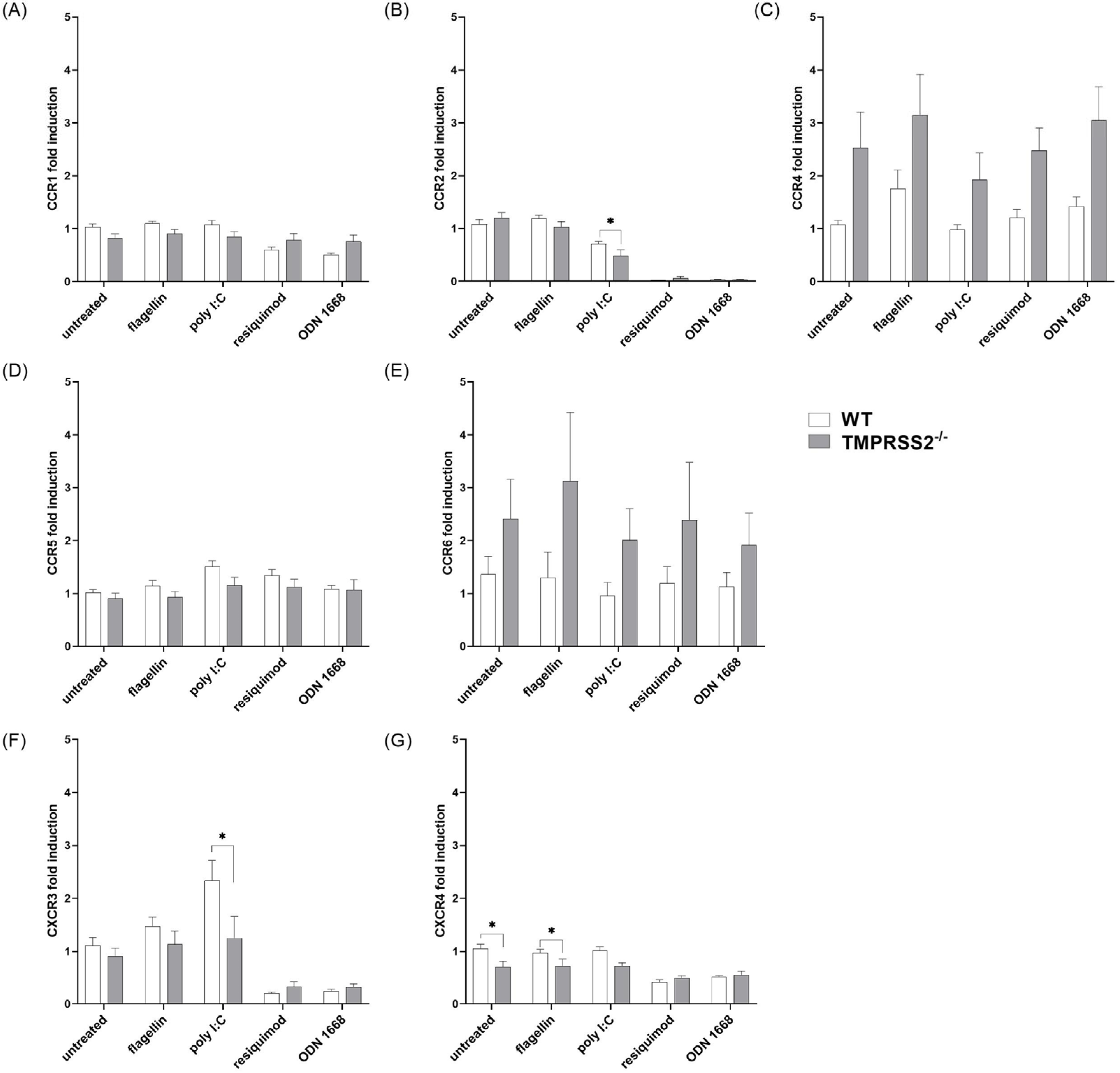
3.3. TMPRSS2 Expression Has Only Minor Effects on Chemokine Receptor Expression in cDC
3.4. TMPRSS2−/− pDCs Show Lower Cytokine Secretion Levels after Resiquimod and ODN 1668 Stimulation Compared to WT
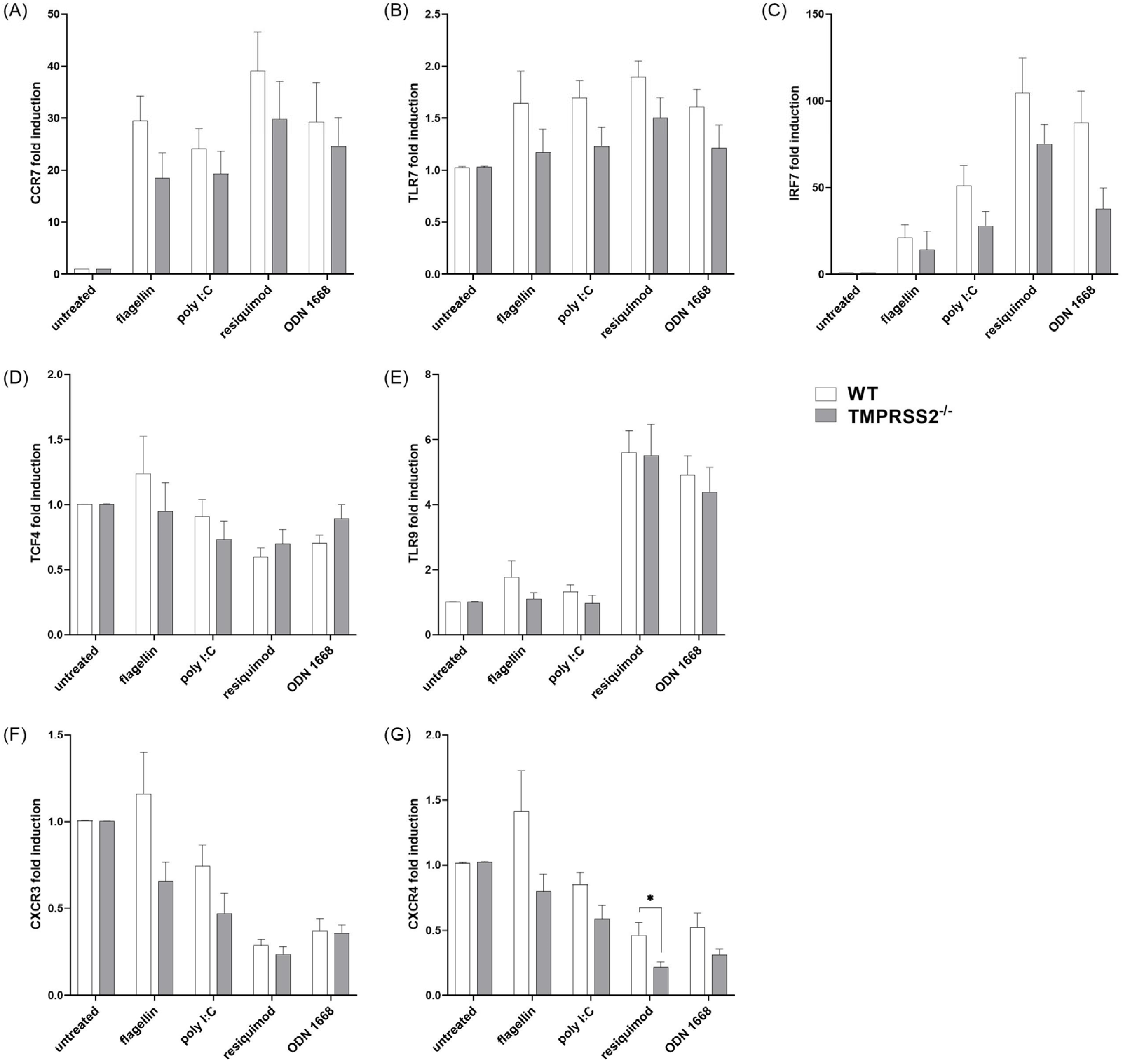
3.5. TMPRSS2−/− pDCs Show Lower Expression of Motility Relevant Receptors and Important Type I IFN Inducing Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bestle, D.; Heindl, M.R.; Limburg, H.; van Lam van, T.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; et al. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 3, e202000786. [Google Scholar] [CrossRef] [PubMed]
- Limburg, H.; Harbig, A.; Bestle, D.; Stein, D.A.; Moulton, H.M.; Jaeger, J.; Janga, H.; Hardes, K.; Koepke, J.; Schulte, L.; et al. TMPRSS2 Is the Major Activating Protease of Influenza A Virus in Primary Human Airway Cells and Influenza B Virus in Human Type II Pneumocytes. J. Virol. 2019, 93, e00649-19. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Heinlein, C.; Hackman, R.C.; Nelson, P.S. Phenotypic analysis of mice lacking the Tmprss2-encoded protease. Mol. Cell. Biol. 2006, 26, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Böttcher-Friebertshäuser, E. Membrane-Anchored Serine Proteases: Host Cell Factors in Proteolytic Activation of Viral Glycoproteins. In Activation of Viruses by Host Proteases; Böttcher-Friebertshäuser, E., Garten, W., Klenk, H.D., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 153–203. ISBN 978-3-319-75473-4. [Google Scholar]
- Lucas, J.M.; True, L.; Hawley, S.; Matsumura, M.; Morrissey, C.; Vessella, R.; Nelson, P.S. The androgen-regulated type II serine protease TMPRSS2 is differentially expressed and mislocalized in prostate adenocarcinoma. J. Pathol. 2008, 215, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.-J.; Huang, C.-C.; Lin, H.-Y.; Juan, C.-P.; Lan, S.-W.; Shyu, H.-Y.; Wu, S.-R.; Hsiao, P.-W.; Huang, H.-P.; Shun, C.-T.; et al. Androgen-Induced TMPRSS2 Activates Matriptase and Promotes Extracellular Matrix Degradation, Prostate Cancer Cell Invasion, Tumor Growth, and Metastasis. Cancer Res. 2015, 75, 2949–2960. [Google Scholar] [CrossRef] [PubMed]
- Devaprasad, A.; Pandit, A. Enrichment of SARS-CoV-2 Entry Factors and Interacting Intracellular Genes in Tissue and Circulating Immune Cells. Viruses 2021, 13, 1757. [Google Scholar] [CrossRef]
- Zankharia, U.; Yadav, A.; Yi, Y.; Hahn, B.H.; Collman, R.G. Highly restricted SARS-CoV-2 receptor expression and resistance to infection by primary human monocytes and monocyte-derived macrophages. J. Leukoc. Biol. 2022, 112, 569–576. [Google Scholar] [CrossRef]
- Liu, X.; Liu, B.; Shang, Y.; Cao, P.; Hou, J.; Chen, F.; Zhang, B.; Fan, Y.; Tan, K. Decreased TMPRSS2 expression by SARS-CoV-2 predicts the poor prognosis of lung cancer patients through metabolic pathways and immune infiltration. Aging 2022, 14, 73–108. [Google Scholar] [CrossRef]
- Bahmad, H.F.; Abou-Kheir, W. Crosstalk between COVID-19 and prostate cancer. Prostate Cancer Prostatic Dis. 2020, 23, 561–563. [Google Scholar] [CrossRef]
- Gkogkou, E.; Barnasas, G.; Vougas, K.; Trougakos, I.P. Expression profiling meta-analysis of ACE2 and TMPRSS2, the putative anti-inflammatory receptor and priming protease of SARS-CoV-2 in human cells, and identification of putative modulators. Redox Biol. 2020, 36, 101615. [Google Scholar] [CrossRef]
- Pandey, R.K.; Srivastava, A.; Singh, P.P.; Chaubey, G. Genetic association of TMPRSS2 rs2070788 polymorphism with COVID-19 case fatality rate among Indian populations. Infect. Genet. Evol. 2022, 98, 105206. [Google Scholar] [CrossRef]
- Schönfelder, K.; Breuckmann, K.; Elsner, C.; Dittmer, U.; Fistera, D.; Herbstreit, F.; Risse, J.; Schmidt, K.; Sutharsan, S.; Taube, C.; et al. Transmembrane serine protease 2 Polymorphisms and Susceptibility to Severe Acute Respiratory Syndrome Coronavirus Type 2 Infection: A German Case-Control Study. Front. Genet. 2021, 12, 667231. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Zhou, J.; To, K.K.-W.; Chu, H.; Li, C.; Wang, D.; Yang, D.; Zheng, S.; Hao, K.; Bossé, Y.; et al. Identification of TMPRSS2 as a Susceptibility Gene for Severe 2009 Pandemic A(H1N1) Influenza and A(H7N9) Influenza. J. Infect. Dis. 2015, 212, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Iwata-Yoshikawa, N.; Okamura, T.; Shimizu, Y.; Hasegawa, H.; Takeda, M.; Nagata, N. TMPRSS2 Contributes to Virus Spread and Immunopathology in the Airways of Murine Models after Coronavirus Infection. J. Virol. 2019, 93, e01815-18. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.; Yang, J.; Deng, H.; Chen, D.; Yang, X.; Tang, Z.-H. Depletion and Dysfunction of Dendritic Cells: Understanding SARS-CoV-2 Infection. Front. Immunol. 2022, 13, 843342. [Google Scholar] [CrossRef]
- Lambertz, R.L.O.; Pippel, J.; Gerhauser, I.; Kollmus, H.; Anhlan, D.; Hrincius, E.R.; Krausze, J.; Kühn, N.; Schughart, K. Exchange of amino acids in the H1-haemagglutinin to H3 residues is required for efficient influenza A virus replication and pathology in Tmprss2 knock-out mice. J. Gen. Virol. 2018, 99, 1187–1198. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Quan, N. Immune Cell Isolation from Mouse Femur Bone Marrow. Bio Protoc. 2015, 5, e1631. [Google Scholar] [CrossRef]
- Muñoz, J.J.; Anauate, A.C.; Amaral, A.G.; Ferreira, F.M.; Watanabe, E.H.; Meca, R.; Ormanji, M.S.; Boim, M.A.; Onuchic, L.F.; Heilberg, I.P. Ppia is the most stable housekeeping gene for qRT-PCR normalization in kidneys of three Pkd1-deficient mouse models. Sci. Rep. 2021, 11, 19798. [Google Scholar] [CrossRef]
- Curtis, K.M.; Gomez, L.A.; Rios, C.; Garbayo, E.; Raval, A.P.; Perez-Pinzon, M.A.; Schiller, P.C. EF1alpha and RPL13a represent normalization genes suitable for RT-qPCR analysis of bone marrow derived mesenchymal stem cells. BMC Mol. Biol. 2010, 11, 61. [Google Scholar] [CrossRef]
- Lam, D.K.; Dang, D.; Flynn, A.N.; Hardt, M.; Schmidt, B.L. TMPRSS2, a novel membrane-anchored mediator in cancer pain. Pain 2015, 156, 923–930. [Google Scholar] [CrossRef]
- Wettstein, L.; Kirchhoff, F.; Münch, J. The Transmembrane Protease TMPRSS2 as a Therapeutic Target for COVID-19 Treatment. Int. J. Mol. Sci. 2022, 23, 1351. [Google Scholar] [CrossRef] [PubMed]
- Lester, S.N.; Li, K. Toll-like receptors in antiviral innate immunity. J. Mol. Biol. 2014, 426, 1246–1264. [Google Scholar] [CrossRef]
- Schülke, S. Induction of Interleukin-10 Producing Dendritic Cells as a Tool to Suppress Allergen-Specific T Helper 2 Responses. Front. Immunol. 2018, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-D.; Cheng, M.; Shang, P.-P.; Yang, Y.-Q. Role of IL-6 in dendritic cell functions. J. Leukoc. Biol. 2022, 111, 695–709. [Google Scholar] [CrossRef]
- Ma, X.; Yan, W.; Zheng, H.; Du, Q.; Zhang, L.; Ban, Y.; Li, N.; Wei, F. Regulation of IL-10 and IL-12 production and function in macrophages and dendritic cells. F1000Research 2015, 4, 1465. [Google Scholar] [CrossRef]
- Maroof, A.; Penny, M.; Kingston, R.; Murray, C.; Islam, S.; Bedford, P.A.; Knight, S.C. Interleukin-4 can induce interleukin-4 production in dendritic cells. Immunology 2006, 117, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Zhang, M.; Wang, J.; Wang, Q.; Xia, D.; Sun, W.; Zhang, L.; Yu, H.; Liu, Y.; Cao, X. Interferon-gamma is an autocrine mediator for dendritic cell maturation. Immunol. Lett. 2004, 94, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Berger, A. Th1 and Th2 responses: What are they? BMJ 2000, 321, 424. [Google Scholar] [CrossRef]
- Pavel, A.B.; Glickman, J.W.; Michels, J.R.; Kim-Schulze, S.; Miller, R.L.; Guttman-Yassky, E. Th2/Th1 Cytokine Imbalance Is Associated With Higher COVID-19 Risk Mortality. Front. Genet. 2021, 12, 706902. [Google Scholar] [CrossRef]
- Lambertz, R.L.O.; Gerhauser, I.; Nehlmeier, I.; Leist, S.R.; Kollmus, H.; Pöhlmann, S.; Schughart, K. Tmprss2 knock-out mice are resistant to H10 influenza A virus pathogenesis. J. Gen. Virol. 2019, 100, 1073–1078. [Google Scholar] [CrossRef]
- Sichien, D.; Lambrecht, B.N.; Guilliams, M.; Scott, C.L. Development of conventional dendritic cells: From common bone marrow progenitors to multiple subsets in peripheral tissues. Mucosal Immunol. 2017, 10, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.R.; Jung, D.; Gu, G.J.; Seok, S.H. GM-CSF Grown Bone Marrow Derived Cells Are Composed of Phenotypically Different Dendritic Cells and Macrophages. Mol. Cells 2016, 39, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Helft, J.; Böttcher, J.; Chakravarty, P.; Zelenay, S.; Huotari, J.; Schraml, B.U.; Goubau, D.; Reis e Sousa, C. GM-CSF Mouse Bone Marrow Cultures Comprise a Heterogeneous Population of CD11c(+)MHCII(+) Macrophages and Dendritic Cells. Immunity 2015, 42, 1197–1211. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Shinoda, M.; Nishizaki, Y.; Shiraki, K.; Hirai, Y.; Kichikawa, Y.; Tsushima, K.; Sinkai, M.; Komura, N.; Yoshida, K.; et al. A multicenter, double-blind, randomized, parallel-group, placebo-controlled study to evaluate the efficacy and safety of camostat mesilate in patients with COVID-19 (CANDLE study). BMC Med. 2022, 20, 342. [Google Scholar] [CrossRef]
- Gunst, J.D.; Staerke, N.B.; Pahus, M.H.; Kristensen, L.H.; Bodilsen, J.; Lohse, N.; Dalgaard, L.S.; Brønnum, D.; Fröbert, O.; Hønge, B.; et al. Efficacy of the TMPRSS2 inhibitor camostat mesilate in patients hospitalized with COVID-19-a double-blind randomized controlled trial. EClinicalMedicine 2021, 35, 100849. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| RPL13A (reference) | TGCCCCACAAGACCAAGAGAG | TGAGAGCAGCAGGGACCACC |
| PPIA (reference) | GCTGGACCAAACACAAACGG | GCCATTCCTGGACCCAAAAC |
| CCR1 | CCATCATCATACAGGAAGCCAAG | TCTGTGAAATCTGAAATCTCCATCC |
| CCR2 | ATCCACGGCATACTATCAACATCTC | GACAAGGCTCACCATCATCGTAG |
| CCR4 | ATCACTTTCAGAAGAGCAAGGCAG | CAGTTTCATCCTGGGTGGTGTC |
| CCR5 | CAGAGGAGGTGAGACATCCGTTC | TTTTGGCAGGGTGCTGACATAC |
| CCR6 | GTCCTGGGTGCTTAGAGACT | CAAGGGCATGACTCCTTTCCA |
| CCR7 | CCAGCAAGCAGCTCAACATT | GCCGATGAAGGCATACAAGA |
| CXCR3 | TGAGGTTAGTGAACGTCAAGTGCT | CCCCATAATCGTAGGGAGAGGT |
| CXCR4 | CACGGCTGTAGAGCGAGTGTTG | ATGAAGTAGATGGTGGGCAGGAAG |
| TLR7 | TTCTCTTCAGCATGTGCCCC | GACATACCCCTTGACACGCA |
| TLR9 | GAGAAGCAACCCTCTGCACT | ATCGAACACCACGAAGGCAT |
| IRF7 | TCCAGTTGATCCGCATAAGGT | CTTCCCTATTTTCCGTGGCTG |
| TCL4 (B2-2) | CCAGGAACCCTTTCGCCCACCAAAC | TGCTGGCTGCTGGCTTGGAGGAA |
| TMPRSS2 | AAGGGATACCAGACCAGAGC | GCGAACCCCGCATTCTATAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gunne, S.; Schwerdtner, M.; Henke, M.; Schneider, A.-K.; Keutmann, L.; Böttcher-Friebertshäuser, E.; Schiffmann, S. TMPRSS2 Impacts Cytokine Expression in Murine Dendritic Cells. Biomedicines 2023, 11, 419. https://doi.org/10.3390/biomedicines11020419
Gunne S, Schwerdtner M, Henke M, Schneider A-K, Keutmann L, Böttcher-Friebertshäuser E, Schiffmann S. TMPRSS2 Impacts Cytokine Expression in Murine Dendritic Cells. Biomedicines. 2023; 11(2):419. https://doi.org/10.3390/biomedicines11020419
Chicago/Turabian StyleGunne, Sandra, Marie Schwerdtner, Marina Henke, Ann-Kathrin Schneider, Lucas Keutmann, Eva Böttcher-Friebertshäuser, and Susanne Schiffmann. 2023. "TMPRSS2 Impacts Cytokine Expression in Murine Dendritic Cells" Biomedicines 11, no. 2: 419. https://doi.org/10.3390/biomedicines11020419
APA StyleGunne, S., Schwerdtner, M., Henke, M., Schneider, A.-K., Keutmann, L., Böttcher-Friebertshäuser, E., & Schiffmann, S. (2023). TMPRSS2 Impacts Cytokine Expression in Murine Dendritic Cells. Biomedicines, 11(2), 419. https://doi.org/10.3390/biomedicines11020419