
Quantitative Structure-Activity Relationship of Fluorescent Probes and Their Intracellular Localizations



Abstract
1. Introduction
2. Materials and Methods
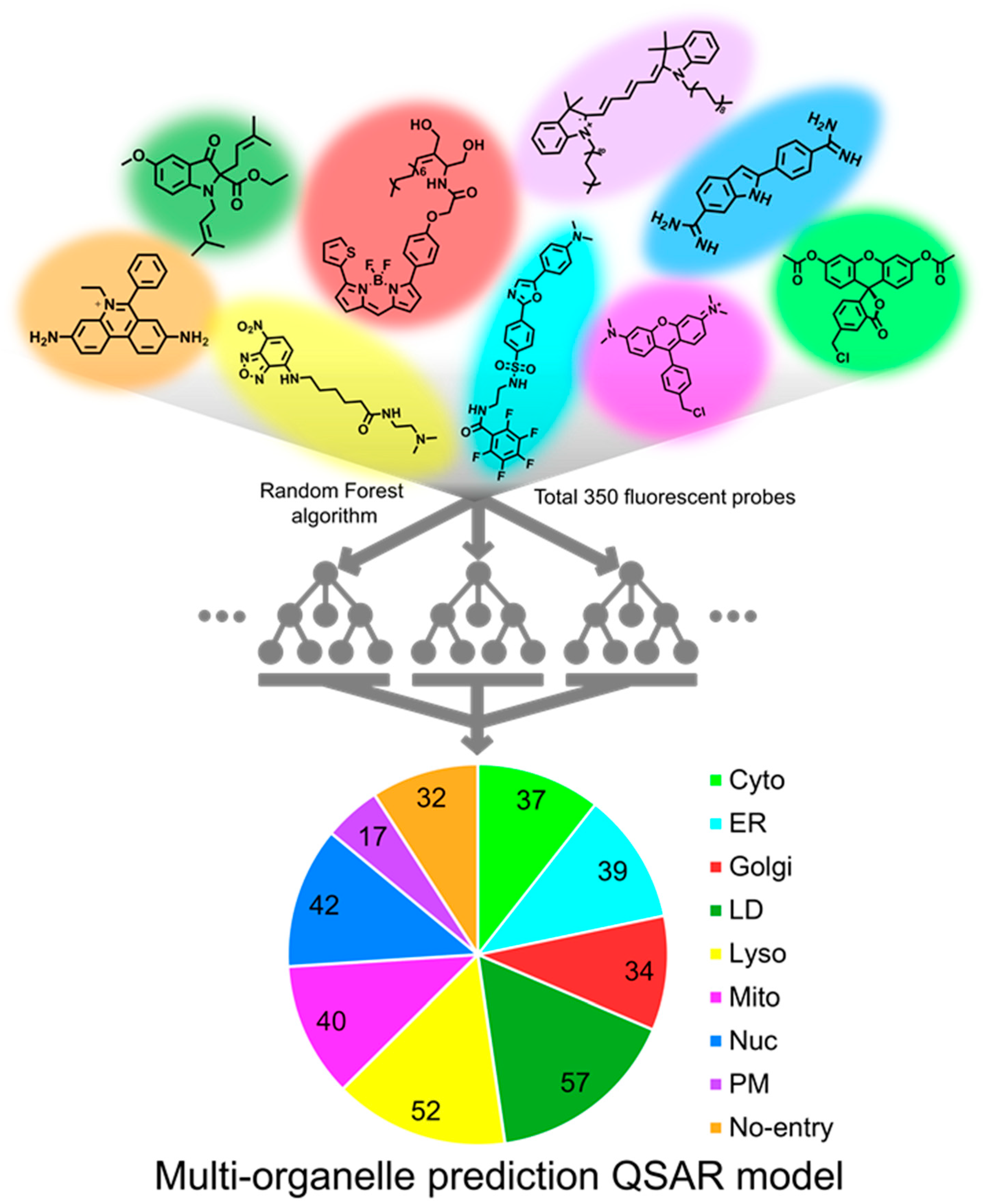
2.1. Dataset
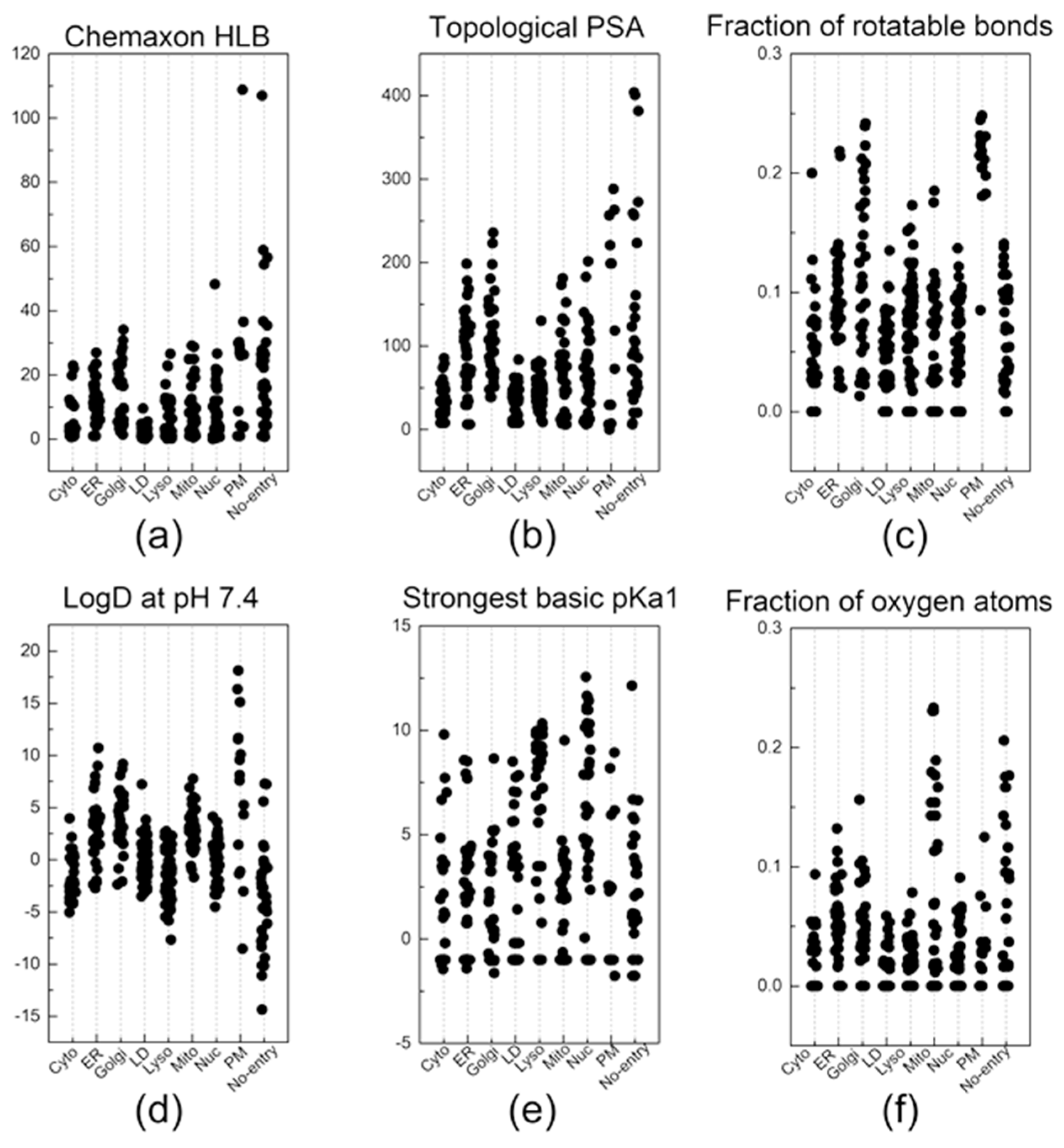
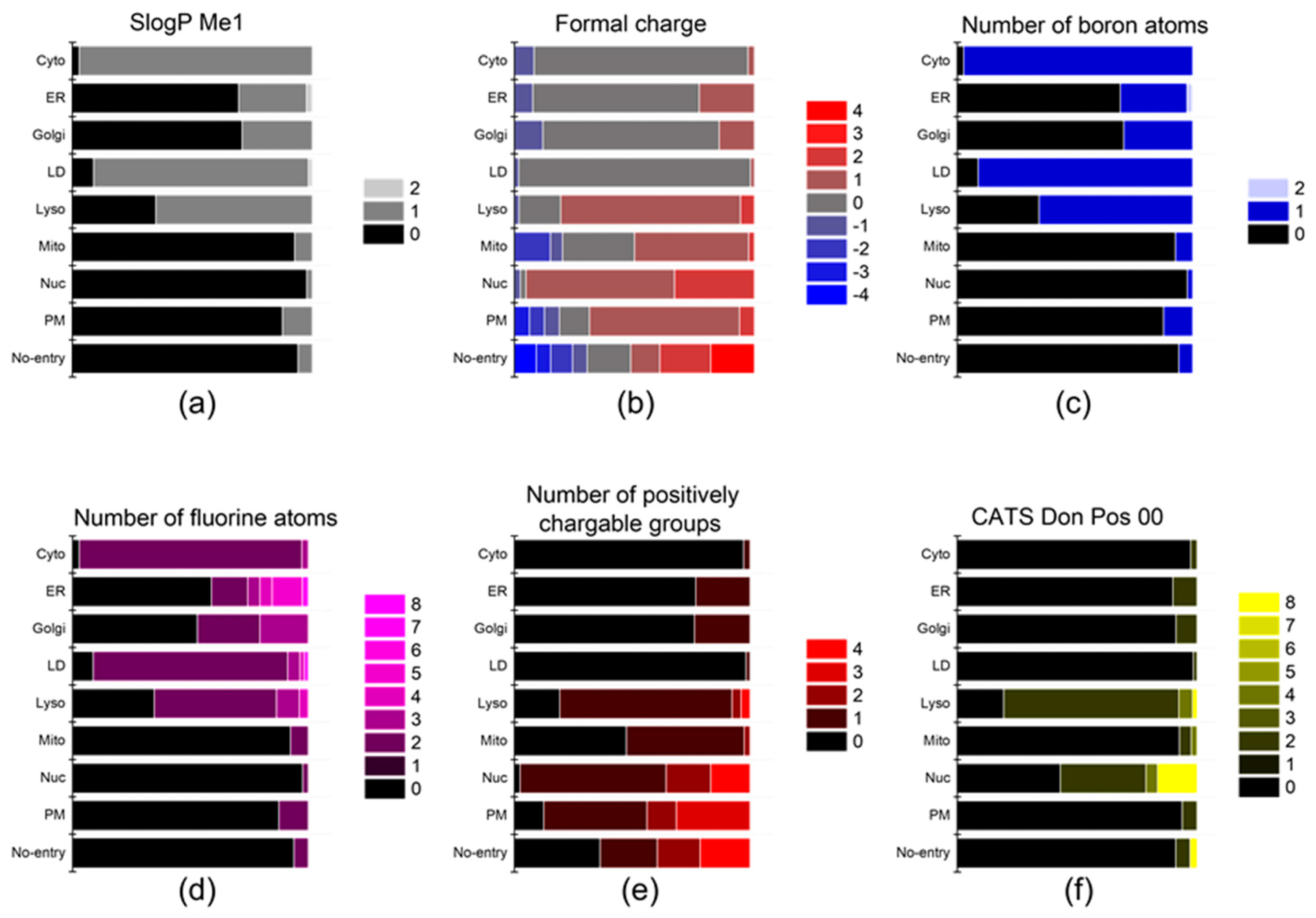
2.2. Descriptors
2.3. Machine Learning Techniques
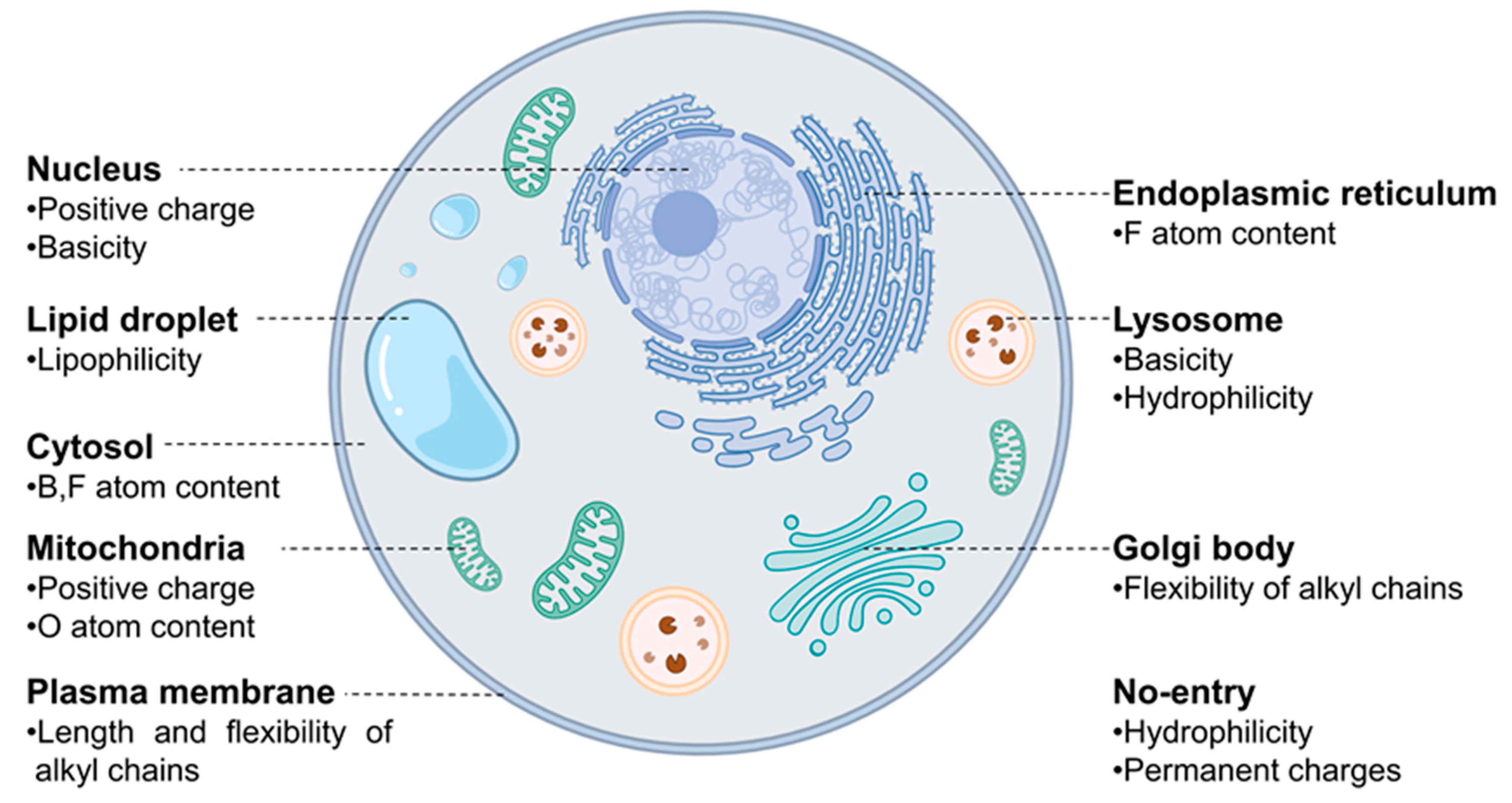
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Casasampere, M.; Izquierdo, E.; Casas, J.; Abad, J.L.; Liu, X.; Xu, R.; Mao, C.; Chang, Y.-T.; Delgado, A.; Fabrias, G. Click and count: Specific detection of acid ceramidase activity in live cells. Chem. Sci. 2020, 11, 13044–13051. [Google Scholar] [CrossRef]
- Choi, Y.-K.; Kim, J.-J.; Chang, Y.-T. Holding-Oriented versus Gating-Oriented Live-Cell Distinction: Highlighting the Role of Transporters in Cell Imaging Probe Development. Acc. Chem. Res. 2019, 52, 3097–3107. [Google Scholar] [CrossRef]
- Gao, P.; Pan, W.; Li, N.; Tang, B. Fluorescent probes for organelle-targeted bioactive species imaging. Chem. Sci. 2019, 10, 6035–6071. [Google Scholar] [CrossRef]
- Kwon, H.-Y.; Kumar Das, R.; Jung, G.T.; Lee, H.-G.; Lee, S.H.; Berry, S.N.; Tan, J.K.S.; Park, S.; Yang, J.-S.; Park, S.; et al. Lipid-Oriented Live-Cell Distinction of B and T Lymphocytes. J. Am. Chem. Soc. 2021, 143, 5836–5844. [Google Scholar] [CrossRef]
- Liu, X.; Chang, Y.-T. Fluorescent probe strategy for live cell distinction. Chem. Soc. Rev. 2022, 51, 1573–1591. [Google Scholar] [CrossRef]
- Yasuda, M.; Ma, Y.; Okabe, S.; Wakabayashi, Y.; Su, D.; Chang, Y.-T.; Seimiya, H.; Tera, M.; Nagasawa, K. Target identification of a macrocyclic hexaoxazole G-quadruplex ligand using post-target-binding visualization. Chem. Commun. 2020, 56, 12905–12908. [Google Scholar] [CrossRef]
- Zhu, H.; Fan, J.; Du, J.; Peng, X. Fluorescent Probes for Sensing and Imaging within Specific Cellular Organelles. Acc. Chem. Res. 2016, 49, 2115–2126. [Google Scholar] [CrossRef]
- Lim, S.; Haque, M.M.; Su, D.; Kim, D.; Lee, J.-S.; Chang, Y.-T.; Kim, Y.K. Development of a BODIPY-based fluorescent probe for imaging pathological tau aggregates in live cells. Chem. Commun. 2017, 53, 1607–1610. [Google Scholar] [CrossRef]
- Horobin, R.; Stockert, J.; Rashid-Doubell, F. Uptake and localization mechanisms of fluorescent and colored lipid probes. Part 2. QSAR models that predict localization of fluorescent probes used to identify (“specifically stain”) various biomembranes and membranous organelles. Biotech. Histochem. 2015, 90, 241–254. [Google Scholar] [CrossRef]
- Horobin, R.; Stockert, J.; Rashid-Doubell, F. Uptake and localization mechanisms of fluorescent and colored lipid probes. Part 3. Protocols for predicting intracellular localization of lipid probes using QSAR models. Biotech. Histochem. 2015, 90, 255–263. [Google Scholar] [CrossRef]
- Horobin, R.W.; Rashid, F. Interactions of molecular probes with living cells and tissues. Part 1. Some general mechanistic proposals, making use of a simplistic Chinese box model. Histochemistry 1990, 94, 205–209. [Google Scholar] [CrossRef]
- Horobin, R.W.; Rashid-Doubell, F. Predicting small molecule fluorescent probe localization in living cells using QSAR modeling. 2. Specifying probe, protocol and cell factors; selecting QSAR models; predicting entry and localization. Biotech. Histochem. 2013, 88, 461–476. [Google Scholar] [CrossRef]
- Horobin, R.W.; Rashid-Doubell, F.; Pediani, J.D.; Milligan, G. Predicting small molecule fluorescent probe localization in living cells using QSAR modeling. 1. Overview and models for probes of structure, properties and function in single cells. Biotech. Histochem. 2013, 88, 440–460. [Google Scholar] [CrossRef]
- Cruz, V.L.; Martinez, S.; Ramos, J.; Martinez-Salazar, J. 3D-QSAR as a Tool for Understanding and Improving Single-Site Polymerization Catalysts. A Review. Organometallics 2014, 33, 2944–2959. [Google Scholar] [CrossRef]
- Ding, B.; Yu, Y.; Geng, S.; Liu, B.; Hao, Y.; Liang, G. Computational Methods for the Interaction between Cyclodextrins and Natural Compounds: Technology, Benefits, Limitations, and Trends. J. Agric. Food Chem. 2022, 70, 2466–2482. [Google Scholar] [CrossRef]
- Muratov, E.N.; Bajorath, J.; Sheridan, R.P.; Tetko, I.V.; Filimonov, D.; Poroikov, V.; Oprea, T.I.; Baskin, I.I.; Varnek, A.; Roitberg, A.; et al. QSAR without borders. Chem. Soc. Rev. 2020, 49, 3525–3564. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Learnings from quantitative structure–activity relationship (QSAR) studies with respect to food protein-derived bioactive peptides: A review. RSC Adv. 2016, 6, 75400–75413. [Google Scholar] [CrossRef]
- Radaeva, M.; Dong, X.; Cherkasov, A. The Use of Methods of Computer-Aided Drug Discovery in the Development of Topoisomerase II Inhibitors: Applications and Future Directions. J. Chem. Inf. Model. 2020, 60, 3703–3721. [Google Scholar] [CrossRef]
- Liu, R.; Jiang, W.; Walkey, C.D.; Chan, W.C.W.; Cohen, Y. Prediction of nanoparticles-cell association based on corona proteins and physicochemical properties. Nanoscale 2015, 7, 9664–9675. [Google Scholar] [CrossRef]
- Ambure, P.; Roy, K. Understanding the structural requirements of cyclic sulfone hydroxyethylamines as hBACE1 inhibitors against Aβ plaques in Alzheimer’s disease: A predictive QSAR approach. RSC Adv. 2016, 6, 28171–28186. [Google Scholar] [CrossRef]
- Rashid, F.; Horobin, R.W. Interaction of molecular probes with living cells and tissues. Part 2. Histochemistry 1990, 94, 303–308. [Google Scholar] [CrossRef]
- Rashid, F.; Horobin, R.W.; Williams, M.A. Predicting the behaviour and selectivity of fluorescent probes for lysosomes and related structures by means of structure-activity models. Histochemistry 1991, 23, 450–459. [Google Scholar] [CrossRef]
- Colston, J.; Horobin, R.; Rashid-Doubell, F.; Pediani, J.; Johal, K. Why fluorescent probes for endoplasmic reticulum are selective: An experimental and QSAR-modelling study. Biotech. Histochem. 2003, 78, 323–332. [Google Scholar] [CrossRef]
- Horobin, R.W.; Stockert, J.C.; Rashid-Doubell, F. Fluorescent cationic probes for nuclei of living cells: Why are they selective? A quantitative structure–activity relations analysis. Histochem. Cell Biol. 2006, 126, 165–175. [Google Scholar] [CrossRef]
- Horobin, R.W.; Stockert, J.C.; Rashid-Doubell, F. Uptake and localisation of small-molecule fluorescent probes in living cells: A critical appraisal of QSAR models and a case study concerning probes for DNA and RNA. Histochem. Cell Biol. 2013, 139, 623–637. [Google Scholar] [CrossRef]
- Horobin, R.W.; Stockert, J.C. Uptake and localization mechanisms of fluorescent and colored lipid probes. 1. Physicochemistry of probe uptake and localization, and the use of QSAR models for selectivity prediction. Biotech. Histochem. 2011, 86, 379–393. [Google Scholar] [CrossRef]
- Uchinomiya, S.; Horobin, R.W.; Alvarado-Martinez, E.; Pena-Cabrera, E.; Chang, Y.T. Prediction of Intracellular Localization of Fluorescent Dyes Using QSAR Models. Comb. Chem. High Throughput Screen. 2016, 19, 378–383. [Google Scholar] [CrossRef]
- Singh, D.; Rajput, D.; Kanvah, S. Fluorescent probes for targeting endoplasmic reticulum: Design strategies and their applications. Chem. Commun. 2022, 58, 2413–2429. [Google Scholar] [CrossRef]
- Liu, C.Y.; Zhu, H.C.; Zhang, Y.; Su, M.J.; Liu, M.Y.; Zhang, X.H.; Wang, X.; Rong, X.D.; Wang, K.; Li, X.W.; et al. Recent advances in Golgi-targeted small-molecule fluorescent probes. Coord. Chem. Rev. 2022, 462, 214504. [Google Scholar] [CrossRef]
- Rashid, F.; Horobin, R.W. Accumulation of Fluorescent Non-Cationic Probes in Mitochondria of Cultured-Cells—Observations, a Proposed Mechanism, and Some Implications. J. Microsc. 1991, 163, 233–241. [Google Scholar] [CrossRef]
- Lansiaux, A.; Tanious, F.; Mishal, Z.; Dassonneville, L.; Kumar, A.; Stephens, C.E.; Hu, Q.; Wilson, W.D.; Boykin, D.W.; Bailly, C. Distribution of furamidine analogues in tumor cells: Targeting of the nucleus or mitochondria depending on the amidine substitution. Cancer Res. 2002, 62, 7219–7229. [Google Scholar]
- Collot, M.; Ashokkumar, P.; Anton, H.; Boutant, E.; Faklaris, O.; Galli, T.; Mely, Y.; Danglot, L.; Klymchenko, A.S. MemBright: A Family of Fluorescent Membrane Probes for Advanced Cellular Imaging and Neuroscience. Cell Chem. Biol. 2019, 26, 600. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Svetnik, V.; Liaw, A.; Tong, C.; Culberson, J.C.; Sheridan, R.P.; Feuston, B.P. Random forest: A classification and regression tool for compound classification and QSAR modeling. J. Chem. Inf. Comput. Sci. 2003, 43, 1947–1958. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Kier, L.B.; Hall, L.H. A Differential Molecular Connectivity Index. Quant. Struct. Act. Relatsh. 1991, 10, 134–140. [Google Scholar] [CrossRef]
- Wildman, S.A.; Crippen, G.M. Prediction of Physicochemical Parameters by Atomic Contributions. J. Chem. Inf. Comput. Sci. 1999, 39, 868–873. [Google Scholar] [CrossRef]
- Schneider, G.; Neidhart, W.; Giller, T.; Schmid, G. “Scaffold-Hopping” by Topological Pharmacophore Search: A Contribution to Virtual Screening. Angew. Chem. Int. Ed. 1999, 38, 2894–2896. [Google Scholar] [CrossRef]
- Phaniraj, S.; Gao, Z.; Rane, D.; Peterson, B.R. Hydrophobic resorufamine derivatives: Potent and selective red fluorescent probes of the endoplasmic reticulum of mammalian cells. Dyes Pigm. 2016, 135, 127–133. [Google Scholar] [CrossRef]
- Yuriev, E.; Kong, D.C.M.; Iskander, M.N. Investigation of structure-activity relationships in a series of glibenclamide analogues. Eur. J. Med. Chem. 2004, 39, 835–847. [Google Scholar] [CrossRef]
- Lu, S.; Dai, Z.; Cui, Y.; Kong, D.-M. Recent Development of Advanced Fluorescent Molecular Probes for Organelle-Targeted Cell Imaging. Biosensors 2023, 13, 360. [Google Scholar] [CrossRef]
- Li, S.; Yang, K.; Zeng, J.; Ding, Y.; Cheng, D.; He, L. Golgi-Targeting Fluorescent Probe for Monitoring CO-Releasing Molecule-3 In Vitro and In Vivo. ACS Omega 2022, 7, 9929–9935. [Google Scholar] [CrossRef]
- Scorrano, L.; Petronilli, V.; Colonna, R.; Di Lisa, F.; Bernardi, P. Chloromethyltetramethylrosamine (Mitotracker OrangeTM) Induces the Mitochondrial Permeability Transition and Inhibits Respiratory Complex I: IMPLICATIONS FOR THE MECHANISM OF CYTOCHROME c RELEASE*. J. Biol. Chem. 1999, 274, 24657–24663. [Google Scholar] [CrossRef]
- Poot, M.; Zhang, Y.Z.; Krämer, J.A.; Wells, K.S.; Jones, L.J.; Hanzel, D.K.; Lugade, A.G.; Singer, V.L.; Haugland, R.P. Analysis of mitochondrial morphology and function with novel fixable fluorescent stains. J. Histochem. Cytochem. 1996, 44, 1363–1372. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # 1 | Descriptor | Information |
|---|---|---|
| 10 | LogD at pH 7.4 | Distribution coefficient at pH 7.4 |
| 10 | Chemaxon HLB | Hydrophilic-lipophilic balance of a molecule, consensus value from Griffin and Davies methods calculated by the Chemaxon program |
| 10 | Topological PSA | Topological polar surface area of a molecule |
| 10 | Strongest basic pKa1 | The first dissociation constant of a molecule |
| 9 | 2D VSA H-bond | Distribution of hydrogen bonding sites within a molecule |
| 9 | Fraction of 2D VSA H-bond | Fraction of the surface area covered by hydrogen bonds to the total surface area of the molecule |
| 9 | SlogP 3 | Octanol-water partition coefficient |
| 9 | SlogP Me1 3 | Contribution of p-block elements to the partition coefficient, excluding ones described by other SlogP atom types |
| 9 | Fraction of N atoms | Fraction or proportion of nitrogen atoms within a molecule |
| 8 | 2D VSA H-bond acceptor | Distribution of hydrogen bond acceptor sites within a molecule |
| 8 | CATS Hyd Pos 02 4 | Number of atom pairs within a molecule: hydrophobic-positive charge, separated by distance of two bonds |
| 8 | CATS Don Acc 02 4 | Number of atom pairs within a molecule: hydrogen bond donor-acceptor, separated by distance of two bonds |
| 8 | CATS Don Pos 00 4 | Number of atoms within a molecule that exhibit both a hydrogen donor and a positive charge |
| 7 | 2D VSA polar | Contribution of polar surface area within a molecule |
| 7 | SC 04 path cluster 2 | Kier and Hall’s topological descriptor: subgraph count of order 4, path cluster |
| 7 | Formal charge | Formal charge of a molecule |
| 7 | CATS Aro Pos 02 4 | Number of atom pairs within a molecule: aromatic-positive charge, separated by distance of two bonds |
| 7 | CATS Pos Pos 00 4 | Number of positively charged atoms within a molecule |
| 7 | CATS binary Don Pos 00 4 | Presence of atoms within a molecule that exhibit both a hydrogen donor and a positive charge |
| 6 | Number of positively chargeable groups | Number of positively chargeable groups within a molecule |
| 6 | Number of positively charged groups | Number of positive charged groups within a molecule |
| 6 | Fraction of 2D VSA H-bond acceptor | The fraction of the 2D surface area that corresponds to hydrogen bond acceptor sites within a molecule |
| 6 | SlogP H1 3 | Number of hydrocarbon hydrogens |
| 6 | Fraction of H atoms | Fraction of hydrogen atoms within a molecule |
| 6 | Fraction of rotatable bonds | Fraction of rotatable bonds within a molecule |
| 5 | Fraction of 2D VSA polar | Fraction of 2D Van der Waals polar surface area |
| 5 | Number of fluorine atoms | The number of fluorine atoms within a molecule |
| 5 | Number of C2 | The number of secondary carbon atoms within a molecule |
| 4 | 2D VSA hydrophobic | The contribution of hydrophobic surface area within a molecule |
| 4 | Fraction of double bonds | Fraction of double bonds within a molecule |
| 4 | Fraction of aromatic bonds | Fraction of aromatic bonds within a molecule |
| 4 | CATS binary Don Acc 02 4 | Presence of atom pairs within a molecule: hydrogen bond donor-acceptor, separated by distance of two bonds |
| 4 | Number of CsH | Number of single bonds between carbon and hydrogen atoms within a molecule |
| 3 | Number of inorganic atoms | Number of inorganic atoms within a molecule |
| 3 | SC 04 path 2 | Kier and Hall’s topological descriptor: subgraph count of order 4, path |
| 3 | Number of boron atoms | Number of boron atoms within a molecule |
| 3 | Fraction of oxygen atoms | Fraction of oxygen atoms within a molecule |
| 3 | Fraction of rigid bonds | Fraction of chemical bonds that exhibit limited or negligible flexibility or rotational freedom |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-H.; Lee, H.-G.; Liu, X.; Lee, S.K.; Chang, Y.-T. Quantitative Structure-Activity Relationship of Fluorescent Probes and Their Intracellular Localizations. Chemosensors 2023, 11, 310. https://doi.org/10.3390/chemosensors11050310
Park S-H, Lee H-G, Liu X, Lee SK, Chang Y-T. Quantitative Structure-Activity Relationship of Fluorescent Probes and Their Intracellular Localizations. Chemosensors. 2023; 11(5):310. https://doi.org/10.3390/chemosensors11050310
Chicago/Turabian StylePark, Seong-Hyeon, Hong-Guen Lee, Xiao Liu, Sung Kwang Lee, and Young-Tae Chang. 2023. "Quantitative Structure-Activity Relationship of Fluorescent Probes and Their Intracellular Localizations" Chemosensors 11, no. 5: 310. https://doi.org/10.3390/chemosensors11050310
APA StylePark, S.-H., Lee, H.-G., Liu, X., Lee, S. K., & Chang, Y.-T. (2023). Quantitative Structure-Activity Relationship of Fluorescent Probes and Their Intracellular Localizations. Chemosensors, 11(5), 310. https://doi.org/10.3390/chemosensors11050310