Lyme Disease Transmission Risk: Seasonal Variation in the Built Environment


Abstract
1. Introduction
2. Materials and Methods
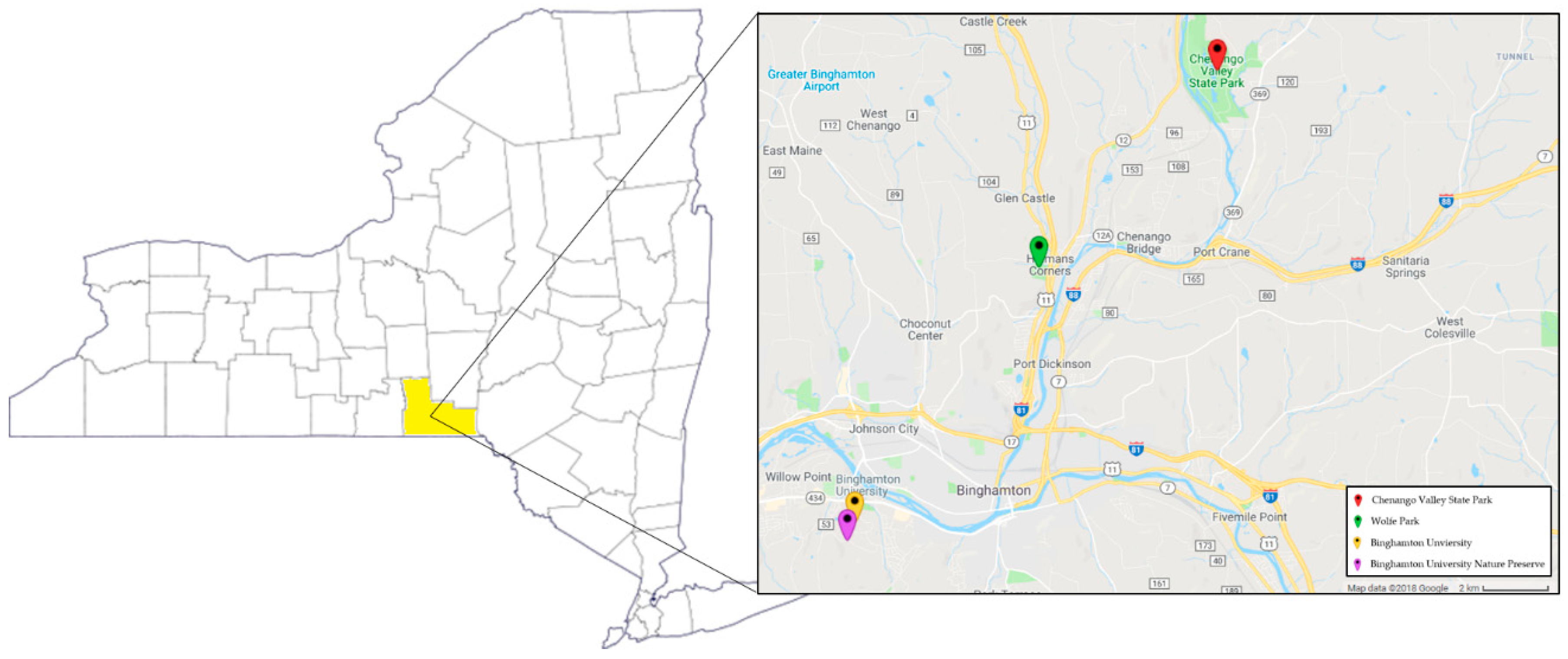
2.1. Tick Collection
2.2. Prevalence of B. burgdorferi in Ticks
2.3. Statistical Analyses
3. Results
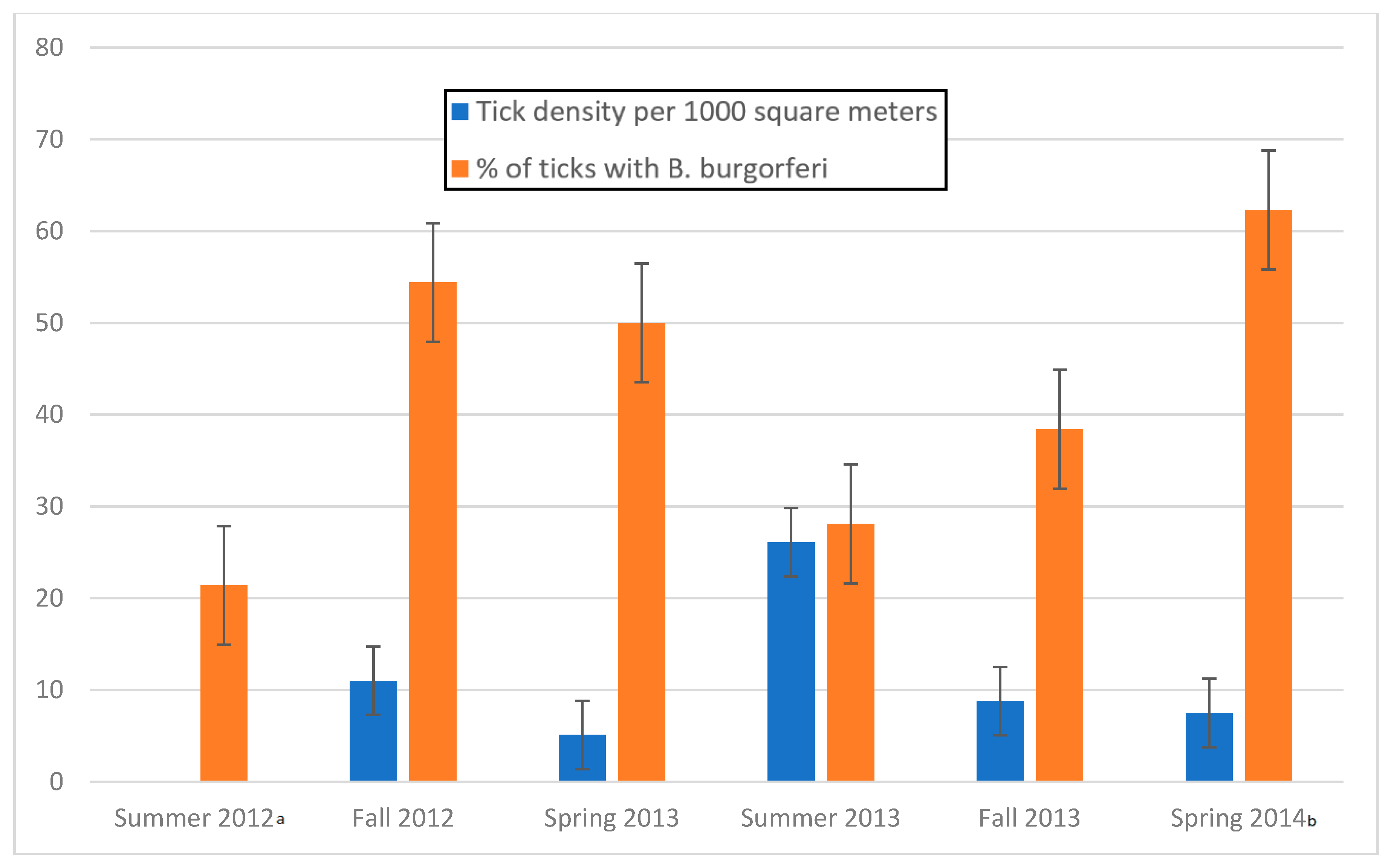
3.1. Tick Density
3.2. Tick Infection Rate
3.3. Seasonality
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Binder, S.; Levitt, A.M.; Sacks, J.J.; Hughes, J.M. Emerging infectious diseases: Public health issues for the 21st century. Science 1999, 284, 1311–1313. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- King, D.A.; Peckham, C.; Waage, J.K.; Brownlie, J.; Woolhouse, M.E.J. Infectious diseases: Preparing for the future. Science 2006, 313, 1392–1393. [Google Scholar] [CrossRef] [PubMed]
- Morens, D.M.; Folkers, G.K.; Fauci, A.S. The challenge of emerging and re-emerging infectious diseases. Nature 2004, 430, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Smolinski, M.S.; Hamburg, M.A.; Lederberg, J. Microbial Threats to Health: Emergence, Detection and Response; The National Academies Press: Washington, DC, USA, 2003; p. 367. ISBN 0-309-50730-8. [Google Scholar]
- Daszak, P.; Cuningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A.; Daszak, P.; Tabor, G.M.; Aquirre, A.A.; Pearl, M.; Epstein, J.; Wolfe, N.D.; Kilpatrick, A.M.; Foufopoulos, J.; Molyneux, D.; et al. Unhealthy landscapes: Policy recommendations on land use change and infectious disease emergence. Environ. Health Perspect. 2004, 112, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.H.; Latham, S.M.; Woolhouse, M.E.J. Risk factors for human disease emergence. Philos. Trans. R. Soc. Lond. 2001, 356, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg. Infect. Dis. 2005, 11, 1842–1847. [Google Scholar] [CrossRef] [PubMed]
- Lederberg, J.; Shope, R.E.; Oakes, S.C.J. Emerging Infections: Microbial Threats to Health in the United States; Institute of Medicine, The National Academies Press: Washington, DC, USA, 1992; p. 294. ISBN 0-309-04741-2. [Google Scholar]
- Weiss, R.A.; McMichael, A.J. Social and environmental risk factors in the emergence of infectious diseases. Nat. Med. 2004, 10, S70–S76. [Google Scholar] [CrossRef] [PubMed]
- Altizer, S.; Ostfeld, R.S.; Johnson, T.J.; Kutz, S.; Harvell, C.D. Climate Change and Infectious Diseases: From Evidence to a Predictive Framework. Science 2013, 341, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. Resurgent vector-borne diseases as a global health pattern. Emerg. Infect. Dis. 1998, 4, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Randolph, S.E. Drivers, dynamics, and control of emerging vector-borne zoonotic diseases. Lancet 2012, 380, 1946–4955. [Google Scholar] [CrossRef]
- McMichael, A.J.; Woodruff, R.E.; Hales, S. Climate change and human health: Present and future risks. Lancet 2006, 367, 859–869. [Google Scholar] [CrossRef]
- Burgdorfer, W.A.; Barbour, A.G.; Hayes, S.F.; Benach, J.L.; Grunwaldt, E.; Davis, J.P. Lyme disease—A tick-borne spirochetosis? Science 1982, 216, 1317–1319. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Piesman, J.; Zielinski-Gutierrez, E.; Eisen, L. What do we need to know about disease ecology to prevent Lyme disease in the Northeastern United States? J. Med. Entomol. 2012, 49, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H., Jr. Updates on Borrelia burgdorferi sensu lato complex with respect to public health. Ticks Tick Borne Dis. 2011, 2, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Burgdorfer, W.; Lane, R.S.; Barbour, A.G.; Gresbrink, R.A.; Anderson, J.R. The Western Black-Legged Tick, Ixodes pacificus: A Vector of Borrelia Burgdorferi. Am. J. Trop. Med. Hyg. 1985, 34, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Shadick, N.A.; Phillips, C.B.; Shangha, O.; Logigian, E.L.; Kaplan, R.F.; Wright, E.A. Musculoskeletal and Neurological Outcomes in Patients with Previously Treated Lyme Disease. Ann. Intern. Med. 1999, 131, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Shadick, N.A.; Phillips, C.B.; Logigian, E.L.; Steere, A.C.; Kaplan, R.F.; Berardi, V.P.; Duray, P.H.; Larson, M.G.; Wright, E.A.; Katz, J.N.; et al. The long-term clinical outcomes of Lyme disease. A population-based retrospective cohort study. Ann. Intern. Med. 1994, 121, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Citrea, M.; Freeman, P.R.; Horowitz, R.I. Empirical validation of the Horowitz Multiple Systemic Infectious Disease Syndrome Questionnaire for suspected Lyme disease. Int. J. Intern. Med. 2017, 10, 249–273. [Google Scholar] [CrossRef] [PubMed]
- Rebman, A.W.; Bechtold, K.T.; Yang, T.; Mihm, E.A.; Soloski, M.J.; Novak, C.B.; Aucott, J.N. The Clinical, Symptom, and Quality-of-Life Characterization of a Well-Defined Group of Patients with Posttreatment Lyme Disease Syndrome. Front. Med. 2017, 4, 224. [Google Scholar] [CrossRef] [PubMed]
- Aucott, J.N.; Crowder, L.A.; Yedlin, V.; Kortte, K.B. Bull’s-Eye and Nontarget Skin Lesions of Lyme Disease: An Internat Survey of Identification of Erythema Migrans. Dermatol. Res. Pract. 2012, 2012, 451727. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Bartenhagen, N.H.; Craft, J.E.; Hutchinson, G.J.; Newman, J.H.; Rahn, D.W.; Sigal, L.H.; Spieler, P.N.; Stenn, K.S.; Malawista, S.E. The early clinical manifestations of Lyme disease. Ann. Intern. Med. 1983, 99, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Sikand, W.V. The presenting manifestations of Lyme disease. Ann. Intern. Med. 2003, 348, 2472–2474. [Google Scholar] [CrossRef]
- Kostic, T.; Momcilovic, S.; Perisic, Z.D.; Apostolovic, S.R.; Cvetkovic, J.; Jovanovic, A.; Barac, A.; Salinger-Martinovic, S.; Tasic-Otasevic, S. Manifestations of Lyme carditis. Int. J. Cardiol. 2017, 232, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, D.M. CDC estimates 300,000 cases of Lyme disease annually. J. Am. Med. Assoc. 2013, 310, 1110. [Google Scholar] [CrossRef] [PubMed]
- Hinckley, A.F.; Connally, N.P.; Meek, J.I.; Johnson, B.J.; Kemperman, M.M.; Feldman, K.A.; White, J.L.; Mead, P.S. Lyme disease testing by large commercial laboratories in the United States. Clin. Infect. Dis. 2014, 59, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.A.; Saha, S.; Kugeler, K.J.; Delorey, M.J.; Shankar, M.B.; Hinckley, A.F.; Mead, P.S. Incidence of Clinician-Diagnosed Lyme Disease, United States, 2005–2010. Emerg. Infect. Dis. 2015, 21, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, R.; Lindsey, N.P.; Fischer, M.; Gregory, C.J.; Hinckley, A.F.; Mead, P.S.; Paz-Bailey, G.; Waterman, S.H.; Drexler, N.A.; Kersh, G.J.; et al. Vital Signs: Trends in Reported Vectorborne Disease Cases- United States and Territories, 2004–2016. Morb. Mortal. Wkly. Rep. 2018, 67, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Eisen, L. The Blacklegged Tick, Ixodes scapularis: An Increasing Public Health Concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef] [PubMed]
- McMichael, A.J. The urban environment and health in a world of increasing globalization: Issues for developing countries. Bull. World Health Organ. 2000, 78, 1117–1126. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; O’Fallon, L.R.; Dearry, A. Creating healthy communities, healthy homes, healthy people: Initiating a research agenda on the built environment and public health. Am. J. Public Health 2003, 93, 1446–1450. [Google Scholar] [CrossRef] [PubMed]
- Cruz, T.; Keppler, H.; Thomas, J.; Kommareddy, D.; Hempstead, S.; Valentine, E.; Spathis, R.; Darcy, J.M., II; Garruto, R.M. Assessing prevalence of tick-borne infectious agents on a university campus. Am. J. Hum. Biol. 2013, 25, 254–255. [Google Scholar]
- Darcy, J.M., II; Spathis, R.; Schmidt, J.; Keppler, H.; Hempstead, S.; Cruz, T.; Kommareddy, D.; Thomas, J.; Riddle, M.; Sayama, H.; et al. Emergence, transmission and risk of Lyme disease and other tick-borne infections: A community based natural experimental model. Am. J. Hum. Biol. 2013, 25, 255. [Google Scholar]
- Allan, B.F.; Keesing, F.; Ostfeld, R.S. Effect of Forest Fragmentation on Lyme Disease Risk. Conserv. Biol. 2003, 17, 267–272. [Google Scholar] [CrossRef]
- Brownstein, J.S.; Skelly, D.K.; Holford, T.R.; Fish, D. Forest fragmentation predicts local scale heterogeneity of Lyme disease risk. Oecologia 2005, 146, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.F.; Magnarelli, L.A. Biology of ticks. Infect. Dis. Clin. N. Am. 2008, 22, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Hengge, U.R.; Tannapfel, A.; Tyring, S.K.; Erbel, R.; Arendt, G.; Ruzicka, T. Lyme borreliosis. Lancet Infect. Dis. 2003, 3, 489–500. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Keesing, F. Biodiversity and disease risk: The case of Lyme disease. Conserv. Biol. 2000, 14, 722–728. [Google Scholar] [CrossRef]
- Rollend, L.; Fish, D.; Childs, J.E. Transovarial transmission of Borrelia spirochetes by Ixodes scapularis: A summary of the literature and recent observations. Ticks Tick-Borne Dis. 2013, 4, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Crowder, C.D.; Carolan, H.E.; Rounds, M.A.; Honig, V.; Mothes, B.; Haag, H.; Nolte, O.; Luft, B.J.; Grubhoffer, L.; Ecker, D.J.; et al. Prevalence of Borrelia miyamotoi in Ixodes Ticks in Europe and the United States. Emerg. Infect. Dis. 2014, 20, 1678–1682. [Google Scholar] [CrossRef] [PubMed]
- Salkeld, D.J.; Cinkovich, S.; Nieto, N.C. Tick-borne Pathogens in Northwestern California, USA. Emerg. Infect. Dis. 2014, 20, 493–494. [Google Scholar] [CrossRef] [PubMed]
- Kurtenbach, K.; Hanincova, K.; Tsao, J.I.; Margos, G.; Fish, D.; Ogden, N.H. Fundamental processes in the evolutionary ecology of Lyme borreliosis. Nat. Rev. Microbiol. 2006, 4, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.; Salinas, A.; Martinez, F.; Cantu, A.; Miller, D.K. Serosurvey for selected disease agents in white-tailed deer from Mexico. J. Wildl. Dis. 1999, 35, 799–803. [Google Scholar] [CrossRef] [PubMed]
- A History of Lyme Disease, Symptoms, Diagnosis, Treatment and Prevention. Available online: http://www.niaid.nih.gov/topics/lymeDisease/understanding/Pages/intro.aspx (accessed on 8 August 2017).
- Barbour, A.G.; Fish, D. The biological and social phenomenon of Lyme disease. Science 1993, 260, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- LoGuidice, K.; Ostfeld, R.S.; Schmidt, K.A.; Keesing, F. The ecology of infectious disease: Effects of host diversity and community composition on Lyme disease risk. Proc. Natl. Acad. Sci. USA 2003, 100, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Be Tick Free—A Guide for Preventing Lyme Disease. Available online: http://www.health.ny.gov/publications/2825/ (accessed on 8 August 2017).
- Diuk-Wasser, M.A.; Hoen, A.G.; CIslo, P.; Brinkerhoff, R.; Hamer, S.A.; Rowland, M.; Cortinas, R.; Vourc’h, G.; Melton, F.; Hickling, G.J.; et al. Human risk of infection with Borrelia burgdorferi, the Lyme disease agent, in Eastern United States. Am. J. Trop. Med. Hyg. 2012, 86, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Hamer, S.A.; Tsao, J.I.; Walker, E.D.; Hickling, G.J. Invasion of the Lyme disease vector Ixodes scapularis: Implications for Borrelia burgdorferi endemicity. EcoHealth 2010, 7, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Roome, A.; Bower, K.; Murnock, C.G.; Hill, L.; Ho, B.; Tyurin, S.; Al-Feghali, V.; Zeitz, H.; Rios, D.; Parwez, R.; et al. Prevalence of tick-borne pathogens and human behavioral risk factors in built environments of upstate New York suggest a necessity for the development of risk management models. Am. J. Hum. Biol. 2014, 26, 279. [Google Scholar]
- Daniels, T.J.; Ralco, R.C.; Fish, D. Estimating population size and drag sampling efficiency for the blacklegged tick (Acari: Ixodidae). J. Med. Entomol. 2000, 37, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Falco, R.C.; Fish, D. A comparison of methods for sampling the deer tick, Ixodes dammini, in a Lyme disease endemic area. Exp. Appl. Acarol. 1992, 14, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Hersh, M.H.; Ostfeld, R.S.; McHenry, D.J.; Tibbett, M.; Brunner, J.L.; Killilea, M.E.; LoGiudice, K.; Schmidt, K.A.; Keesing, F. Co-infection of blacklegged ticks with Babesia microti and Borrelia burgdorferi is higher than expected and acquired from small mammal hosts. PLoS ONE 2014, 9, e99348. [Google Scholar] [CrossRef] [PubMed]
- Marconi, R.T.; Garon, C.F. Development of polymerase chain reaction primer sets for diagnosis of Lyme disease and for species-specific identification of Lyme disease isolates by 16S rRNA signature nucleotide analysis. J. Clin. Microbiol. 1992, 30, 2830–2834. [Google Scholar] [PubMed]
- Lee, S.H.; Vigliotti, V.S.; Vigliotti, J.S.; Jones, W.; Pappu, S. Increased Sensitivity of Borrelia burgdorferi 16S Ribosomal DNA Detection. Am. J. Clin. Pathol. 2010, 133, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Vuong, H.B.; Canham, C.D.; Fonesca, D.M.; Brisson, D.; Morin, P.J.; Smouse, P.E.; Ostfeld, R.S. Occurrence and transmission efficiencies of Borrelia burgdorferi OspC types in avian and mammalian wildlife. Infect. Genet. Evolut. 2014, 27, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.H.; Fish, D.; Moy, F.H. Landscape features associated with Lyme disease risk in a suburban residential environment. Landsc. Ecol. 1998, 13, 27–36. [Google Scholar] [CrossRef]
- Maupin, G.O.; Fish, D.; Zultowsky, J.; Campos, E.G.; Piesman, J. Landscape ecology of Lyme disease in a residential area of Westchester County, New York. Am. J. Epidemiol. 1991, 133, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Communicable Disease in New York State—Rate Per 100,000 Population: Lyme Disease to Shigellosis: 2000. Available online: https://www.health.ny.gov/statistics/diseases/communicable/2000/rates3.htm (accessed on 8 August 2017).
- Newman, E.A.; Eisen, L.; Eisen, R.J.; Fedorova, N.; Hasty, J.M.; Vaughn, C.; Lane, R.S. Borrelia burgdorferi sensu lato spirochetes in wild birds in northwestern California: Associations with ecological factors, bird behavior and tick infestation. PLoS ONE 2015, 10, e0118146. [Google Scholar] [CrossRef] [PubMed]
- Prusinski, M.A.; Kokas, J.E.; Hukey, K.T.; Kogut, S.J.; Lee, J.; Backenson, P.B. Prevalence of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae), Anaplasma phagocytophilum (Rickettsiales: Anaplasmataceae), and Babesia microti (Piroplasmida: Babesiidae) in Ixodes scapularis (Acari: Ixodidae) collected from recreational lands in the Hudson Valley Region, New York State. J. Med. Entomol. 2014, 51, 226–236. [Google Scholar] [PubMed]
- Lyme Disease Incidence per 100,000. Available online: https://www.health.ny.gov/statistics/chac/general/g40.htm (accessed on 8 August 2017).
- Ebel, G.D.; Kramer, L.D. Short report: Duration of tick attachment required for transmission of Powassan virus by deer ticks. Am. J. Trop. Med. Hyg. 2004, 71, 268–271. [Google Scholar] [PubMed]
- Hynote, E.D.; Mervine, P.C.; Stricker, R.B. Clinical evidence for rapid transmission of lyme disease following a tickbite. Diagn. Microbiol. Infect. Dis. 2012, 72, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Piesman, J.; Schneider, B.S.; Zeidner, N.S. Use of quantitative PCR to measure density of Borrelia burgdorferi in the midgut and salivary glands of feeding tick vectors. J. Clin. Microbiol. 2001, 39, 4145–4148. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.J. Lyme borreliosis: A review of data on transmission time after tick attachment. Int. J. Gen. Med. 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jouda, F.; Perret, J.L.; Gern, L. Density of questing Ixodes ricinus nymphs and adults infected by Borrelia burgdorferi sensu lato in Switzerland: Spatio-temporal pattern at a regional scale. Vector Borne Zoonotic Dis. 2004, 4, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Tälleklint, L.; Jaenson, T.G. Seasonal variations in density of questing Ixodes ricinus (Acari: Ixodidae) nymphs and prevalence of infection with B. burgdorferi s.l. in south central Sweden. J. Med. Entomol. 1996, 33, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, R.; Jain, K.; Bennett, A.; Briese, T.; Lipkin, W.I. Assessment of polymicrobial infections in ticks in New York state. Vector Borne Zoonotic Dis. 2010, 10, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Couch, P.; Johnson, C.E. Prevention of Lyme disease. Am. J. Health-Syst. Pharmacy. 1992, 49, 1164–1173. [Google Scholar]
- Vazquez, M.; Muehlenbein, C.; Cartter, M.; Hayes, E.B.; Ertel, S.; Shapiro, E.D. Effectiveness of Personal Protective Measures to Prevent Lyme Disease. Emerg. Infect. Dis. 2008, 14, 210–216. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Season | Month | Total Ticks Collected | Total Area Dragged | Tick Density per 1000 m2 | # Ticks Tested | % Ticks Infected | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Nymphs | Adults | Square Meters | Nymphs (95% CI) | Adults (95% CI) | Nymphs | Adults | Nymphs (95% CI) | Adults (95% CI) | ||
| Summer 2012 | June | Data not collected | 15 | 3 | 46.7 | 66.7 | ||||
| July | 54 | 12 | 14.8 | 8.3 | ||||||
| Overall | 69 | 15 | 21.7 | 20.0 | ||||||
| Fall 2012 | October | 4 | 68 | 5304.4 | 0.8 | 12.8 | 4 | 68 | 50.0 | 47.1 |
| November | 0 | 20 | 3051.6 | 0.0 | 6.6 | 0 | 18 | n/a | 83.3 | |
| Overall | 4 | 88 | 8356.0 | 0.5 | 10.5 | 4 | 86 | 50.0 | 54.7 | |
| Spring 2013 | April | 0 | 32 | 3172.1 | 0.0 | 10.1 | 0 | 29 | n/a | 37.9 |
| May | 0 | 9 | 4829.0 | 0.0 | 1.9 | 0 | 9 | n/a | 33.3 | |
| Overall | 0 | 41 | 8001.1 | 0.0 | 5.1 | 0 | 38 | n/a | 36.8 | |
| Summer 2013 | June | 467 | 1 | 17,958.9 | 26.0 | 0.1 | 419 | 1 | 28.2 | 0.0 |
| Overall | 467 | 1 | 17,958.9 | 26.0 | 0.1 | 419 | 1 | 28.2 | 0.0 | |
| Fall 2013 | September | 0 | 13 | 11,286.3 | 0.0 | 1.2 | 0 | 13 | n/a | 30.8 |
| October | 10 | 525 | 53,048.4 | 0.2 | 9.9 | 10 | 521 | 30.0 | 38.2 | |
| November | 0 | 24 | 927.0 | 0.0 | 25.9 | 0 | 24 | n/a | 50.0 | |
| Overall | 10 | 562 | 65,261.7 | 0.2 | 8.6 | 10 | 558 | 30.0 | 38.5 | |
| Spring 2014 | April | 0 | 131 | 20,627.9 | 0.0 | 6.4 | 0 | 130 | n/a | 62.3 a |
| May | 0 | 71 | 6407.1 | 0.0 | 11.1 | 0 | 65 | n/a | 72.3 a | |
| Overall | 0 | 202 | 27,035.0 | 0.0 | 7.5 | 0 | 195 | n/a | 65.6 a | |
| Summer 2012–Spring 2014 | 481 | 894 | 126,612.6 | 3.8 (±3.1) | 7.1 (±3.1) | 502 | 893 | 27.5 (±4.38) | 45.5 (±2.83) | |
| Overall | 1375 * | 126,612.6 | 10.9 (±2.19) | 1395 * | 39.0 (±2.38) | |||||
| Season | Month | Density of Infected Ticks | |
|---|---|---|---|
| Nymphs | Adults | ||
| Fall 2012 | October | 0.4 | 6.6 |
| November | 0.0 | 4.9 | |
| Ovearll | 0.2 | 5.5 | |
| Spring 2013 | April | 0.0 | 3.5 |
| May | 0.0 | 0.6 | |
| Overall | 0.0 | 1.7 | |
| Summer 2013 | June | 6.6 | 0.0 |
| July | Data not collected for July and August. Cannot be calculated | ||
| August | |||
| Overall | 6.6 | 0.0 | |
| Fall 2013 | September | 0.0 | 0.4 |
| October | 0.1 | 3.8 | |
| November | 0.0 | 12.9 | |
| Overall | 0.1 | 3.3 | |
| Spring 2014 | April | 0.0 | 3.9 |
| May | 0.0 | 7.3 | |
| Overall | 0.0 | 4.7 | |
| Fall 2012–Spring 2014 | 1.1 (±3.1) | 3.2 (±3.1) | |
| Total Ticks Fall 2012–Spring 2014 | 4.3 (±3.1) | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roome, A.; Spathis, R.; Hill, L.; Darcy, J.M.; Garruto, R.M. Lyme Disease Transmission Risk: Seasonal Variation in the Built Environment. Healthcare 2018, 6, 84. https://doi.org/10.3390/healthcare6030084
Roome A, Spathis R, Hill L, Darcy JM, Garruto RM. Lyme Disease Transmission Risk: Seasonal Variation in the Built Environment. Healthcare. 2018; 6(3):84. https://doi.org/10.3390/healthcare6030084
Chicago/Turabian StyleRoome, Amanda, Rita Spathis, Leah Hill, John M. Darcy, and Ralph M. Garruto. 2018. "Lyme Disease Transmission Risk: Seasonal Variation in the Built Environment" Healthcare 6, no. 3: 84. https://doi.org/10.3390/healthcare6030084
APA StyleRoome, A., Spathis, R., Hill, L., Darcy, J. M., & Garruto, R. M. (2018). Lyme Disease Transmission Risk: Seasonal Variation in the Built Environment. Healthcare, 6(3), 84. https://doi.org/10.3390/healthcare6030084