
Synergistic Impact of Active Case Detection and Early Hospitalization for Controlling the Spread of Yellow Fever Outbreak in Nigeria: An Epidemiological Modeling and Optimal Control Analysis

 , , and
, , and 
Abstract
1. Introduction
2. Methods
2.1. Yellow Fever Epidemic Model
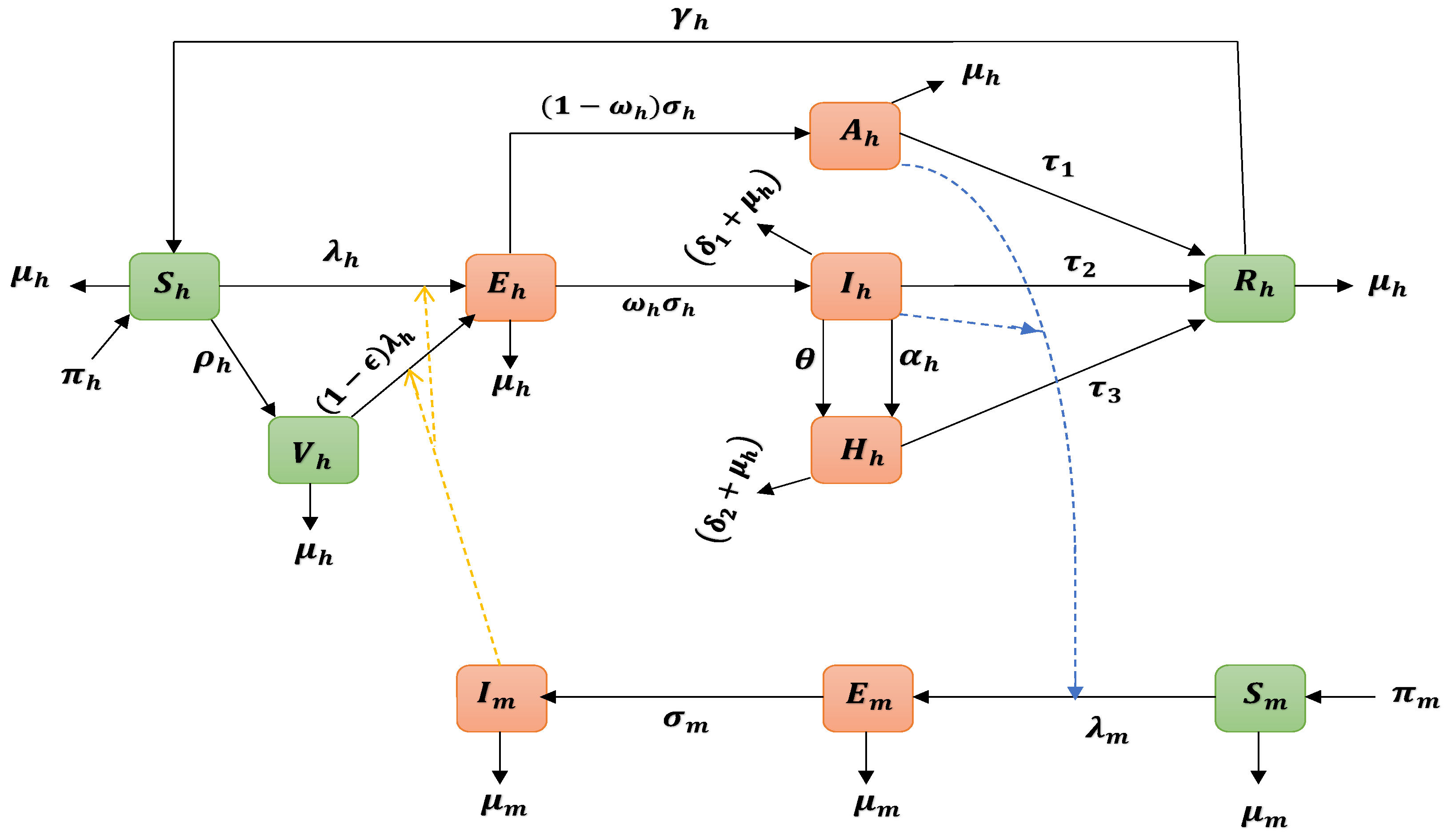

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description |
|---|---|
| Total human population | |
| Susceptible humans who are at risk of contracting the YFV | |
| Population of vaccinated susceptible humans against YFV infection | |
| Population of humans exposed to YFV | |
| Population of asymptomatically infected humans with YFV | |
| Population of symptomatically infected humans with YFV | |
| Population of hospitalized/isolated humans | |
| Population of recovered humans | |
| Total mosquito population | |
| Population of susceptible mosquitoes | |
| Population of mosquitoes exposed to YFV | |
| Population of YFV-infected mosquitoes |
| Parameter | Interpretation/Description |
|---|---|
| Recruitment rate of humans | |
| Recruitment rate of mosquitoes | |
| Natural death rate of humans | |
| Transmission probability from infectious mosquitoes to susceptible humans | |
| b | Mosquito biting rate |
| Modification parameter for the decrease in infectiousness of | |
| Vaccination rate | |
| Vaccine efficacy | |
| Fraction of humans exposed to YFV | |
| Rate of exposed humans becoming infected humans | |
| Hospitalization rate | |
| Rate of active case detection | |
| YFV-induced death rate | |
| Recovery rate of infected humans | |
| Rate of immunity loss | |
| Recruitment rate of mosquitoes | |
| Mosquito natural death rate | |
| Probability of transmission from infected humans to susceptible mosquitos | |
| Rate of exposed mosquitoes becoming infected mosquitoes | |
| m | Average mosquito to human ratio |
2.2. Model’s Preliminary Qualitative Properties
3. Theoretical Analysis
3.1. Disease-Free Equilibrium and Basic Reproduction Number
- i.
- The term represents the rate at which new mosquitoes become infected due to contact with an infected human host during the host’s period of exposure to the YFV. Here, b is the rate at which mosquitoes bite humans, is the probability of YFV transmission from humans to mosquitoes per contact, is the rate at which mosquitoes become infectious after acquiring the virus, is the initial population of susceptible mosquitoes, is a model parameter related to the mosquito population dynamics, and is the natural death rate of mosquitoes.
- ii.
- The expression represents the rate of new human hosts becoming infected due to contact with a diseased mosquito during the mosquito’s expected infectious period. Here, b is the biting rate of mosquitoes, is the probability of YFV transmission from mosquitoes to humans per contact, is the natural death rate of humans, and is the recruitment rate of humans into the susceptible population.
- iii.
- The term signifies the average duration of the infectious stage for humans, where is the rate at which infected individuals develop clinical symptoms of YF, and is a model parameter related to the duration of the infectious period.
- iv.
- The expression represents the probability of vaccinated humans transitioning to the exposed class, with being the vaccination ratio of susceptible humans, related to vaccination dynamics, the natural death rate of humans, and a model parameter associated with transitions between compartments.
- v.
- The term indicates the likelihood that an individual, after exposure to YFV, survives the asymptomatic infectious stage before moving into the recovery class. Here, and are model parameters related to the progression of the disease and recovery.
- vi.
- The expression calculates the probability that an individual exposed to YFV survives the symptomatic infectious stage and transitions into the recovery class, facilitated by ACD. represents the fraction of individuals progressing to recovery, while and are model parameters associated with disease progression and recovery facilitated by interventions.
3.2. Endemic Equilibrium
3.3. Stability Analysis of the Endemic Equilibrium
3.4. Optimal Control Analysis
3.4.1. Existence of Optimal Control
3.4.2. Hamiltonian and Optimality System
- The optimality conditions is given as ,
- Furthermore, the control is given as
4. Numerical Results
4.1. Fitting of Biological Parameters
4.2. Numerical Simulations of the Dynamic Model
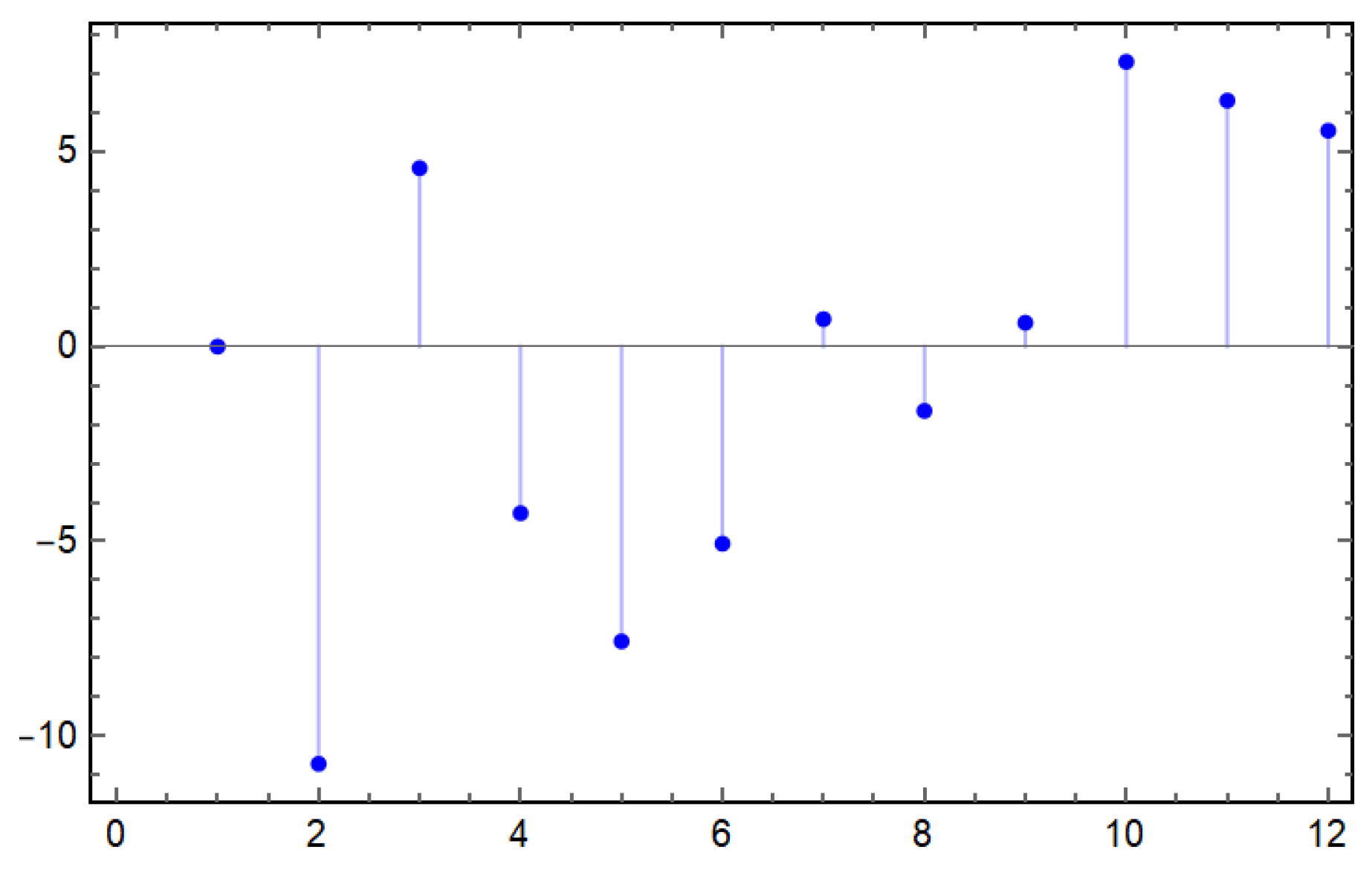
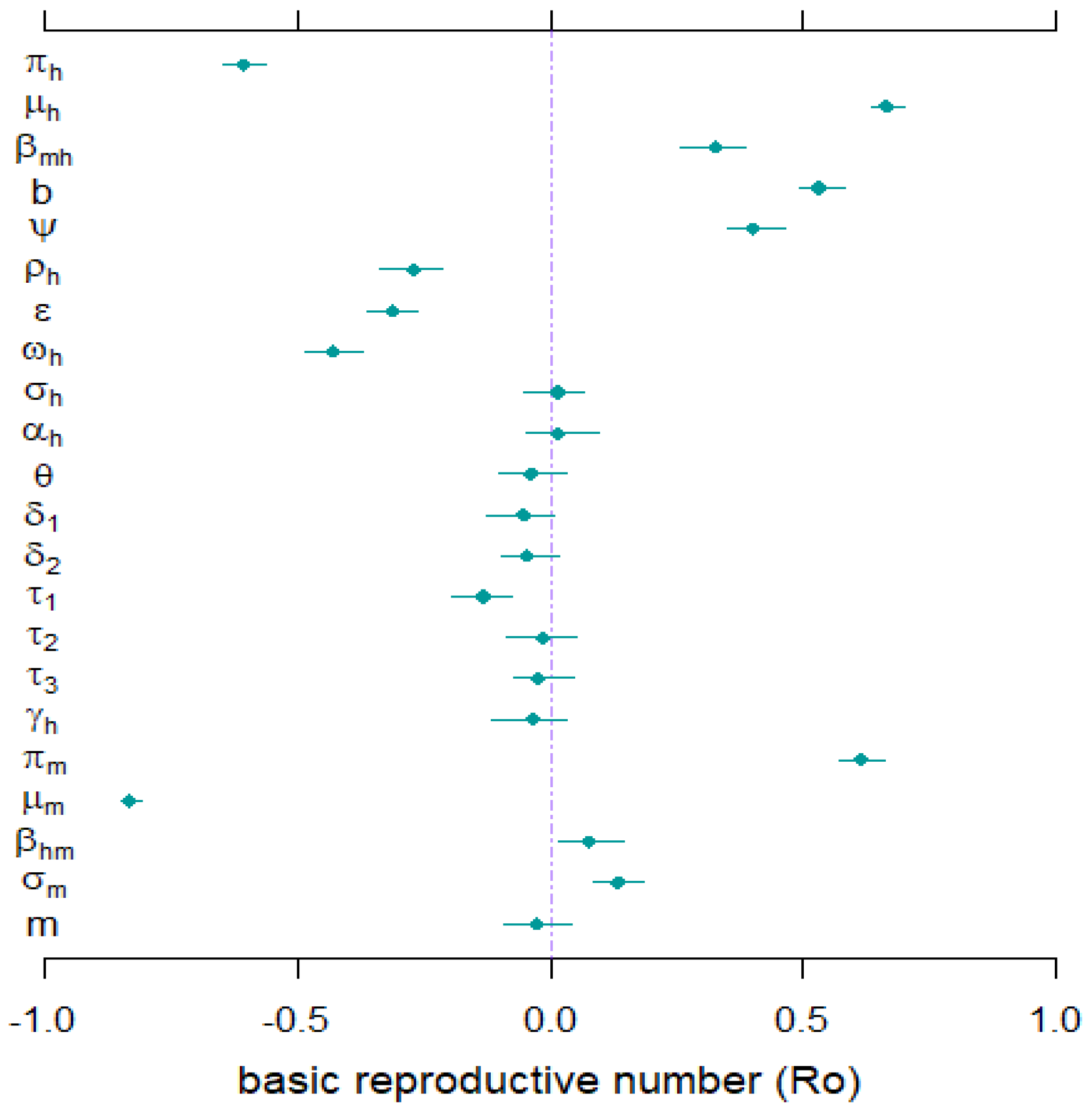
4.3. Sensitivity Analysis
5. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Coefficient of the Polynomial (7)
Appendix B. Summary Tables of the Number of Possible Positive Real Roots of Equation (7)
| Case | No. of Sign Changes | Possible + Real Roots | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | + | + | + | + | + | 0 | 0 | |
| 2 | + | + | + | + | − | 1 | 1 | |
| 3 | + | − | − | − | − | 1 | 1 | |
| 4 | + | + | − | − | − | 1 | 1 | |
| 5 | + | + | + | − | − | 1 | 1 | |
| 6 | + | − | − | − | + | 2 | 0, 2 | |
| 7 | + | − | − | + | + | 2 | 0, 2 | |
| 8 | + | − | − | + | + | 2 | 0, 2 | |
| 9 | + | + | + | − | + | 2 | 0, 2 | |
| 10 | + | + | − | + | + | 2 | 0, 2 | |
| 11 | + | − | + | + | + | 2 | 0, 2 | |
| 12 | + | − | + | − | − | 3 | 1, 3 | |
| 13 | + | − | − | + | − | 3 | 1, 3 | |
| 14 | + | + | − | + | − | 3 | 1, 3 | |
| 15 | + | − | + | + | − | 3 | 1, 3 | |
| 16 | + | − | + | − | + | 4 | 0, 2, 4 | |
| 17 | − | − | − | − | − | 0 | 0 | |
| 18 | − | + | + | + | + | 1 | 1 | |
| 19 | − | − | − | − | + | 1 | 1 | |
| 20 | − | − | − | + | + | 1 | 1 | |
| 21 | − | − | − | + | + | 1 | 1 | |
| 22 | − | − | + | + | + | 1 | 1 | |
| 23 | − | + | + | + | − | 2 | 0, 2 | |
| 24 | − | + | − | − | − | 2 | 0, 2 | |
| 25 | − | − | + | − | − | 2 | 0, 2 | |
| 26 | − | − | − | + | − | 2 | 0, 2 | |
| 27 | − | + | + | − | − | 2 | 0, 2 | |
| 28 | − | − | + | + | − | 2 | 0, 2 | |
| 29 | − | − | + | − | + | 3 | 1, 3 | |
| 29 | − | + | + | − | + | 3 | 1, 3 | |
| 31 | − | + | − | + | + | 3 | 1, 3 | |
| 32 | − | + | − | + | − | 4 | 0, 2, 4 |
Appendix C. The Proof of Theorem 2
Appendix D. Proof of Theorem 3
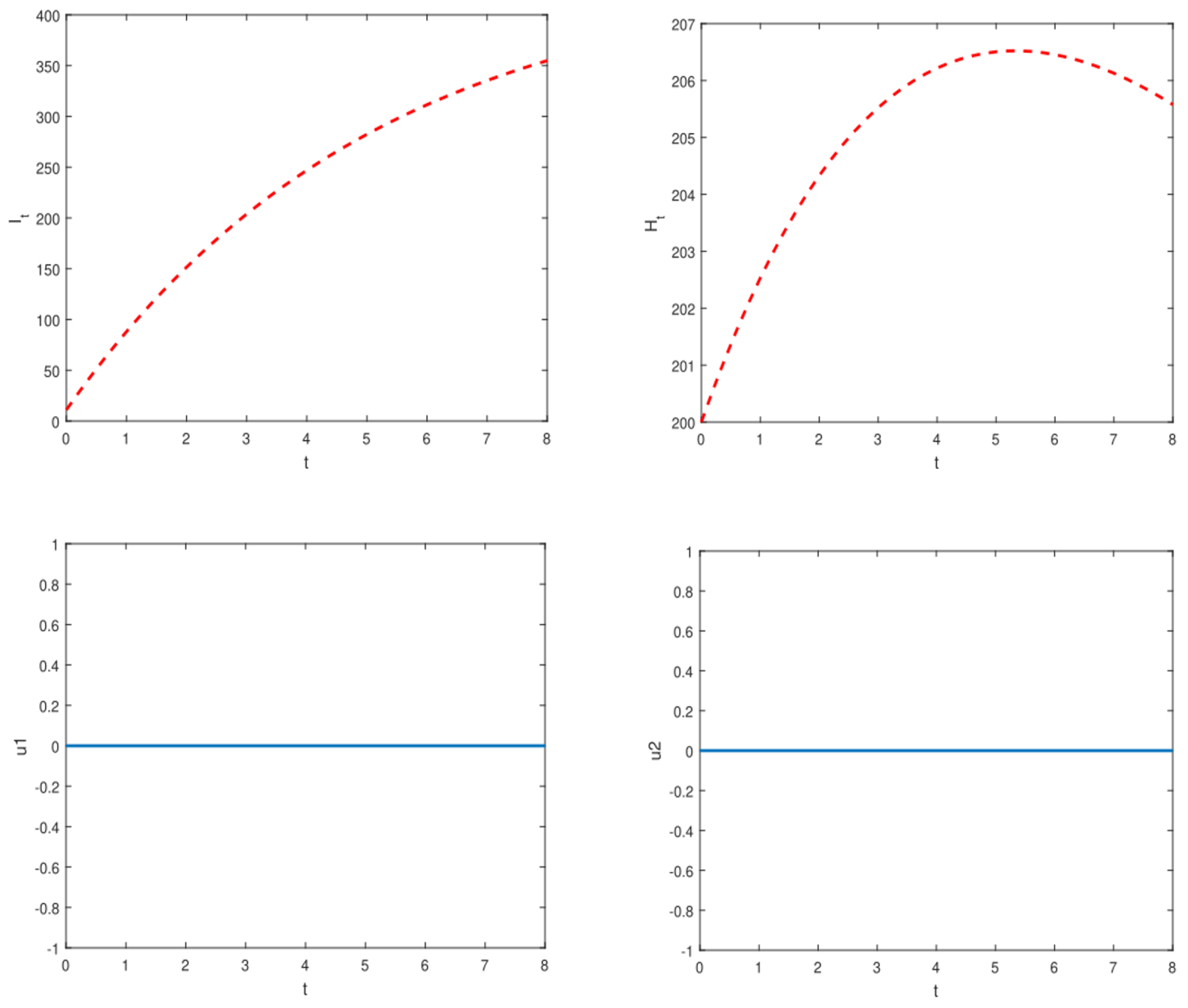
Appendix E. Numerical Illustration of the Optimal Control Problems

Appendix F. Summary Tables for Parameter Estimates
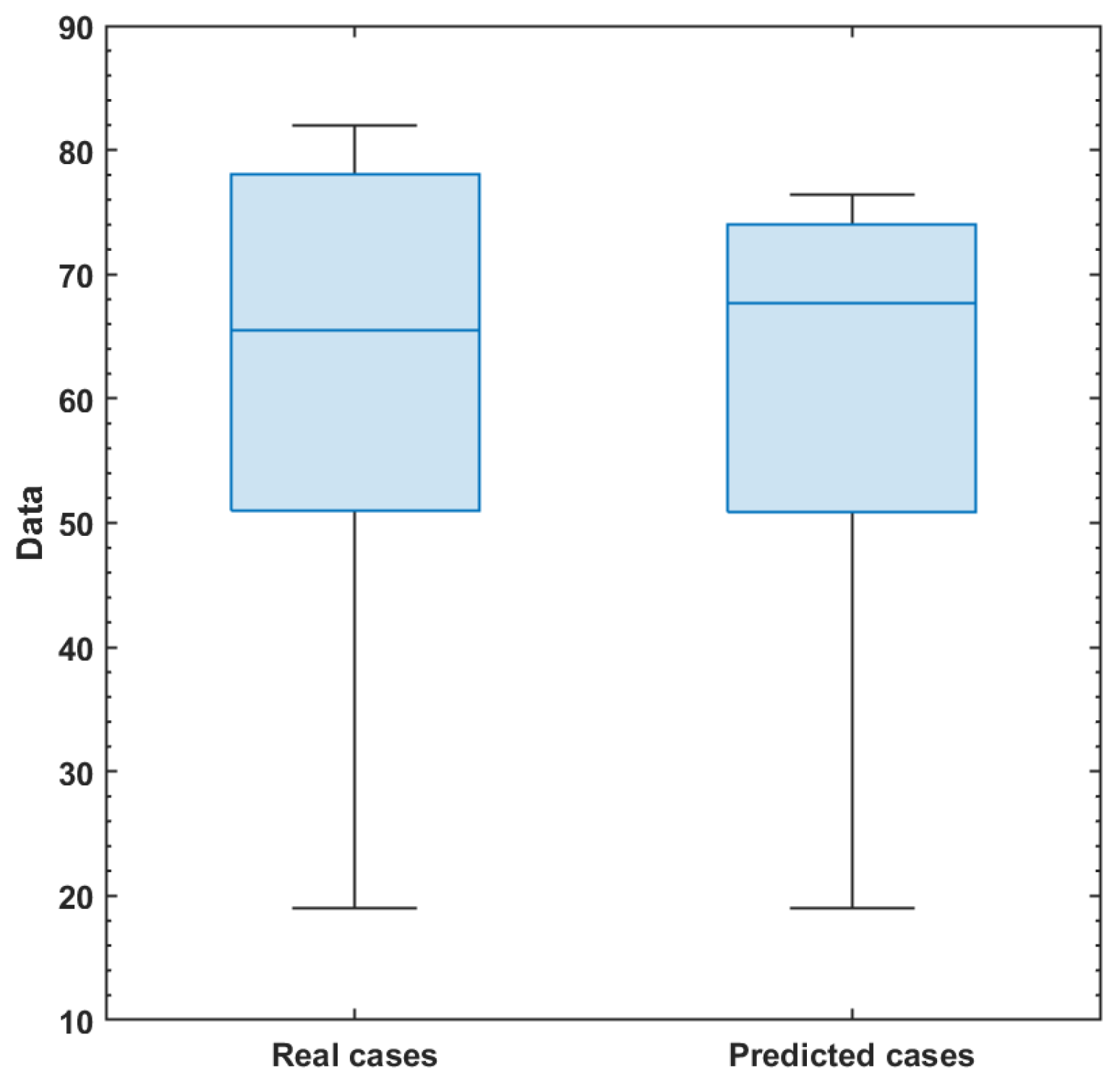
Appendix F.1. Estimates of Comparison Between Number of Actual Symptomatically Infected Individuals and Predicted Ones
| Real Cases | Predicted | Standard Error | Confidence Interval |
|---|---|---|---|
| 19. | 19. | 6.74915 | {3.43642, 34.5636} |
| 23. | 33.7107 | 6.92959 | {17.7311, 49.6904} |
| 51. | 46.4193 | 7.82933 | {28.3648, 64.4737} |
| 51. | 55.278 | 7.09286 | {38.9219, 71.6342} |
| 54. | 61.5672 | 7.38555 | {44.5361, 78.5983} |
| 61. | 66.0599 | 8.95316 | {45.4139, 86.706} |
| 70. | 69.296 | 7.95725 | {50.9465, 87.6454} |
| 70. | 71.6518 | 7.96207 | {53.2912, 90.0124} |
| 74. | 73.3897 | 8.38355 | {54.0572, 92.7223} |
| 82. | 74.6924 | 8.30785 | {55.5345, 93.8503} |
| 82. | 75.6867 | 7.5411 | {58.2969, 93.0765} |
| 82. | 76.4607 | 8.12709 | {57.7196, 95.2018} |
Appendix F.2. Summary Results for Actual and Predicted Data
| Data | Min. | 1st Qu. | Median | Mean | 3rd Qu. | Max. | SD |
|---|---|---|---|---|---|---|---|
| Real cases | 1.90 | 5.10 | 6.55 | 5.99 | 7.80 | 8.20 | 2.15 |
| Predicted | 1.90 | 5.08 | 6.77 | 6.03 | 7.40 | 7.65 | 1.85 |
References
- World Health Organization. Yellow Fever. WHO Fact Sheets. 2019. Available online: https://www.who.int/news-room/fact-sheets/detail/yellow-fever (accessed on 11 November 2019).
- Kung’Aro, M.; Luboobi, L.S.; Shahada, F. Modelling and stability analysis of SVEIRS yellow fever two-host model. Gulf J. Math. 2015, 3, 106–129. [Google Scholar] [CrossRef]
- Monath, T.P.; Vasconcelos, P.F.C. Yellow fever. J. Clinical Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef]
- Zhao, S.; Stone, L.; Gao, D.; He, D. Modelling the large-scale yellow fever outbreak in Luanda, Angola, and the impact of vaccination. PLoS Negl. Trop. Dis. 2018, 12, e0006158. [Google Scholar] [CrossRef]
- World Health Organization. Yellow Fever-Nigeria. WHO Reports. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2020-DON299 (accessed on 11 November 2019).
- Duijzer, L.E.; van Jaasveld, W.; Dekker, R. Literature review: The vaccine supply chain. Eur. J. Oper. Res. 2018, 268, 174–192. [Google Scholar] [CrossRef]
- Robbins, M.J.; Lunday, B.J. A bilevel formulation of the pediatric vaccine pricing problem. Eur. J. Oper. Res. 2016, 248, 634–645. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Faria, N.R.; Reiner, J.R.; Golding, N.; Nikolay, B.; Stasse, S.; Johansson, M.A.; Salje, H.; Faye, O.; Wint, G.W.; et al. Spread of yellow fever virus outbreak in Angola and the Democratic Republic of the Congo 2015–16: A modelling study. Lancet Infect. Dis. 2017, 17, 330–338. [Google Scholar] [CrossRef]
- Abdulrashid, I.; Tomas, C.; Xiaoying, H. Effects of delays in mathematical models of cancer chemotherapy. Pure Appl. Funct. Anal. 2022, 7, 1103–1126. [Google Scholar]
- Abdulrashid, I.; Han, X. A mathematical model of chemotherapy with variable infusion. Commun. Pure Appl. Anal. 2020, 19, 1875–1890. [Google Scholar] [CrossRef]
- Abdulrashid, I.; Alsammani, A.M.; Han, X. Stability analysis of a chemotherapy model with delays. Disc. Cont. Dyn. Syst.-B 2019, 24, 989–1005. [Google Scholar] [CrossRef]
- Abdulrashid, I.; Ghazzai, H.; Han, X.; Massoud, Y. Optimal Control Treatment Analysis for the Predator-Prey Chemotherapy Model. In Proceedings of the 2019 31st International Conference on Microelectronics (ICM), Cairo, Egypt, 15–18 December 2019; pp. 296–299. [Google Scholar]
- Al-Shomrani, M.M.; Musa, S.S.; Yusuf, A. Unfolding the transmission dynamics of monkeypox virus: An epidemiological modelling analysis. Mathematics 2023, 11, 1121. [Google Scholar] [CrossRef]
- Gu, Y.; Khan, M.; Zarin, R.; Khan, A.; Yusuf, A.; Humphries, U.W. Mathematical analysis of a new nonlinear dengue epidemic model via deterministic and fractional approach. Alexandria Eng. J. 2023, 67, 1–21. [Google Scholar] [CrossRef]
- Zarin, R.; Khan, M.; Khan, A.; Yusuf, A. Deterministic and fractional analysis of a newly developed dengue epidemic model. Waves Random Complex Media 2023, 1–34. [Google Scholar] [CrossRef]
- Adel, M.; Khader, M.M.; Ahmad, H.; Assiri, T.A. Approximate analytical solutions for the blood ethanol concentration system and predator-prey equations by using variational iteration method. AIMS Math. 2023, 8, 19083–19096. [Google Scholar] [CrossRef]
- Shearer, F.M.; Longbottom, J.; Browne, A.J.; Pigott, D.M.; Brady, O.J.; Kraemer, M.U.; Marinho, F.; Yactayo, S.; de Araújo, V.E.; da Nóbrega, A.A.; et al. Existing and potential infection risk zones of yellow fever worldwide: A modelling analysis. Lancet Glob. Health 2018, 6, e270–e278. [Google Scholar] [CrossRef]
- Teklu, S.W.; Rao, K.P. HIV/AIDS–Pneumonia Codynamics Model Analysis with Vaccination and Treatment. Comput. Math. Methods Med. 2022, 2022, 3105734. [Google Scholar] [CrossRef]
- Alshehri, A.; El Hajji, M. Mathematical study for Zika virus transmission with general incidence rate. AIMS Math. 2022, 7, 7117–7142. [Google Scholar] [CrossRef]
- Fraser, K.; Hamlet, A.; Jean, K.; Ramos, D.G.; Romano, A.; Horton, J.; Cibrelus, L.; Ferguson, N.; Gaythorpe, K.A. Assessing yellow fever outbreak potential and implications for vaccine strategy. medRxiv 2023. [Google Scholar] [CrossRef]
- Kung’aro, M. Mathematical Modelling of Intra and Inter Dynamics and Control of Yellow Fever in Primate and Human Populations. Ph.D. Thesis, The Nelson Mandela African Institution of Science and Technology, Arusha, Tanzania, 2016. [Google Scholar] [CrossRef]
- Caasi, J.A.; Joseph, B.M.; Kodiyamplakkal, H.J.; Manibusan, J.R.; Camacho Aquino, L.J.; Oh, H.; Rychtář, J.; Taylor, D. A game-theoretic model of voluntary yellow fever vaccination to prevent urban outbreaks. Games 2022, 13, 55. [Google Scholar] [CrossRef]
- Ghosh, I.; Tiwari, P.K.; Chattopadhyay, J. Effect of active case finding on dengue control: Implications from a mathematical model. J. Theoret. Biol. 2019, 464, 50–62. [Google Scholar] [CrossRef]
- Nigeria Centre for Disease Control and Prevention. Yellow Fever. Available online: https://www.ncdc.gov.ng/ncdc.gov.ng/diseases/sitreps/?cat=10&name=An%20update%20of%20Yellow%20Fever%20outbreak%20in%20Nigeria (accessed on 4 August 2022).
- Nigeria Centre for Disease Control. Disease Situation Report: An Update of Lassa Fever Outbreak in Nigeria. 2018. Available online: https://www.ncdc.gov.ng/ncdc.gov.ng/diseases/sitreps (accessed on 6 March 2022).
- World Health Organization. Yellow Fever. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/yellow-fever (accessed on 2 December 2021).
- Hussaini, N.; Lubuma, J.M.; Barley, K.; Gumel, A.B. Mathematical analysis of a model for AVL-HIV co-endemicity. Math. Biosci. 2016, 271, 80–95. [Google Scholar] [CrossRef]
- Diekmann, O.; Heesterbeek, J.; Metz, J. On the definition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations. J. Math. Biol. 1990, 28, 365–382. [Google Scholar] [CrossRef] [PubMed]
- van den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Chowell, G.; Viboud, C.; Simonsen, L.; Moghadas, S.M. Characterizing the reproduction number of epidemics with early subexponential growth dynamics. J. R. Soc. Interface 2016, 13, 0659. [Google Scholar] [CrossRef]
- Yang, C.; Wang, X.; Gao, D.; Wang, J. Impact of awareness programs on cholera dynamics: Two modeling approaches. Bull. Math. Biol. 2017, 79, 2109–2131. [Google Scholar] [CrossRef]
- Musa, S.S.; Zhao, S.; Hussaini, N.; Habib, A.G.; He, D. Mathematical modeling and analysis of meningococcal meningitis transmission dynamics. Int. J. Biomath. 2020, 13, 2050006. [Google Scholar] [CrossRef]
- Wang, X. A simple proof of Descartes’s rule of signs. Am. Math. Mon. 2004, 111, 525. [Google Scholar] [CrossRef]
- Molnar, S.; Szidarovsky, F. Some notes on the time-variant Lyapunov theory. Eur. J. Oper. Res. 1996, 89, 668–670. [Google Scholar] [CrossRef]
- Tadic, V. Stochastic gradient algorithm with random truncations. Eur. J. Oper. Res. 1997, 101, 261–284. [Google Scholar] [CrossRef]
- Sun, G.Q.; Xie, J.H.; Huang, S.H.; Jin, Z.; Li, M.T.; Liu, L. Transmission dynamics of cholera: Mathematical modeling and control strategies. Commun. Non. Sci. Numer. Simulat. 2017, 45, 235–244. [Google Scholar] [CrossRef]
- Farouk, T.S.; Evren, H. An optimal control approach for the interaction of immune checkpoints, immune system, and BCG in the treatment of superficial bladder cancer. Eur. Phys. J. Plus 2018, 133, 241. [Google Scholar]
- Kopp, R.E. Pontryagin maximum principle. In Mathematics in Science and Engineering; Elsevier: Amsterdam, The Netherlands, 1962; Volume 5, pp. 255–279. [Google Scholar]
- Garba, S.M.; Lubuma, J.M.; Tsanou, B. Modeling the transmission dynamics of the COVID-19 Pandemic in South Africa. Math. Biosci. 2020, 328, 108441. [Google Scholar] [CrossRef] [PubMed]
- Hussaini, N.; Okuneye, K.; Gumel, A.B. Mathematical analysis of a model for zoonotic visceral leishmaniasis. Infect. Dis. Model. 2017, 2, 455–474. [Google Scholar] [CrossRef] [PubMed]
- Musa, S.S.; Zhao, S.; Hussaini, N.; Usaini, S.; He, D. Dynamics analysis of typhoid fever with public health education programs and final epidemic size relation. Results Appl. Math. 2021, 10, 100153. [Google Scholar] [CrossRef]
- He, D.; Zhao, S.; Lin, Q.; Musa, S.S.; Stone, L. New estimates of the Zika virus epidemic attack rate in Northeastern Brazil from 2015 to 2016: A modelling analysis based on Guillain-Barré Syndrome (GBS) surveillance data. PLoS Negl. Trop. Dis. 2020, 14, e0007502. [Google Scholar] [CrossRef]
- Roop-O, P.; Chinviriya, W.; Chinviriyasit, S. The effect of incidence function in backward bifurcation for malaria model with temporary immunity. Math. Biosci. 2015, 265, 47–64. [Google Scholar] [CrossRef]
- Musa, S.S.; Zhao, S.; Chan, H.S.; Jin, Z.; He, D. A mathematical model to study the 2014–2015 large-scale dengue epidemics in Kaohsiung and Tainan cities in Taiwan, China. Math. Biosci. Eng. 2019, 16, 3841–3863. [Google Scholar] [CrossRef] [PubMed]
- Garba, S.M.; Gumel, A.B.; Bukar, M.R.A. Backward bifurcations in dengue transmission dynamics. Math. Biosci. 2008, 215, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Musa, S.S.; Baba, I.A.; Yusuf, A.; Sulaiman, T.A.; Aliyu, A.I.; Zhao, S.; He, D. Transmission dynamics of SARS-CoV-2: A modeling analysis with high- and moderate-risk populations. Results Phys. 2021, 26, 104290. [Google Scholar] [CrossRef] [PubMed]
- Baba, I.A.; Yusuf, A.; Nisar, K.S.; Abdel-Atye, A.H.; Nofal, T.A. Mathematical model to assess the imposition of lockdown during COVID-19 pandemic. Results Phys. 2021, 20, 103716. [Google Scholar] [CrossRef]
- Lu, X.; Borgonovo, E. Global sensitivity analysis in epidemiological modeling. Eur. J. Oper. Res. 2023, 304, 9–24. [Google Scholar] [CrossRef]
- Gao, D.; Lou, Y.; He, D.; Porco, T.C.; Kuang, Y.; Chowell, G.; Ruan, S. Prevention and control of Zika as a mosquito-borne and sexually transmitted disease: A mathematical modeling analysis. Sci. Rep. 2016, 6, 28070. [Google Scholar] [CrossRef] [PubMed]
- Musa, S.S.; Zhao, S.; Gao, D.; Lin, Q.; Chowell, G.; He, D. Mechanistic modelling of the large-scale Lassa fever epidemics in Nigeria from 2016 to 2019. J. Theoret. Biol. 2020, 493, 110209. [Google Scholar] [CrossRef] [PubMed]
- Handari, B.D.; Aldila, D.; Dewi, B.O.; Rosuliyana, H.; Khosnaw, S. Analysis of yellow fever prevention strategy from the perspective of mathematical model and cost-effectiveness analysis. Math. Biosci. Eng. 2022, 19, 1786–1824. [Google Scholar] [CrossRef] [PubMed]
- Gaythorpe, K.A.; Jean, K.; Cibrelus, L.; Garske, T. Quantifying model evidence for yellow fever transmission routes in Africa. PLoS Comput. Biol. 2019, 15, e1007355. [Google Scholar] [CrossRef] [PubMed]
- Laiton-Donato, K.; Quintero-Cortés, P.; Franco-Salazar, J.P.; Peláez-Carvajal, D.; Navas, M.C.; Junglen, S.; Parra-Henao, G.; Usme-Ciro, J.A. Usefulness of an in vitro-transcribed RNA control for the detection and quantification of Yellow fever virus through real-time reverse transcription-polymerase chain reaction. Infect. Dis. Now 2023, 53, 104654. [Google Scholar] [CrossRef]
- Bonin, C.R.; dos Santos, R.W.; Fernandes, G.C.; Lobosco, M. Computational modeling of the immune response to yellow fever. J. Comput. Appl. Math. 2016, 295, 127–138. [Google Scholar] [CrossRef]
- Jiang, N.; Chu, W.; Li, Y. Modeling, Inference, and Prediction in Mobility-Based Compartmental Models for Epidemiology. arXiv 2024, arXiv:2406.12002. [Google Scholar]
- Menkir, T.F.; Jbaily, A.; Verguet, S. Incorporating equity in infectious disease modeling: Case study of a distributional impact framework for measles transmission. Vaccine 2021, 39, 2894–2900. [Google Scholar] [CrossRef]
- LaSalle, J.P. The Stability of Dynamical Systems; Regional Conference Series in Applied Mathematics; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 1976. [Google Scholar]



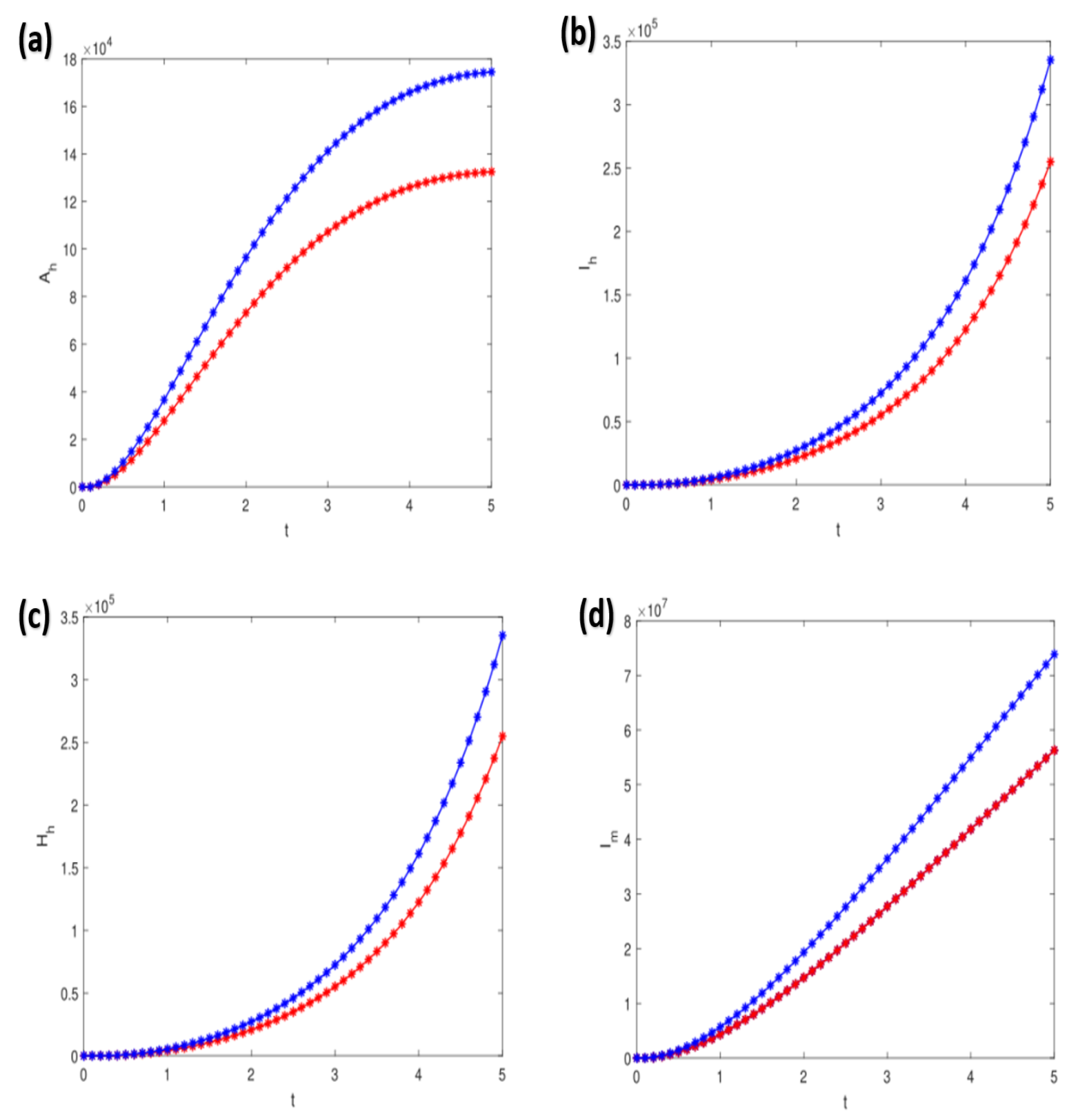
| Estimate | Standard Error | t-Statistic | p-Value | Confidence Interval | |
|---|---|---|---|---|---|
| 0.630716 | 0.174866 × | 3.60686 | 6.91309 × | {2.27475 × , 1.03396} | |
| 0.803653 | 4.88085 × | 16.4654 | 1.86704 × | {6.911 × , 9.16205 × } | |
| 2.03782 | 4.06303 × | 5.01551 | 1.03264 × | {1.10088, 2.97476} | |
| 2.87329 | 2.77785 × | 10.3436 | 6.59241 × | {2.23271, 3.51386} |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsowait, N.L.; Al-Shomrani, M.M.; Abdulrashid, I.; Musa, S.S. Synergistic Impact of Active Case Detection and Early Hospitalization for Controlling the Spread of Yellow Fever Outbreak in Nigeria: An Epidemiological Modeling and Optimal Control Analysis. Mathematics 2024, 12, 3817. https://doi.org/10.3390/math12233817
Alsowait NL, Al-Shomrani MM, Abdulrashid I, Musa SS. Synergistic Impact of Active Case Detection and Early Hospitalization for Controlling the Spread of Yellow Fever Outbreak in Nigeria: An Epidemiological Modeling and Optimal Control Analysis. Mathematics. 2024; 12(23):3817. https://doi.org/10.3390/math12233817
Chicago/Turabian StyleAlsowait, Nawaf L., Mohammed M. Al-Shomrani, Ismail Abdulrashid, and Salihu S. Musa. 2024. "Synergistic Impact of Active Case Detection and Early Hospitalization for Controlling the Spread of Yellow Fever Outbreak in Nigeria: An Epidemiological Modeling and Optimal Control Analysis" Mathematics 12, no. 23: 3817. https://doi.org/10.3390/math12233817
APA StyleAlsowait, N. L., Al-Shomrani, M. M., Abdulrashid, I., & Musa, S. S. (2024). Synergistic Impact of Active Case Detection and Early Hospitalization for Controlling the Spread of Yellow Fever Outbreak in Nigeria: An Epidemiological Modeling and Optimal Control Analysis. Mathematics, 12(23), 3817. https://doi.org/10.3390/math12233817