

Coexisting Attractors and Multistate Noise-Induced Intermittency in a Cycle Ring of Rulkov Neurons




{kind=link}
{kind=link}
{kind=link}
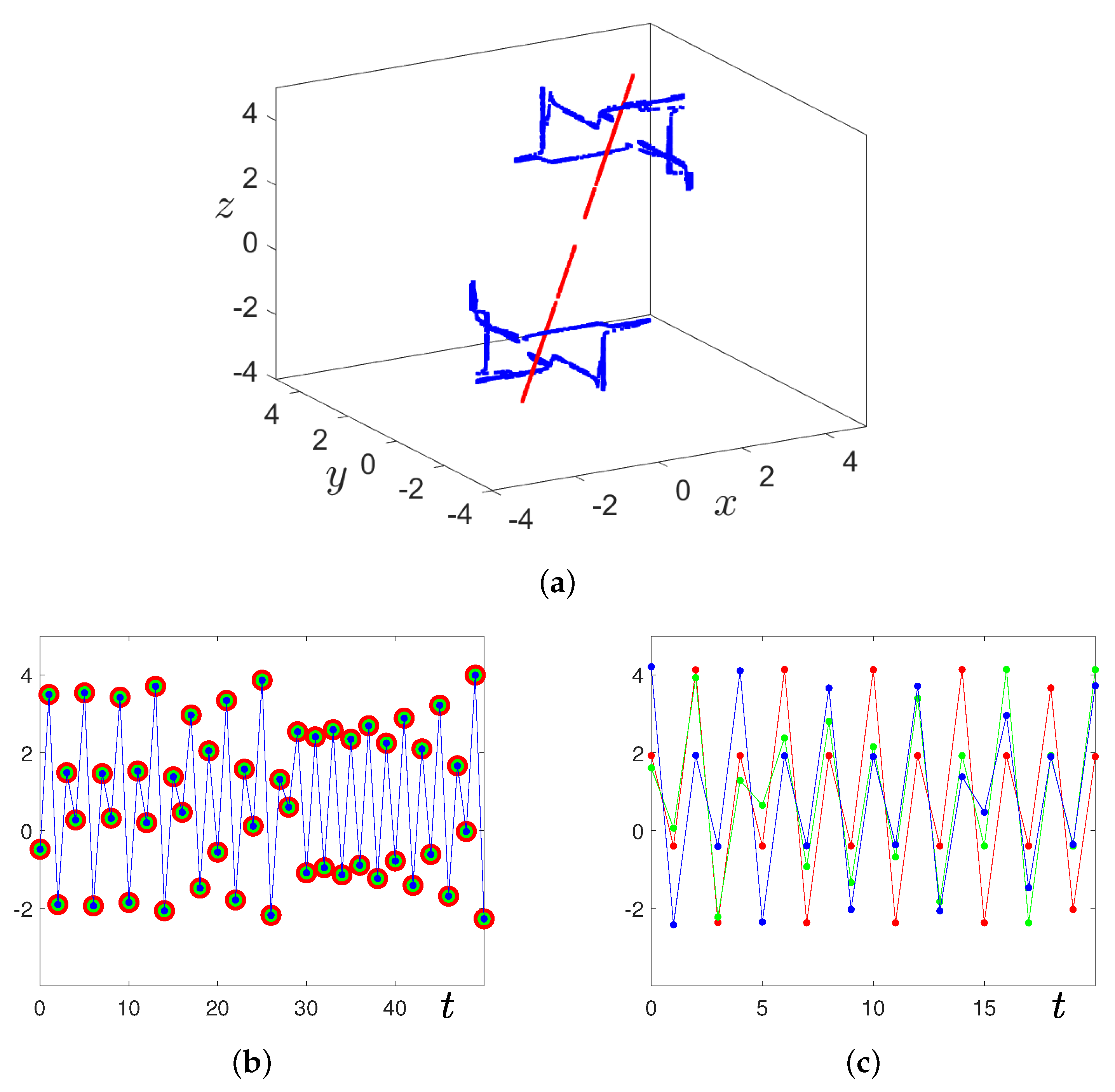{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Deterministic System
3. Stochastic System
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pisarchik, A.N.; Hramov, A.E. Multistability in Physical and Living Systems: Characterization and Applications; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Postnov, D.E.; Vadivasova, T.E.; Sosnovtseva, O.V.; Balanov, A.G.; Anishchenko, V.S.; Mosekilde, E. Role of multistability in the transition to chaotic phase synchronization. Phys. Rev. Lett. 1999, 9, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Astakhov, V.; Shabunin, A.; Uhm, W.; Kim, S. Multistability formation and synchronization loss in coupled Hénon maps: Two sides of the single bifurcational mechanism. Phys. Rev. E 2001, 63, 056212. [Google Scholar] [CrossRef]
- Carvalho, R.; Fernandez, B.; Mendes, R.M. From synchronization to multistability in two coupled quadratic maps. Phys. Lett. A 2001, 285, 327–338. [Google Scholar] [CrossRef]
- Barba-Franco, J.J.; Gallegos, A.; Jaimes-Reátegui, R.; Gerasimova, S.A.; Pisarchik, A.N. Dynamics of a ring of three unidirectionally coupled Duffing oscillators with time-dependent damping. Europhys. Lett. 2021, 134, 30005. [Google Scholar] [CrossRef]
- Bashkirtseva, I.; Ryashko, L.; Pisarchik, A.N. Stochastic transitions between in-phase and anti-phase synchronization in coupled map-based neural oscillators. Commun. Nonlinear Sci. Numer. Simulat. 2021, 95, 105611. [Google Scholar] [CrossRef]
- Pisarchik, A.N.; Jaimes-Reátegui, R.; Villalobos-Salazar, J.R.; García-López, J.H.; Boccaletti, S. Synchronization of chaotic systems with coexisting attractors. Phys. Rev. Lett. 2006, 96, 244102. [Google Scholar] [CrossRef]
- Boccaletti, S.; Pisarchik, A.N.; del Genio, C.I.; Amann, A. Synchronization: From Coupled Systems to Complex Networks; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]
- Elson, R.C.; Selverston, A.I.; Huerta, R.; Rulkov, N.F.; Rabinovich, M.I.; Abarbanel, H.D.I. Synchronous behavior of two coupled biological neurons. Phys. Rev. Lett. 1998, 81, 5692. [Google Scholar] [CrossRef]
- Rulkov, N.F.; Timofeev, I.; Bazhenov, M. Oscillations in large-scale cortical networks: Map-based model. J. Comput. Neurosci. 2004, 17, 203. [Google Scholar] [CrossRef]
- Sun, X.; Lu, Q.; Kurths, J.; Wang, Q. Spatiotemporal coherence resonance in a map lattice. Int. J. Bifurc. Chaos 2009, 19, 737. [Google Scholar] [CrossRef]
- Sausedo-Solorio, J.M.; Pisarchik, A.N. Synchronization of map-based neurons with memory and synaptic delay. Phys. Lett. A 2014, 378, 2108. [Google Scholar] [CrossRef]
- Lodato, I.; Boccaletti, S.; Latora, V. Synchronization properties of network motifs. Europhys. Lett. 2007, 78, 28001. [Google Scholar] [CrossRef]
- Lang, X.; Lu, Q.; Kurths, J. Phase synchronization in noise-driven bursting neurons. Phys. Rev. E 2010, 82, 021909. [Google Scholar] [CrossRef]
- Matias, F.S.; Carelli, P.V.; Mirasso, C.; Copelli, R.M. Anticipated synchronization in a biologically plausible model of neuronal motifs. Phys. Rev. E 2011, 84, 021922. [Google Scholar] [CrossRef]
- Andreev, A.V.; Maksimenko, V.A.; Pisarchik, A.N.; Hramov, A.E. Synchronization of interacted spiking neuronal networks with inhibitory coupling. Chaos Solitons Fractals 2021, 146, 110812. [Google Scholar] [CrossRef]
- Turing, A.M. The chemical basis of morphogenesis. Philos. Trans. R. Soc. B 1952, 237, 37–72. [Google Scholar]
- Collins, J.J.; Stewart, I. Hexapodal gaits and coupled nonlinear oscillator models. Biol. Cybern. 1993, 68, 287–298. [Google Scholar] [CrossRef]
- Sausedo-Solorio, J.M.; Pisarchik, A.N. Synchronization in network motifs of delay-coupled map-based neurons. Eur. Phys. J. Spec. Top. 2017, 226, 1911–1920. [Google Scholar] [CrossRef]
- Grillner, S.; Wallen, P. Central pattern generators for locomotion, with special reference to vertebrates. Annu. Rev. Neurosci. 1985, 8, 233–261. [Google Scholar] [CrossRef]
- Collins, J.J.; Stewart, I. A group-theoretic approach to rings of coupled biological oscillators. Biol. Cybern. 1994, 71, 95–103. [Google Scholar] [CrossRef]
- Abarbanel, H.D.I.; Rabinovich, M.I.; Selverston, A.; Bazhenov, M.V.; Huerta, R.; Sushchik, M.M.; Rubchinskii, L.L. Synchronisation in neural networks. Physics-Uspekhi 1996, 39, 337–362. [Google Scholar] [CrossRef]
- Shen-Orr, S.S.; Milo, R.; Mangan, S.; Alon, U. Network motifs in the transcriptional regulation network of Escherichia coli. Nat. Genet. 2002, 31, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Pisarchik, A.N.; Hramov, A.E. Coherence resonance in neural networks: Theory and experiments. Phys. Rep. 2023, 1000, 1–57. [Google Scholar] [CrossRef]
- Horsthemke, W.; Lefever, R. Noise-Induced Transitions; Springer: Berlin/Heidelberg, Germany, 1984; p. 338. [Google Scholar]
- Anishchenko, V.S.; Astakhov, V.V.; Neiman, A.B.; Vadivasova, T.E.; Schimansky-Geier, L. Nonlinear Dynamics of Chaotic and Stochastic Systems. Tutorial and Modern Development; Springer: Berlin/Heidelberg, Germany, 2007; p. 535. [Google Scholar]
- Pikovsky, A.S.; Kurths, J. Coherence resonance in a noise-driven excitable system. Phys. Rev. Lett. 1997, 78, 775–778. [Google Scholar] [CrossRef]
- McDonnell, M.D.; Stocks, N.G.; Pearce, C.E.M.; Abbott, D. Stochastic Resonance: From Suprathreshold Stochastic Resonance to Stochastic Signal Quantization; Cambridge University Press: Cambridge, UK, 2008; p. 446. [Google Scholar]
- Bashkirtseva, I.; Pankratov, A.; Ryashko, L. Noise-induced formation of heterogeneous patterns in the Turing stability zones of diffusion systems. J. Phys. 2022, 34, 444001. [Google Scholar] [CrossRef] [PubMed]
- Arnold, L. Random Dynamical Systems; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Gassmann, F. Noise-induced chaos-order transitions. Phys. Rev. E. 1997, 55, 2215–2221. [Google Scholar] [CrossRef]
- Gao, J.B.; Hwang, S.K.; Liu, J.M. When can noise induce chaos? Phys. Rev. Lett. 1999, 82, 1132–1135. [Google Scholar] [CrossRef]
- Lai, Y.C.; Tel, T. Transient Chaos. Complex Dynamics on Finite Time Scales; Springer: New York, NY, USA, 2011; p. 502. [Google Scholar]
- Lindner, B.; Garcia-Ojalvo, J.; Neiman, A.; Schimansky-Geier, L. Effects of noise in excitable systems. Phys. Rep. 2004, 392, 321–424. [Google Scholar] [CrossRef]
- Kloeden, P.E.; Platen, E. Numerical Solution of Stochastic Differential Equations; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Milstein, G.N.; Tretyakov, M.V. Stochastic Numerics for Mathematical Physics; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Freidlin, M.I.; Wentzell, A.D. Random Perturbations of Dynamical Systems; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Bashkirtseva, I.; Neiman, A.B.; Ryashko, L. Stochastic sensitivity analysis of noise-induced suppression of firing and giant variability of spiking in a Hodgkin-Huxley neuron model. Phys. Rev. E 2015, 91, 052920. [Google Scholar] [CrossRef]
- Nekorkin, V.I.; Makarov, V.A.; Velarde, M.G. Spatial disorder and waves in a ring chain of bistable oscillators. Int. J. Bifurc. Chaos 1996, 6, 1845–1858. [Google Scholar] [CrossRef]
- Matías, M.A.; Pérez-Muñuzuri, V.; Lorenzo, M.N.; Mariño, I.P.; Pérez-Villar, V.V. Observation o fa fast rotating wave in rings of coupled chaotic oscillators. Phys. Rev. Lett. 1997, 78, 219–222. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, G.; Cerdeira, H.A. How does a periodic rotating wave emerge from high-dimensional chaos in a ring of coupled chaotic oscillators? Phys. Rev. E 2001, 64, 037203. [Google Scholar] [CrossRef]
- Shimizu, K.; Komuro, M.; Endo, T. Onset of the propagating pulse wave in a ring of coupled bistable oscillators. Nonlinear Theory Its Appl. IEICE 2011, 2, 139–151. [Google Scholar] [CrossRef]
- Kamiyama, K.; Endo, T. Chaos of the propagating pulse wave in a ring of six-coupled bistable oscillators. Int. J. Bifurc. Chaos 2012, 22, 1250091. [Google Scholar] [CrossRef]
- Rulkov, N.F. Regularization of synchronized chaotic bursts. Phys. Rev. Lett. 2001, 86, 183–186. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashkirtseva, I.A.; Pisarchik, A.N.; Ryashko, L.B. Coexisting Attractors and Multistate Noise-Induced Intermittency in a Cycle Ring of Rulkov Neurons. Mathematics 2023, 11, 597. https://doi.org/10.3390/math11030597
Bashkirtseva IA, Pisarchik AN, Ryashko LB. Coexisting Attractors and Multistate Noise-Induced Intermittency in a Cycle Ring of Rulkov Neurons. Mathematics. 2023; 11(3):597. https://doi.org/10.3390/math11030597
Chicago/Turabian StyleBashkirtseva, Irina A., Alexander N. Pisarchik, and Lev B. Ryashko. 2023. "Coexisting Attractors and Multistate Noise-Induced Intermittency in a Cycle Ring of Rulkov Neurons" Mathematics 11, no. 3: 597. https://doi.org/10.3390/math11030597
APA StyleBashkirtseva, I. A., Pisarchik, A. N., & Ryashko, L. B. (2023). Coexisting Attractors and Multistate Noise-Induced Intermittency in a Cycle Ring of Rulkov Neurons. Mathematics, 11(3), 597. https://doi.org/10.3390/math11030597