
Z-Type Control Methods on a Three-Species Model with an Invasive Prey

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Highlights on the Z Control
2.2. The Reference Model
2.3. Invasive Species Removal: Single Indirect Control on Foxes
2.3.1. Z-Controlled Model Design on Foxes
2.3.2. Equilibria
2.3.3. Local Stability Analysis
- If feasible, the hare-free equilibrium is locally asymptotically stable if and only if
- The coexistence equilibrium , when feasible, is instead locally asymptotically stable for every choice of parameters, including λ.
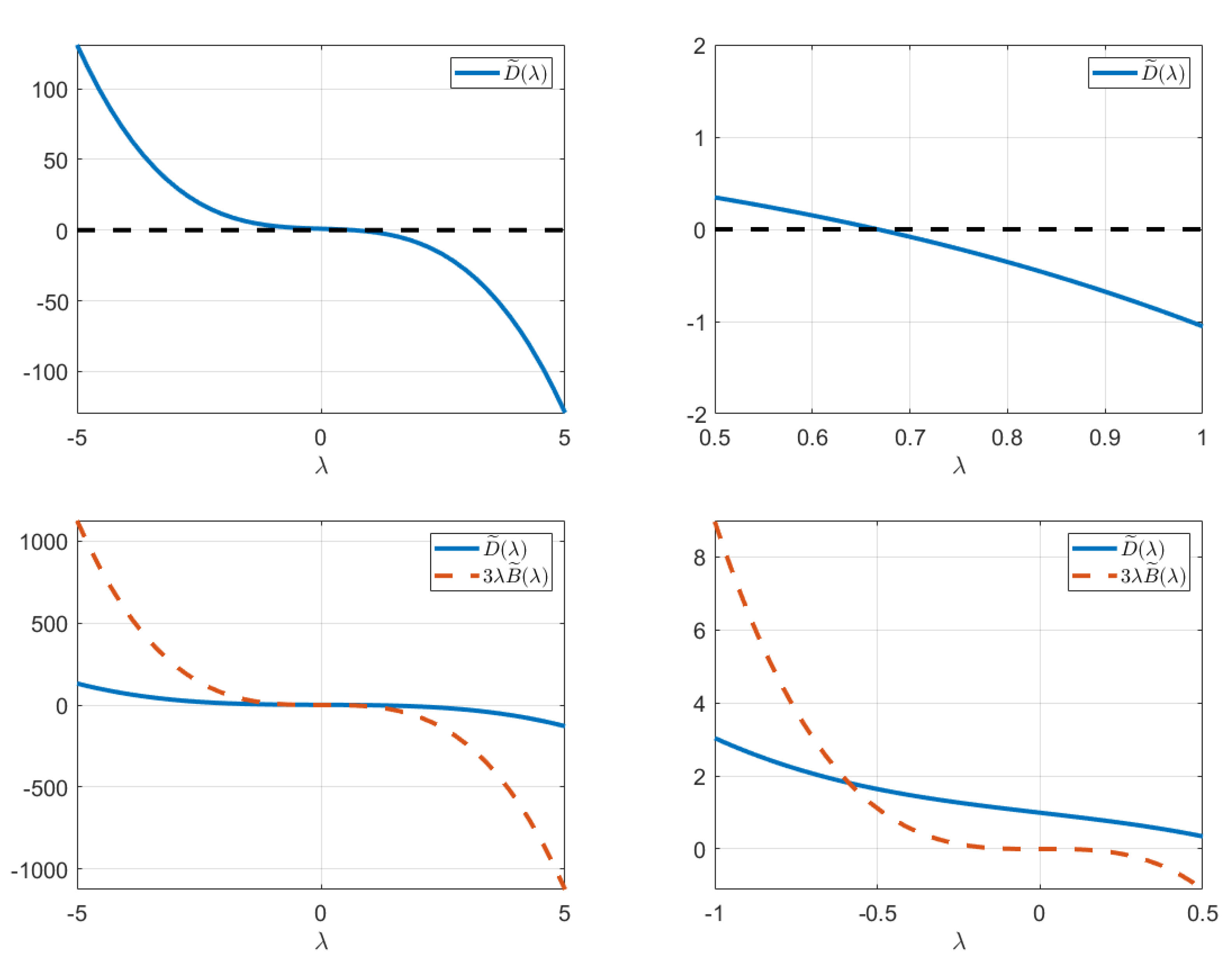
2.3.4. Existence of the Transcritical Bifurcation
2.4. Action on Hares
2.4.1. Equilibria
2.4.2. Local Stability Analysis
2.5. Combined Control over Two Species
2.5.1. Direct and Indirect Control Combination Providing Cottontails’ Extinction and Hares’ Survival
2.5.2. How to Choose the Hares’ Desired State
- 1.
- should be chosen in such a way that . Here, denotes the carrying capacity related to this class, as well as the population level attained by hares in the absence of the other two species, i.e., the value in the two-vanishing populations equilibrium [24]. Consistently with the previous sections, in the simulations we will use a value for the carrying capacity within the range of approximately 26–40 hares/km2 [32], and so hares/km2.
- 2.
2.6. Equilibria
2.7. Local Stability Analysis
3. Results
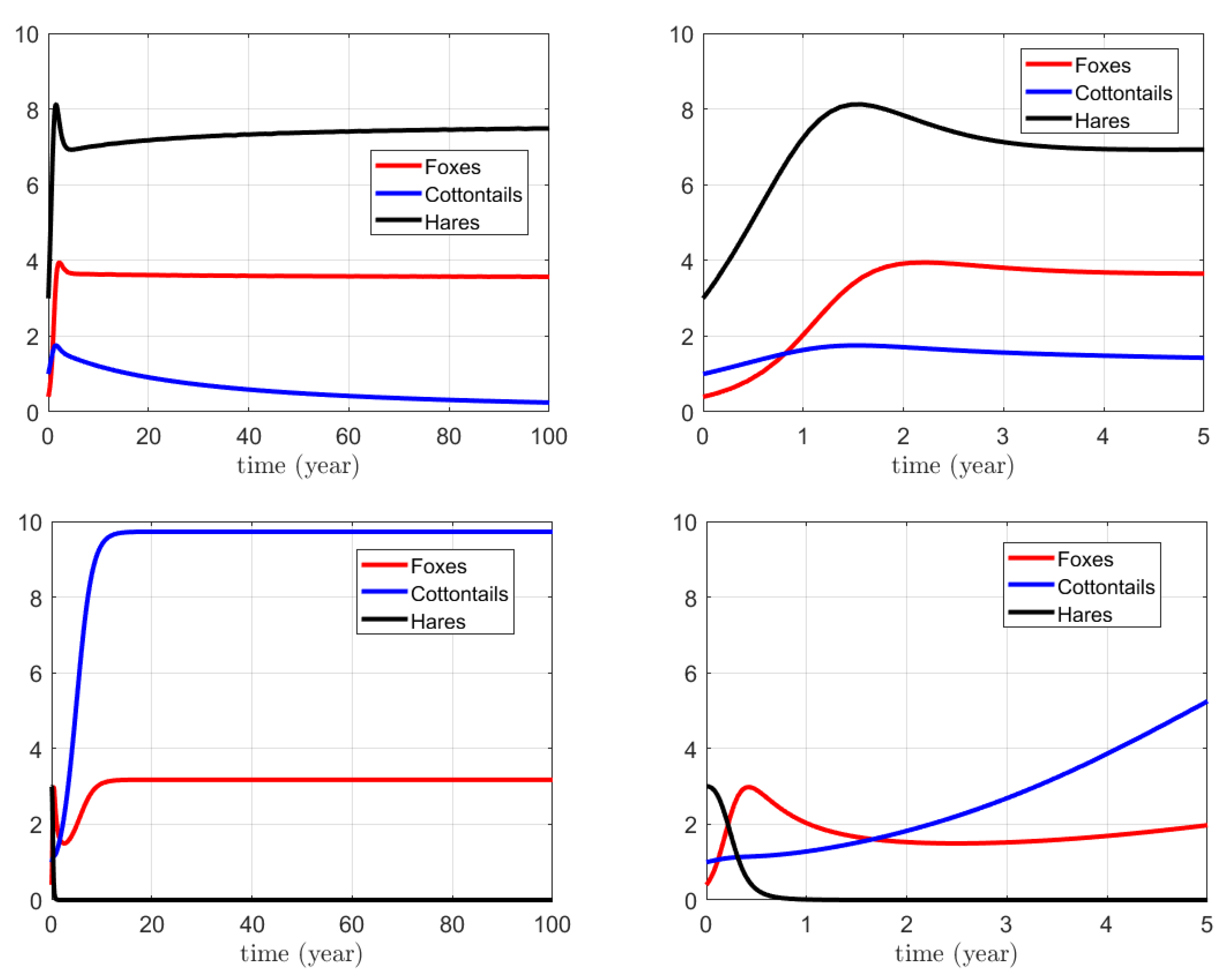
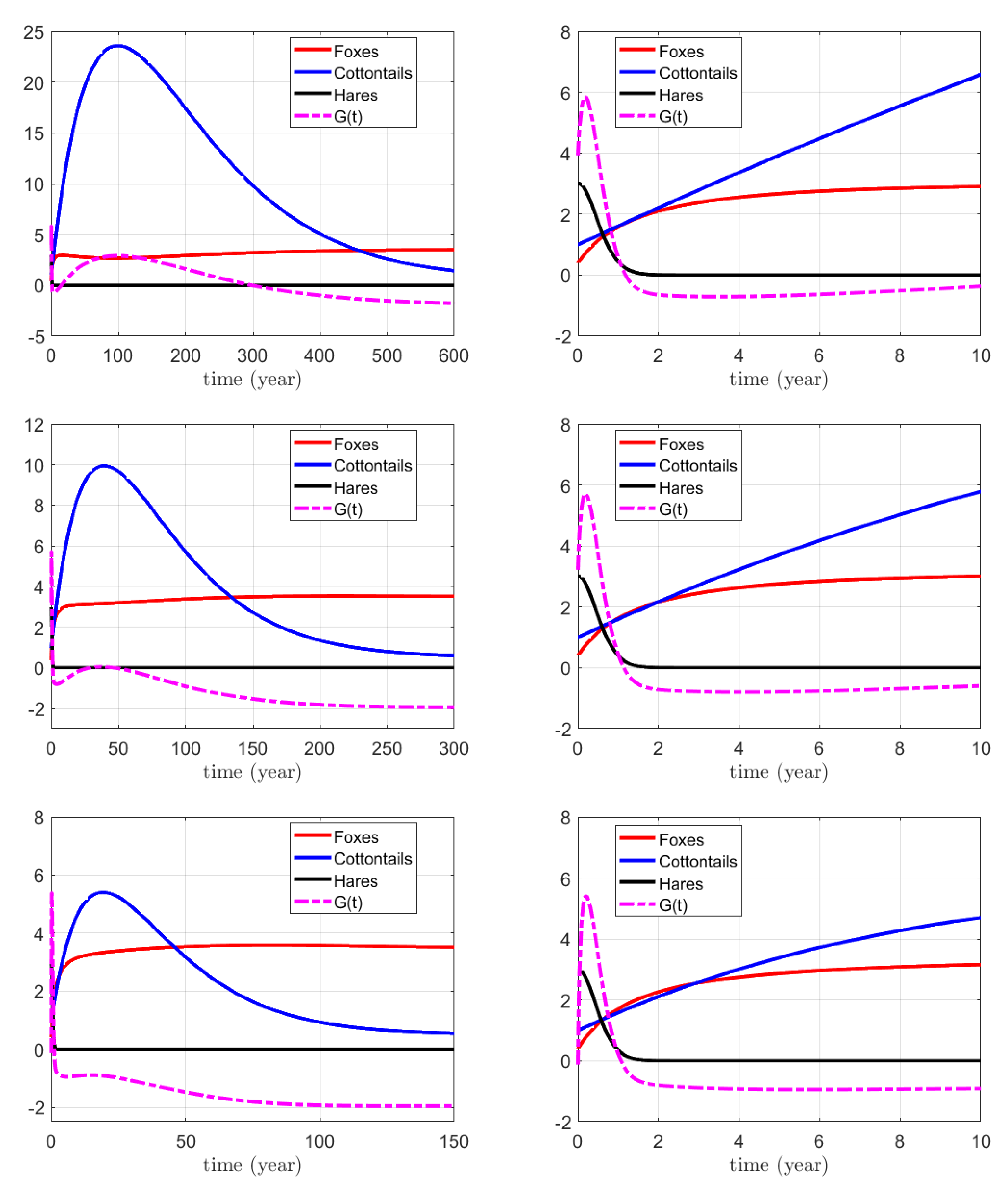
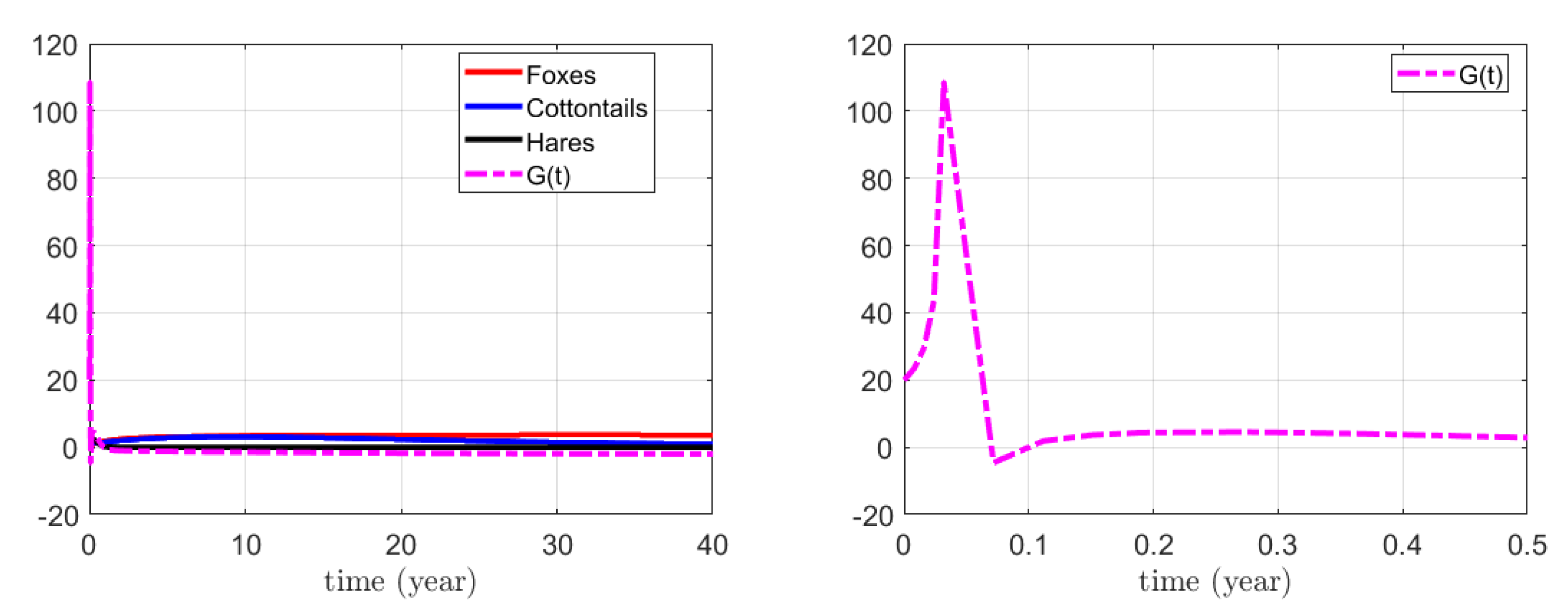
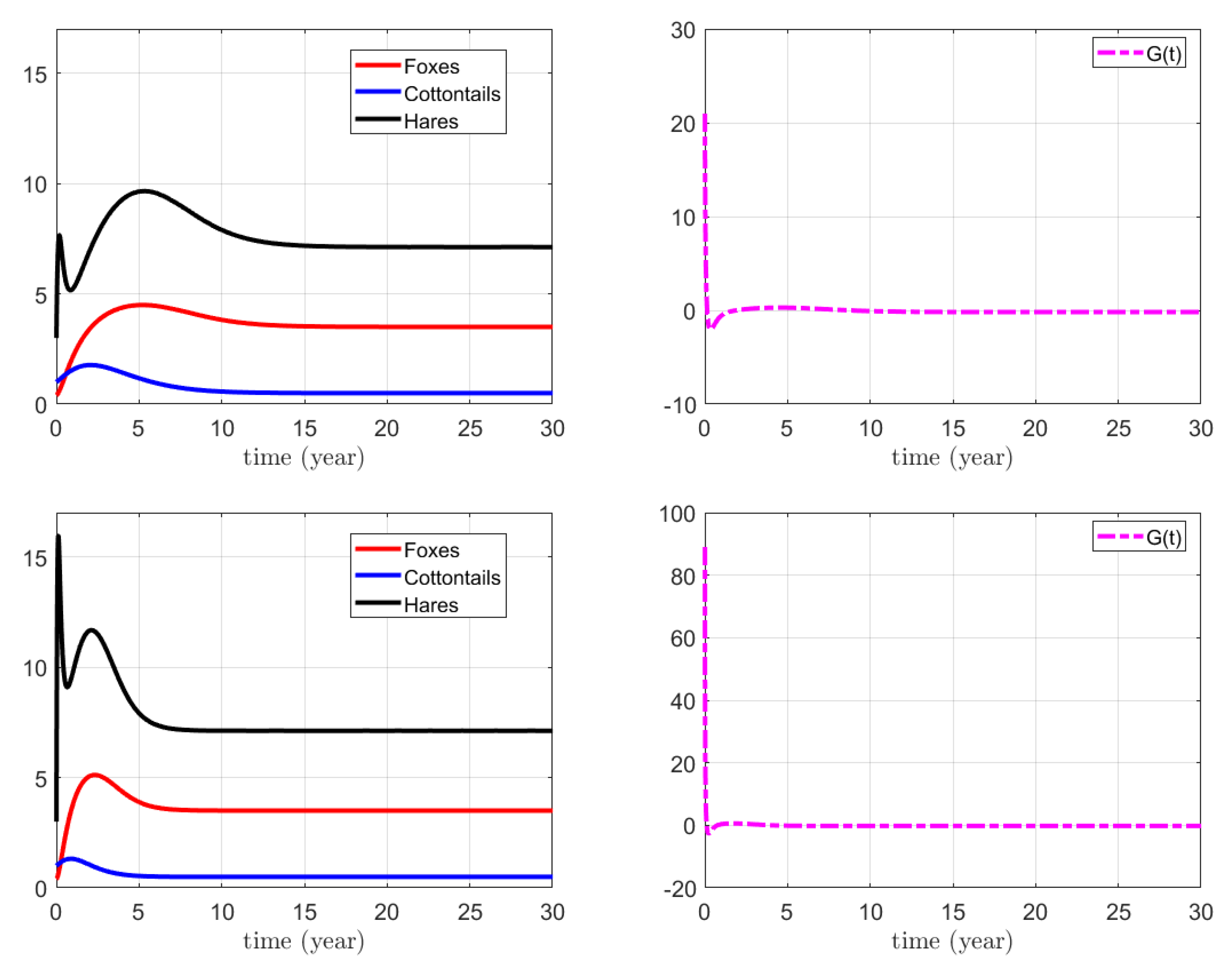
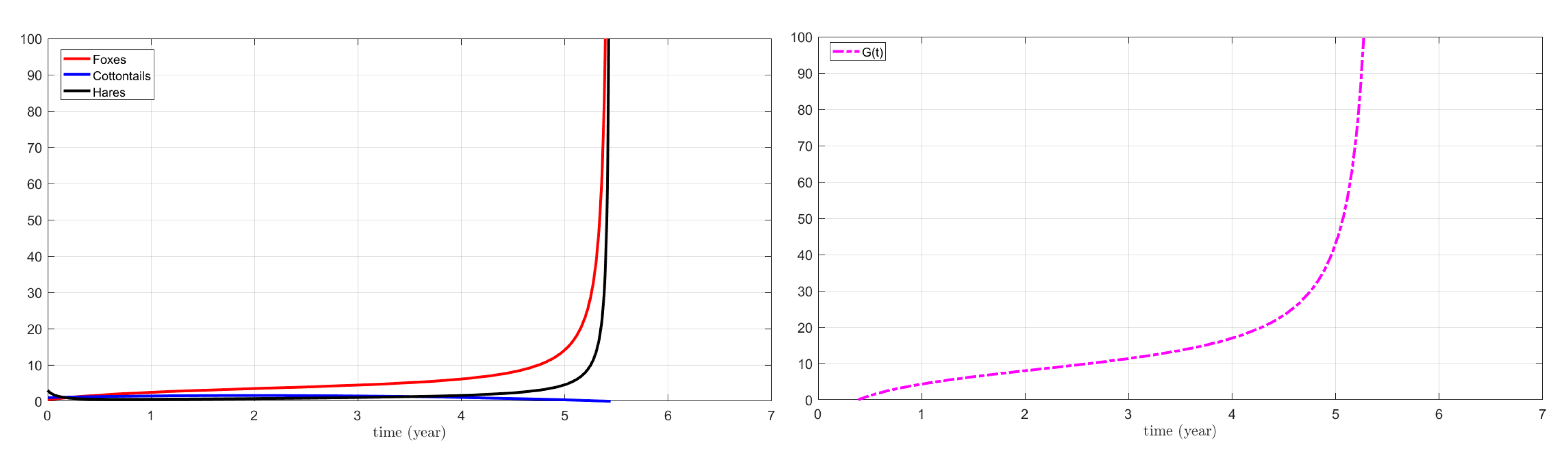
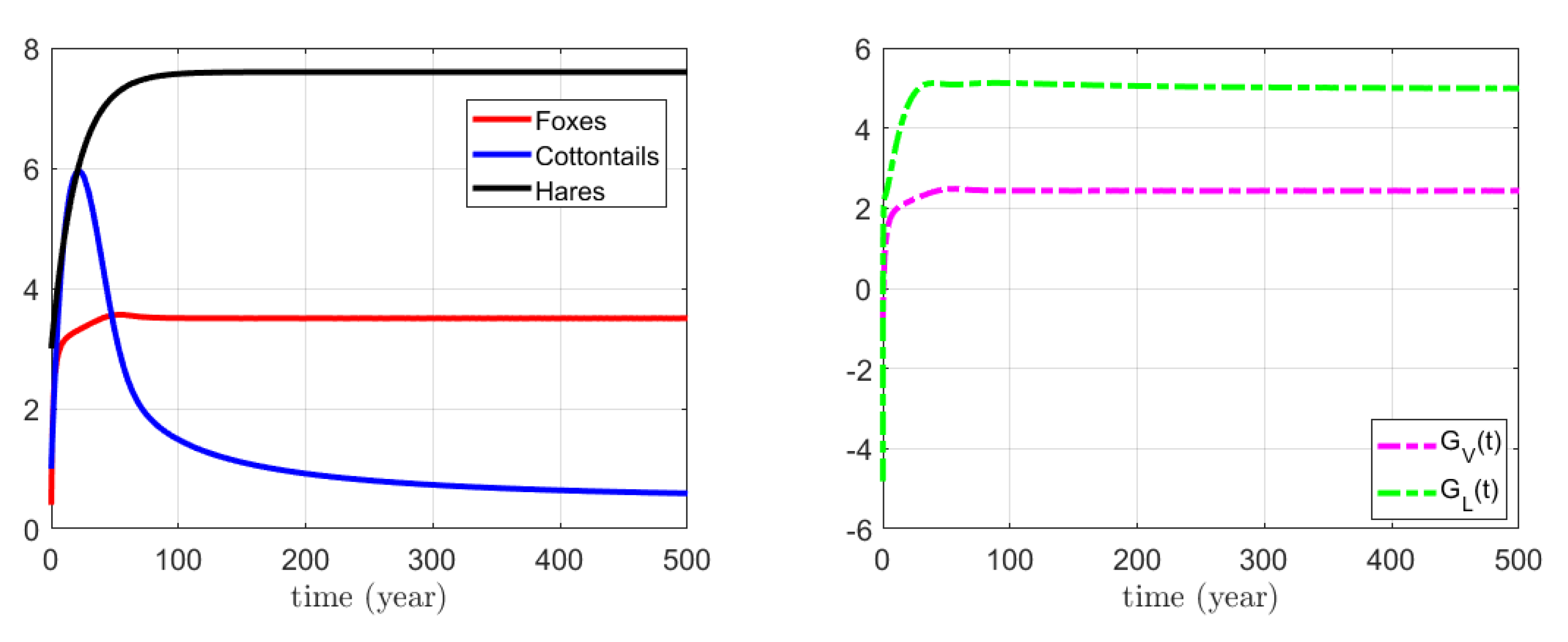
3.1. Numerical Simulations for the Action on Foxes
3.2. Numerical Simulations for the Action on Hares
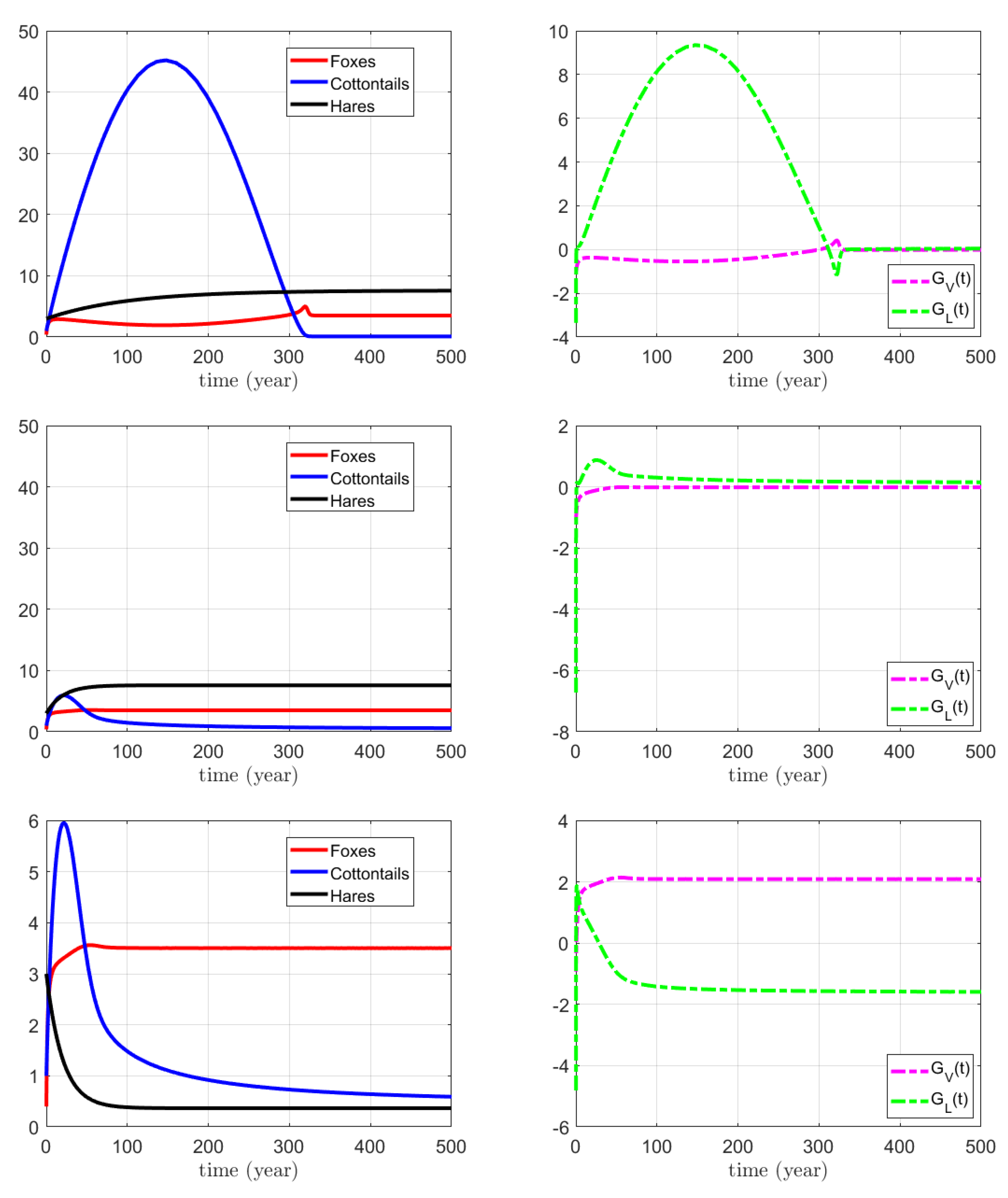
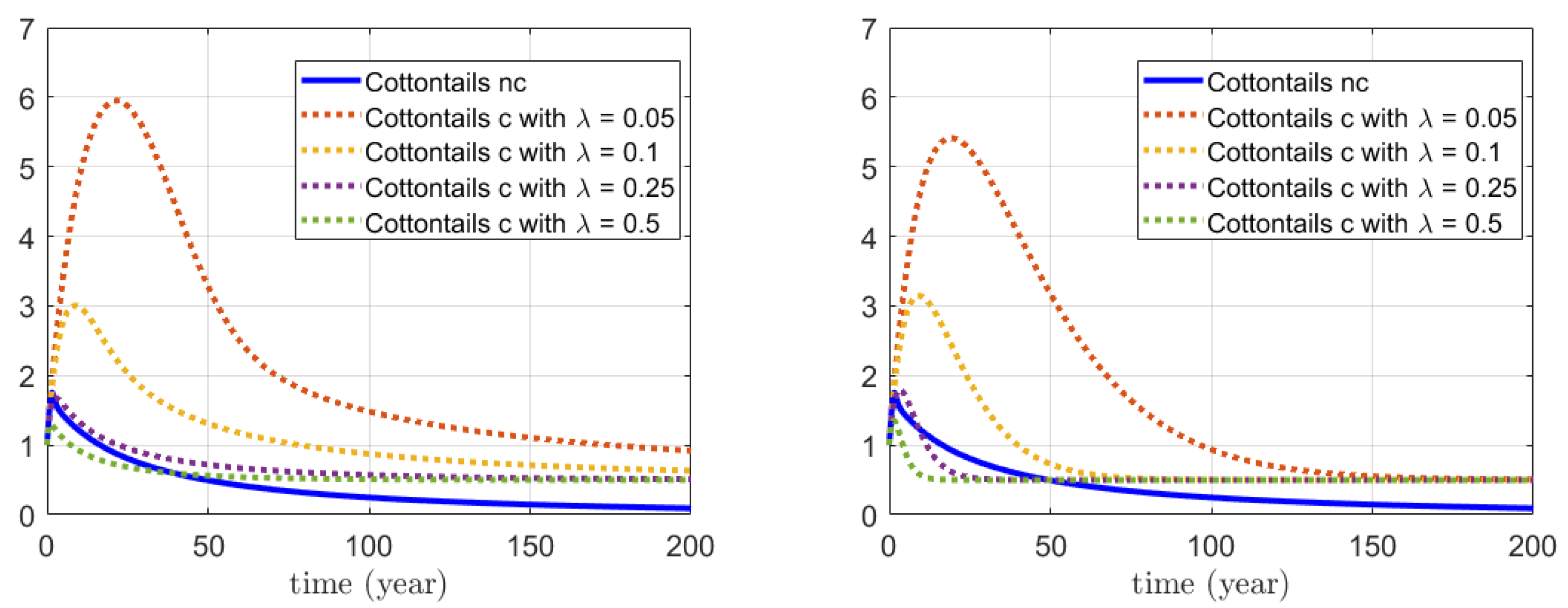
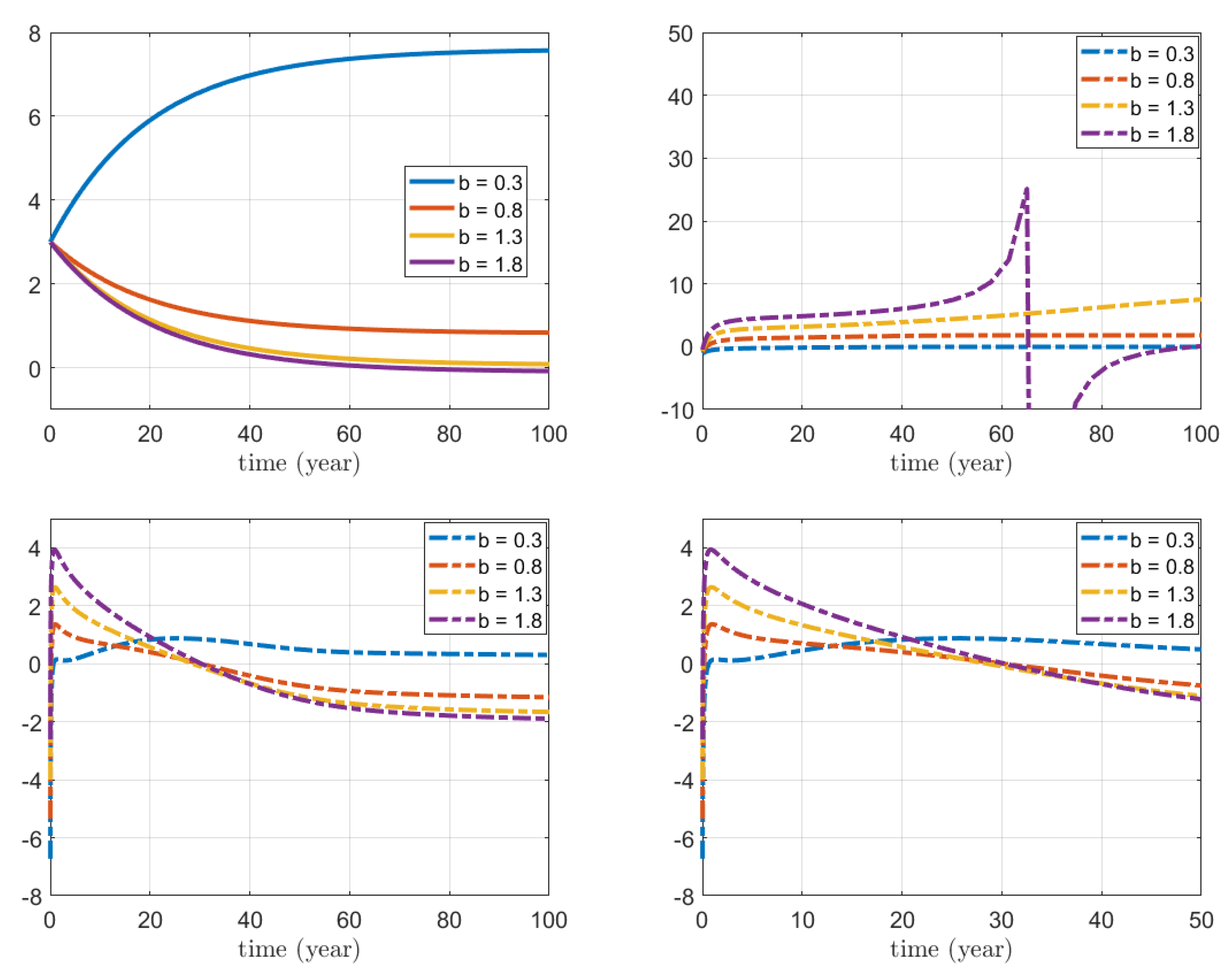
3.3. Numerical Simulations for the Combined Action on Foxes and Hares
4. Discussion
4.1. Biological Interpretation for Indirect Control on Foxes
4.2. Biological Interpretation for Indirect Control on Hares
4.3. Biological Interpretation for Combined Indirect Control on Foxes and Hares
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Bellavere, E.; Venturino, E. Crossing the desert: A model for alien species invasion containment or to lessen habitat disruption effects. J. Biol. Syst. 2023, 31, 557–589. [Google Scholar] [CrossRef]
- Harms, W.B.; Stortelder, A.H.F.; Vos, W. Effects of intensification of agriculture on nature and landscape in the Netherlands. In Land Transformation in Agriculture; Wolman, M.G., Fournier, F.G.A., Eds.; SCOPE Publication 32; John Wiley: New York, NY, USA, 1987; pp. 357–380. [Google Scholar]
- Huffaker, C.B.; Kennett, C.E.; Finney, G.L. Biological control of olive scale. Parlatoria oleae in California by imported Aphytis maculiformis (Hymenoptera; Aphelinidae). Hilgardia 1962, 32, 541–636. [Google Scholar] [CrossRef]
- Isaia, M.; Bona, F.; Badino, G. Influence of landscape diversity and agricultural practices on spiders assemblage in Italian vineyards of Langa Astigiana (Northwest Italy). Environ. Entomol. 2006, 35, 297–307. [Google Scholar] [CrossRef]
- Brown, K.C.; Lawton, J.H.; Shires, S. Effects of insecticides on invertebrate predators and their cereal aphid (Hemiptera: Aphidae) prey: Laboratory experiments. Environ. Entomol. 1983, 12, 1747–1750. [Google Scholar] [CrossRef]
- Nyffeler, M.; Benz, G. Spiders in natural pest control: A review. J. Appl. Entomol. 1987, 103, 321–339. [Google Scholar] [CrossRef]
- Sunderland, K.D.; Samu, F. Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders: A review. Entomol. Exp. Appl. 2000, 95, 113. [Google Scholar] [CrossRef]
- Wise, D.H. Spiders in Ecological Webs; Cambridge University Press: London, UK, 1993. [Google Scholar]
- DeBach, P.; Rosen, D. Biological Control by Natural Enemies; Cambridge University Press: London, UK, 1991. [Google Scholar]
- Flaherty, D.L.; Huffaker, C.B. Biological control of pacific mites and Willamette mites in San Joaquin Valley vineyards, I. Role of Metaseilus occidentalis. Hilgardia 1970, 40, 267–308. [Google Scholar] [CrossRef]
- Marc, P.; Canard, A. Maintaining spider biodiversity in agroecosystems as a tool in pest control. Agric. Ecosyst. Environ. 1997, 62, 229–235. [Google Scholar] [CrossRef]
- Marc, P.; Canard, A.; Ysnel, F. Spiders (Aranee) useful for pest limitation and bioindication. Agric. Ecosyst. Environ. 1999, 74, 229–273. [Google Scholar] [CrossRef]
- Wood, B.J. Development of integrated control programs for pests of tropical perennial crops in Malaysia. In Biological Control; Huffaker, C.B., Ed.; Plenum Press: New York, NY, USA, 1971; pp. 423–457. [Google Scholar]
- Pappalardo, S.; Villa, M.; Santos, S.A.P.; Benhadi-Marín, J.; Pereira, J.A.; Venturino, E. A tritrophic interaction model for an olive tree pest, the olive moth Prays oleae (Bernard). Ecol. Model. 2021, 462, 10977. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Clavero, M.; García-Berthou, E. Invasive species are a leading cause of animal extinctions. Trends Ecol. Evol. 2005, 20, 110. [Google Scholar] [CrossRef] [PubMed]
- Rilov, G. Predator-Prey Interactions of Marine Invaders. In Biological Invasions in Marine Ecosystems: Ecological, Management, and Geographic Perspectives; Springer: Berlin/Heidelberg, Germany, 2009; pp. 261–285. [Google Scholar] [CrossRef]
- Norbury, G. Conserving dryland lizards by reducing predator-mediated apparent com- petition and direct competition with introduced rabbits. J. Appl. Ecol. 2001, 38, 1350–1361. [Google Scholar] [CrossRef]
- Bertolino, S.; Perrone, A.; Gola, L.; Viterbi, R. Population density and habitat use of the introduced Eastern Cottontail (Sylvilagus floridanus) compared to the native European Hare (Lepus europaeus). Zool. Stud. 2011, 50, 315–326. [Google Scholar]
- Bertolino, S.; di Montezemolo, N.C.; Perrone, A. Daytime habitat selection by introduced Eastern Cottontail Sylvilagus floridanus and native European hare Lepus Europaeus in Northern Italy. Zool. Sci. 2011, 28, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Bertolino, S.; di Montezemolo, N.C.; Perrone, A. Habitat use of coexisting introduced eastern cottontail and native European hare. Mamm. Biol. 2013, 78, 235–240. [Google Scholar] [CrossRef]
- Cerri, J.; Ferretti, M.; Bertolino, S. Rabbits killing hares: An invasive mammal modifies native predator-prey dynamics. Anim. Conserv. 2017, 20, 511–519. [Google Scholar] [CrossRef]
- Caudera, E.; Viale, S.; Bertolino, S.; Cerri, J.; Venturino, E. A mathematical model supporting a hyperpredation effect in the apparent competition between invasive Eastern cottontail and native European hare. Bull. Math. Biol. 2021, 83, 51. [Google Scholar] [CrossRef]
- Laticignola, D.; Diele, F. On the Z-type control of backward bifurcations in epidemic models. Math. Biosci. 2019, 315, 108215. [Google Scholar]
- Laticignola, D.; Diele, F. Using awareness to Z-control a SEIR model with overexposure: Insights od Covid-19 pandemic. Chaos Solitons Fractals 2021, 150, 111063. [Google Scholar] [CrossRef]
- Laticignola, D.; Diele, F.; Marangi, C.; Provenzale, A. On the dynamics of a generalized predator-prey system with Z-type control. Math. Biosci. 2016, 280, 10–23. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, X.; Liao, B.; Zhang, Y.; Ding, Y. Z-type control of populations for Lotka-Volterra model with exponencial convergence. Math. Biosci. 2016, 272, 15–23. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Y. Design and experimentation of acceleration-level drift-free scheme aided by two recurrent neural networks. IET Control Theory Appl. 2013, 7, 25–42. [Google Scholar] [CrossRef]
- Perko, L. Differential Equations and Dynamical Systems; Springer: New York, NY, USA, 2001. [Google Scholar]
- La Morgia, V.; Venturino, E. Understanding hybridization and competition processes between hare species: Implications for conservation and management on the basis of a mathematical model. Ecol. Model. 2017, 364, 13–24. [Google Scholar] [CrossRef]
- Pandini, W.; Tosi, G.; Meriggi, A.; Lepre, L.; Bianca, C.; Selvatico, S.; Simonetta, A.M.; Dessì-Fulgheri, F. Principi e tecniche di gestione faunistico-venatoria. Greentime 1998, 195–220. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camattari, F.; Acotto, F.; Venturino, E. Z-Type Control Methods on a Three-Species Model with an Invasive Prey. Mathematics 2023, 11, 4182. https://doi.org/10.3390/math11194182
Camattari F, Acotto F, Venturino E. Z-Type Control Methods on a Three-Species Model with an Invasive Prey. Mathematics. 2023; 11(19):4182. https://doi.org/10.3390/math11194182
Chicago/Turabian StyleCamattari, Fabiana, Francesca Acotto, and Ezio Venturino. 2023. "Z-Type Control Methods on a Three-Species Model with an Invasive Prey" Mathematics 11, no. 19: 4182. https://doi.org/10.3390/math11194182
APA StyleCamattari, F., Acotto, F., & Venturino, E. (2023). Z-Type Control Methods on a Three-Species Model with an Invasive Prey. Mathematics, 11(19), 4182. https://doi.org/10.3390/math11194182