
Dynamics of a Predator–Prey Model with the Additive Predation in Prey


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- (i)
- If the ratio exceeds the largest size that the prey u may eventually achieve, then the predator species goes extinct.
- (ii)
- When model (2) has the weak Allee effect or no Allee effect, if the ratio is below the largest size that the prey u may eventually achieve, both the predator and prey species may coexist. In the weak Allee effect case, the coexistence exhibits an oscillatory mode (when there exists a stable limit cycle) or steady state mode (when the interior equilibrium is globally asymptotically stable), while in the no Allee effect case, it is only the steady state mode. In the strong Allee effect, the extinction and coexistence depend on the initial population sizes of the prey and predator.
- (iii)
- For the strong Allee effect case, model (2) exhibits a more complex bifurcation phenomenon than the other cases, such as the Hopf and heteroclinic bifurcations. Due to the strong Allee effect caused by the additive predation term , the initial population sizes play an extremely important role in the dynamics of (2). For a set of parameter values, the extinction, coexistence, and population oscillations may be the result of different initial conditions.
- (iv)
- The main dynamics of model (2) can be described conclusively in a bifurcation diagram with respect to .
- (v)
- Compared to the well-known Lotka–Volterra type model (when ), the additive predation term leads model (2) to have much richer and more complex dynamical behaviors, such as the Allee effect, oscillatory behavior, a more complex bifurcation phenomenon and sensitivity to the initial conditions. The Lotka–Volterra type model only has the similar dynamic structure of model (2) in the case of no Allee effect.
2. Dynamics of the Single Population Model
- (i)
- (ii)
- If , i.e., , then the species u and hence v will be extinct since for all .
- (a)
- Assume .
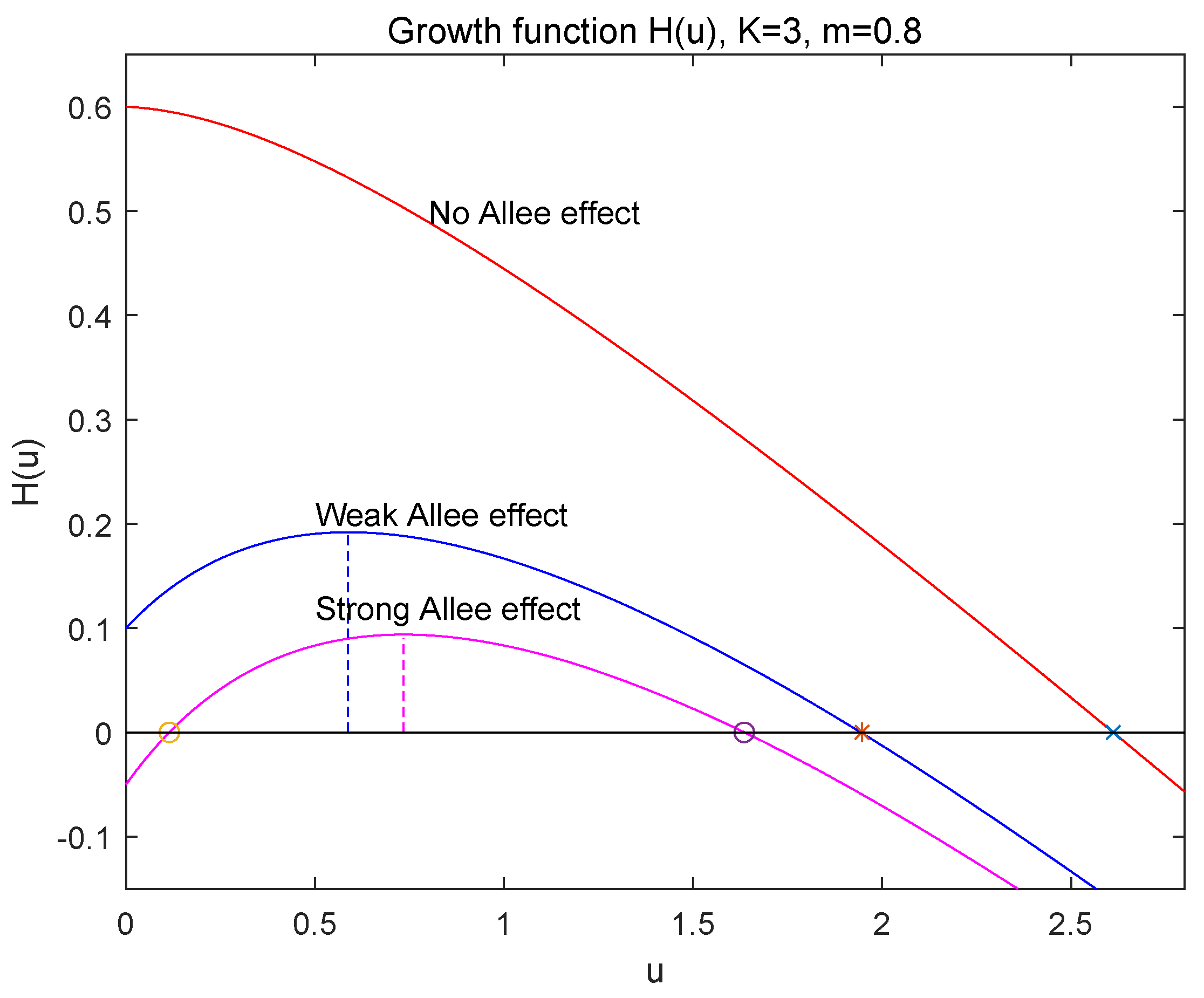
- If , which implies that , then , has two positive roots and , which are given by (8) and satisfy that and since (see Figure 1). Thus, is locally asymptotically stable, is unstable and is locally asymptotically stable. So, in this case, the strong Allee effect appears in population growth of (1) and is the Allee threshold. The species u will be extinct when the initial population size is below , i.e., , and persistent when .
- (b)
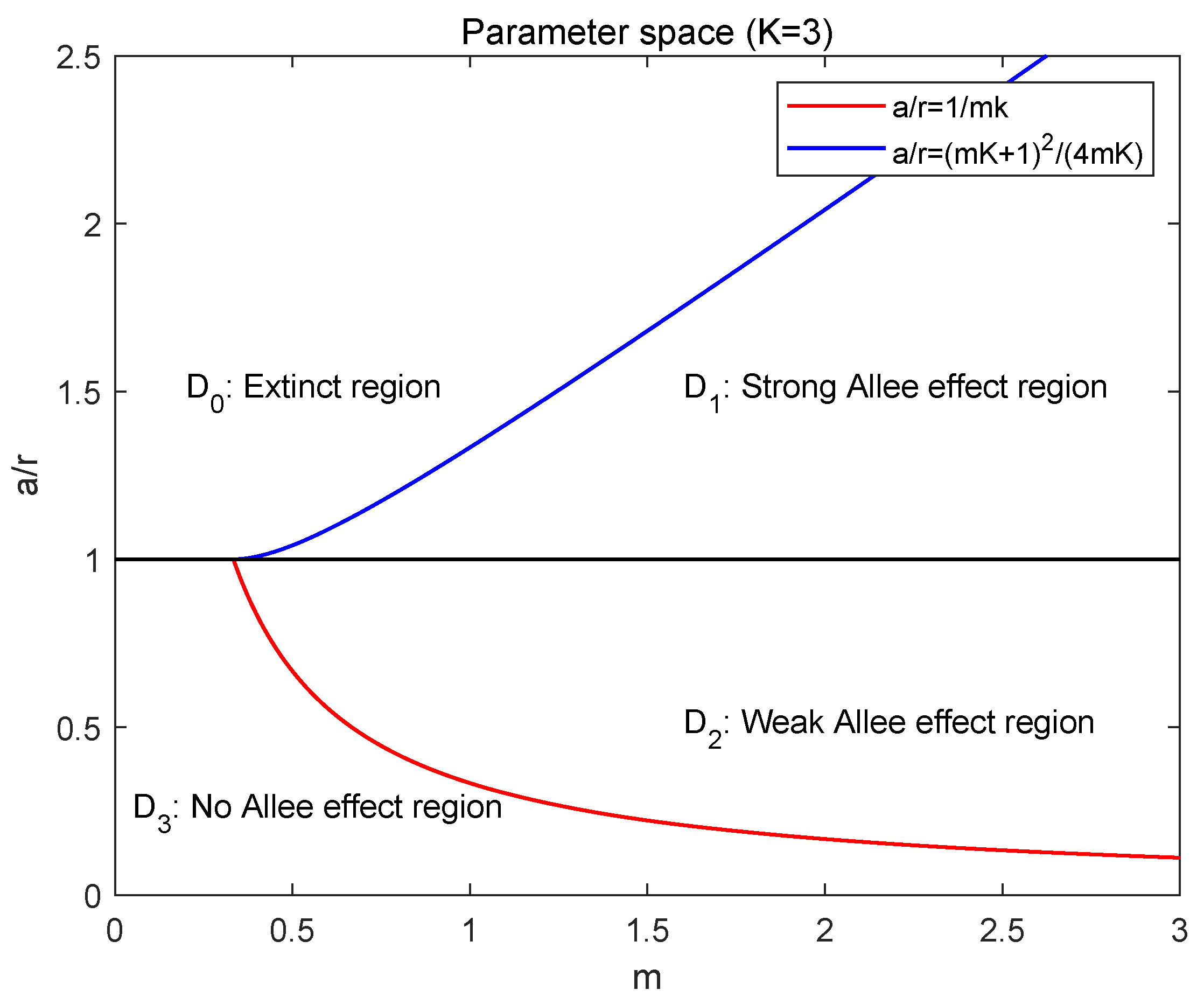
- Extinct region: ,
- Strong Allee effect region: ,
- Weak Allee effect region: ,
- No Allee effect region: .
3. Dynamics of the Predator–Prey Model
3.1. Positivity and Boundedness
3.2. Equilibria
3.3. Dynamics of the Strong Allee Effect Case
- 1.
- 2.
- is a stable node.
- 3.
- If , is a saddle with its unstable manifold along the u axis and its stable manifold (denoted by ) entering from the interior of ; if , is an unstable node.
- 4.
- If , is saddle with its stable manifold along the u axis and its unstable manifold (denoted by ) entering the interior of ; if , is a stable node.
- 5.
- The positive equilibrium is locally asymptotically stable when and unstable when . When , model (2) undergoes the Hopf bifurcation at .
- i.
- The first part is about the extinction of predator species due to the large ratio of the death rate d to the conversion rate β of predator species.
- ii.
- The second part is about the extinction due to the strong Allee effect of the prey population driven by the additive predation . implies that the invasion or reproduction of the predator v is excessive while the reproduction of the prey u is not fast enough to sustain its own population. Thus, the excessive invasion or reproduction of the predator v drives the population of the prey u to below its Allee threshold and eventually to zero, which consequently drives the predator population to extinction.
- iii.
- The third statement indicates that when the population density of the prey is below its Allee threshold , all species will be extinct.
- (i)
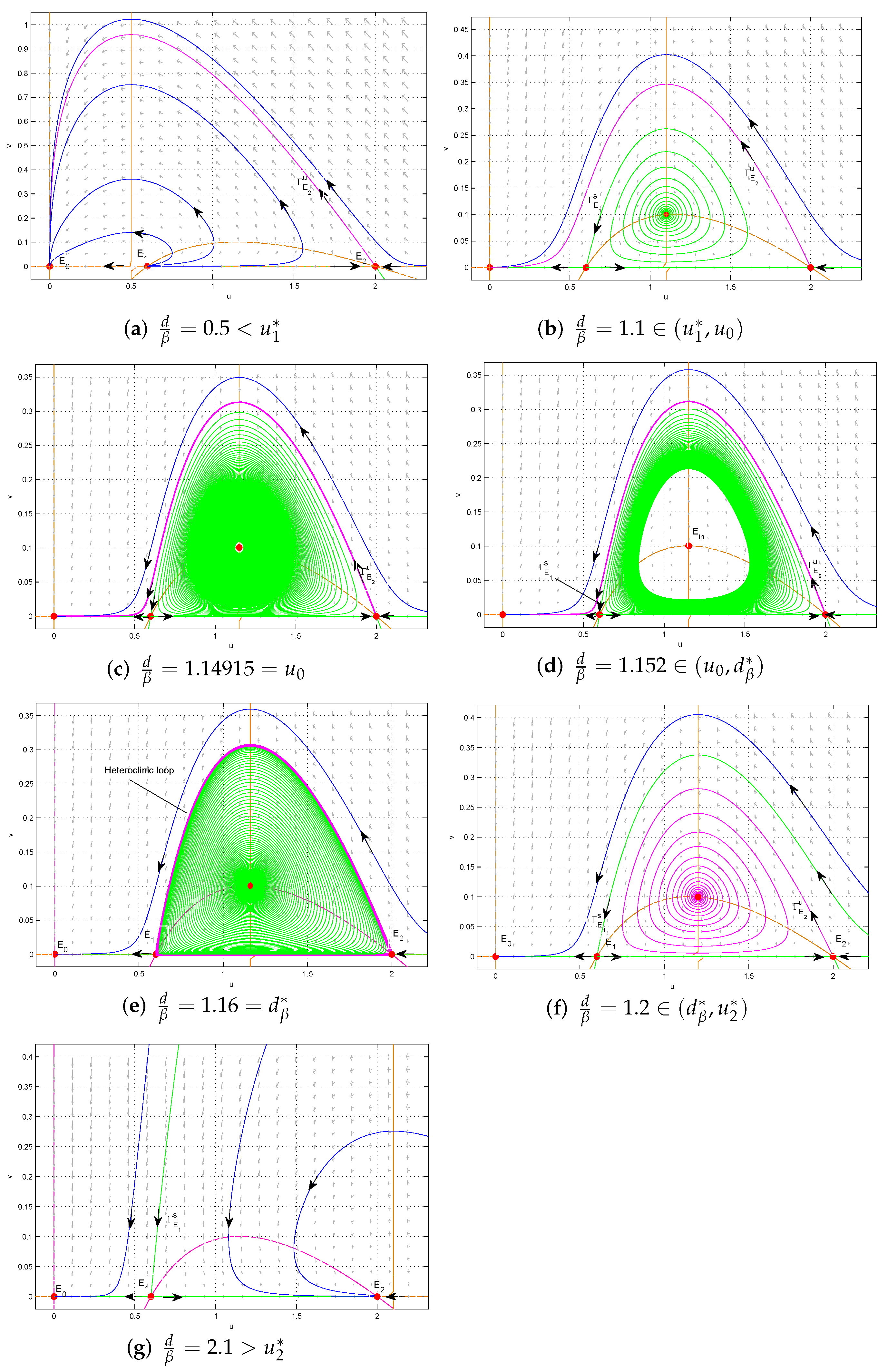
- . is a stable node, is an unstable node, is a saddle. There is no interior equilibrium. All trajectories in the interior of converge to ; two species will be extinct (see Figure 3a).
- (ii)
- . becomes a saddle and an unstable interior equilibrium appears. Around , there is no limit cycle. Except for the stable manifold of , all trajectories converge to (see Figure 3b).
- (iii)
- . A forward subcritical Hopf bifurcation occurs (see Figure 3c).
- (iv)
- . Both and are saddle and the interior equilibrium become locally asymptotically stable. There exists a threshold value such that
- (a)
- when , there exists an unstable limit cycle surrounding . Outside the limit cycle, two species cannot coexist, while inside the limit cycle trajectories converge towards the stable interior equilibrium (see Figure 3d);
- (b)
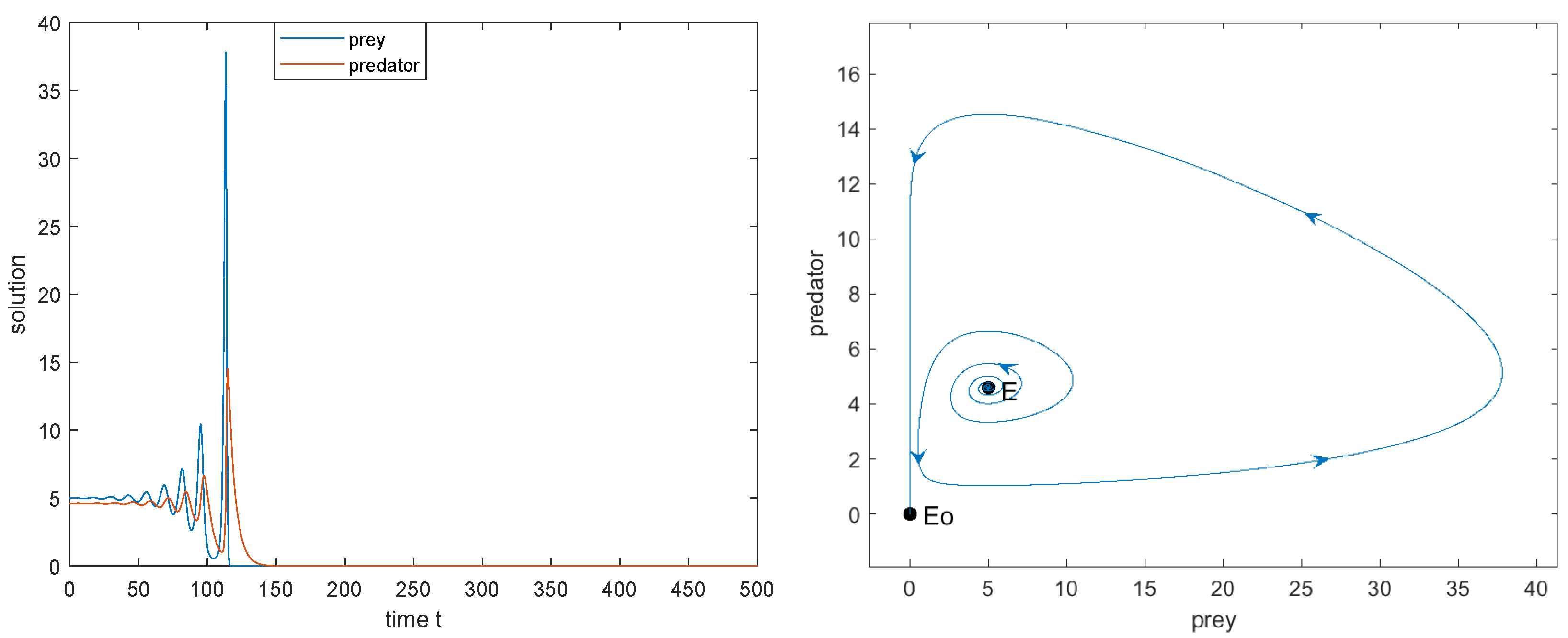
- when , the limit cycle disappears and a heteroclinic loop connecting to appears. Outside the heteroclinic loop, trajectories converge to , and two species will be extinct, while inside the heteroclinic loop trajectories converge towards the heteroclinic loop, and two species coexist (see Figure 3e);
- (c)
- when , the heteroclinic orbit is broken. Above the stable manifold of , trajectories converge to , and two species will be extinct, while below , trajectories converge towards the stable interior equilibrium , and two species coexist (see Figure 3f).
- (v)
- . There is a transcritical bifurcation at . When increasing the value of from , becomes a stable node and the interior equilibrium disappears. This leads to the predator free dynamics with as attractors (see Figure 3g).
3.4. Dynamics of the Weak Allee Effect Case
- 1.
- 2.
- is always an unstable saddle.
- 3.
- is a stable node when and an unstable saddle when .
- 4.
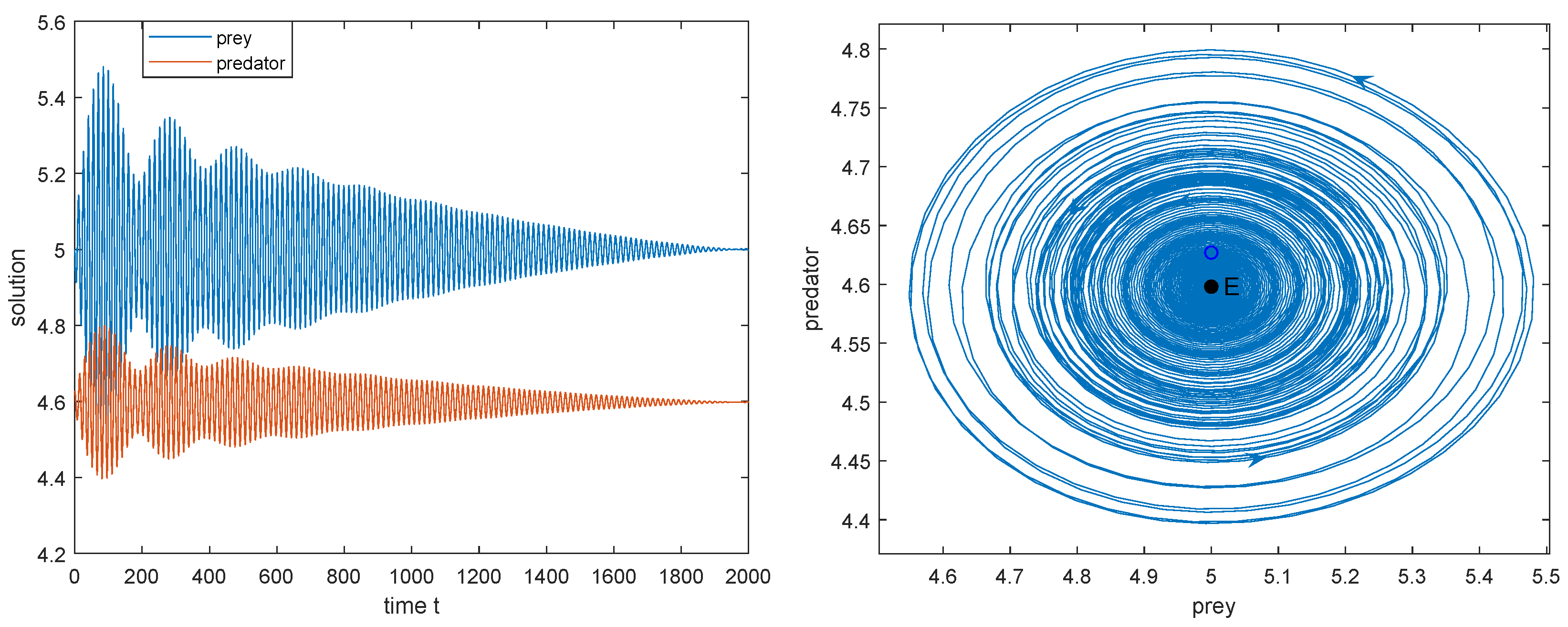
- The positive equilibrium is locally asymptotically stable when and unstable when . When , system (2) undergoes the Hopf bifurcation at .
3.5. Dynamics of No Allee Effect Case
- 1.
- 2.
- is always an unstable saddle.
- 3.
- is a stable node when and an unstable saddle when .
- 4.
- If the positive equilibrium exists, it is locally asymptotically stable.
- 1.
- If , then is globally asymptotically stable.
- 2.
- If , then is globally asymptotically stable.
- i.
- If the ratio is large such that , then the predator species u will be extinct.
- ii.
- If , both species u and v can coexist.
3.6. Summary of the Dynamics of Model
- (1)
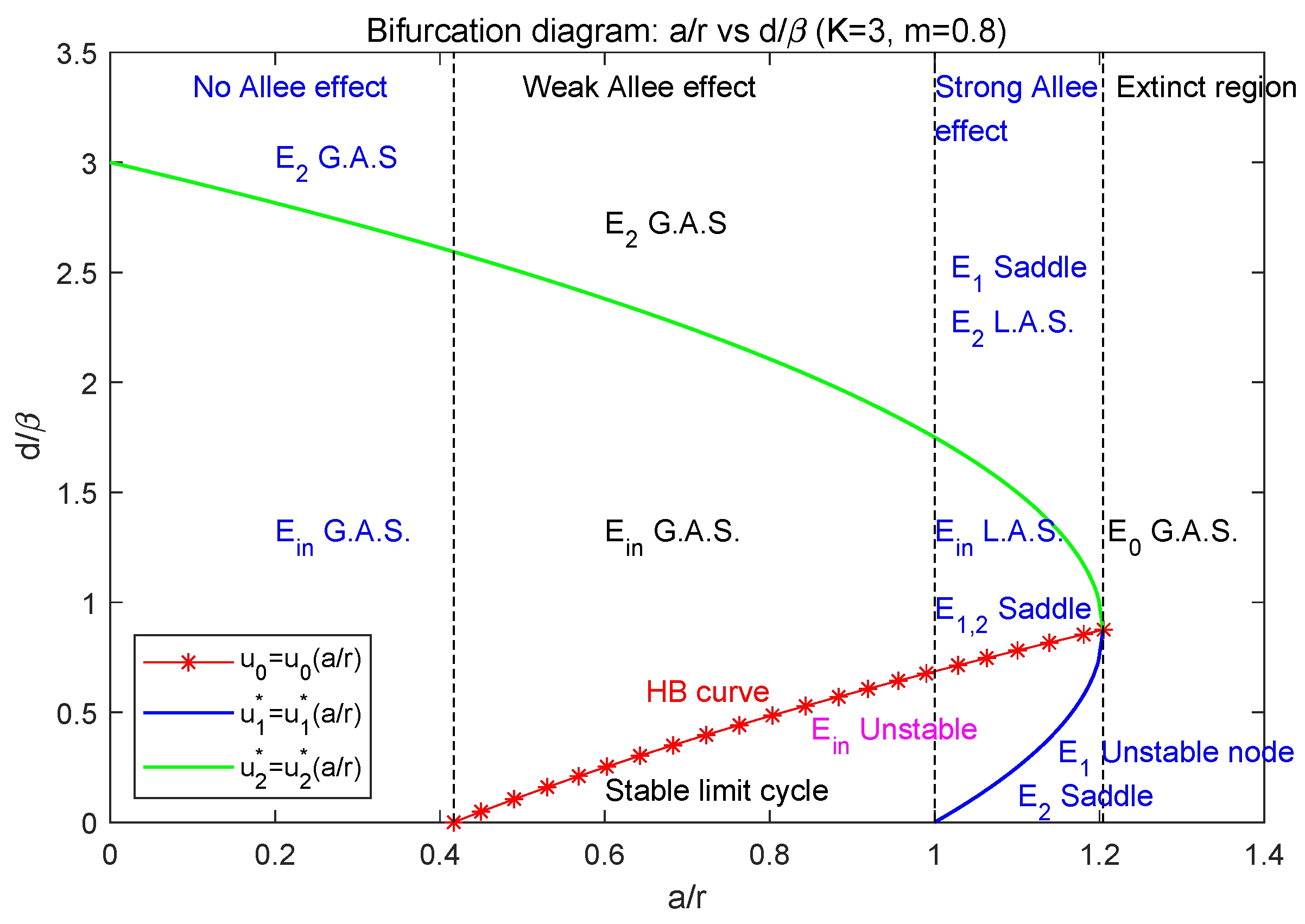
- When , the parameter lies in the extinct region, i.e., ; model (2) only has the extinction equilibrium which is globally asymptotically stable. In this case, both the predator and prey species will be extinct.
- (2)
- When , model (2) admits the strong Allee effect. In this case, model (2) has three boundary equilibria , where is a stable node (see Section 3.3).
- (2.1)
- (2.2)
- Below the blue curve (i.e., ), is a saddle, is an unstable node, and is globally asymptotically stable. So, two species will be extinct (see Figure 3a).
- (2.3)
- Between the blue and green curves (i.e., ), the interior equilibrium exists, and both and are saddle. Between the red and green curve (i.e., ), is locally asymptotically stable; between the blue and red curve (i.e., ), it is unstable; along the red curve (i.e., ), Hopf bifurcation occurs. The two species may coexist but it depends on their initial population sizes. In this case, model (2) may have the heteroclinic loop connecting and (see Figure 3b–f).
- (3)
- When , model (2) has weak Allee effect. In this case, model (2) has two boundary equilibria , where is a saddle (see Section 3.4).
- (3.1)
- (3.2)
- Below the green curve (i.e., ), the interior equilibrium exists, is a saddle. Between the red and green curve (i.e., ), is globally asymptotically stable; below the red curve (i.e., ), model (2) has a stable limit cycle surrounding ; along the red curve (i.e., ), Hopf bifurcation occurs. So, in this case, two species can coexist (see Figure 4a–c).
- (4)
- When , model (2) has no Allee effect. In this case, model (2) has two boundary equilibria , where is a saddle (see Section 3.5).
- (4.1)
- Above the green curve (i.e., ), model (2) has no positive equilibrium; is globally asymptotically stable; that is, the predator will be extinct while the prey is permanent.
- (4.2)
- Below the blue curve (i.e., ), the interior equilibrium exists and is globally asymptotically stable. So, in this case, two species can coexist.
4. The Influence of Delay
4.1. Stability and Hopf Bifurcation Induced by Delay
- 1.
- For all , is locally asymptotically stable if , and unstable if .
- 2.
- If exists, it is unstable for all .
- 3.
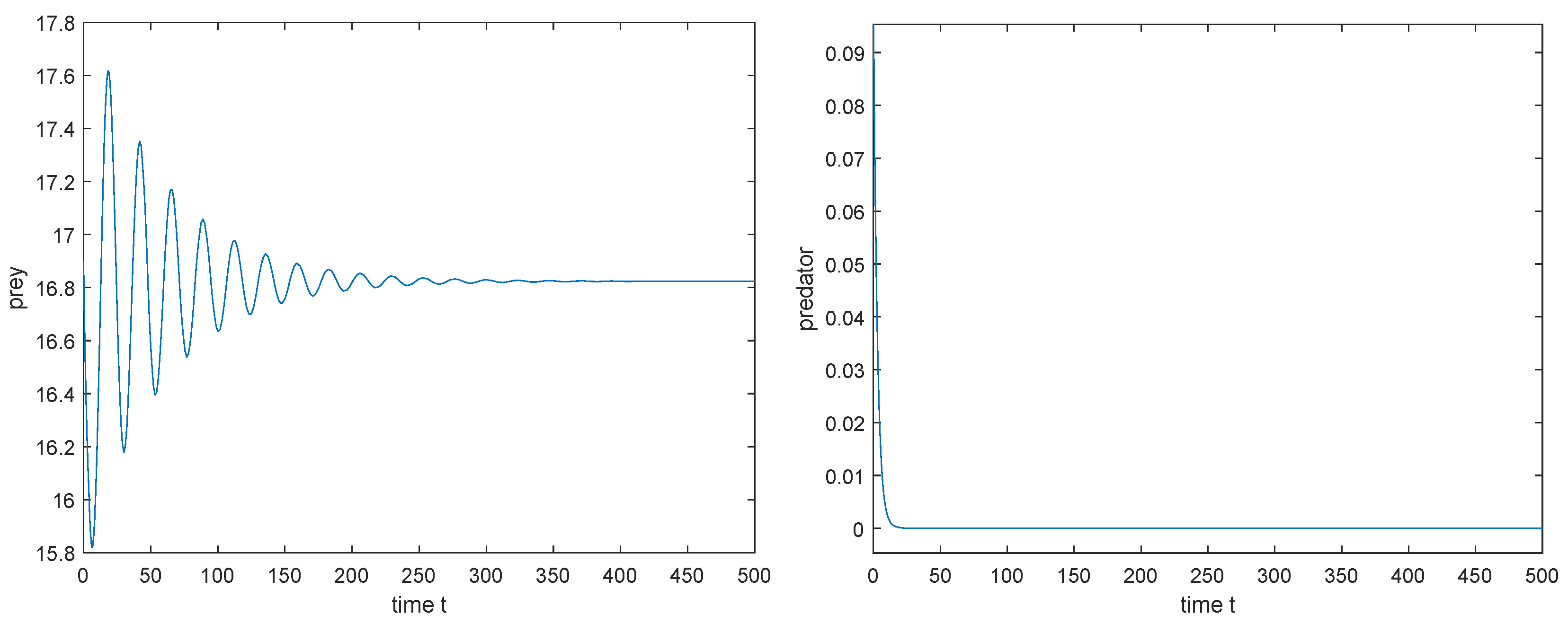
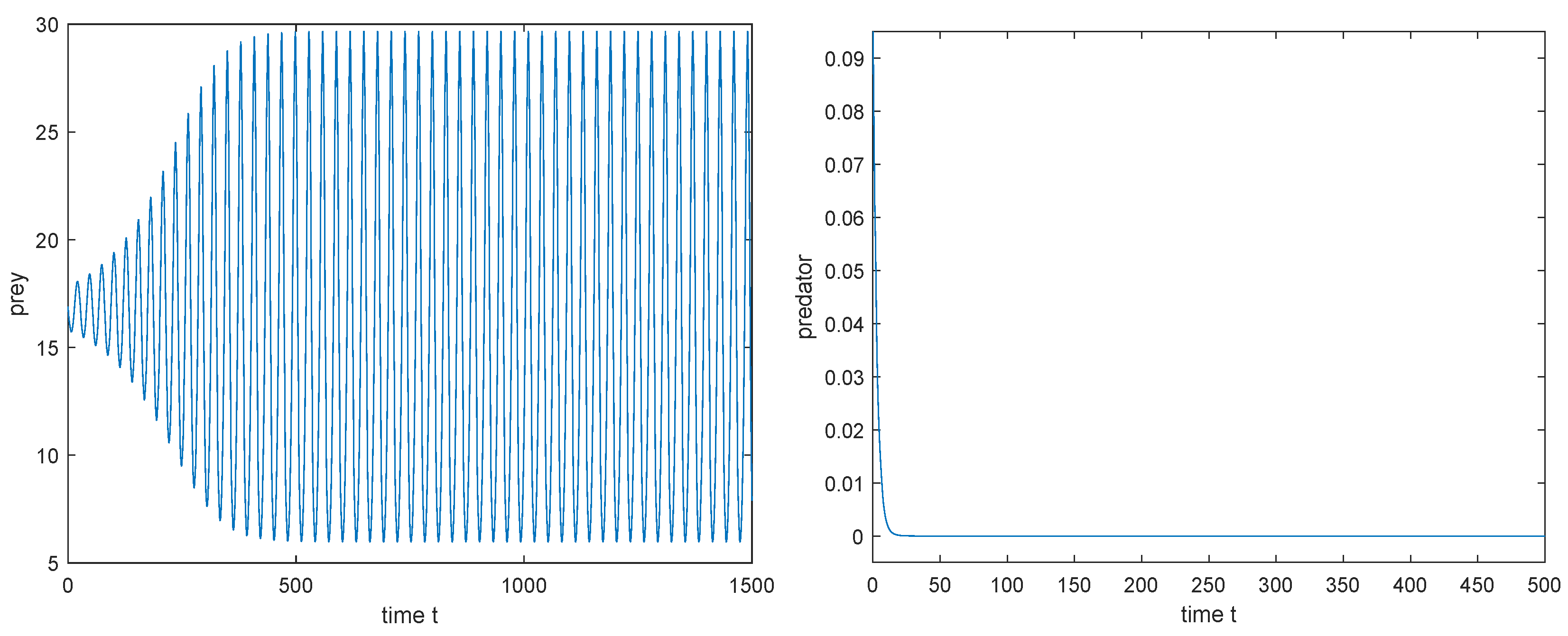
- If exists, then (i) when , there exists a such that is locally asymptotically stable for , unstable for , and undergoes Hopf bifurcation at ; (ii) when , is unstable for all .
4.2. Direction and the Stability of Hopf-Bifurcating Periodic Solutions
- (1)
- If (resp. ), then the Hopf bifurcation is supercritical (resp. subcritical).
- (2)
- If (resp. ), then the periodic solution is orbitally asymptotically stable (resp. unstable).
- (3)
- If (resp. ), then the period of bifurcating periodic solutions increases (resp. decreases) as the bifurcation parameter (delay time ) keeps away from the critical value.
4.2.1. At the Positive Equilibrium
4.2.2. At the Semi-Trivial Equilibrium
5. Conclusions
- (i)
- Due to the additive predation term , model (2) may have the Allee effect;
- (ii)
- The dynamics of the Lotka–Volterra type model (40) only have the similar dynamic structure of (2) in the case of no Allee effect (see Section 3.4), but have no dynamics of the strong and weak Allee effect cases;
- (iii)
- Model (2) may have oscillatory behavior;
- (iv)
- The strong Allee effect increases the extinction risk of the prey and predator species. The initial populations of the prey and predator play an important role in the persistence of (2). Not only the low initial prey density, but also the high initial predator density which causes the over-exploitation of the prey, will lead to the extinction of all species. For a set of parameter values, both community extinction, coexistence, and population oscillations may be the result of different initial conditions;
- (v)
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kang, Y.; Udiani, O. Dynamics of a single species evolutionary model with Allee effects. J. Math. Anal. Appl. 2014, 418, 492–515. [Google Scholar] [CrossRef]
- Dennis, B. Allee effect: Population growth, critical density, and chance of extinction. Nat. Resour. Model. 1989, 3, 481–538. [Google Scholar] [CrossRef]
- Dercole, F.; Ferriére, R.; Rinaldi, S. Ecological bistability and evolutionary reversals under asymmetrical competition. Evolution 2002, 56, 1081–1090. [Google Scholar] [CrossRef][Green Version]
- Allee, W.C. Animal Aggregations: A Study in General Sociology; University of Chicago Press: Chicago, IL, USA, 1931. [Google Scholar]
- Allee, W.C. Studies in animal aggregations: Mass protection against colloidal silver among goldfishes. J. Exp. Zool. 1932, 61, 185–207. [Google Scholar] [CrossRef]
- Kramer, A.M.; Dennis, B.; Liebhold, A.M.; Drake, J.M. The evidence for Allee effects. Popul. Ecol. 2009, 51, 341–354. [Google Scholar] [CrossRef]
- Berec, L.; Angulo, E.; Courchamp, F. Multiple Allee effects and population management. Trends Ecol. Evol. 2006, 22, 185–191. [Google Scholar] [CrossRef]
- Courchamp, F.; Berec, L.; Gascoigne, J. Allee Effects in Ecology and Conservation; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Mooring, M.S.; Fitzpatrick, T.A.; Nishihira, T.T.; Reisig, D.D. Vigilance, predation risk, and the Allee effect in desert bighorn sheep. J. Wildl. Manag. 2004, 68, 519–532. [Google Scholar] [CrossRef]
- Rinella, D.J.; Wipfli, M.S.; Stricker, C.A.; Heintz, R.A.; Rinella, M.J. Pacific Salmon (Oncorhynchus sp.) runs and consumer fitness: Growth and energy storage in stream-dwelling salmonids increase with salmon spawner density. Can. J. Fish. Aquat. Sci. 2012, 69, 73–84. [Google Scholar] [CrossRef]
- Stephens, P.A.; Sutherland, W.J. Consequences of the Allee effect for behaviour, ecology and conservation. Trends Ecol. Evol. 1999, 14, 401–405. [Google Scholar] [CrossRef]
- Stephens, P.A.; Sutherland, W.J.; Freckleton, R.P. What is the Allee effect? Oikos 1999, 87, 185–190. [Google Scholar] [CrossRef]
- Wang, M.E.; Kot, M. Speeds of invasion in a model with strong or weak Allee effects. Math. Biosci. 2001, 171, 83–97. [Google Scholar] [CrossRef]
- Courchamp, F.; Clutton-Brock, T.; Grenfell, B. Inverse density dependence and the Allee effect. Trends Ecol. Evol. 1999, 14, 405–410. [Google Scholar] [CrossRef]
- Conway, E.D.; Smoller, J.A. Global analysis of a system of predator–prey equations. SIAM J. Appl. Math. 1986, 46, 630–642. [Google Scholar] [CrossRef]
- Wang, J.; Shi, J.; Wei, J. Predator–prey system with strong Allee effect in prey. J. Math. Biol. 2011, 62, 291–331. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.A.; Kareiva, P. Allee dynamics and the spread of invading organisms. Theor. Popul. Biol. 1993, 43, 141–158. [Google Scholar] [CrossRef]
- Amarasekare, P. Allee effects in metapopulation dynamics. Am. Nat. 1998, 152, 298–302. [Google Scholar] [CrossRef]
- Zhou, S.; Liu, Y.; Wang, G. The stability of predator–prey systems subject to the Allee effects. Theor. Popul. Bio. 2005, 67, 23–31. [Google Scholar] [CrossRef]
- Gascoigne, J.C.; Lipcius, R.N. Allee effects driven by predation. J. Appl. Ecol. 2004, 41, 801–810. [Google Scholar] [CrossRef]
- Wilson, E.O.; Bossert, W.H. A Primer of Population Biology; Sinauer Assiates, Inc.: Sunderland, UK, 1971. [Google Scholar]
- Boukal, D.S.; Berec, L. Single-species models and the Allee effect: Extinction boundaries, sex ratios and mate encounters. J. Theoret. Biol. 2002, 218, 375–394. [Google Scholar] [CrossRef]
- Bai, D.; Kang, Y.; Ruan, S.; Wang, L. Dynamics of an intraguild predation food web model with strong Allee effect in the basal prey. Nonlinear Anal. RWA 2021, 58, 103206. [Google Scholar] [CrossRef]
- Kostitzin, V.A. Sur la loi logistique et ses generalizations. Acta Biotheor. 1940, 5, 155–159. [Google Scholar] [CrossRef]
- Wang, W. Population dispersal and Allee effect. Ric. Mat. 2016, 65, 535–548. [Google Scholar] [CrossRef]
- Dennis, B. The Dynamics of Low Density Populations. Ph.D. Thesis, The Pennsylvania State University, State College, PA, USA, 1982. [Google Scholar]
- Jacobs, J. Cooperation, optimal density and low density thresholds: Yet another modification of the logistic model. Oecologia 1984, 64, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Song, Y.; Yu, P. Delay-induced triple-zero bifurcation in a delayed Leslie-type predator–prey model with additive Allee effect. Int. J. Bifurc. Chaos 2016, 26, 1650117. [Google Scholar] [CrossRef]
- Aguirre, P.; González-Olivares, E.; Sáez, E. Two limit cycles in a Leslie-Gower predator–prey model with additive Allee effect. Nonlinear Anal. RWA 2009, 10, 1401–1416. [Google Scholar] [CrossRef]
- Aguirre, P.; González-Olivares, E.; Sáez, E. Three limit cycles in a Leslie-Gower predator–prey model with additive Allee effect. SIAM J. Appl. Math. 2009, 69, 1244–1269. [Google Scholar] [CrossRef]
- Terry, A.J. Predator–prey models with component Allee effect for predator reproduction. J. Math. Biol. 2015, 71, 1325–1352. [Google Scholar] [CrossRef]
- Cai, L.; Zhao, C.; Wang, W.; Wang, J. Dynamics of a Leslie-Gower predator–prey model with additive Allee effect. Appl. Math. Model. 2015, 39, 2092–2106. [Google Scholar] [CrossRef]
- Smith, H. An Introduction to Delay Differential Equations with Applications to the Life Sciences; Springer: New York, NY, USA, 2011. [Google Scholar]
- Kuang, Y. Delay Differential Equations: With Applications in Population Dynamics; Academic Press: New York, NY, USA, 1993. [Google Scholar]
- Wei, J.; Wang, H.; Jiang, W. Bifurcation Theory of Delay Differential Equations; Science Press: Beijing, China, 2012. (In Chinese) [Google Scholar]
- Hassard, B.D.; Kazarinoff, N.D.; Wan, Y.H. Theory and Applications of Hopf Bifurcation; Cambridge University Press: Cambridge, UK, 1981. [Google Scholar]
- Ma, Z. Mathematical Modeling and Research of Population Ecology; Anhui Education Press: Hefei, China, 1996. (In Chinese) [Google Scholar]
- Chen, L. Mathematical Ecological Models and Research Methods; Science Press: Beijing, China, 1988. (In Chinese) [Google Scholar]
- Wiggins, S. Introduction to Applied Nonlinear Dynamical Systems and Chaos, Texts in Applied Mathematics, 2nd ed.; Springer: New York, NY, USA, 1990. [Google Scholar]
- Guckenheimer, J.; Holmes, P. Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields; Springer: New York, NY, USA, 1983. [Google Scholar]
- Banerjee, J.; Sasmal, S.K.; Layek, R.K. Supercritical and subcritical Hopf-bifurcations in a two-delayed prey–predator system with density-dependent mortality of predator and strong Allee effect in prey. BioSystems 2019, 180, 19–37. [Google Scholar] [CrossRef]
- Meng, X.; Li, J. Dynamical Behavior of a Delayed Prey-Predator-Scavenger System with Fear Effect and Linear Harvesting. Int. J. Biomath. 2020, 14, 2150024. [Google Scholar] [CrossRef]
- Deng, L.; Wang, X.; Peng, M. Hopf bifurcation analysis for a ratio-dependent predator–prey system with two delays and stage structure for the predator. Appl. Math. Comput. 2014, 231, 214–230. [Google Scholar] [CrossRef]
- Panday, P.; Samanta, S.; Pal, N.; Chattopadhyay, J. Delay induced multiple stability switch and chaos in a predator–prey model with fear effect. Math. Comput. Simulat. 2020, 172, 134–158. [Google Scholar] [CrossRef]
- Xu, C.; Tang, X.; Liao, M.; He, X. Bifurcation analysis in a delayed Lokta-Volterra predator–prey model with two delays. Nonlinear Dyn. 2011, 66, 169–183. [Google Scholar] [CrossRef]















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, D.; Zhang, X. Dynamics of a Predator–Prey Model with the Additive Predation in Prey. Mathematics 2022, 10, 655. https://doi.org/10.3390/math10040655
Bai D, Zhang X. Dynamics of a Predator–Prey Model with the Additive Predation in Prey. Mathematics. 2022; 10(4):655. https://doi.org/10.3390/math10040655
Chicago/Turabian StyleBai, Dingyong, and Xiaoxuan Zhang. 2022. "Dynamics of a Predator–Prey Model with the Additive Predation in Prey" Mathematics 10, no. 4: 655. https://doi.org/10.3390/math10040655
APA StyleBai, D., & Zhang, X. (2022). Dynamics of a Predator–Prey Model with the Additive Predation in Prey. Mathematics, 10(4), 655. https://doi.org/10.3390/math10040655