
Dynamics of a Reduced System Connected to the Investigation of an Infinite Network of Identical Theta Neurons



Abstract
:1. Introduction
2. Description of the Mathematical Model
- Set , so that , for any . As pointed out in [22], this approach is not suitable if one wants to understand the global behavior of the system, as each initial condition yields different system constants .
- Consequently, this choice is more appropriate for a global dynamical analysis.
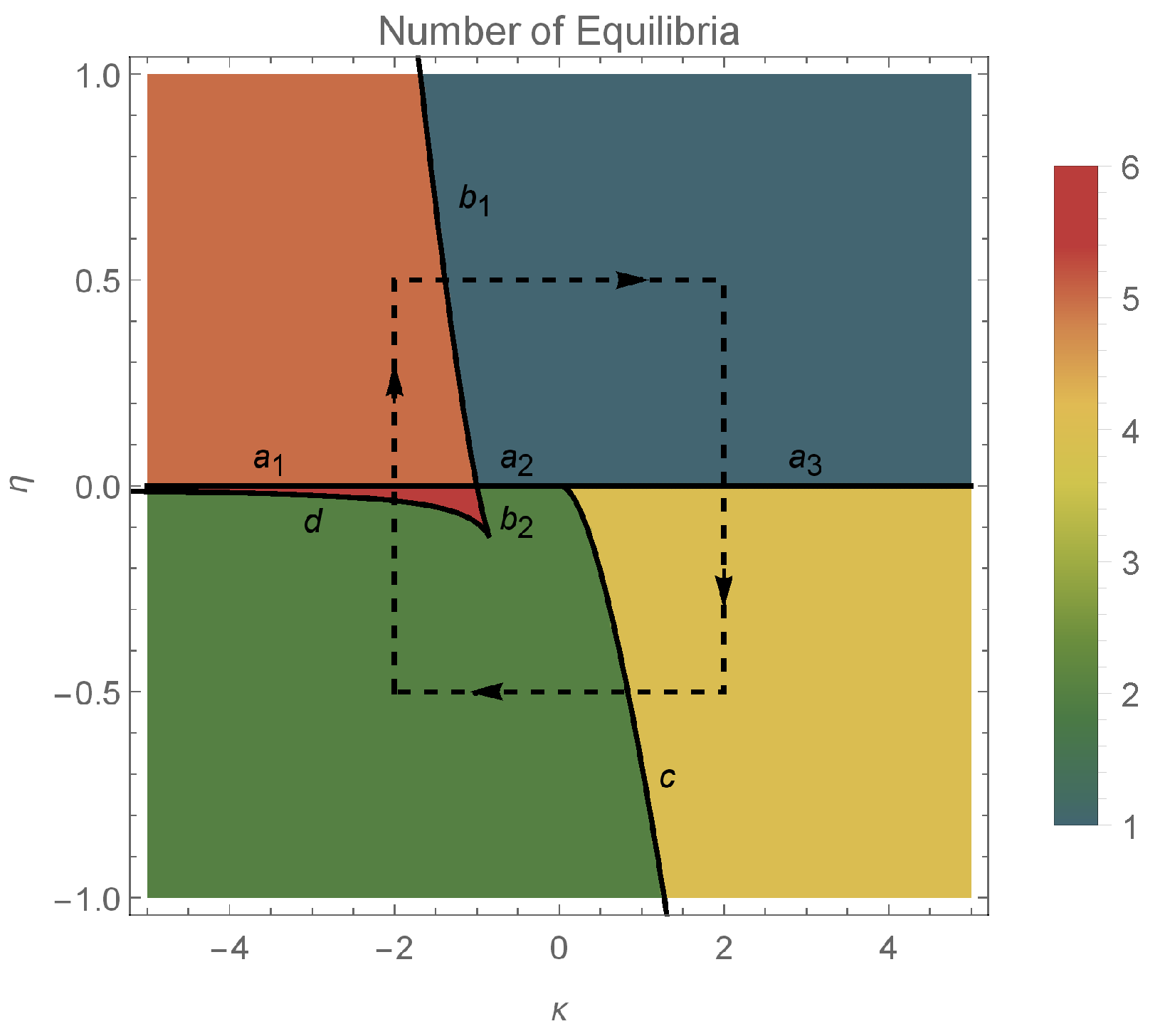
3. Stability of Equilibria
- Type 1 equilibria with (on the unit circle). For the infinite network of identical neurons, corresponds to full locking [21], and, hence, are all equal to .
- Type 2 equilibria with (on the real axis). These equilibrium points correspond to splay states, in which all the neurons of the infinite network follow the same trajectory but are equally displaced from one another in time.
3.1. Type 1 Equilibria
- excitatory coupling:
- if , the system has exactly one pair of equilibrium points of type 1: a source and a sink;
- if , there are no equilibria of type 1.
- inhibitory coupling:
- if , we have two subcases:
- –
- if belongs to the green area in Figure 1 (left), then the system has one pair of equilibirium points, a source and a sink;
- –
- if , we have two subcases:
3.2. Type 2 Equilibria
- excitatory coupling:
- if , there are two subcases:
- if , there is a unique type 2 equilibrium which is a center.
- inhibitory coupling:
- if , there are no type 2 equilibria;
- if , there is a unique type 2 equilibrium which is a center.
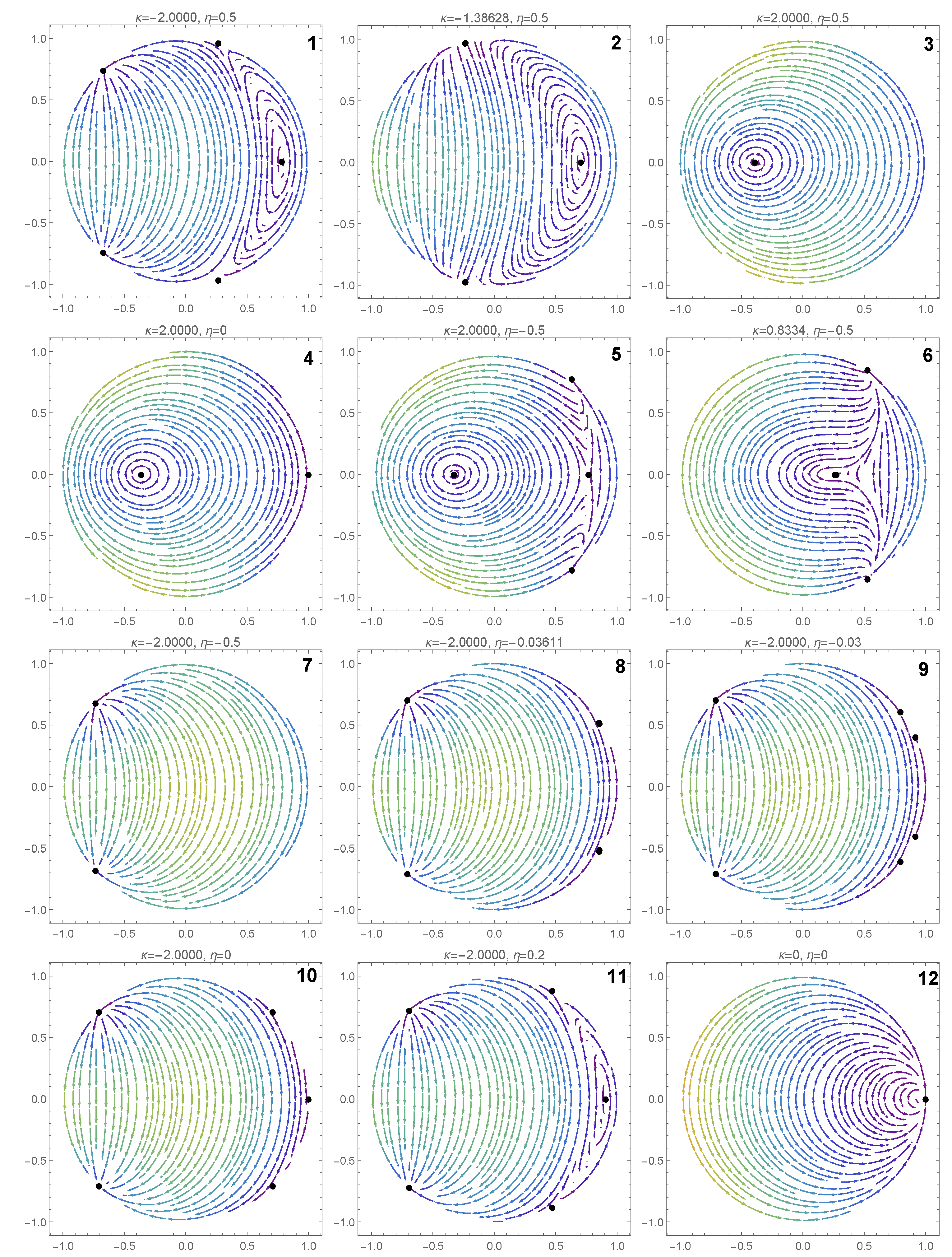
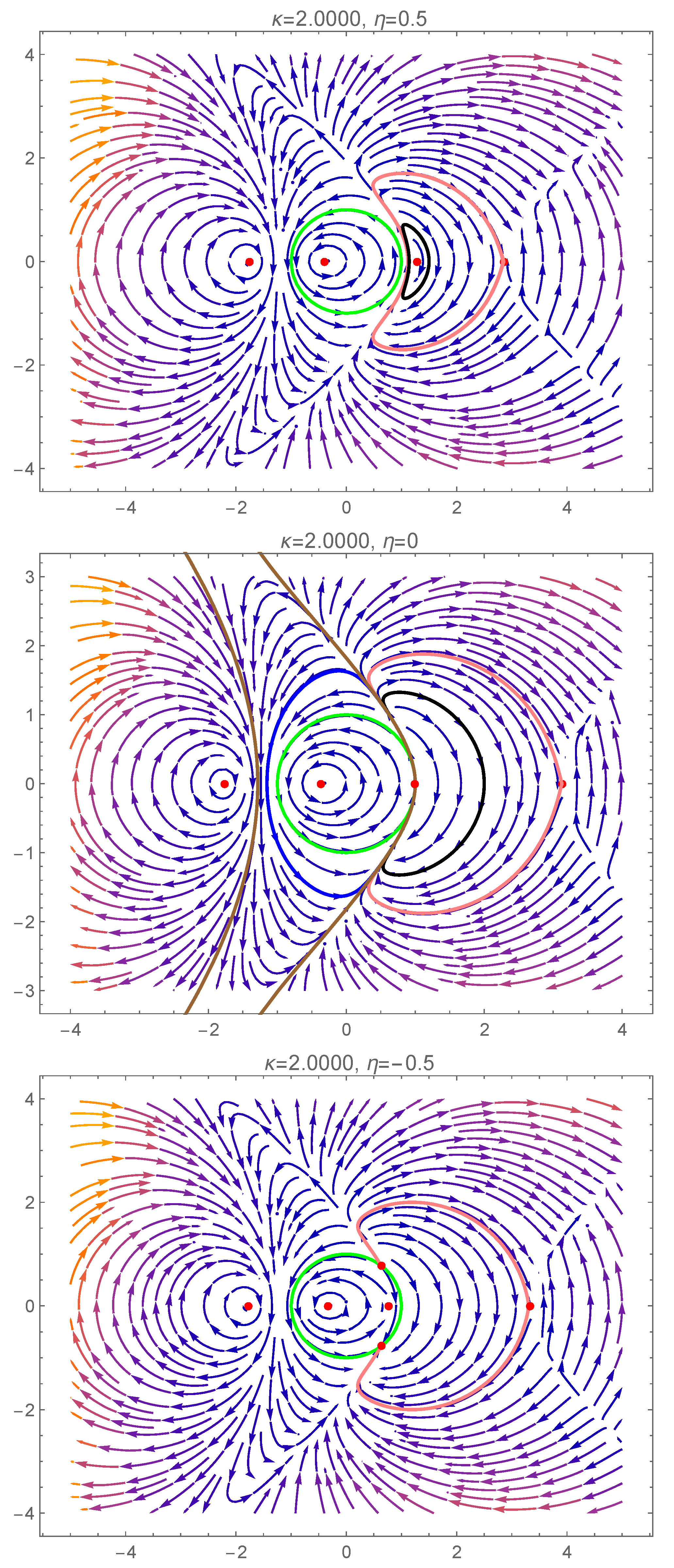
4. Bifurcation Phenomena and Dynamics of the Reduced System
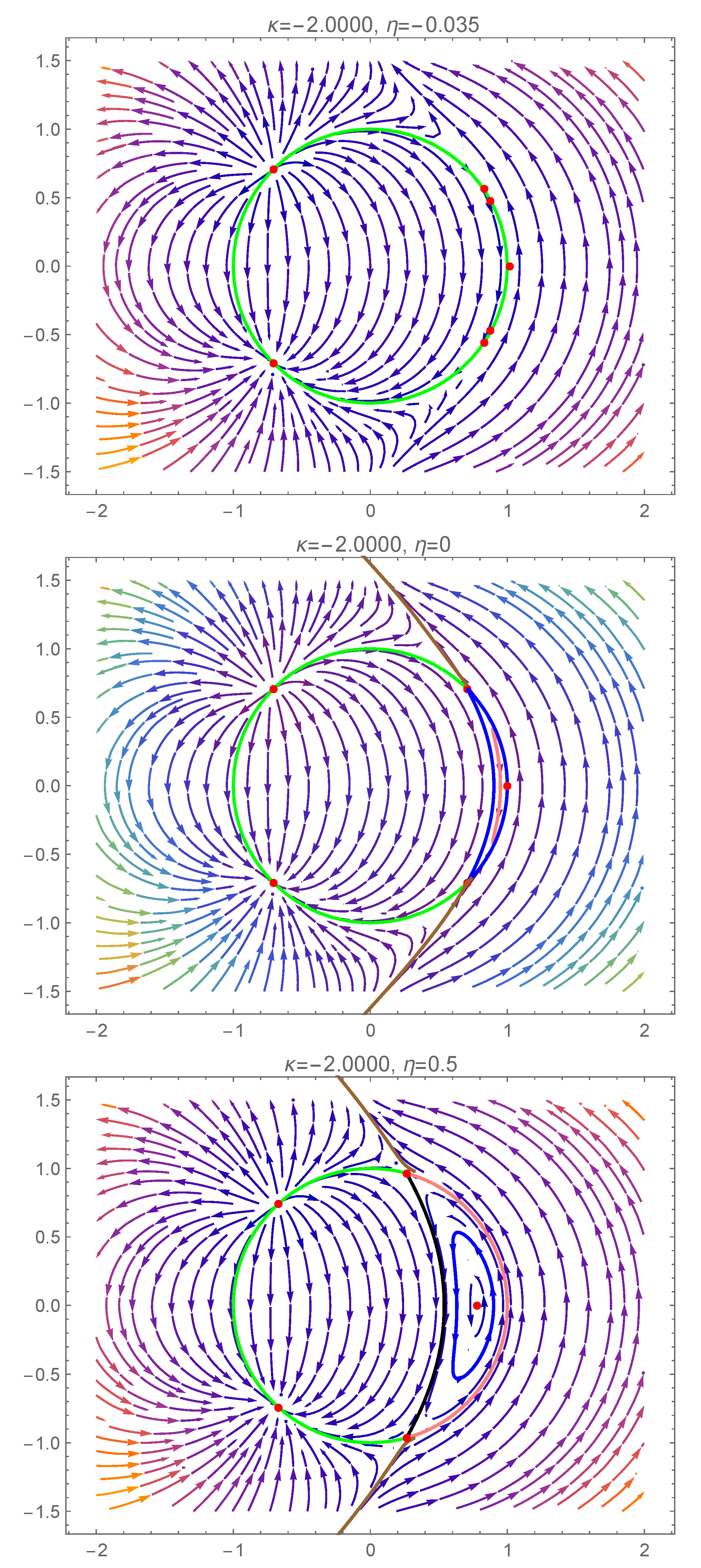
5. Bifurcations Involving the Degenerate Center
5.1. First Scenario
5.2. Second Scenario
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Watanabe–Strogatz Transformations
Appendix B. Reduction of Equations for the Infinite Dimensional Case
References
- Bem, T.; Rinzel, J. Short duty cycle destabilizes a half-center oscillator, but gap junctions can restabilize the anti-phase pattern. J. Neurophysiol. 2004, 91, 693–703. [Google Scholar] [CrossRef]
- Bressloff, P.C. Spatiotemporal dynamics of continuum neural fields. J. Phys. A Math. Theor. 2011, 45, 033001. [Google Scholar] [CrossRef]
- Coombes, S. Waves, bumps, and patterns in neural field theories. Biol. Cybern. 2005, 93, 91–108. [Google Scholar] [CrossRef]
- Coombes, S.; beim Graben, P.; Potthast, R.; Wright, J. Neural Fields: Theory and Applications; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Hoppensteadt, F.C.; Izhikevich, E.M. Associative memory of weakly connected oscillators. In Proceedings of the International Conference on Neural Networks (ICNN’97), Houston, TX, USA, 9–12 June 1997; Volume 2, pp. 1135–1138. [Google Scholar]
- Fenichel, N.; Moser, J. Persistence and smoothness of invariant manifolds for flows. Indiana Univ. Math. J. 1971, 21, 193–226. [Google Scholar] [CrossRef]
- Guckenheimer, J.; Holmes, P. Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; Volume 42. [Google Scholar]
- Kopell, N.; Ermentrout, G.B. Coupled oscillators and the design of central pattern generators. Math. Biosci. 1988, 90, 87–109. [Google Scholar] [CrossRef]
- Cabral, J.; Hugues, E.; Sporns, O.; Deco, G. Role of local network oscillations in resting-state functional connectivity. Neuroimage 2011, 57, 130–139. [Google Scholar] [CrossRef]
- Strogatz, S.H. From Kuramoto to Crawford: Exploring the onset of synchronization in populations of coupled oscillators. Phys. D Nonlinear Phenom. 2000, 143, 1–20. [Google Scholar] [CrossRef]
- Popovych, O.V.; Maistrenko, Y.L.; Tass, P.A. Phase chaos in coupled oscillators. Phys. Rev. E 2005, 71, 065201. [Google Scholar] [CrossRef] [PubMed]
- Bick, C.; Timme, M.; Paulikat, D.; Rathlev, D.; Ashwin, P. Chaos in symmetric phase oscillator networks. Phys. Rev. Lett. 2011, 107, 244101. [Google Scholar] [CrossRef]
- Li, K.; Bao, H.; Li, H.; Ma, J.; Hua, Z.; Bao, B. Memristive Rulkov Neuron Model With Magnetic Induction Effects. IEEE Trans. Ind. Inform. 2022, 18, 1726–1736. [Google Scholar] [CrossRef]
- Hua, M.; Bao, H.; Wu, H.; Xu, Q.; Bao, B. A single neuron model with memristive synaptic weight. Chin. J. Phys. 2022, 76, 217–227. [Google Scholar] [CrossRef]
- Bao, H.; Ding, R.; Hua, M.; Wu, H.; Chen, B. Initial-Condition Effects on a Two-Memristor-Based Jerk System. Mathematics 2022, 10, 411. [Google Scholar] [CrossRef]
- Chen, B.; Cheng, X.; Bao, H.; Chen, M.; Xu, Q. Extreme Multistability and Its Incremental Integral Reconstruction in a Non-Autonomous Memcapacitive Oscillator. Mathematics 2022, 10, 754. [Google Scholar] [CrossRef]
- Ermentrout, B. Type I membranes, phase resetting curves, and synchrony. Neural Comput. 1996, 8, 979–1001. [Google Scholar] [CrossRef] [PubMed]
- Laing, C.R. Derivation of a neural field model from a network of theta neurons. Phys. Rev. E 2014, 90, 010901. [Google Scholar] [CrossRef] [PubMed]
- Luke, T.B.; Barreto, E.; Thus, P. Complete classification of the macroscopic behavior of a heterogeneous network of theta neurons. Neural Comput. 2013, 25, 3207–3234. [Google Scholar] [CrossRef]
- Gutkin, B. Theta neuron model. In Encyclopedia of Computational Neuroscience; Springer: New York, NY, USA, 2015; pp. 2958–2965. [Google Scholar]
- Laing, C.R. The dynamics of networks of identical theta neurons. J. Math. Neurosci. 2018, 8, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Strogatz, S. Constants of motion for superconducting Josephson arrays. Phys. D 1991, 74, 197–253. [Google Scholar] [CrossRef]
- Watanabe, S.; Strogatz, S.H. Integrability of a globally coupled oscillator array. Phys. Rev. Lett. 1993, 70, 2391. [Google Scholar] [CrossRef] [PubMed]
- Ott, E.; Antonsen, T.M. Low dimensional behavior of large systems of globally coupled oscillators. Chaos Interdiscip. J. Nonlinear Sci. 2008, 18, 037113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, E.; Antonsen, T.M. Long time evolution of phase oscillator systems. Chaos: Interdiscip. J. Nonlinear Sci. 2009, 19, 023117. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.A.; Barreto, E.; Strogatz, S.H.; Ott, E.; Thus, P.; Antonsen, T.M. Exact results for the Kuramoto model with a bimodal frequency distribution. Phys. Rev. E 2009, 79, 026204. [Google Scholar] [CrossRef]
- Pazó, D.; Montbrió, E. Low-dimensional dynamics of populations of pulse-coupled oscillators. Phys. Rev. X 2014, 4, 011009. [Google Scholar] [CrossRef]
- Laing, C.R. Exact neural fields incorporating gap junctions. SIAM J. Appl. Dyn. Syst. 2015, 14, 1899–1929. [Google Scholar] [CrossRef]
- Montbrió, E.; Pazó, D.; Roxin, A. Macroscopic description for networks of spiking neurons. Phys. Rev. X 2015, 5, 021028. [Google Scholar] [CrossRef]
- Thus, P.; Luke, T.B.; Barreto, E. Networks of theta neurons with time-varying excitability: Macroscopic chaos, multistability, and final-state uncertainty. Phys. D Nonlinear Phenom. 2014, 267, 16–26. [Google Scholar]
- Pikovsky, A.; Rosenblum, M. Partially integrable dynamics of hierarchical populations of coupled oscillators. Phys. Rev. Lett. 2008, 101, 264103. [Google Scholar] [CrossRef] [PubMed]
- Llibre, J.; Pantazi, C. Limit cycles bifurcating from a degenerate center. Math. Comput. Simul. 2016, 120, 1–11. [Google Scholar] [CrossRef]
- Childs, L.M.; Strogatz, S.H. Stability diagram for the forced Kuramoto model. Chaos: Interdiscip. J. Nonlinear Sci. 2008, 18, 043128. [Google Scholar] [CrossRef]
- Marvel, S.A.; Mirollo, R.E.; Strogatz, S.H. Identical phase oscillators with global sinusoidal coupling evolve by Möbius group action. Chaos 2009, 19, 043104. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bîrdac, L.; Kaslik, E.; Mureşan, R. Dynamics of a Reduced System Connected to the Investigation of an Infinite Network of Identical Theta Neurons. Mathematics 2022, 10, 3245. https://doi.org/10.3390/math10183245
Bîrdac L, Kaslik E, Mureşan R. Dynamics of a Reduced System Connected to the Investigation of an Infinite Network of Identical Theta Neurons. Mathematics. 2022; 10(18):3245. https://doi.org/10.3390/math10183245
Chicago/Turabian StyleBîrdac, Lavinia, Eva Kaslik, and Raluca Mureşan. 2022. "Dynamics of a Reduced System Connected to the Investigation of an Infinite Network of Identical Theta Neurons" Mathematics 10, no. 18: 3245. https://doi.org/10.3390/math10183245
APA StyleBîrdac, L., Kaslik, E., & Mureşan, R. (2022). Dynamics of a Reduced System Connected to the Investigation of an Infinite Network of Identical Theta Neurons. Mathematics, 10(18), 3245. https://doi.org/10.3390/math10183245