
Spatiotemporal Dynamics in a Predator–Prey Model with Functional Response Increasing in Both Predator and Prey Densities


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Model Formulation
- 1.
- There is no coexistence equilibrium if .
- 2.
- There is a unique coexistence equilibrium if , where .
- 3.
- There are two coexistence equilibria and if , where are the two positive roots of (6) and .
3. Bifurcation Analysis
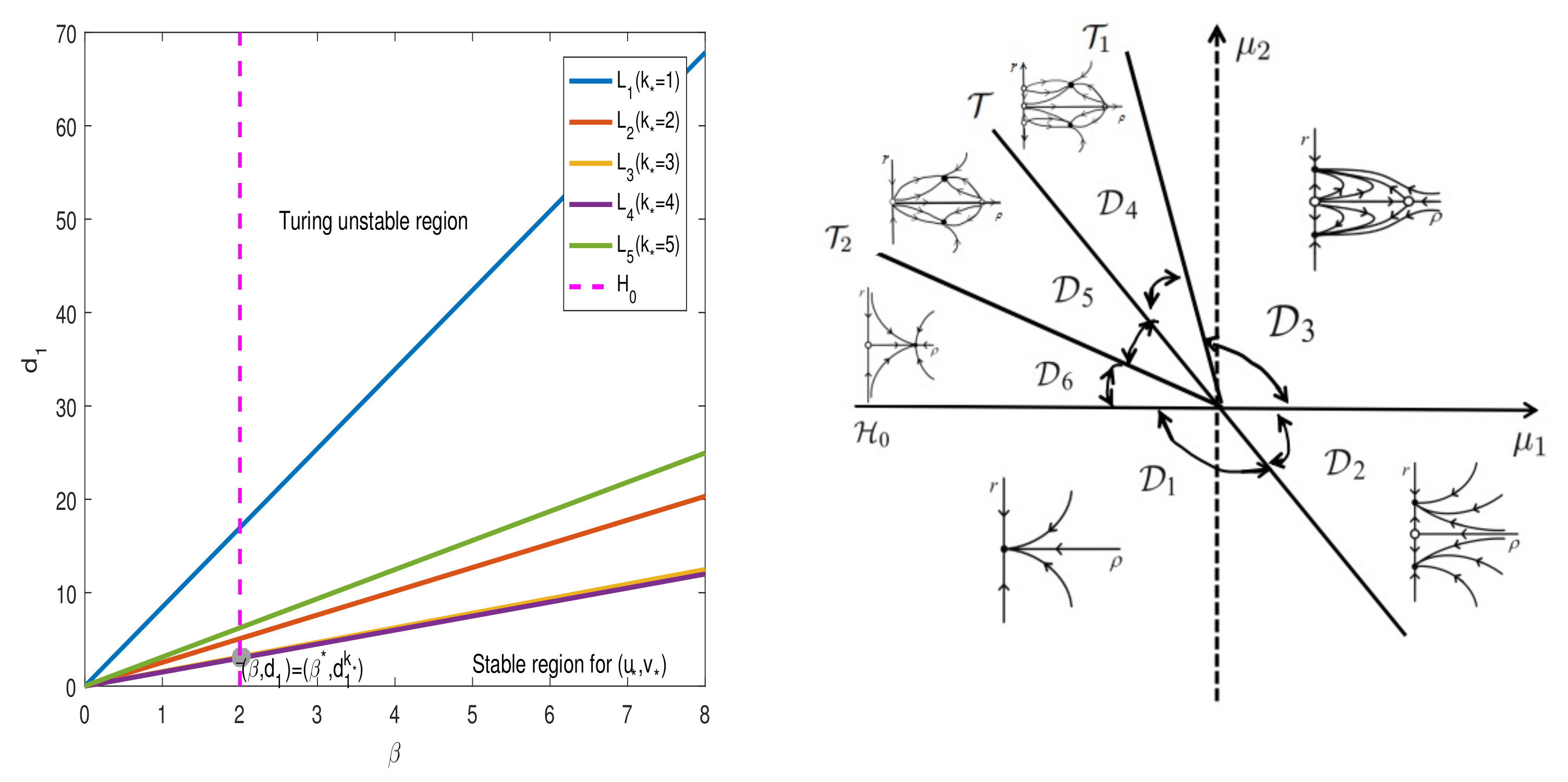
3.1. Turing Instability
3.2. Hopf Bifurcation
3.3. Turing–Hopf Bifurcation
4. Normal Forms for Turing–Hopf Bifurcation
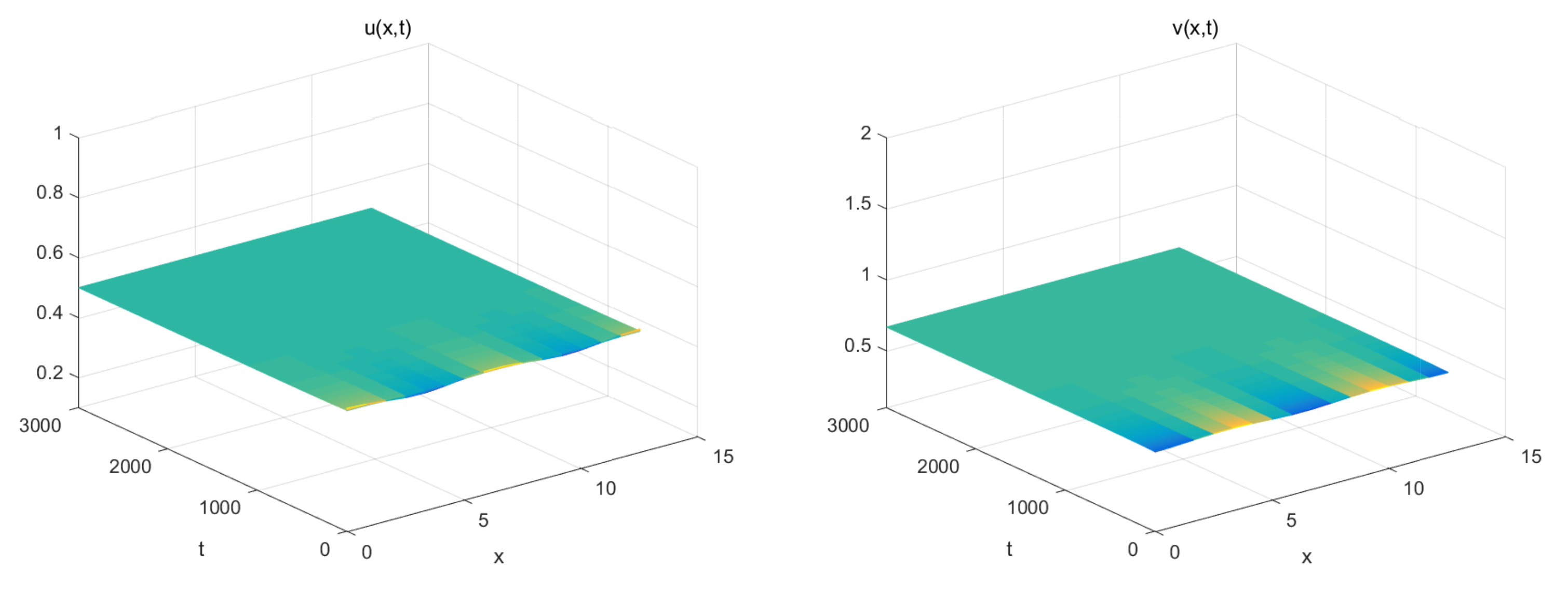
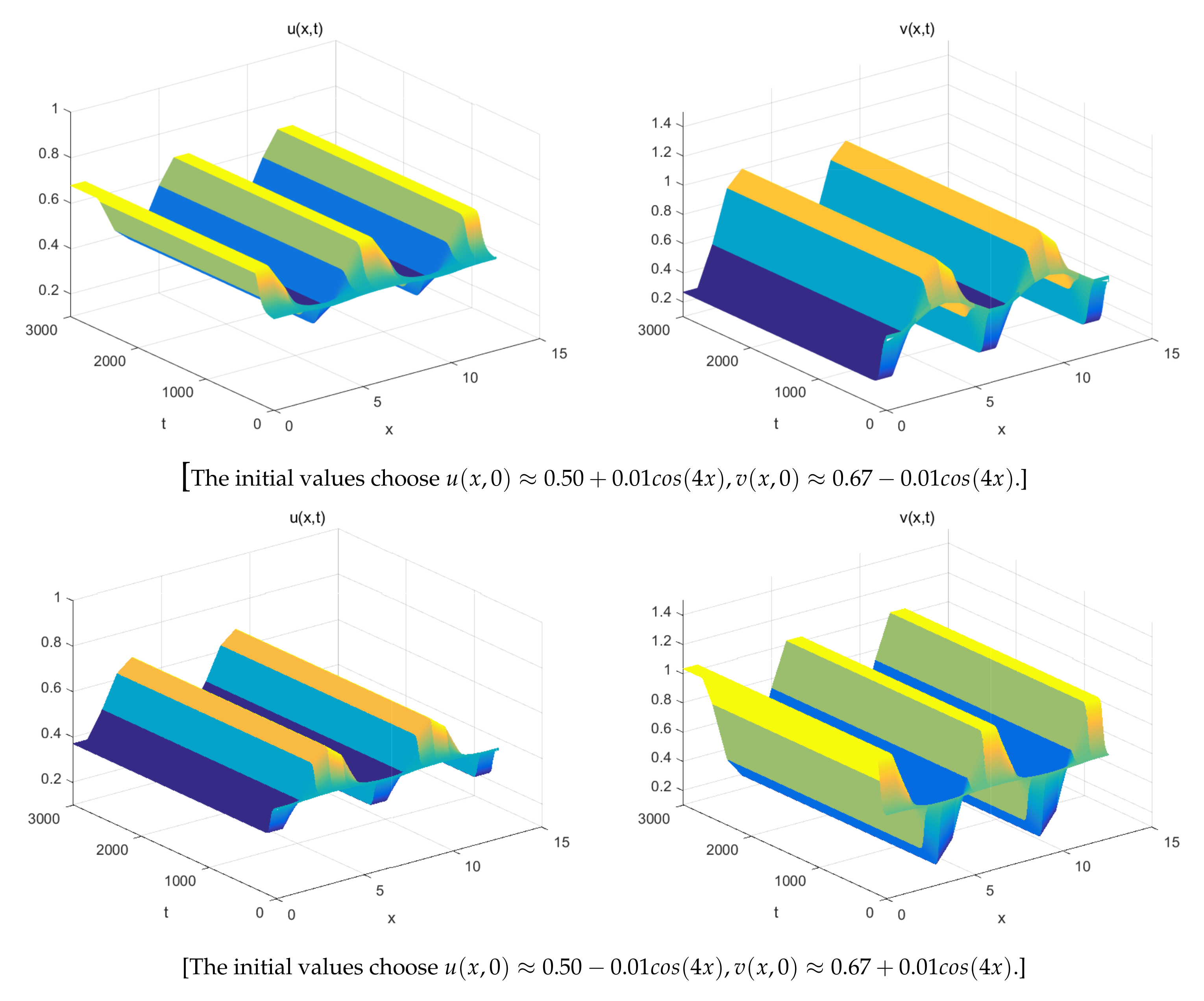
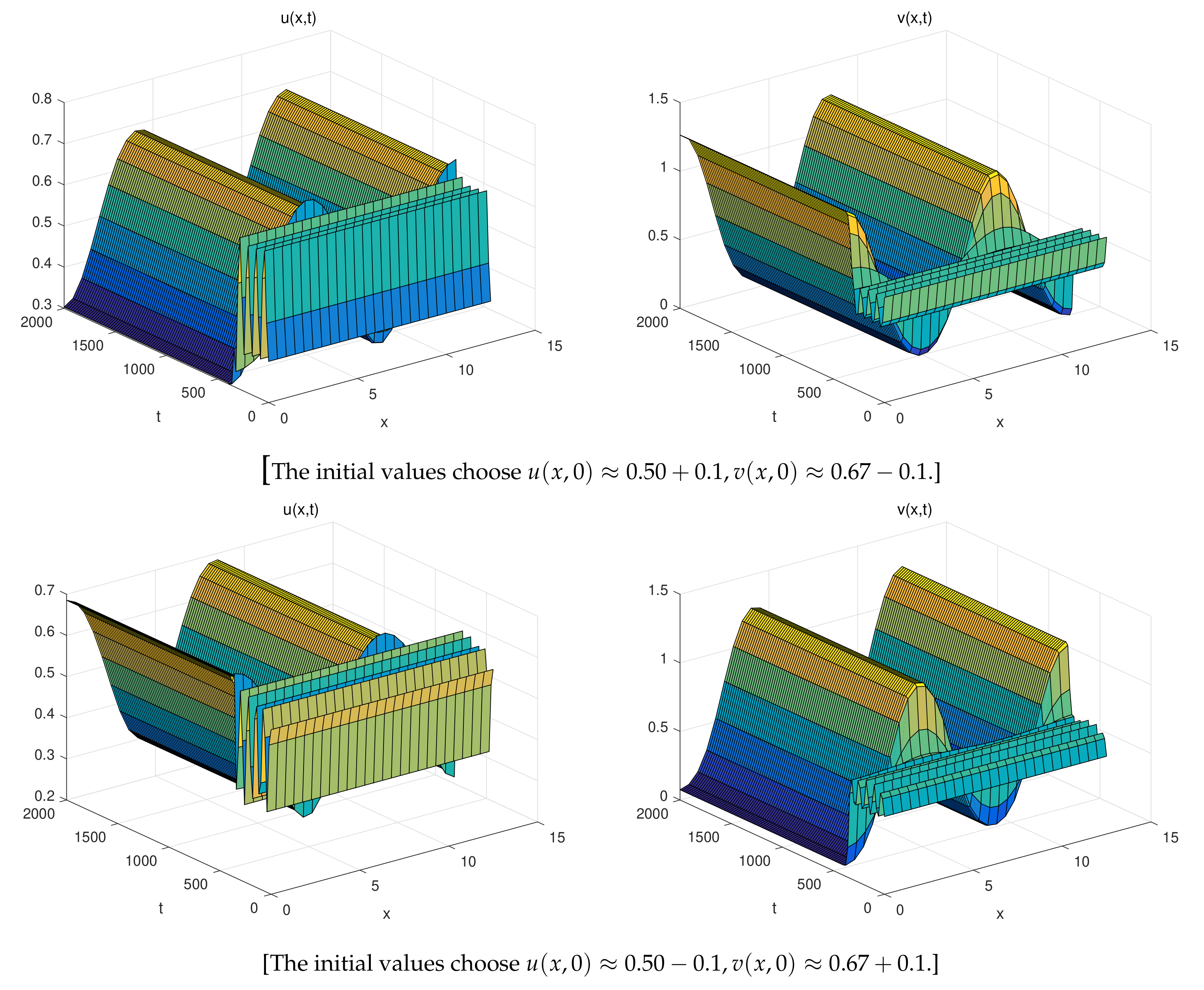
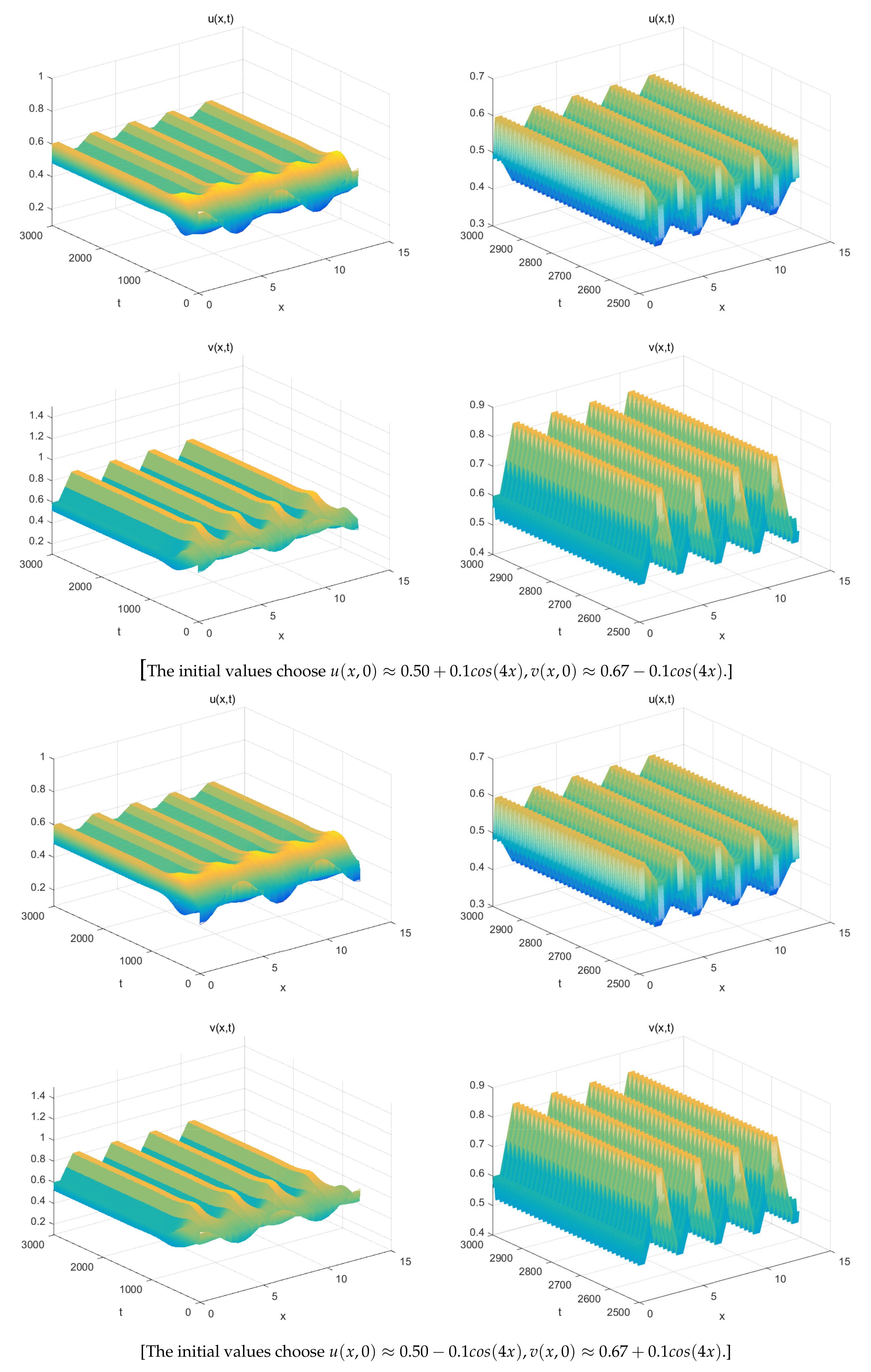
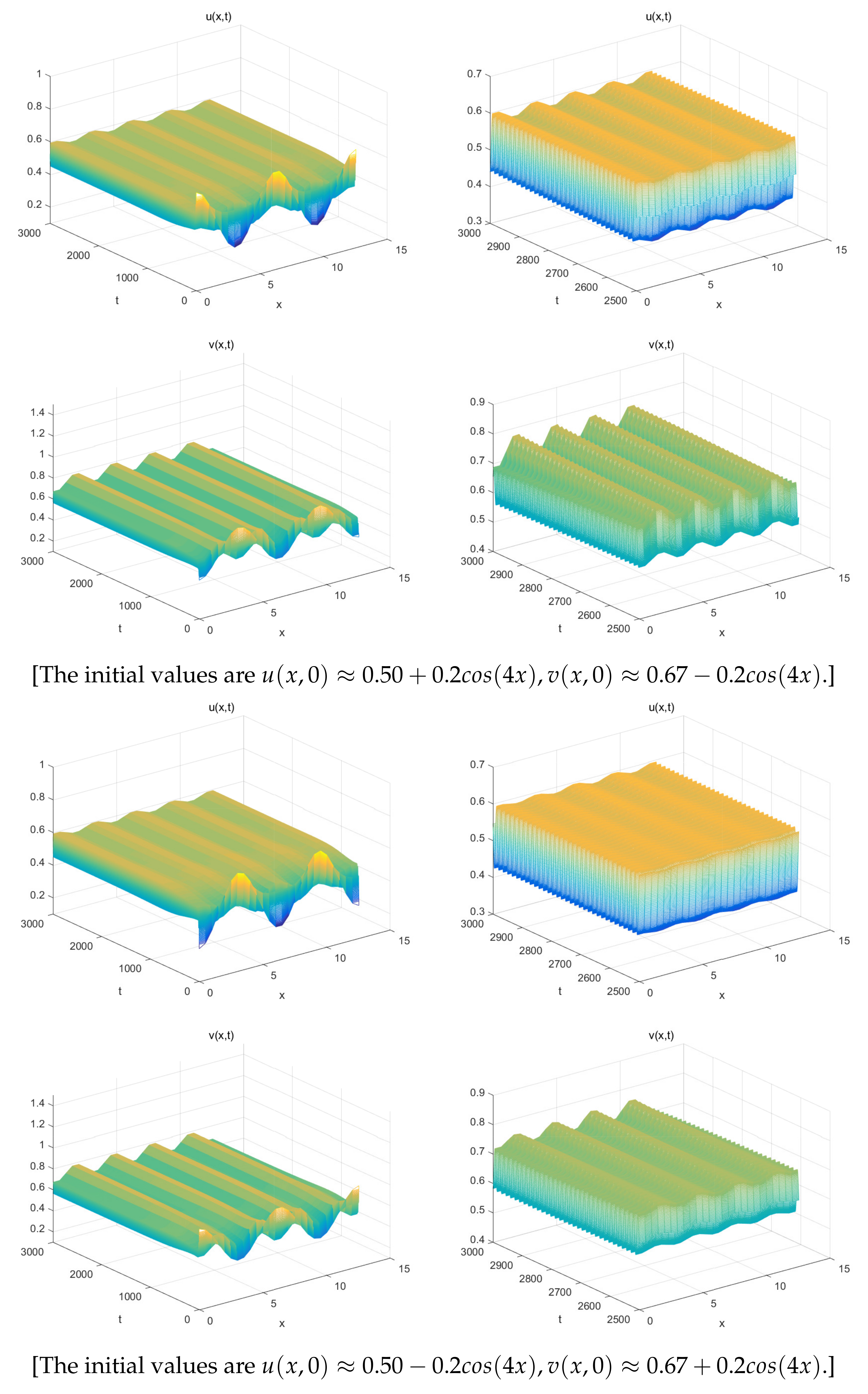
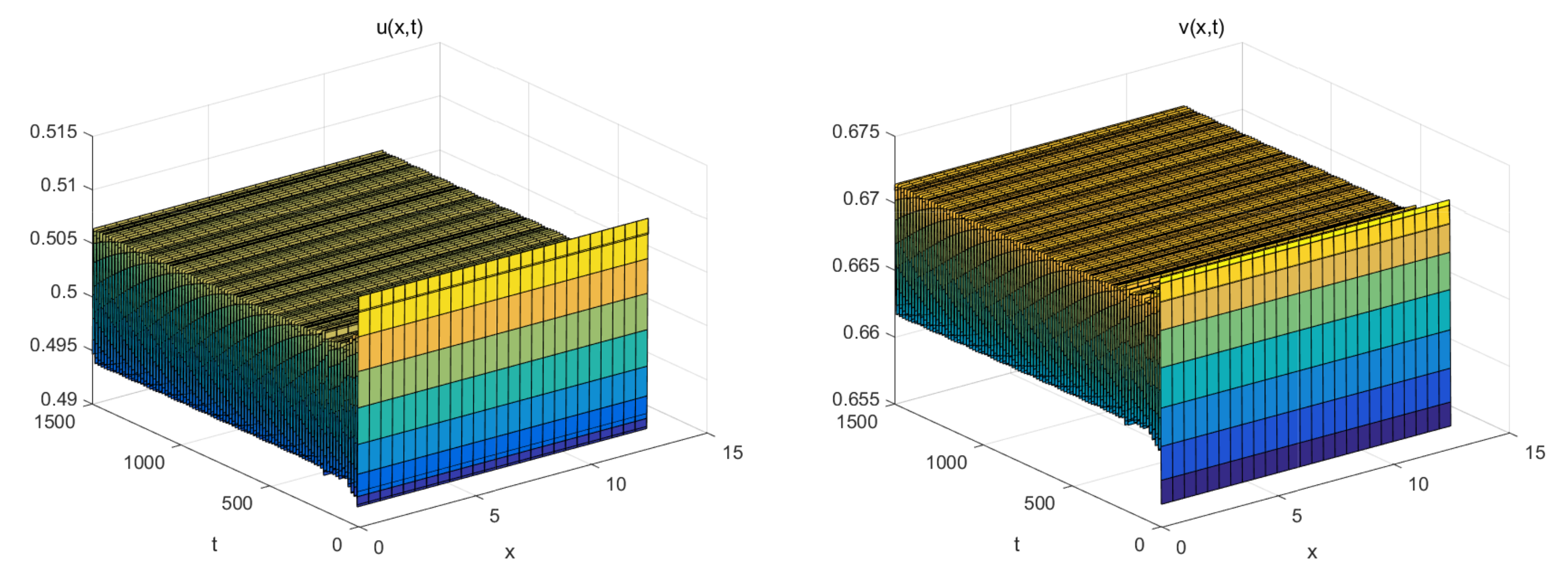
5. Numerical Simulations
- (1)
- The coexistence equilibrium: .
- (2)
- The spatially inhomogeneous steady states:
- (3)
- The spatially homogeneous periodic solution:
- (4)
- The spatially inhomogeneous periodic solutions:
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sabin, G.C.W.; Summers, D. Chaos in a periodically forced predator-prey ecosystem model. Math. Biosci. 1993, 113, 113. [Google Scholar] [CrossRef]
- Sadhu, S.; Kuehn, C. Stochastic mixed-mode oscillations in a three-species predator-prey model. Chaos 2018, 28, 033606. [Google Scholar] [CrossRef] [PubMed]
- Gilioli, G.; Pasquali, S.; Ruggeri, F. Nonlinear functional response parameter estimation in a stochastic predator-prey model. Math. Biosci. Eng. 2017, 9, 75–96. [Google Scholar]
- Shi, Q.; Shi, J.; Song, Y. Hopf bifurcation in a reaction-diffusion equation with distributed delay and Dirichlet boundary condition. J. Differ. Equ. 2017, 263, 6537–6575. [Google Scholar] [CrossRef]
- Bica, A.M.; Muresan, S. Smooth Dependence by LAG of the Solution of a Delay Integro-Differential Equation from Biomathematics. Commun. Math. Anal. 2006, 1, 64–74. [Google Scholar]
- Xiao, D.; Ruan, S. Multiple Bifurcations in a Delayed Predator-Prey System with Nonmonotonic Functional Response. J. Differ. Equ. 2001, 176, 494–510. [Google Scholar] [CrossRef] [Green Version]
- Lamontagne, Y.; Coutu, C.; Rousseau, C. Bifurcation analysis of a predator-prey system with generalised Holling type III functional response. J. Dyn. Differ. Equ. 2008, 20, 535–571. [Google Scholar] [CrossRef]
- Yang, R.; Ming, L.; Zhang, C. A delayed-diffusive predator-prey model with a ratio-dependent functional response. Commun. Nonlinear Sci. Numer. Simul. 2017, 53, S1007570417301508. [Google Scholar]
- Yang, R.; Zhang, C. Dynamics in a diffusive predator-prey system with a constant prey refuge and delay. Nonlinear Anal. Real World Appl. 2016, 31, 1–22. [Google Scholar] [CrossRef]
- Holling, C.S. The functional response of predators to prey density and its role in mimicry and population regulation. Mem. Entomol. Soc. Can. 1965, 97, 1–60. [Google Scholar] [CrossRef] [Green Version]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Holling, C.S.; Sra, B. A behavioral model of predator-prey functional responses. Syst. Res. Behav. Sci. 2010, 21, 183–195. [Google Scholar] [CrossRef]
- Partridge, B.L.; Johansson, J.; Kalish, J. The structure of schools of giant bluefin tuna in Cape Cod Bay. Environ. Biol. Fishes 1983, 9, 253–262. [Google Scholar] [CrossRef]
- Cosner, C.; Deangelis, D.L.; Ault, J.S. Effects of spatial grouping on the functional response of predators. Theor. Popul. Biol. 1999, 56, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Ryu, K.; Ko, W.; Haque, M. Bifurcation analysis in a predator-prey system with a functional response increasing in both predator and prey densities. Nonlinear Dyn. 2018, 94, 1639–1656. [Google Scholar] [CrossRef] [Green Version]
- Faria, T. Stability and bifurcation for a delayed predator-prey model and the effect of diffusion. J. Math. Anal. Appl. 2001, 254, 433–463. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Song, Y. Bifurcation analysis of a diffusive predator-prey system with a herd behavior and quadratic mortality. Math. Methods Appl. Sci. 2015, 38, 2994–3006. [Google Scholar] [CrossRef]
- Wolkowicz, G.S.K. Bifurcation analysis of a predator-prey system involving group defence. Siam J. Appl. Math. 1988, 48, 592–606. [Google Scholar] [CrossRef]
- Singh, M.K.; Bhadauria, B.S.; Singh, B.K. Bifurcation analysis of modified Leslie-Gower predator-prey model with double Allee effect. Ain Shams Eng. J. 2016, 9, S2090447916301095. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Peng, Y.; Zhang, T. The spatially inhomogeneous Hopf bifurcation induced by memory delay in a memory-based diffusion system. J. Differ. Equ. 2021, 300, 597–624. [Google Scholar] [CrossRef]
- Jiang, W.; An, Q.; Shi, J. Formulation of the normal forms of Turing-Hopf bifurcation in reaction-diffusion systems with time delay. J. Differ. Equ. 2020, 268, 6067–6102. [Google Scholar] [CrossRef]
- Yi, F. Turing instability of the periodic solutions for reaction-diffusion systems with cross-diffusion and the patch model with cross-diffusion-like coupling. J. Differ. Equ. 2021, 281, 379–410. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, R.; Song, Q.; An, Y. Spatiotemporal Dynamics in a Predator–Prey Model with Functional Response Increasing in Both Predator and Prey Densities. Mathematics 2022, 10, 17. https://doi.org/10.3390/math10010017
Yang R, Song Q, An Y. Spatiotemporal Dynamics in a Predator–Prey Model with Functional Response Increasing in Both Predator and Prey Densities. Mathematics. 2022; 10(1):17. https://doi.org/10.3390/math10010017
Chicago/Turabian StyleYang, Ruizhi, Qiannan Song, and Yong An. 2022. "Spatiotemporal Dynamics in a Predator–Prey Model with Functional Response Increasing in Both Predator and Prey Densities" Mathematics 10, no. 1: 17. https://doi.org/10.3390/math10010017
APA StyleYang, R., Song, Q., & An, Y. (2022). Spatiotemporal Dynamics in a Predator–Prey Model with Functional Response Increasing in Both Predator and Prey Densities. Mathematics, 10(1), 17. https://doi.org/10.3390/math10010017