Author Contributions
Conceptualization, B.E., K.B., R.S., C.S.C. and K.M.; Data curation, B.E. and M.T.; Formal analysis, B.E. and K.S.; Funding acquisition, M.E., R.S., C.S.C. and K.M.; Investigation, B.E., K.S. and M.T.; Methodology, B.E., K.S., M.T., R.S. and C.S.C.; Project administration, R.S., C.S.C. and K.M.; Resources, M.E., R.S. and C.S.C.; Software, B.E., K.S., M.T. and M.E.; Supervision, R.S., C.S.C. and K.M.; Validation, B.E.; Visualization, B.E., K.S. and M.T.; Writing—original draft, B.E., K.S. and M.T.; Writing—review and editing, B.E., M.T., K.B., R.S., C.S.C. and K.M. All authors have read and agreed to the published version of the manuscript.
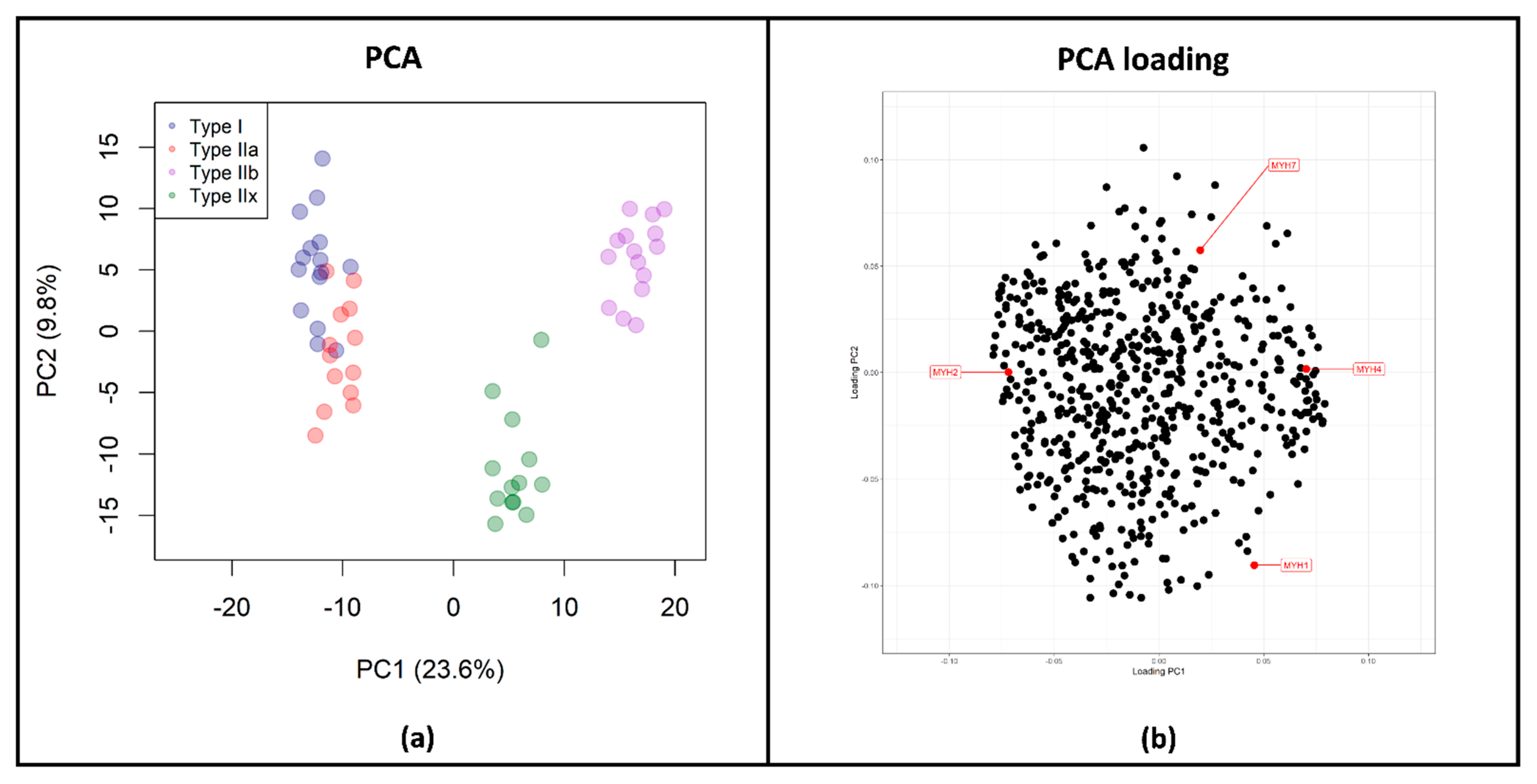
Figure 1.
Differentiation among the four fiber types. (a) Principal component analysis (PCA) of fiber types (n = 56). Significant separation of type IIb (purple) and type IIx fibers (green) can be observed in principle component (PC2), while type I (blue) and type IIa fibers (red) formed a combined, but significantly separated, cluster from type IIx and type IIb fibers in PC2 with moderate segregation into type I and type IIa fibers. (b) PCA loadings, visualizing the proteins separating the four groups. The four fiber type characteristic myosin heavy chain (MYH) isoforms are indicated in red and were identified as major separators.
Figure 1.
Differentiation among the four fiber types. (a) Principal component analysis (PCA) of fiber types (n = 56). Significant separation of type IIb (purple) and type IIx fibers (green) can be observed in principle component (PC2), while type I (blue) and type IIa fibers (red) formed a combined, but significantly separated, cluster from type IIx and type IIb fibers in PC2 with moderate segregation into type I and type IIa fibers. (b) PCA loadings, visualizing the proteins separating the four groups. The four fiber type characteristic myosin heavy chain (MYH) isoforms are indicated in red and were identified as major separators.
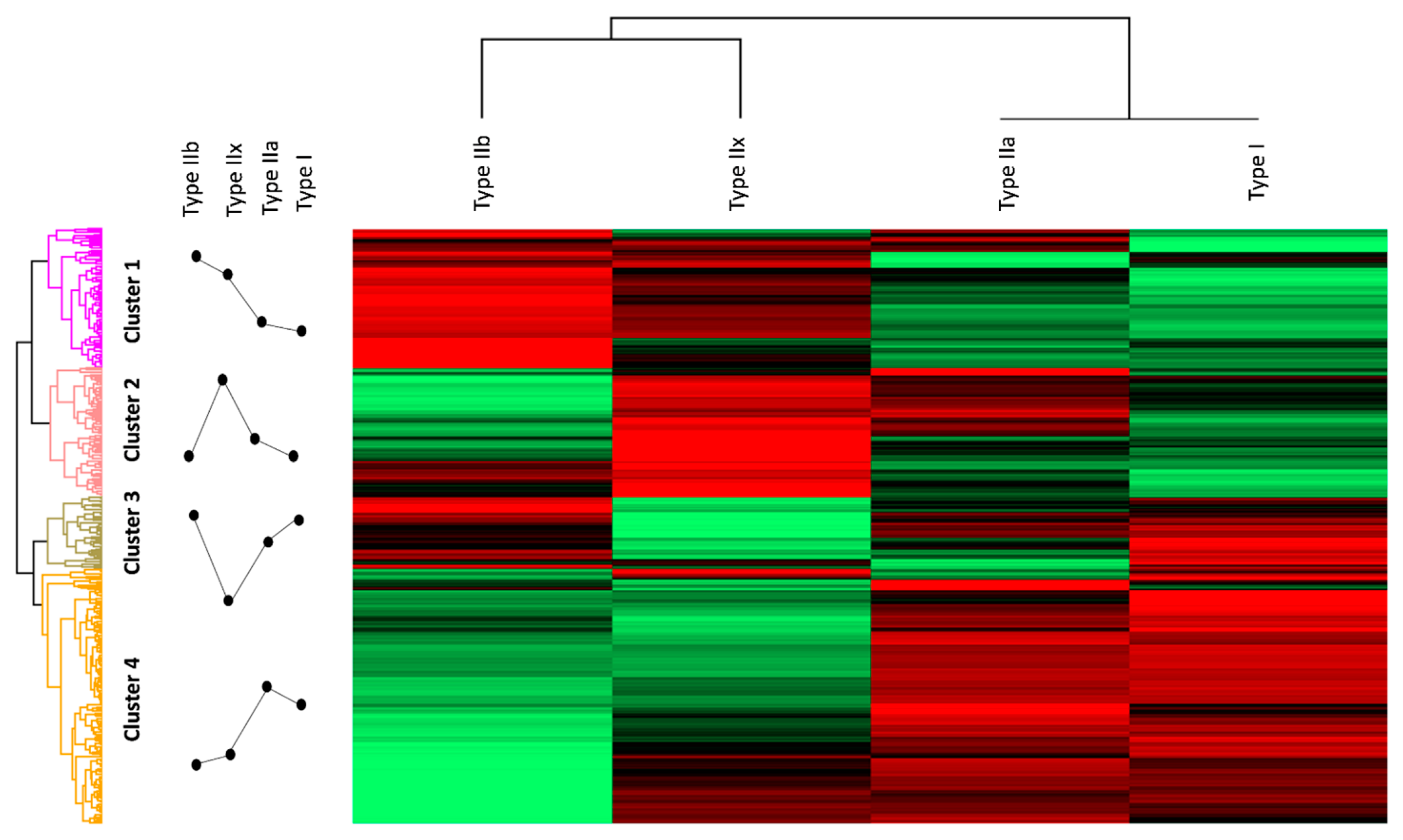
Figure 2.
Hierarchical clustering: high protein intensities are shown in red, and low protein intensities are shown in green. The profile plots indicate trends of protein expression intensity in the different fiber types. Cluster 1 displays high intensities in type IIx and type IIb fibers. Proteins of cluster 1 were annotated to glycolysis, calcium signaling, and the sarcoplasmic reticulum. High intensities in cluster 2 were annotated to type IIa and type IIx fibers and could be associated with mitochondrial processes such as the tricarboxylic acid (TCA) cycle the oxidative phosphorylation (OXPHOS) pathway and the mitochondrial respiratory chain complexes. Cluster 3 shows high-intensity values in type I, type IIa, and type IIb and is composed of ribosomal and proteasomal proteins. Cluster 4 can be annotated to structural components of the sarcomere as well as mitochondrial processes, such as the OXPHOS pathway, the TCA cycle, and the respiratory chain but also has a unique annotation to fatty acid beta oxidation and high intensities in GO terms and pathways associated with structural components of the sarcomere and the cytoskeleton.
Figure 2.
Hierarchical clustering: high protein intensities are shown in red, and low protein intensities are shown in green. The profile plots indicate trends of protein expression intensity in the different fiber types. Cluster 1 displays high intensities in type IIx and type IIb fibers. Proteins of cluster 1 were annotated to glycolysis, calcium signaling, and the sarcoplasmic reticulum. High intensities in cluster 2 were annotated to type IIa and type IIx fibers and could be associated with mitochondrial processes such as the tricarboxylic acid (TCA) cycle the oxidative phosphorylation (OXPHOS) pathway and the mitochondrial respiratory chain complexes. Cluster 3 shows high-intensity values in type I, type IIa, and type IIb and is composed of ribosomal and proteasomal proteins. Cluster 4 can be annotated to structural components of the sarcomere as well as mitochondrial processes, such as the OXPHOS pathway, the TCA cycle, and the respiratory chain but also has a unique annotation to fatty acid beta oxidation and high intensities in GO terms and pathways associated with structural components of the sarcomere and the cytoskeleton.

Figure 3.
(a) Principal component analysis (PCA) for all fiber types (n = 56) based on our complete proteomic data. Clear discrimination of type IIb and T2X fibers is possible, whereas type I and type IIa fibers cluster together. (b) PCA based on our alpha-actinin-3 and myosin-1 peptide panel. Discrimination of type I and type IIa fibers is especially enhanced. (c,d) Boxplots of the intensity value distributions on a log2-scale of the four fiber types, type I, type IIa, type IIb, and type IIx, for alpha-actinin-3 peptide (c) and the myosin-1 peptide (d), which were used in our panel. The alpha-actinin-3 peptide TINEVENQVLTR differentiates well between the type IIa, type IIb, and type IIx fibers, and the myosin-1-peptide IAEQELLDASER differentiates well between type I and all other fiber types, especially type IIa. Thus, only these two peptides are needed to differentiate all four fiber types.
Figure 3.
(a) Principal component analysis (PCA) for all fiber types (n = 56) based on our complete proteomic data. Clear discrimination of type IIb and T2X fibers is possible, whereas type I and type IIa fibers cluster together. (b) PCA based on our alpha-actinin-3 and myosin-1 peptide panel. Discrimination of type I and type IIa fibers is especially enhanced. (c,d) Boxplots of the intensity value distributions on a log2-scale of the four fiber types, type I, type IIa, type IIb, and type IIx, for alpha-actinin-3 peptide (c) and the myosin-1 peptide (d), which were used in our panel. The alpha-actinin-3 peptide TINEVENQVLTR differentiates well between the type IIa, type IIb, and type IIx fibers, and the myosin-1-peptide IAEQELLDASER differentiates well between type I and all other fiber types, especially type IIa. Thus, only these two peptides are needed to differentiate all four fiber types.
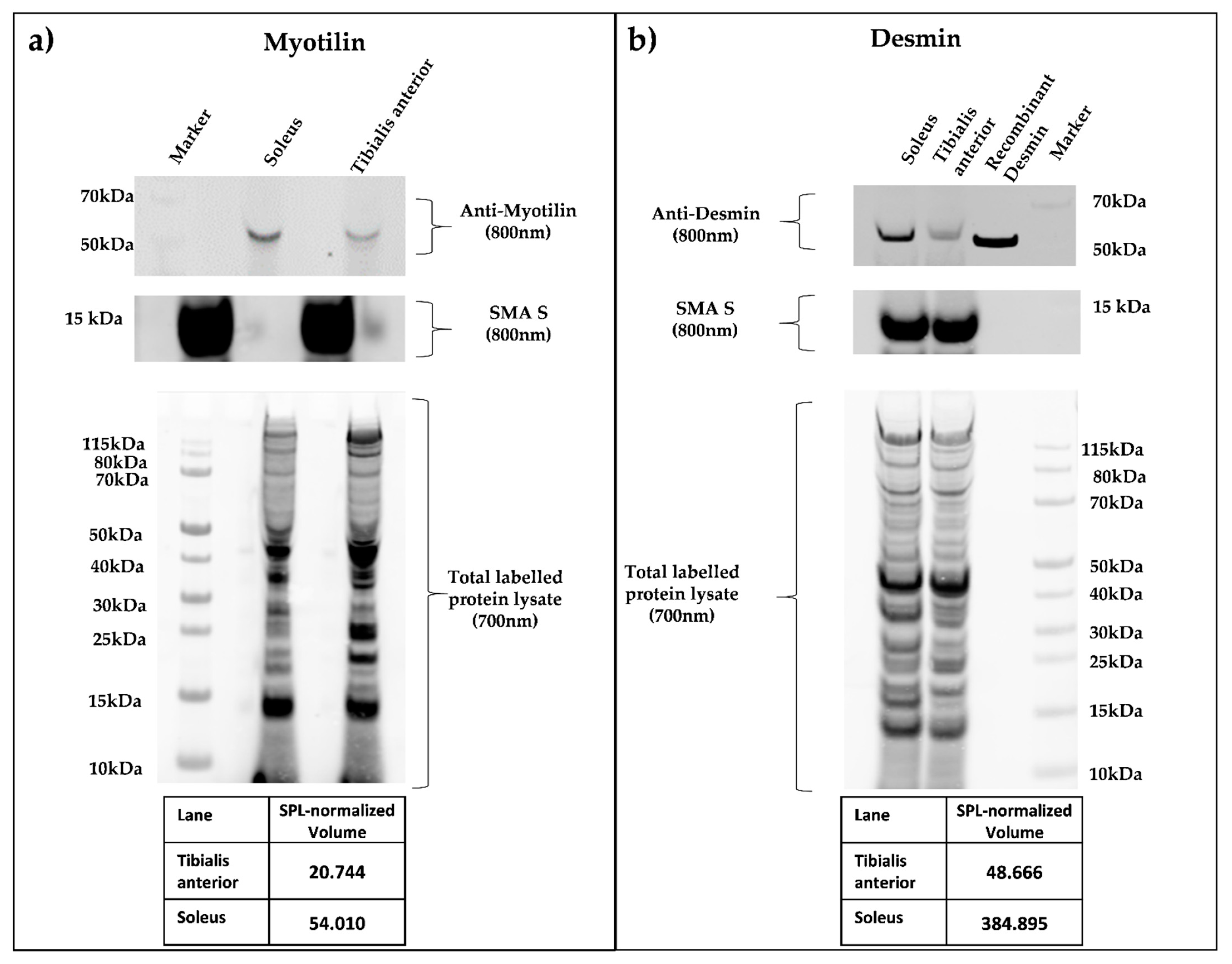
Figure 4.
(a,b) Quantitative Smart Protein Layers (SPL) Western blot analysis of whole murine soleus and tibialis anterior muscle. SPL Western blotting system (NH DyeAGNOSTICS GmbH) was used for the detection of quantitative differences between selected candidate proteins. Total protein was pre-labeled with a red fluorescent fluorophore (700 nm), and a 12.5 kDa green fluorescence-labeled standard protein (SMA S, 800 nm) was spiked in, enabling error correction of differing sample loading and data normalization between experiments. Primary antibodies against myotilin (a), myotilin (E-10), sc-393957), and desmin (b), DAKO, M0760 Clone D33) were visualized by infrared secondary antibodies (IRDye goat anti-mouse 800 nm). SPL system analysis resulted in quantitative protein volumes (SPL normalized volume), determining a higher concentration in the soleus muscle for both proteins.
Figure 4.
(a,b) Quantitative Smart Protein Layers (SPL) Western blot analysis of whole murine soleus and tibialis anterior muscle. SPL Western blotting system (NH DyeAGNOSTICS GmbH) was used for the detection of quantitative differences between selected candidate proteins. Total protein was pre-labeled with a red fluorescent fluorophore (700 nm), and a 12.5 kDa green fluorescence-labeled standard protein (SMA S, 800 nm) was spiked in, enabling error correction of differing sample loading and data normalization between experiments. Primary antibodies against myotilin (a), myotilin (E-10), sc-393957), and desmin (b), DAKO, M0760 Clone D33) were visualized by infrared secondary antibodies (IRDye goat anti-mouse 800 nm). SPL system analysis resulted in quantitative protein volumes (SPL normalized volume), determining a higher concentration in the soleus muscle for both proteins.
Table 1.
Detailed information on myosin isoforms (MYH): accession (Uniprot), protein name, protein name abbreviation, gene name, and corresponding fiber type.
Table 1.
Detailed information on myosin isoforms (MYH): accession (Uniprot), protein name, protein name abbreviation, gene name, and corresponding fiber type.
| Accession (Uniprot) | Protein Name | Protein Name Abbreviation | Gene Name | Corresponding
Fiber Type |
|---|
| Q91Z83 | Myosin-7 | MYH7 | Myh7 | Type I |
| G3UW82 | Myosin, heavy polypeptide 2, skeletal muscle, adult * | MYH2 | Myh2 | Type IIa |
| Q5SX39 | Myosin-4 | MYH4 | Myh4 | Type IIb |
| Q5SX40 | Myosin-1 | MYH1 | Myh1 | Type IIx |
Table 2.
Sample information. Mouse identifier (ID), sex, genotype, and age in months and days for each mouse used in the present study are displayed.
Table 2.
Sample information. Mouse identifier (ID), sex, genotype, and age in months and days for each mouse used in the present study are displayed.
| Mouse ID | Sex | Genotype | Age in Months | Age in Days |
|---|
| 1 | f | wt | 21 | 657 |
| 2 | f | wt | 21 | 657 |
| 3 | m | wt | 15 | 466 |
| 4 | m | wt | 14 | 448 |
| 5 | m | wt | 14 | 448 |
| 6 | m | wt | 14 | 448 |
| 7 | m | wt | 13 | 401 |
| 8 | m | wt | 11 | 361 |
| 9 | m | wt | 11 | 361 |
| 10 | m | wt | 11 | 361 |
| 11 | m | wt | 3 | 95 |
| 12 | m | wt | 3 | 95 |
| 13 | m | wt | 3 | 96 |
| 14 | f | wt | 3 | 96 |
| 15 | f | wt | 3 | 96 |
Table 3.
Primary antibodies (product numbers Developmental Studies Hybridoma Bank (DSHB), Iowa) and dilutions used for the detection of fiber types (type I, type IIa, type IIb, and type IIx) using the fiber type-specific myosin (MYH) isoform. Secondary antibodies and dilutions necessary for the detection of primary antibody signals.
Table 3.
Primary antibodies (product numbers Developmental Studies Hybridoma Bank (DSHB), Iowa) and dilutions used for the detection of fiber types (type I, type IIa, type IIb, and type IIx) using the fiber type-specific myosin (MYH) isoform. Secondary antibodies and dilutions necessary for the detection of primary antibody signals.
| Primary Antibody/Dilution | Secondary Antibody/Dilution | Fiber Type | MYH Isoform |
|---|
| BA-F8/1:20 | TexasRed/1:60 | Type I | MYH7 |
| SC-71/1:20 | Cy2/1:200 | Type IIa | MYH2 |
| BF-F3/1:20 | AlexaFluor594/1:500 | Type IIb | MYH4 |
| 6H1/1:15 | AlexaFluor594/1:500 | Type IIx | MYH1 |
Table 4.
Summed intensities (Sum) of unique peptides (proteotypicity = TRUE) detected for each myosin heavy chain (MYH) isoform that is indicative of fiber types (type I, type IIa, type IIb, and type IIx).
Table 4.
Summed intensities (Sum) of unique peptides (proteotypicity = TRUE) detected for each myosin heavy chain (MYH) isoform that is indicative of fiber types (type I, type IIa, type IIb, and type IIx).
| Protein | Peptide Sequence | Proteotypicity | Sum Type I (10 × 106) | Sum Type IIa (10 × 106) | Sum Type IIb (10 × 106) | Sum Type IIx (10 × 106) |
| MYH7 | DTQIQLDDAVR | TRUE | 226.57 | 45.22 | 0.00 | 0.00 |
| MYH7 | IEDEQALGSQLQK | TRUE | 162.65 | 34.33 | 0.05 | 0.19 |
| MYH7 | ANDDLKENIAIVER | TRUE | 137.41 | 27.70 | 0.01 | 0.01 |
| MYH7 | GQNVQQVSYAIGALAK | TRUE | 80.46 | 15.95 | 0.04 | 0.03 |
| MYH7 | SLQSLLK | TRUE | 74.67 | 15.45 | 0.02 | 0.03 |
| Protein | Peptide Sequence | Proteotypicity | Sum Type I (10 × 106) | Sum Type IIa (10 × 106) | Sum Type IIb (10 × 106) | Sum Type IIx (10 × 106) |
| MYH2 | LINDLTTQR | TRUE | 95.23 | 216.18 | 8.32 | 14.98 |
| MYH2 | AAYLQGLNSADLLK | TRUE | 77.60 | 179.82 | 8.17 | 11.76 |
| MYH2 | VLNASAIPEGQYIDSK | TRUE | 74.62 | 173.66 | 6.23 | 11.20 |
| MYH2 | GQTVEQVTNAVGALAK | TRUE | 39.80 | 106.67 | 3.76 | 7.45 |
| MYH2 | IEDEQAIGIQLQK | TRUE | 25.58 | 71.06 | 2.53 | 5.65 |
| Protein | Peptide Sequence | Proteotypicity | Sum Type I (10 × 106) | Sum Type IIa (10 × 106) | Sum Type IIb (10 × 106) | Sum Type IIx (10 × 106) |
| MYH4 | VAEQELLDASER | TRUE | 3.78 | 3.82 | 221.71 | 49.27 |
| MYH4 | LINELSTQK | TRUE | 4.17 | 4.55 | 175.11 | 37.76 |
| MYH4 | AAYLTSLNSADLLK | TRUE | 1.87 | 2.98 | 129.68 | 24.67 |
| MYH4 | LQDAEEHVEAVNSK | TRUE | 1.42 | 1.34 | 75.10 | 17.27 |
| MYH4 | RVAEQELLDASER | TRUE | 1.52 | 2.48 | 74.18 | 19.12 |
| Protein | Peptide Sequence | Proteotypicity | Sum Type I (10 × 106) | Sum Type IIa (10 × 106) | Sum Type IIb (10 × 106) | Sum Type IIx (10 × 106) |
| MYH1 | AAYLQNLNSADLLK | TRUE | 14.02 | 20.54 | 25.48 | 122.96 |
| MYH1 | SALAHALQSSR | TRUE | 14.19 | 20.28 | 17.76 | 97.12 |
| MYH1 | DSLVSQLSR | TRUE | 8.37 | 11.42 | 13.79 | 49.63 |
| MYH1 | NAYEESLDHLETLKR | TRUE | 4.60 | 6.71 | 8.68 | 41.85 |
| MYH1 | QLDEKDSLVSQLSR | TRUE | 4.59 | 6.82 | 8.77 | 41.53 |
Table 5.
Percentage values of sarcomeric protein markers and their indication for either slow or fast fibers in all four skeletal muscle fiber types (type I, type IIa, type IIb, and type IIx).
Table 5.
Percentage values of sarcomeric protein markers and their indication for either slow or fast fibers in all four skeletal muscle fiber types (type I, type IIa, type IIb, and type IIx).
| Protein Names | Indicative For | % Type I | % Type IIa | % Type IIb | % Type IIx |
|---|
| Alpha-actinin-2 | Slow | 2.60 | 1.89 | 0.39 | 1.15 |
| Alpha-actinin-3 | Fast | 0.10 | 0.10 | 0.93 | 0.63 |
| Myomesin 2 | Fast | 0.05 | 0.11 | 0.36 | 0.34 |
| Myomesin-1 | Slow | 0.04 | 0.02 | 0.00 | 0.00 |
| Myosin-binding protein C. slow type | Slow | 0.09 | 0.09 | 0.04 | 0.06 |
| Myosin-binding protein C. fast type | Fast | 0.08 | 0.11 | 0.64 | 0.33 |
| Myozenin-1 | Fast | 0.10 | 0.19 | 0.30 | 0.31 |
| Myozenin-2 | Slow | 0.20 | 0.14 | 0.01 | 0.02 |
| Troponin C. slow skeletal and cardiac muscles (TN-C) | Slow | 0.45 | 0.17 | 0.00 | 0.00 |
| Troponin I. fast skeletal muscle (troponin I. fast-twitch isoform) | Fast | 0.06 | 0.12 | 0.23 | 0.26 |
Table 6.
Percentages of manually annotated proteins into 13 groups (Sarcomere/Cytoskeleton, Respiratory chain, Tricarboxylic acid cycle, Glycolysis, Glycogenolysis, Mitochondria, Proteasome, Heat shock, Nucleus, Fatty acid beta oxidation, Calcium regulation/transport, Ribosome, Others) for each cluster.
Table 6.
Percentages of manually annotated proteins into 13 groups (Sarcomere/Cytoskeleton, Respiratory chain, Tricarboxylic acid cycle, Glycolysis, Glycogenolysis, Mitochondria, Proteasome, Heat shock, Nucleus, Fatty acid beta oxidation, Calcium regulation/transport, Ribosome, Others) for each cluster.
| Annotation | Cluster 1 (%) | Cluster 2 (%) | Cluster 3 (%) | Cluster 4 (%) |
|---|
| Sarcomere/Cytoskeleton | 18.24 | 5.11 | 14.29 | 19.56 |
| Respiratory chain | 3.38 | 37.23 | 5.19 | 7.38 |
| Tricarboxylic acid cycle | 0.68 | 7.30 | 1.30 | 1.48 |
| Glycolysis | 4.73 | 0.00 | 0.00 | 0.74 |
| Glycogenolysis | 5.41 | 0.73 | 0.00 | 0.00 |
| Mitochondria | 0.00 | 9.49 | 1.30 | 5.90 |
| Proteasome | 0.00 | 0.00 | 9.09 | 4.43 |
| Heat shock | 0.00 | 0.00 | 2.60 | 3.69 |
| Nucleus | 0.68 | 0.00 | 2.60 | 3.32 |
| Fatty acid beta oxidation | 2.03 | 0.73 | 1.30 | 5.17 |
| Calcium regulation/transport | 8.78 | 0.73 | 0.00 | 1.85 |
| Ribosome | 8.78 | 4.38 | 28.57 | 4.43 |
| Other | 47.30 | 34.31 | 33.77 | 42.07 |
Table 7.
Classification of fiber types according to the traditional classification based on ATPase staining, the oxidative phosphorylation capacity, the immunohistochemical classification based on myosin heavy chain (MYH) isoforms, and the classification based on the proteomic data.
Table 7.
Classification of fiber types according to the traditional classification based on ATPase staining, the oxidative phosphorylation capacity, the immunohistochemical classification based on myosin heavy chain (MYH) isoforms, and the classification based on the proteomic data.
| Fiber Type | Traditional Classification (Based on ATPase Staining) | Oxidative Phosphorylation Capacity | Immunohistochemical
Classification via MYH Isoform | Proteomic Classification |
|---|
| Type I | Slow | Very high | MYH7 | Highly oxidative |
| Type IIa | Fast | High | MYH2 | Highly oxidative |
| Type IIb | Fast | Very low | MYH4 | Highly glycolytic |
| Type IIx | Fast | Low | MYH1 | Oxidative and glycolytic |
Table 8.
Mass spectrometry-based calculated percentages of disease-related proteins and their distribution (in%) in skeletal muscle fiber types (type I, type IIa, type IIb, and type IIx). All proteins do show a higher abundance in type I and type IIa fibers.
Table 8.
Mass spectrometry-based calculated percentages of disease-related proteins and their distribution (in%) in skeletal muscle fiber types (type I, type IIa, type IIb, and type IIx). All proteins do show a higher abundance in type I and type IIa fibers.
| Protein Names | % Type I | % Type IIa | % Type IIb | % Type IIx |
|---|
| Actin. alpha skeletal muscle | 30.49 | 24.23 | 20.14 | 21.98 |
| Alpha-crystallin B chain | 0.06 | 0.06 | 0.01 | 0.02 |
| BAG family molecular chaperone regulator 3 | 0.01 | 0.01 | 0.00 | 0.01 |
| Desmin | 0.37 | 0.40 | 0.14 | 0.20 |
| Dystrophin | 0.08 | 0.07 | 0.02 | 0.02 |
| Filamin-C | 0.12 | 0.15 | 0.08 | 0.13 |
| Heat shock protein beta-8 | 0.01 | 0.01 | 0.00 | 0.00 |
| Myotilin | 0.38 | 0.41 | 0.13 | 0.25 |


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}



