
Corynebacterium diphtheriae Proteome Adaptation to Cell Culture Medium and Serum




Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria and Growth Conditions
2.2. Sample Preparation of Proteomic Analysis
2.3. Mass Spectrometry
2.4. Data Analysis
3. Results
3.1. Growth Experiments under Different Culture Conditions
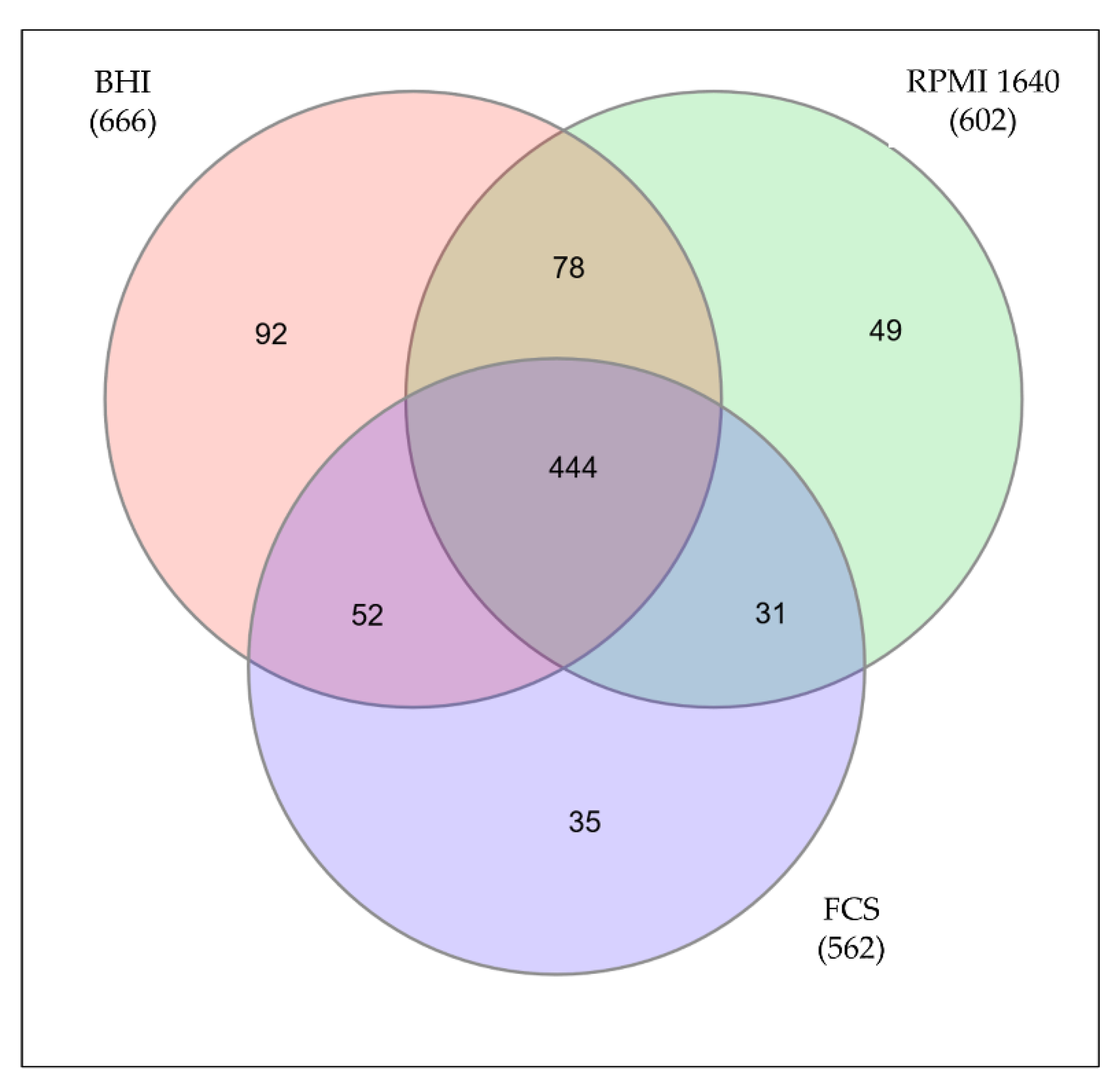
3.2. Mass Spectrometric Analysis
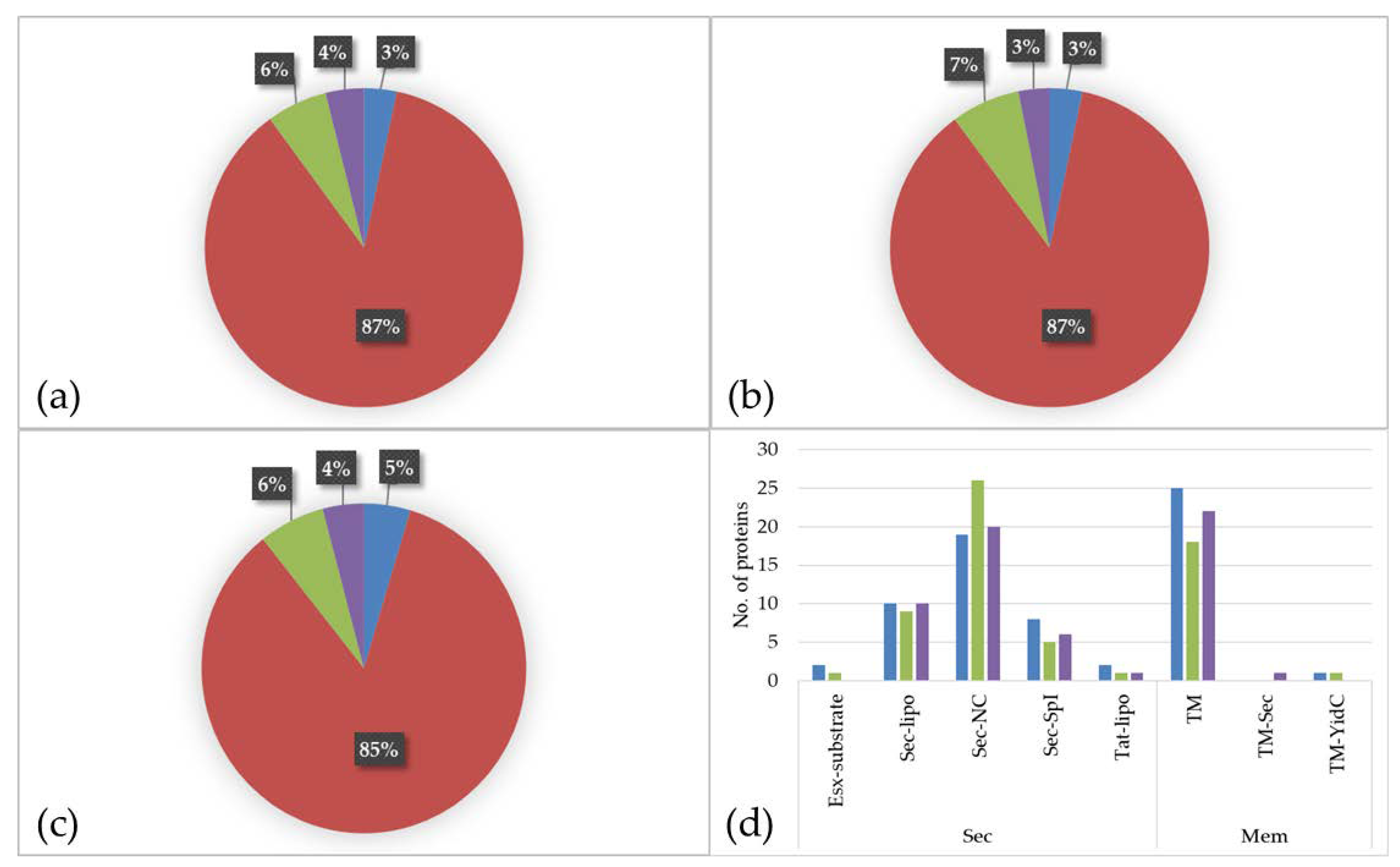
3.3. Differentially Expressed Proteins in Cell Culture Media and Serum
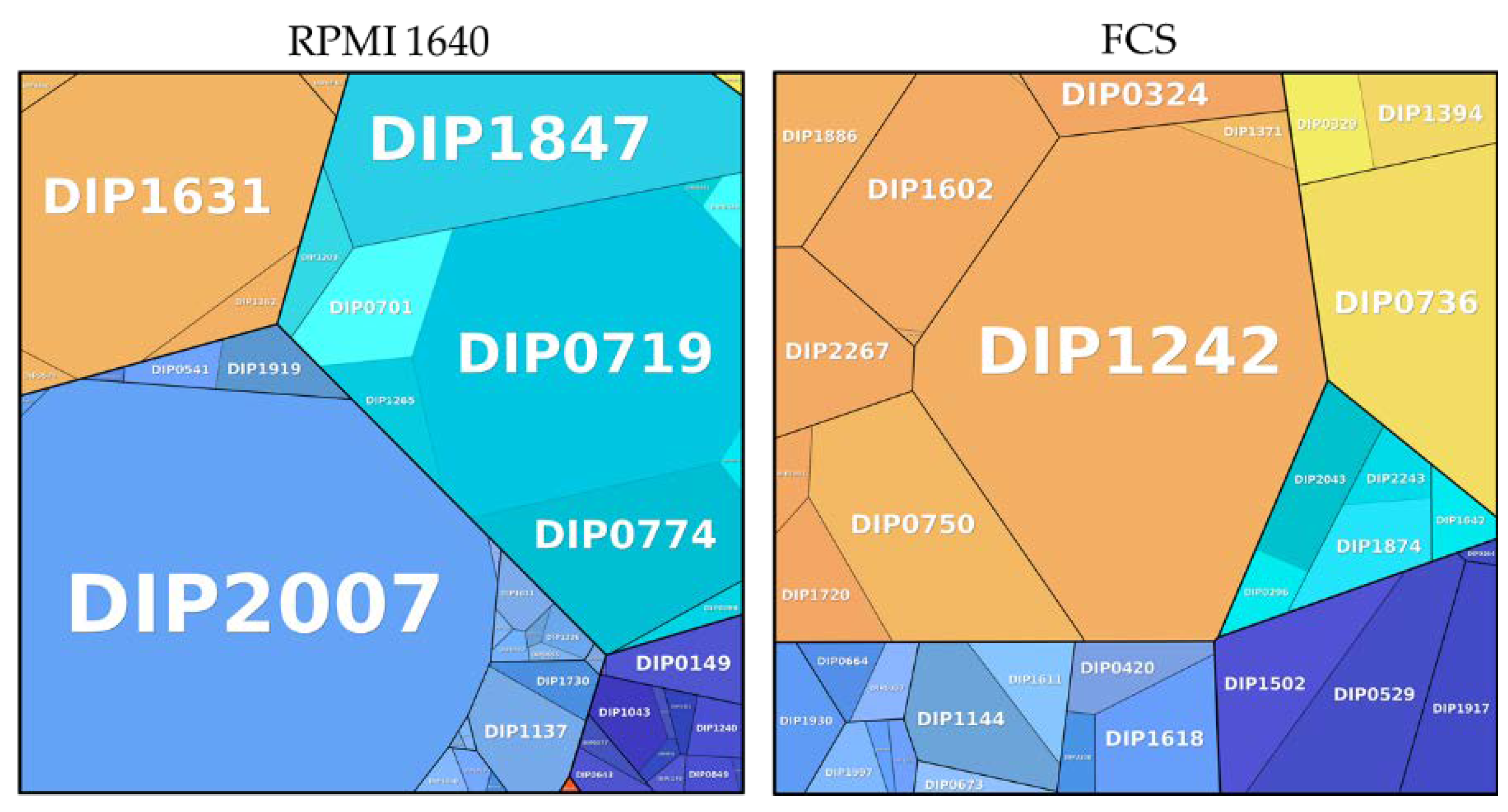
3.4. Newly Synthesised Proteins under Two Different Growth Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AGC | automatic gain control |
| BHI | brain heart infusion |
| CAA | chloroacetamide |
| CFU | colony forming units |
| DT | diphtheria toxin |
| DtxR | diphtheria toxin repressor |
| FCS | fetal calf serum |
| FDR | false discovery rate |
| GO | gene onthology |
| HPLC | High-performance liquid chromatography |
| LC-MS/MS | liquid chromatography mass spectrometry/mass spectrometry |
| OD | optical density |
| PAI | pathogenic island |
| ROS | reactive oxygen species |
| RPMI | Roswell Park Memorial Institute |
| TEAB | triethylammonium bicarbonate |
| TFA | trifluoroacetic acid |
References
- Ventura, M.; Canchaya, C.; Tauch, A.; Chandra, G.; Fitzgerald, G.F.; Chater, K.F.; van Sinderen, D. Genomics of Actinobacteria: Tracing the evolutionary history of an ancient phylum. Microbiol. Mol. Biol. Rev. 2007, 71, 495–548. [Google Scholar] [CrossRef]
- Burkovski, A. Diphtheria and its etiological agents. In Corynebacterium Diphtheriae and Related Toxigenic Species; Burkovski, A., Ed.; Springer International Publishing: Dordrecht, The Netherlands, 2014; pp. 1–14. [Google Scholar]
- Sharma, N.C.; Efstratiou, A.; Mokrousov, I.; Mutreja, A.; Das, B.; Ramamurthy, T. Diphtheria. Nat. Rev. Dis. Prim. 2019, 5. [Google Scholar] [CrossRef]
- World Health Organization Diphtheria Reported Cases. Available online: https://apps.who.int/immunization_monitoring/globalsummary/timeseries/tsincidencediphtheria.html (accessed on 16 March 2020).
- Hadfield, T.L.; McEvoy, P.; Polotsky, Y.; Tzinserling, V.A.; Yakovlev, A.A. The pathology of diphtheria. J. Infect. Dis. 2000, 181, S116–S120. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Hacker, E.; Antunes, C.A.; Weerasekera, D.; Alves, A.; Oliveira, D.S.; Martins, C.A.; Júnior, R.H.; Burkovski, A.; Mattos-guaraldi, A.L. Pathogenic properties of a Corynebacterium diphtheriae strain isolated from a case of osteomyelitis. J. Med. Microbiol. 2016, 65, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Tiley, S.M.; Kociuba, K.R.; Heron, L.G.; Munro, R. Infective endocarditis due to nontoxigenic Corynebacterium diphtheriae: Report of seven cases and review. Clin. Infect. Dis. 1993, 16, 271–275. [Google Scholar] [CrossRef]
- Patey, O.; Bimet, F.; Riegel, P.; Halioua, B.; Emond, J.P.; Estrangin, E.; Dellion, S.; Alonso, J.M.; Kiredjian, M.; Dublanchet, A.; et al. Clinical and molecular study of Corynebacterium diphtheriae systemic infections in France. J. Clin. Microbiol. 1997, 35, 441–445. [Google Scholar] [CrossRef]
- Berger, A.; Teusch, B.; Heinzinger, S.; Sing, A. Corynebacterium ulcerans—ein Emerging Pathogen? Daten des Konsiliarlabors für Diphtherie 2011–2016. Epidemiol. Bull. 2018. [Google Scholar] [CrossRef]
- Puliti, M.; Von Hunolstein, C.; Marangi, M.; Bistoni, F.; Tissi, L. Experimental model of infection with non-toxigenic strains of Corynebacterium diphtheriae and development of septic arthritis. J. Med. Microbiol. 2006, 55, 229–235. [Google Scholar] [CrossRef]
- Weerasekera, D.; Fastner, T.; Lang, R.; Burkovski, A.; Ott, L. Of mice and men: Interaction of Corynebacterium diphtheriae strains with murine and human phagocytes. Virulence 2019, 10, 414–428. [Google Scholar] [CrossRef]
- Sabbadini, P.S.; Assis, M.C.; Trost, E.; Gomes, D.L.R.; Moreira, L.O.; dos Santos, C.S.; Pereira, G.A.; Nagao, P.E.; de Azevedo, V.A.; Hirata Júnior, R.; et al. Corynebacterium diphtheriae 67-72p hemagglutinin, characterized as the protein DIP0733, contributes to invasion and induction of apoptosis in HEp-2 cells. Microb. Pathog. 2012, 52, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, R.S.; Antunes, C.A.; Lourêdo, L.S.; Viana, V.G.; dos Santos, C.S.; da Silva, J.F.R.; Hirata, R.; Hacker, E.; Mattos-Guaraldi, A.L.; Burkovski, A. Functional characterization of the collagen-binding protein DIP2093 and its influence on host–pathogen interaction and arthritogenic potential of Corynebacterium diphtheriae. Microbiology 2017, 163, 692–701. [Google Scholar] [CrossRef]
- Kolodkina, V.; Denisevich, T.; Titov, L. Identification of Corynebacterium diphtheriae gene involved in adherence to epithelial cells. Infect. Genet. Evol. 2011, 11, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.S.; Antunes, C.A.; dos Santos, C.S.; Pereira, J.A.A.; Sabbadini, P.S.; de Luna, M.; Azevedo, V.; Hirata Junior, R.; Burkovski, A.; Asad, L.M.B.; et al. Corynebacterium diphtheriae putative tellurite-resistance protein (CDCE8392_0813) contributes to the intracellular survival in human epithelial cells and lethality of Caenorhabditis elegans. Mem. Inst. Oswaldo Cruz 2015, 110, 662–668. [Google Scholar] [CrossRef]
- Weerasekera, D.; Hahn, J.; Herrmann, M.; Burkovski, A. Live cell imaging of macrophage/bacterium interaction demonstrates cell lysis induced by Corynebacterium diphtheriae and Corynebacterium ulcerans. BMC Res. Notes 2019, 12, 19–21. [Google Scholar] [CrossRef]
- Antunes, C.A.; dos Santos, L.S.; Hacker, E.; Köhler, S.; Bösl, K.; Ott, L.; de Luna, M.G.; Hirata, R.; Azevedo, V.A.; Mattos-Guaraldi, A.L.; et al. Characterization of DIP0733, a multi-functional virulence factor of Corynebacterium diphtheriae. Microbiology 2015, 161, 639–647. [Google Scholar] [CrossRef]
- Bittel, M.; Gastiger, S.; Amin, B.; Hofmann, J.; Burkovski, A. Surface and extracellular proteome of the emerging pathogen Corynebacterium ulcerans. Proteomes 2018, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Möller, J.; Schorlemmer, S.; Hofmann, J.; Burkovski, A. Cellular and extracellular proteome of the animal pathogen Corynebacterium silvaticum, a close relative of zoonotic Corynebacterium ulcerans and Corynebacterium pseudotuberculosis. Proteomes 2020, 8, 19. [Google Scholar] [CrossRef]
- Goring, A.K.; Chen, Y.; Clubb, R.T.; Loo, J.A. Secreted and cell surface proteome analysis of pathogenic Corynebacterium diphtheriae reveals proteins relevant to virulence. bioRxiv 2020, 1–11. [Google Scholar] [CrossRef]
- Hansmeier, N.; Chao, T.C.; Kalinowski, J.; Pühler, A.; Tauch, A. Mapping and comprehensive analysis of the extracellular and cell surface proteome of the human pathogen Corynebacterium diphtheriae. Proteomics 2006, 6, 2465–2476. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.M.; Folador, E.L.; Soares, S.C.; Souza, G.H.M.F.; Santos, A.V.; Sousa, C.S.; Figueiredo, H.; Miyoshi, A.; Le Loir, Y.; Silva, A.; et al. Label-free quantitative proteomics of Corynebacterium pseudotuberculosis isolates reveals differences between Biovars ovis and equi strains. BMC Genom. 2017, 18, 1–14. [Google Scholar] [CrossRef]
- Kraner, M.E.; Müller, C.; Sonnewald, U. Comparative proteomic profiling of the choline transporter-like1 (CHER1) mutant provides insights into plasmodesmata composition of fully developed Arabidopsis thaliana leaves. Plant. J. 2017, 92, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, W.; Eckart, R.A.; Schmid, B.; Cagköylü, H.; Hof, K.; Muller, Y.A.; Amin, B.; Lührmann, A. Nuclear trafficking of the anti-apoptotic Coxiella burnetii effector protein AnkG requires binding to p32 and Importin-α1. Cell. Microbiol. 2016, 19, 43–48. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Hein, M.Y.; Cox, J.; Mann, M. A “Proteomic Ruler” for protein copy number and concentration estimation without spike-in standards. Mol. Cell. Proteom. 2014, 13, 3497–3506. [Google Scholar] [CrossRef] [PubMed]
- Sangal, V.; Blom, J.; Sutcliffe, I.C.; von Hunolstein, C.; Burkovski, A.; Hoskisson, P.A. Adherence and invasive properties of Corynebacterium diphtheriae strains correlates with the predicted membrane-associated and secreted proteome. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Yi, Y.; Fang, Y.; Wu, K.; Liu, Y.; Zhang, W. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, J.; Funke, S.; Hecker, M.; Siebourg, J. Visualizng gene expression data via Voroni treemaps. Int. Symp. Vor. Diagrams 2009, 233–241. [Google Scholar] [CrossRef]
- Otto, A.; Bernhardt, J.; Meyer, H.; Schaffer, M.; Herbst, F.A.; Siebourg, J.; Mäder, U.; Lalk, M.; Hecker, M.; Becher, D. Systems-wide temporal proteomic profiling in glucose-starved Bacillus subtilis. Nat. Commun. 2010, 1. [Google Scholar] [CrossRef]
- Liebermeister, W.; Noor, E.; Flamholz, A.; Davidi, D.; Bernhardt, J.; Milo, R. Visual account of protein investment in cellular functions. Proc. Natl. Acad. Sci. USA 2014, 111, 8488–8493. [Google Scholar] [CrossRef]
- Houben, E.N.G.; Korotkov, K.V.; Bitter, W. Take five—Type VII secretion systems of Mycobacteria. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 1707–1716. [Google Scholar] [CrossRef]
- Dover, L.G.; Cerde, A.M.; Pallen, M.J.; Parkhill, J.; Besra, G.S. Comparative cell wall core biosynthesis in the mycolated pathogens, Mycobacterium tuberculosis and Corynebacterium diphtheriae. FEMS Microbiol Rev. 2004, 28, 225–250. [Google Scholar] [CrossRef] [PubMed]
- Ott, L.; Höller, M.; Gerlach, R.G.; Hensel, M.; Rheinlaender, J.; Schäffer, T.E.; Burkovski, A. Corynebacterium diphtheriae invasion-associated protein (DIP1281) is involved in cell surface organization, adhesion and internalization in epithelial cells. BMC Microbiol. 2010, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Dalbey, R.E.; Wang, P.; Kuhn, A. Assembly of bacterial inner membrane proteins. Annu. Rev. Biochem. 2011, 80, 161–187. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.C.; Hazlitt, W. Antimicrobial reactive oxygen and nitrogen species: Concepts and controverses. Nat. Rev. Microbiol. 2004, 2, 820–832. [Google Scholar] [CrossRef]
- Zahrt, T.C.; Deretic, V. Reactive nitrogen and oxygen intermediates and bacterial defenses: Unusual adaptations in Mycobacterium tuberculosis. Antioxid. Redox Signal. 2002, 4, 141–159. [Google Scholar] [CrossRef]
- Yellaboina, S.; Ranjan, S.; Chakhaiyar, P.; Hasnain, S.E.; Ranjan, A. Prediction of DtxR regulon: Identification of binding sites and operons controlled by diphtheria toxin repressor in Corynebacterium diphtheriae. BMC Microbiol. 2004, 8, 1–8. [Google Scholar] [CrossRef]
- Martinez, A.; Kolter, R. Protection of DNA during oxidative stress by the nonspecific DNA-binding protein Dps. J. Bacteriol. 1997, 179, 5188–5194. [Google Scholar] [CrossRef]
- Merkamm, M.; Guyonvarch, A. Cloning of the sodA Gene from Corynebacterium melassecola and role of superoxide dismutase in cellular viability. J. Bacteriol. 2001, 183, 1284–1295. [Google Scholar] [CrossRef]
- Spinler, J.K.; Zajdowicz, S.L.W.; Haller, J.C.; Oram, D.M.; Ronald, E.; Holmes, R.K. Development and use of a selectable, broad-host-range reporter transposon for identifying environmentally regulated promoters in bacteria. FEMS Microbiol. Lett. 2010, 291, 143–150. [Google Scholar] [CrossRef]
- Ott, L.; Hacker, E.; Kunert, T.; Karrington, I.; Etschel, P.; Lang, R.; Wiesmann, V.; Wittenberg, T.; Singh, A.; Varela, C.; et al. Analysis of Corynebacterium diphtheriae macrophage interaction: Dispensability of corynomycolic acids for inhibition of phagolysosome maturation and identification of a new gene involved in synthesis of the corynomycolic acid layer. PLoS ONE 2017, 12, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Duckworth, B.P.; Nelson, K.M.; Aldrich, C.C. Adenylating enzymes in Mycobacterium tuberculosis as drug targets. Curr. Top. Med. Chem. 2012, 12, 766–796. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Date, M.; Itaya, H.; Matsui, K.; Wu, L. Functional analysis of the Twin-Arginine translocation pathway in Corynebacterium glutamicum ATCC 13869. Appl. Environ. Microbiol. 2006, 72, 7183–7192. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brune, I.; Werner, H.; Hüser, A.T.; Kalinowski, J.; Pühler, A.; Tauch, A. The DtxR protein acting as dual transcriptional regulator directs a global regulatory network involved in iron metabolism of Corynebacterium glutamicum. BMC Genom. 2006, 7, 1–19. [Google Scholar] [CrossRef]
- Bhat, A.H.; Pathak, D.; Rao, A. The alr-groEL1 operon in Mycobacterium tuberculosis: An interplay of multiple regulatory elements. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Hu, Y.; Coates, A.R.M. Acute and persistent Mycobacterium tuberculosis infections depend on the thiol peroxidase TPX. PLoS ONE 2009, 4, e5150. [Google Scholar] [CrossRef] [PubMed]
- Buchmeier, N.A.; Newton, G.L.; Fahey, R.C. A mycothiol synthase mutant of Mycobacterium tuberculosis has an altered thiol-disulfide content and limited tolerance to stress. J. Bacteriol. 2006, 188, 6245–6252. [Google Scholar] [CrossRef]
- Meena, L.S.; Chopra, P.; Bedwal, R.S.; Singh, Y. Nucleoside diphosphate kinase-like activity in adenylate kinase of Mycobacterium tuberculosis. Biotechnol. Appl. Biochem. 2003, 38, 169. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; De Sessions, P.F.; Teoh, G.H.K.; Mohamed, A.N.N.; Zhu, Y.O.; Koh, V.H.Q.; Ang, M.L.T.; Dedon, P.C.; Hibberd, M.L.; Alonsoa, S. Transcriptional profiling of Mycobacterium tuberculosis exposed to in vitro lysosomal stress. Infect. Immun. 2016, 84, 2505–2523. [Google Scholar] [CrossRef]
- Byun, E.H.; Kim, W.S.; Kim, J.S.; Won, C.J.; Choi, H.G.; Kim, H.J.; Cho, S.N.; Lee, K.; Zhang, T.; Hur, G.M.; et al. Mycobacterium paratuberculosis CobT activates dendritic cells via engagement of toll-like receptor 4 resulting in Th1 cell expansion. J. Biol. Chem. 2012, 287, 38609–38624. [Google Scholar] [CrossRef]
- Taniguchi, H.; Busche, T.; Patschkowski, T.; Niehaus, K.; Pátek, M.; Kalinowski, J.; Wendisch, V.F. Physiological roles of sigma factor SigD in Corynebacterium glutamicum. BMC Microbiol. 2017, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nampoothiri, K.M.; Hoischen, C.; Bathe, B.; Möckel, B.; Pfefferle, W.; Krumbach, K.; Sahm, H.; Eggeling, C. Expression of genes of lipid synthesis and altered lipid composition modulates L -glutamate efflux of Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2002, 58, 89–96. [Google Scholar] [CrossRef]
- Arora, D.; Chawla, Y.; Malakar, B.; Singh, A.; Nandicoori, V.K. The transpeptidase PbpA and noncanonical transglycosylase RodA of Mycobacterium tuberculosis play important roles in regulating bacterial cell lengths. J. Biol. Chem. 2018, 293, 6497–6516. [Google Scholar] [CrossRef]
- Kawai, Y.; Marles-Wright, J.; Cleverley, R.M.; Emmins, R.; Ishikawa, S.; Kuwano, M.; Heinz, N.; Bui, N.K.; Hoyland, C.N.; Ogasawara, N.; et al. A widespread family of bacterial cell wall assembly proteins. EMBO J. 2011, 30, 4931–4941. [Google Scholar] [CrossRef] [PubMed]
- Frunzke, J.; Ga, C.; Brocker, M.; Bott, M. Control of heme homeostasis in Corynebacterium glutamicum by the two-component system HrrSA. J. Bacteriol. 2011, 193, 1212–1221. [Google Scholar] [CrossRef]
- Bott, M.; Brocker, M. Two-component signal transduction in Corynebacterium glutamicum and other corynebacteria: On the way towards stimuli and targets. Appl. Microbiol. Biotechnol. 2012, 94, 1131–1150. [Google Scholar] [CrossRef]
- Pfanzagl, V.; Holcik, L.; Maresch, D.; Gorgone, G.; Michlits, H.; Paul, G. Coproheme decarboxylases Phylogenetic prediction versus biochemical experiments. Arch. Biochem. Biophys. 2018, 640, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Rodionova, I.A.; Schuster, B.M.; Guinn, K.M.; Sorci, L.; Scott, D.A.; Li, X.; Kheterpal, I.; Shoen, C.; Cynamon, M.; Locher, C.; et al. Metabolic and bactericidal effects of targeted suppression of NadD and NadE enzymes in mycobacteria. MBio 2014, 5, 1–9. [Google Scholar] [CrossRef]
- Jamal, S.B.; Hassan, S.S.; Tiwari, S.; Viana, M.V.; De Jesus Benevides, L.; Ullah, A.; Turjanski, A.G.; Barh, D.; Ghosh, P.; Costa, D.A.; et al. An integrative in-silico approach for therapeutic target identification in the human pathogen Corynebacterium diphtheriae. PLoS ONE 2017, 12, 1–25. [Google Scholar] [CrossRef]
- Be, N.A.; Bishai, W.R.; Jain, S.K. Role of Mycobacterium tuberculosis pknD in the pathogenesis of central nervous system tuberculosis. BMC Microbiol. 2012, 12, 7. [Google Scholar] [CrossRef]
- Yoon, S.H.; Park, Y.K.; Kim, J.F. PAIDB v2.0: Exploration and analysis of pathogenicity and resistance islands. Nucleic Acids Res. 2015, 43, D624–D630. [Google Scholar] [CrossRef]
- Soares, S.C.; Ramos, R.T.J.; Silva, W.M.; Oliveira, L.C.; Amorim, L.G.; Hirata, R.; Mattos-Guaraldi, A.L.; Miyoshi, A.; Silva, A.; Azevedo, V. Corynebacterium pathogenic species in next-generation genomic era: The use of EDGAR and PIPS software and the importance of pathogenicity islands identification in pan-genomic analyses of pathogenic species. Microb. Pathog. Strateg. Combat. Them Sci. Technol. Educ. 2013. [Google Scholar] [CrossRef]
- Hu, Y.; Morichaud, Z.; Chen, S.; Leonetti, J.P.; Brodolin, K. Mycobacterium tuberculosis RbpA protein is a new type of transcriptional activator that stabilizes the σ a-containing RNA polymerase holoenzyme. Nucleic Acids Res. 2012, 40, 6547–6557. [Google Scholar] [CrossRef] [PubMed]
- Burgos, J.; Schmitt, M. The ChrA response regulator in Corynebacterium diphtheriae controls hemin-regulated gene expression through binding to the hmuO and hrtAB promoter regions. J. Bacteriol. 2012, 194, 1717–1729. [Google Scholar] [CrossRef]
- Schmitt, M. Iron acquisition and iron-dependent gene expression in Corynebacterium diphtheriae. In Corynebacterium diphtheriae and Related Toxigenic Species: Genomics, Pathogenicity and Applications; Springer: Berlin/Heidelberg, Germany, 2014; pp. 95–121. ISBN 978-94-007-7623-4. [Google Scholar]
- Lechardeur, D.; Fernandez, A.; Robert, B.; Gaudu, P.; Trieu-Cuot, P.; Lamberet, G.; Gruss, A. The 2-Cys peroxiredoxin alkyl hydroperoxide reductase C binds heme and participates in its intracellular availability in Streptococcus agalactiae. J. Biol. Chem. 2010, 285, 16032–16041. [Google Scholar] [CrossRef] [PubMed]
- Anzaldi, L.L.; Skaar, E.P. Overcoming the heme paradox: Heme toxicity and tolerance in bacterial pathogens. Infect. Immun. 2010, 78, 4977–4989. [Google Scholar] [CrossRef]
- Qi, Z.; Hamza, I.; O’Brian, M.R. Heme is an effector molecule for iron-dependent degradation of the bacterial iron response regulator (Irr) protein. Proc. Natl. Acad. Sci. USA 1999, 96, 13056–13061. [Google Scholar] [CrossRef]
- Choby, J.E.; Skaar, E.P. Heme synthesis and acquisition in bacterial pathogens. J. Mol. Biol. 2016, 428, 3408–3428. [Google Scholar] [CrossRef] [PubMed]
- Ott, L.; Burkovski, A. Toxigenic corynebacteria: Adhesion, invasion and host response. In Corynebacterium diphtheriae and Related Toxigenic Species: Genomics, Pathogenicity and Applications; Springer: Berlin/Heidelberg, Germany, 2014; Volume 9789400776, pp. 1–293. ISBN 9789400776241. [Google Scholar]
- Nathan, C.; Shiloh, M.U. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci. USA 2000, 97, 8841–8848. [Google Scholar] [CrossRef]
- Tsolis, R.M.; Baumler, A.J.; Heffron, F. Role of Salmonella typhimurium mn-superoxide dismutase (SodA) in protection against early killing by J774 macrophages. Infect. Immun. 1995, 63, 1739–1744. [Google Scholar] [CrossRef]
- Colombo, A.V.; Hirata, R.; De Souza, C.M.R.; Monteiro-Leal, L.H.; Previato, J.O.; Formiga, L.C.D.; Andrade, A.F.B.; Mattos-Guaraldi, A.L. Corynebacterium diphtheriae surface proteins as adhesins to human erythrocytes. FEMS Microbiol. Lett. 2001, 197, 235–239. [Google Scholar] [CrossRef][Green Version]
- Hirata, R.; Souza, S.M.S.; Rocha-de-Souza, C.M.; Andrade, A.F.B.; Monteiro-Leal, L.H.; Formiga, L.C.D.; Mattos-Guaraldi, A.L. Patterns of adherence to HEp-2 cells and actin polymerisation by toxigenic Corynebacterium diphtheriae strains. Microb. Pathog. 2004, 36, 125–130. [Google Scholar] [CrossRef]
- Ott, L.; Mckenzie, A.; Baltazar, M.T.; Britting, S.; Bischof, A.; Burkovski, A.; Hoskisson, P.A. Evaluation of invertebrate infection models for pathogenic corynebacteria. FEMS Immunol. Med. Microbiol. 2012, 65, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Kana, B.D.; Mizrahi, V. Resuscitation-promoting factors as lytic enzymes for bacterial growth and signaling. FEMS Immunol. Med. Microbiol. 2010, 58, 39–50. [Google Scholar] [CrossRef]
- Rosser, A.; Stover, C.; Pareek, M.; Mukamolova, G.V. Resuscitation-promoting factors are important determinants of the pathophysiology in Mycobacterium tuberculosis infection. Crit. Rev. Microbiol. 2017, 43, 621–630. [Google Scholar] [CrossRef]
- Dobrindt, U.; Hochhut, B.; Hentschel, U.; Hacker, J. Genomic islands in pathogenic and environmental microorganisms. Nat. Rev. Microbiol. 2004, 2, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Trost, E.; Tauch, A. Comparative genomics and pathogenicity islands of Corynebacterium diphtheriae, Corynebacterium ulcerans, and Corynebacterium pseudotuberculosis. In Corynebacterium diphtheriae and Related Toxigenic Species: Genomics, Pathogenicity and Applications; Burkovski, A., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 39–65. ISBN 978-94-007-7624-1. [Google Scholar]
- Hacker, E.; Antunes, C.A.; Mattos-guaraldi, A.L.; Burkovski, A.; Tauch, A. Corynebacterium ulcerans, an emerging human pathogen. Future Microbiol. 2016, 11, 1191–1208. [Google Scholar] [CrossRef] [PubMed]
- Perez-riverol, Y.; Csordas, A.; Bai, J.; Bernal-llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, 442–450. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | RPMI 1640 | FCS |
|---|---|---|
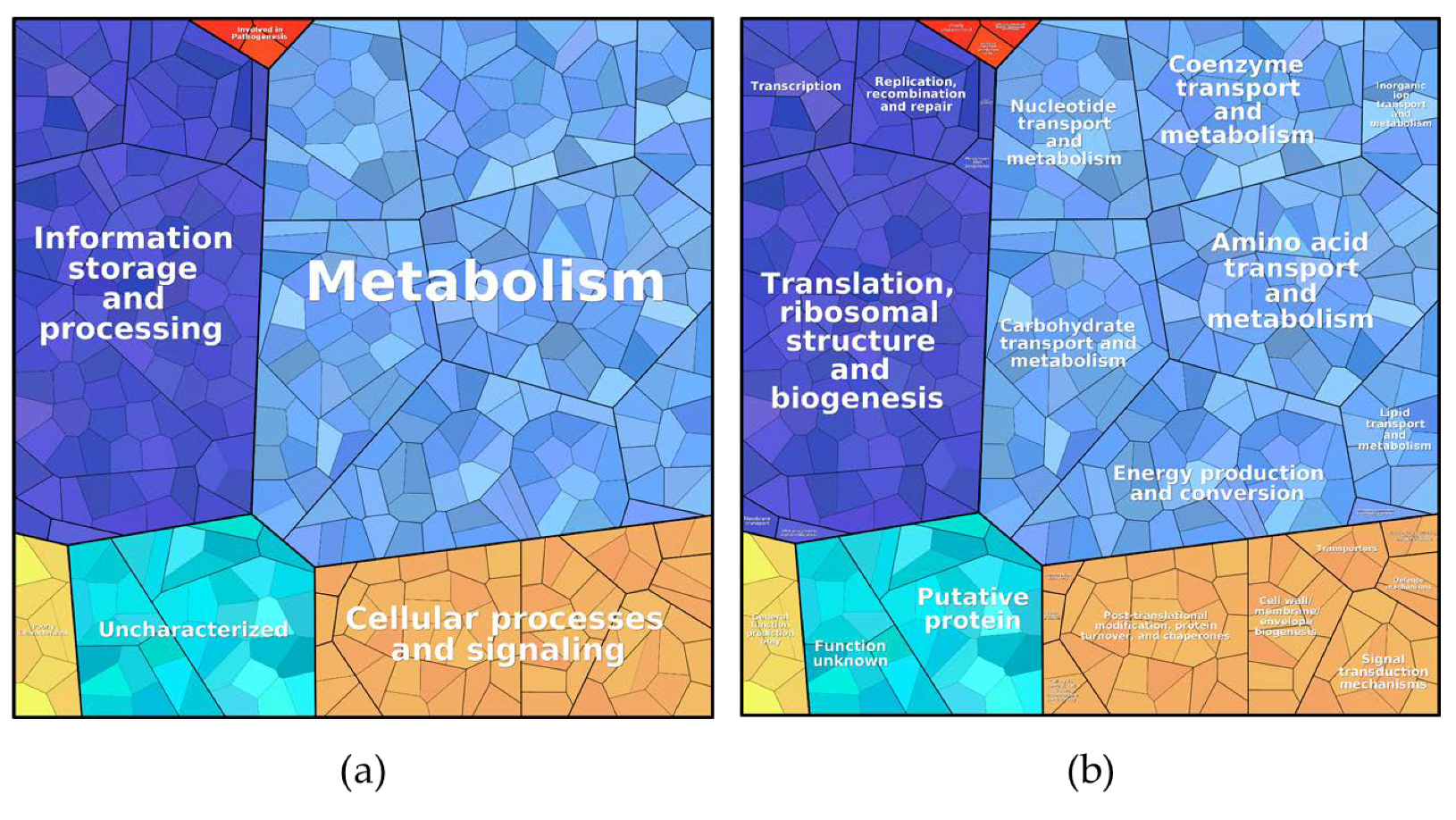
| Cellular processes and signaling | 15.94 | 56.65 |
| Information storage and processing | 4.65 | 10.12 |
| Involved in pathogenesis | 0.03 | - |
| Metabolism | 40.00 | 12.90 |
| Poorly characterized | 0.08 | 14.07 |
| Uncharacterized | 39.31 | 6.24 |
| Metabolic Function | Pathway | RPMI 1640 | FCS |
|---|---|---|---|
| Cell wall/membrane/envelope biogenesis | C | 0.24 | 6.88 |
| Cellular community-prokaryotes | C | - | 2.41 |
| Intracellular trafficking, secretion, and vesicular transport | C | - | 30.73 |
| Post-translational modification, protein turnover, and chaperones | C | 15.50 | 10.71 |
| Transporters | C | 0.20 | - |
| Signal transduction mechanisms | C | - | 3.23 |
| Defense mechanisms | C | - | 2.70 |
| Translation, ribosomal structure, and biogenesis | I | 2.52 | 7.49 |
| Replication, recombination, and repair | I | 1.50 | 0.10 |
| Transcription | I | 0.62 | - |
| Transfer RNA biogenesis | I | - | 2.53 |
| Cell wall/membrane/envelope biogenesis | P | 0.24 | 6.88 |
| Amino acid transport and metabolism | M | 0.06 | 4.04 |
| Coenzyme transport and metabolism | M | 0.50 | 1.13 |
| Nucleotide transport and metabolism | M | 1.83 | 1.26 |
| Inorganic ion transport and metabolism | M | 33.77 | 1.34 |
| Carbohydrate transport and metabolism | M | 1.23 | - |
| Lipid transport and metabolism | M | 2.53 | 4.64 |
| Accession No. | GO | RPMI [%] | FCS [%] | Description |
|---|---|---|---|---|
| Q6NJ28 | DIP0585 | 0.001 | 0.011 | Putative iron transport system ATP-binding protein |
| Q6NJ19 | DIP0597 | 0.056 | 0.108 | Uncharacterized protein |
| Q6NJ16 | DIP0600 | 0.007 | 0.039 | Putative two component system response regulator |
| Q6NIF4 | DIP0823 | 0.081 | 0.205 | Glutaredoxin domain-containing protein |
| Q6NHP5 | DIP1086 | 0.004 | 0.006 | Putative iron transport system exported solute-binding component |
| Q6NHB5 | DIP1225 | 0.079 | 0.003 | RNA polymerase-binding protein RbpA |
| Q6NEE8 | DIP2327 | 0.019 | 0.006 | Two-component response regulator ChrA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Möller, J.; Nosratabadi, F.; Musella, L.; Hofmann, J.; Burkovski, A. Corynebacterium diphtheriae Proteome Adaptation to Cell Culture Medium and Serum. Proteomes 2021, 9, 14. https://doi.org/10.3390/proteomes9010014
Möller J, Nosratabadi F, Musella L, Hofmann J, Burkovski A. Corynebacterium diphtheriae Proteome Adaptation to Cell Culture Medium and Serum. Proteomes. 2021; 9(1):14. https://doi.org/10.3390/proteomes9010014
Chicago/Turabian StyleMöller, Jens, Fatemeh Nosratabadi, Luca Musella, Jörg Hofmann, and Andreas Burkovski. 2021. "Corynebacterium diphtheriae Proteome Adaptation to Cell Culture Medium and Serum" Proteomes 9, no. 1: 14. https://doi.org/10.3390/proteomes9010014
APA StyleMöller, J., Nosratabadi, F., Musella, L., Hofmann, J., & Burkovski, A. (2021). Corynebacterium diphtheriae Proteome Adaptation to Cell Culture Medium and Serum. Proteomes, 9(1), 14. https://doi.org/10.3390/proteomes9010014