Diurnal Differences in Human Muscle Isometric Force In Vivo Are Associated with Differential Phosphorylation of Sarcomeric M-Band Proteins

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
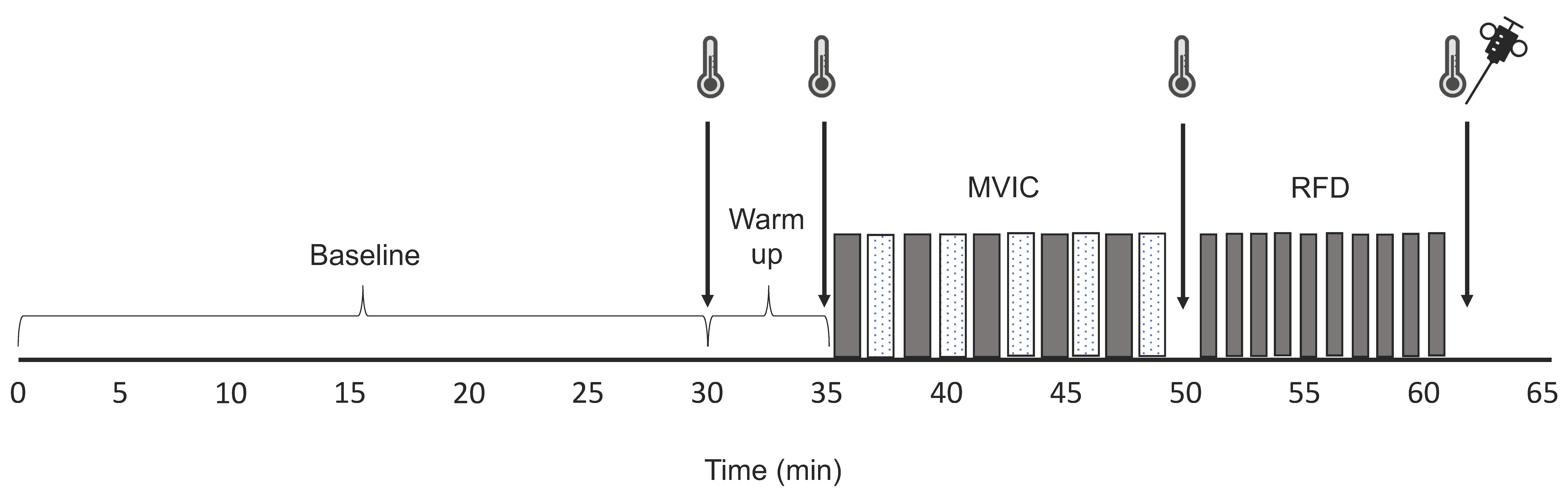
2. Materials and Methods
3. Results
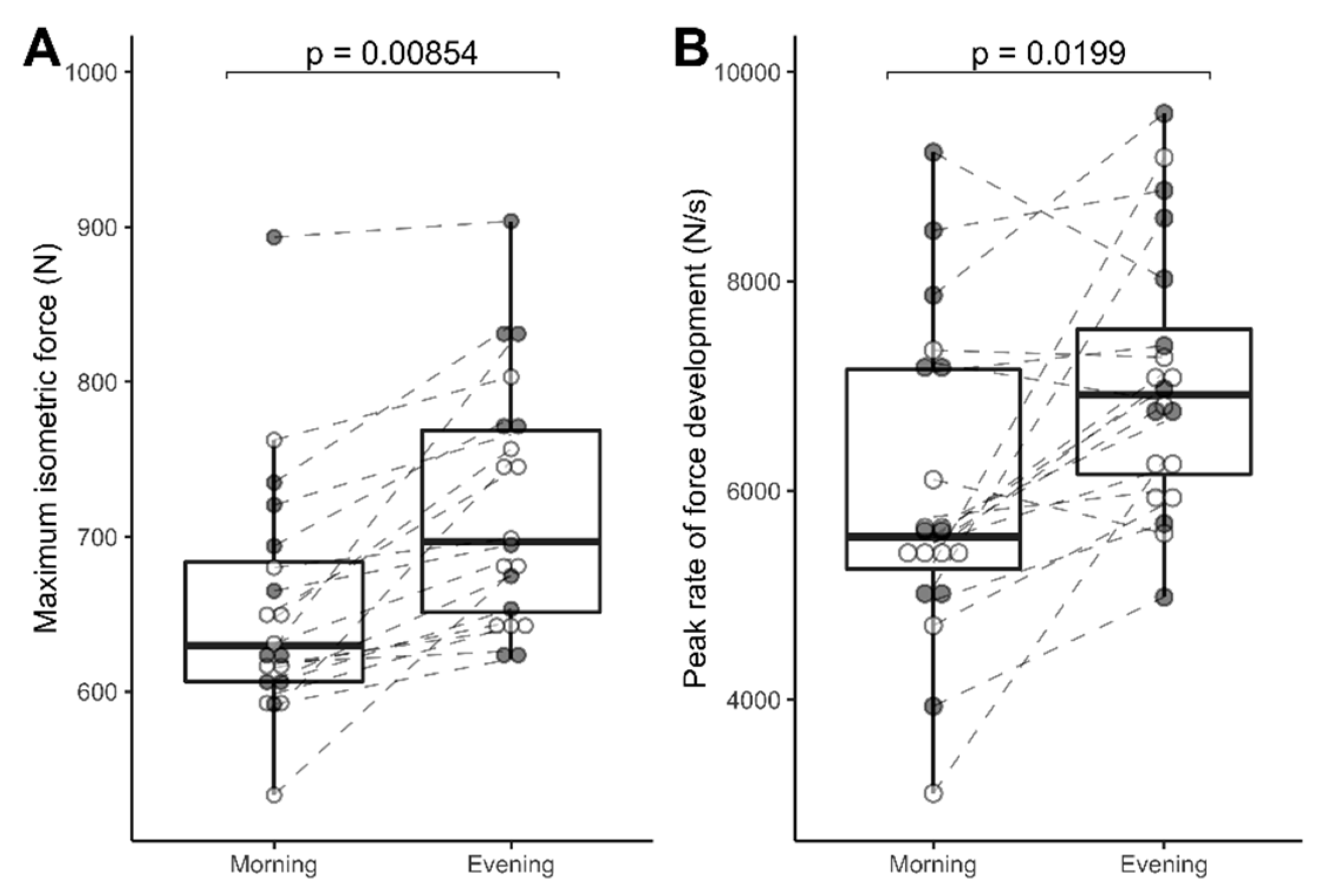
3.1. Reliability and Diurnal Variation Data: MVIC, % Activation, RFD, and Subjects Rated Maximal Effort in n = 20 Subjects
3.2. Data Handling: 10 Subjects from the Cohort of 20 Volunteered for Muscle Biopsy Procedures; Hence, Only Their Data (Temperature, MVIC, and RFD) Are Reported below to Align with the Proteomic Analyses
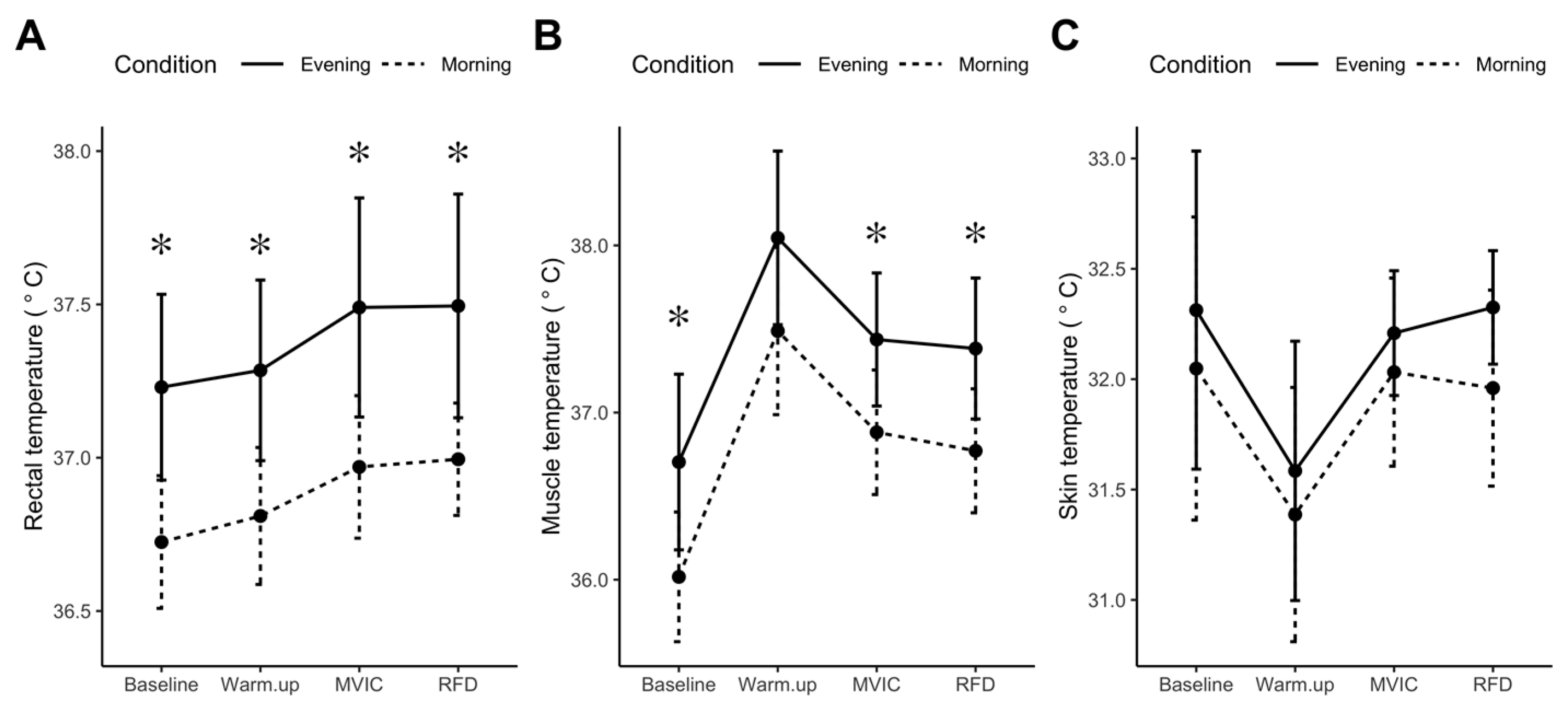
3.2.1. Resting Rectal, Skin, and Muscle Temperatures (n = 10)
3.2.2. MVIC, % Activation, Peak RFD, and Subject-Rated Maximal Effort (n = 10)
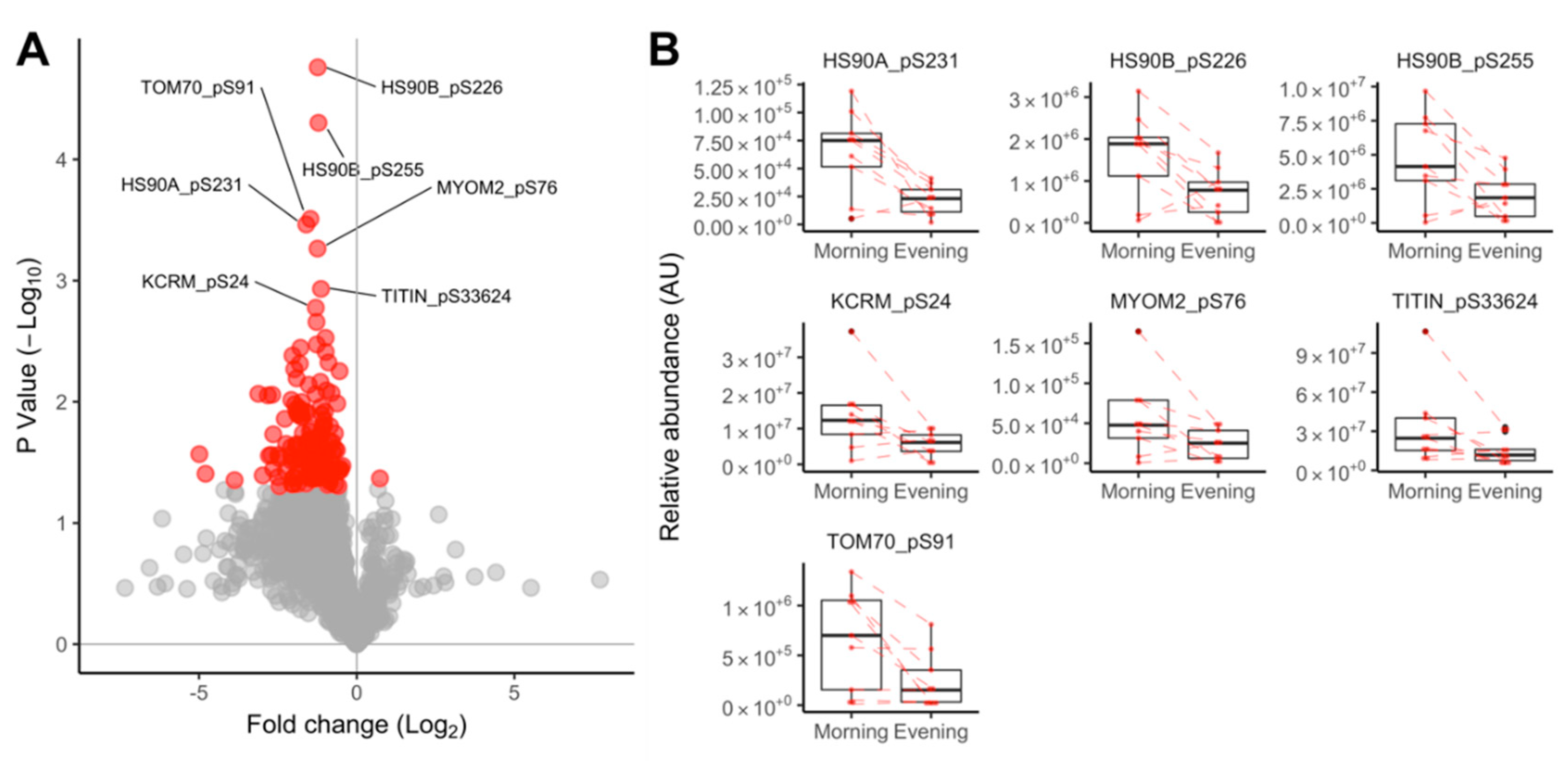
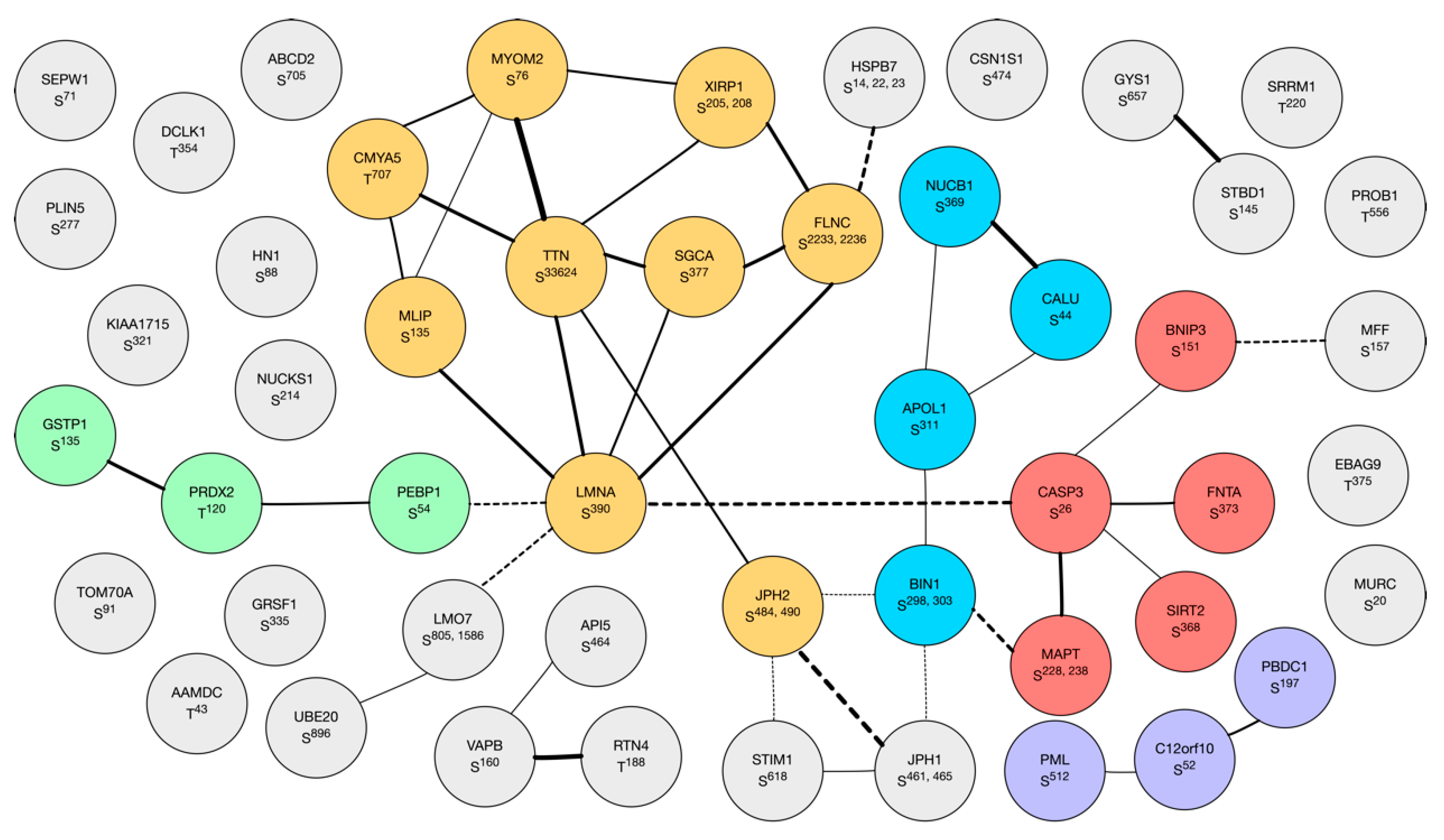
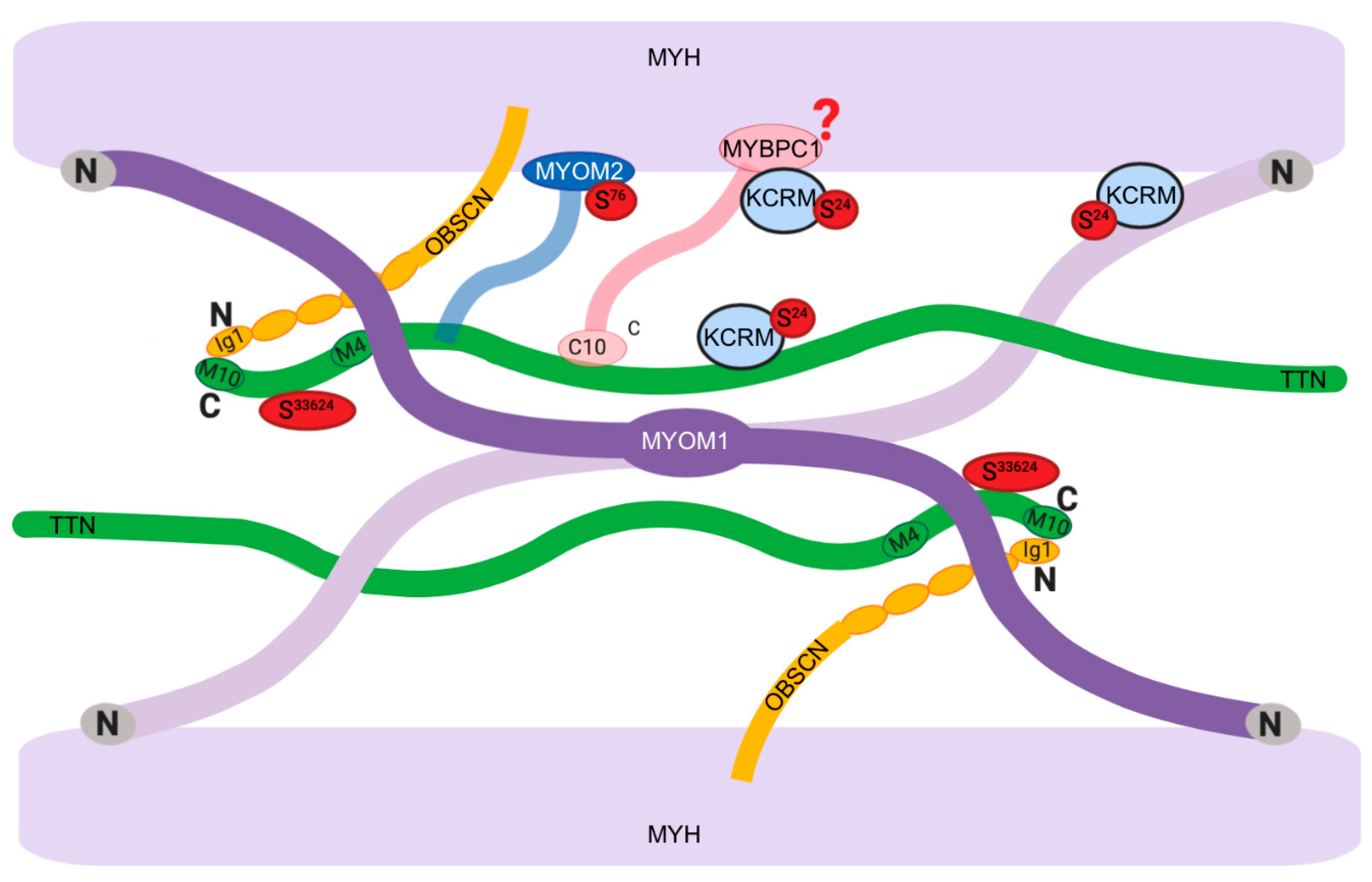
3.2.3. Proteomic Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- McLeod, M.; Breen, L.; Hamilton, D.L.; Philp, A. Live strong and prosper: The importance of skeletal muscle strength for healthy ageing. Biogerontology 2016, 17, 497–510. [Google Scholar] [CrossRef]
- Foldvari, M.; Clark, M.; Laviolette, L.C.; Bernstein, M.A.; Kaliton, D.; Castaneda, C.; Pu, C.T.; Hausdorff, J.M.; Fielding, R.A.; Singh, M.A.F. Association of Muscle Power With Functional Status in Community-Dwelling Elderly Women. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2000, 55, M192–M199. [Google Scholar] [CrossRef] [PubMed]
- Solaro, R.J.; De Tombe, P.P. Review focus series: Sarcomeric proteins as key elements in integrated control of cardiac function. Cardiovasc. Res. 2008, 77, 616–618. [Google Scholar] [CrossRef] [PubMed]
- Drust, B.; Waterhouse, J.; Atkinson, G.; Edwards, B.; Reilly, T. Circadian Rhythms in Sports Performance—An Update. Chronobiol. Int. 2005, 22, 21–44. [Google Scholar] [CrossRef] [PubMed]
- Reilly, T.; Waterhouse, J. Sports performance: Is there evidence that the body clock plays a role? Eur. J. Appl. Physiol. 2009, 106, 321–332. [Google Scholar] [CrossRef]
- Martin, A.; Carpentier, A.; Guissard, N.; van Hoecke, J.; Duchateau, J. Effect of time of day on force variation in a human muscle. Muscle Nerve 1999, 22, 1380–1387. [Google Scholar] [CrossRef]
- Robinson, W.R.; Pullinger, S.A.; Kerry, J.W.; Giacomoni, M.; Robertson, C.M.; Burniston, J.G.; Waterhouse, J.M.; Edwards, B.J. Does lowering evening rectal temperature to morning levels offset the diurnal variation in muscle force production? Chronobiol. Int. 2013, 30, 998–1010. [Google Scholar] [CrossRef]
- Erskine, R.M.; Fletcher, G.; Folland, J.P. The contribution of muscle hypertrophy to strength changes following resistance training. Eur. J. Appl. Physiol. 2014, 114, 1239–1249. [Google Scholar] [CrossRef]
- Sedliak, M.; Finni, T.; Cheng, S.; Haikarainen, T.; Häkkinen, K. Diurnal Variation in Maximal and Submaximal Strength, Power and Neural Activation of Leg Extensors in Men: Multiple Sampling Across Two Consecutive Days. Int. J. Sports Med. 2008, 29, 217–224. [Google Scholar] [CrossRef]
- Edwards, B.J.; Pullinger, S.A.; Kerry, J.W.; Robinson, W.R.; Reilly, T.P.; Robertson, C.M.; Waterhouse, J.M. Does raising morning rectal temperature to evening levels offset the diurnal variation in muscle force production? Chronobiol. Int. 2013, 30, 486–501. [Google Scholar] [CrossRef]
- Albrecht, U. Timing to Perfection: The Biology of Central and Peripheral Circadian Clocks. Neuron 2012, 74, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Zambon, A.C.; McDearmon, E.L.; Salomonis, N.; Vranizan, K.M.; Johansen, K.L.; Adey, D.; Takahashi, J.S.; Schambelan, M.; Conklin, B.R. Time- and exercise-dependent gene regulation in human skeletal muscle. Genome Biol. 2003, 4, R61. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.J.; Andrews, J.L.; McDearmon, E.L.; Campbell, K.S.; Barber, B.K.; Miller, B.H.; Walker, J.R.; Hogenesch, J.B.; Takahashi, J.S.; Esser, K.A. Identification of the circadian transcriptome in adult mouse skeletal muscle. Physiol. Genomics 2007, 31, 86–95. [Google Scholar] [CrossRef]
- Dyar, K.A.; Ciciliot, S.; Wright, L.E.; Biensø, R.S.; Tagliazucchi, G.M.; Patel, V.R.; Forcato, M.; Paz, M.I.P.; Gudiksen, A.; Solagna, F.; et al. Muscle insulin sensitivity and glucose metabolism are controlled by the intrinsic muscle clock. Mol. Metab. 2014, 3, 29–41. [Google Scholar] [CrossRef]
- Ezagouri, S.; Zwighaft, Z.; Sobel, J.; Baillieul, S.; Doutreleau, S.; Ladeuix, B.; Golik, M.; Verges, S.; Asher, G. Physiological and Molecular Dissection of Daily Variance in Exercise Capacity. Cell Metab. 2019, 30, 78–91.e4. [Google Scholar] [CrossRef]
- Andrews, J.L.; Zhang, X.; McCarthy, J.J.; McDearmon, E.L.; Hornberger, T.A.; Russell, B.; Campbell, K.S.; Arbogast, S.; Reid, M.B.; Walker, J.R.; et al. CLOCK and BMAL1 regulate MyoD and are necessary for maintenance of skeletal muscle phenotype and function. Proc. Natl. Acad. Sci. USA 2010, 107, 19090–19095. [Google Scholar] [CrossRef]
- Schroder, E.A.; Harfmann, B.D.; Zhang, X.; Srikuea, R.; England, J.H.; Hodge, B.A.; Wen, Y.; Riley, L.A.; Yu, Q.; Christie, A.; et al. Intrinsic muscle clock is necessary for musculoskeletal health. J. Physiol. 2015, 593, 5387–5404. [Google Scholar] [CrossRef]
- Camera, D.M.; Burniston, J.G.; Pogson, M.A.; Smiles, W.J.; Hawley, J.A. Dynamic proteome profiling of individual proteins in human skeletal muscle after a high-fat diet and resistance exercise. FASEB J. 2017, 31, 5478–5494. [Google Scholar] [CrossRef]
- Ryder, J.W.; Lau, K.S.; Kamm, K.E.; Stull, J.T. Enhanced skeletal muscle contraction with myosin light chain phosphorylation by a calmodulin-sensing kinase. J. Biol. Chem. 2007, 282, 20447–20454. [Google Scholar] [CrossRef]
- Potts, G.K.; McNally, R.M.; Blanco, R.; You, J.-S.; Hebert, A.S.; Westphall, M.S.; Coon, J.J.; Hornberger, T.A. A map of the phosphoproteomic alterations that occur after a bout of maximal-intensity contractions. J. Physiol. 2017, 595, 5209–5226. [Google Scholar] [CrossRef]
- Kampourakis, T.; Sun, Y.; Irving, M. Myosin light chain phosphorylation enhances contraction of heart muscle via structural changes in both thick and thin filaments. Proc. Natl. Acad. Sci. USA 2016, 113, E3039–E3047. [Google Scholar] [CrossRef] [PubMed]
- Burniston, J.G.; Hoffman, E.P. Proteomic responses of skeletal and cardiac muscle to exercise. Expert Rev. Proteom. 2011, 8, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Burniston, J.G. How many phosphoproteins does it take to make muscle grow? J. Physiol. 2017, 595, 5009–5010. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.R.; Thingholm, T.E.; Jensen, O.N.; Roepstorff, P.; Jørgensen, T.J.D. Highly Selective Enrichment of Phosphorylated Peptides from Peptide Mixtures Using Titanium Dioxide Microcolumns. Mol. Cell. Proteom. 2005, 4, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Højlund, K.; Bowen, B.P.; Hwang, H.; Flynn, C.R.; Madireddy, L.; Geetha, T.; Langlais, P.; Meyer, C.; Mandarino, L.J.; Yi, Z. In vivo phosphoproteome of human skeletal muscle revealed by phosphopeptide enrichment and HPLC-ESI-MS/MS. J. Proteome Res. 2009, 8, 4954–4965. [Google Scholar] [CrossRef]
- Hoffman, N.J.; Parker, B.L.; Chaudhuri, R.; Fisher-Wellman, K.H.; Kleinert, M.; Humphrey, S.J.; Yang, P.; Holliday, M.; Trefely, S.; Fazakerley, D.J.; et al. Global Phosphoproteomic Analysis of Human Skeletal Muscle Reveals a Network of Exercise-Regulated Kinases and AMPK Substrates. Cell Metab. 2015, 22, 922–935. [Google Scholar] [CrossRef]
- Smith, C.S.; Reilly, C.; Midkiff, K. Evaluation of three circadian rhythm questionnaires with suggestions for an improved measure of morningness. J. Appl. Psychol. 1989, 74, 728–738. [Google Scholar] [CrossRef]
- Morton, J.P.; Atkinson, G.; MacLaren, D.P.M.; Cable, N.T.; Gilbert, G.; Broome, C.; McArdle, A.; Drust, B. Reliability of maximal muscle force and voluntary activation as markers of exercise-induced muscle damage. Eur. J. Appl. Physiol. 2005, 94, 541–548. [Google Scholar] [CrossRef]
- Murtagh, C.F.; Nulty, C.; Vanrenterghem, J.; O’Boyle, A.; Morgans, R.; Drust, B.; Erskine, R.M. The Neuromuscular Determinants of Unilateral Jump Performance in Soccer Players Are Direction-Specific. Int. J. Sports Physiol. Perform. 2018, 13, 604–611. [Google Scholar] [CrossRef]
- Burniston, J.G. Changes in the rat skeletal muscle proteome induced by moderate-intensity endurance exercise. Biochim. Biophys. Acta Proteins Proteom. 2008, 1784, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Holloway, K.V.; Gorman, M.O.; Woods, P.; Morton, J.P.; Evans, L.; Cable, N.T.; Goldspink, D.F.; Burniston, J.G. Proteomic investigation of changes in human vastus lateralis muscle in response to interval-exercise training. Proteomics 2009, 9, 5155–5174. [Google Scholar] [CrossRef] [PubMed]
- Burniston, J.G.; Kenyani, J.; Gray, D.; Guadagnin, E.; Jarman, I.H.; Cobley, J.N.; Cuthbertson, D.J.; Chen, Y.-W.; Wastling, J.M.; Lisboa, P.J.; et al. Conditional independence mapping of DIGE data reveals PDIA3 protein species as key nodes associated with muscle aerobic capacity. J. Proteom. 2014, 106, 230–245. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Isserlin, R.; Emili, A.; Burniston, J.G. Exercise-responsive phosphoproteins in the heart. J. Mol. Cell. Cardiol. 2017, 111, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; Von Mering, C.; et al. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, 808–815. [Google Scholar] [CrossRef]
- Wolff, C.A.; Esser, K.A. Exercise timing and circadian rhythms. Curr. Opin. Physiol. 2019, 10, 64–69. [Google Scholar] [CrossRef]
- Lange, S.; Pinotsis, N.; Agarkova, I.; Ehler, E. The M-band: The underestimated part of the sarcomere. Biochim. Biophys. Acta Mol. Cell Res. 2019. [Google Scholar] [CrossRef]
- Pernigo, S.; Fukuzawa, A.; Beedle, A.E.M.; Holt, M.; Round, A.; Pandini, A.; Garcia-Manyes, S.; Gautel, M.; Steiner, R.A. Binding of Myomesin to Obscurin-Like-1 at the Muscle M-Band Provides a Strategy for Isoform-Specific Mechanical Protection. Structure 2017. [Google Scholar] [CrossRef]
- Lange, S.; Himmel, M.; Auerbach, D.; Agarkova, I.; Hayess, K.; Fürst, D.O.; Perriard, J.C.; Ehler, E. Dimerisation of myomesin: Implications for the structure of the sarcomeric M-band. J. Mol. Biol. 2005, 345, 289–298. [Google Scholar] [CrossRef]
- Mayans, O.; van der Ven, P.F.M.; Wilm, M.; Mues, A.; Young, P.; Fürst, D.O.; Wilmanns, M.; Gautel, M. Structural basis for activation of the titin kinase domain during myofibrillogenesis. Nature 1998, 395, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Obermann, W.M.J.; Gautel, M.; Weber, K.; Fürst, D.O. Molecular structure of the sarcomeric M band: Mapping of titin and myosin binding domains in myomesin and the identification of a potential regulatory phosphorylation site in myomesin. EMBO J. 1997. [Google Scholar] [CrossRef] [PubMed]
- Obermann, W.M.J.; van der Ven, P.F.M.; Steiner, F.; Weber, K.; Fürst, D.O. Mapping of a Myosin-binding Domain and a Regulatory Phosphorylation Site in M-Protein, a Structural Protein of the Sarcomeric M Band. Mol. Biol. Cell 1998, 9, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.A.; Hu, L.Y.R.; Bowman, A.L.; Bloch, R.J.; Kontrogianni-Konstantopoulos, A. Obscurin interacts with a novel isoform of MyBP-C slow at the periphery of the sarcomeric M-band and regulates thick filament assembly. Mol. Biol. Cell 2009. [Google Scholar] [CrossRef] [PubMed]
- Podobed, P.; Pyle, W.G.; Ackloo, S.; Alibhai, F.J.; Tsimakouridze, E.V.; Ratcliffe, W.F.; Mackay, A.; Simpson, J.; Wright, D.C.; Kirby, G.M.; et al. The day/night proteome in the murine heart. Am. J. Physiol. Integr. Comp. Physiol. 2014, 307, R121–R137. [Google Scholar] [CrossRef]
- Hornemann, T.; Kempa, S.; Himmel, M.; Hayeß, K.; Fürst, D.O.; Wallimann, T. Muscle-type Creatine Kinase Interacts with Central Domains of the M-band Proteins Myomesin and M-protein. J. Mol. Biol. 2003, 332, 877–887. [Google Scholar] [CrossRef]
- Carlsson, E.; Grove, B.K.; Wallimann, T.; Eppenberger, H.M.; Thornell, L.E. Myofibrillar M-band proteins in rat skeletal muscles during development. Histochemistry 1990, 95, 27–35. [Google Scholar] [CrossRef]
- Puchner, E.M.; Alexandrovich, A.; Kho, A.L.; Hensen, U.; Schafer, L.V.; Brandmeier, B.; Grater, F.; Grubmuller, H.; Gaub, H.E.; Gautel, M. Mechanoenzymatics of titin kinase. Proc. Natl. Acad. Sci. USA 2008, 105, 13385–13390. [Google Scholar] [CrossRef]
- Lange, S. The Kinase Domain of Titin Controls Muscle Gene Expression and Protein Turnover. Science 2005, 308, 1599–1603. [Google Scholar] [CrossRef]
- McGrath, M.J.; Cottle, D.L.; Nguyen, M.-A.; Dyson, J.M.; Coghill, I.D.; Robinson, P.A.; Holdsworth, M.; Cowling, B.S.; Hardeman, E.C.; Mitchell, C.A.; et al. Four and a Half LIM Protein 1 Binds Myosin-binding Protein C and Regulates Myosin Filament Formation and Sarcomere Assembly. J. Biol. Chem. 2006, 281, 7666–7683. [Google Scholar] [CrossRef]
- Gresham, K.S.; Stelzer, J.E. The contributions of cardiac myosin binding protein C and troponin I phosphorylation to β-adrenergic enhancement of in vivo cardiac function. J. Physiol. 2016, 594, 669–686. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.A.; Kontrogianni-Konstantopoulos, A. Myosin Binding Protein-C Slow is a Novel Substrate for Protein Kinase A (PKA) and C (PKC) in Skeletal Muscle. J. Proteome Res. 2011, 10, 4547–4555. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, T.-J.; Li, J.; Gao, Y.-S.; Meng, F.-G.; Yan, Y.-B.; Zhou, H.-M. Slow skeletal muscle myosin-binding protein-C (MyBPC1) mediates recruitment of muscle-type creatine kinase (CK) to myosin. Biochem. J. 2011, 436, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Srikakulam, R.; Winkelmann, D.A. Chaperone-mediated folding and assembly of myosin in striated muscle. J. Cell Sci. 2004. [Google Scholar] [CrossRef]
- Ojima, K.; Ichimura, E.; Suzuki, T.; Oe, M.; Muroya, S.; Nishimura, T. HSP90 modulates the myosin replacement rate in myofibrils. Am. J. Physiol. Physiol. 2018, 315, C104–C114. [Google Scholar] [CrossRef]
- Li, J.; Soroka, J.; Buchner, J. The Hsp90 chaperone machinery: Conformational dynamics and regulation by co-chaperones. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 624–635. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Akashi, M.; Matsuda, M.; Goto, K.; Miyata, Y.; Node, K.; Nishida, E. Involvement of the Protein Kinase CK2 in the Regulation of Mammalian Circadian Rhythms. Sci. Signal. 2009, 2, ra26. [Google Scholar] [CrossRef]
- Robles, M.S.; Humphrey, S.J.; Mann, M. Phosphorylation Is a Central Mechanism for Circadian Control of Metabolism and Physiology. Cell Metab. 2017, 25, 118–127. [Google Scholar] [CrossRef]
- Schmidt, O.; Harbauer, A.B.; Rao, S.; Eyrich, B.; Zahedi, R.P.; Stojanovski, D.; Schönfisch, B.; Guiard, B.; Sickmann, A.; Pfanner, N.; et al. Regulation of Mitochondrial Protein Import by Cytosolic Kinases. Cell 2011, 144, 227–239. [Google Scholar] [CrossRef]
- Tahara, Y.; Aoyama, S.; Shibata, S. The mammalian circadian clock and its entrainment by stress and exercise. J. Physiol. Sci. 2017, 67, 1–10. [Google Scholar] [CrossRef]
- Oster, H.; Challet, E.; Ott, V.; Arvat, E.; de Kloet, E.R.; Dijk, D.-J.; Lightman, S.; Vgontzas, A.; Van Cauter, E. The functional and clinical significance of the 24-h rhythm of circulating glucocorticoids. Endocr. Rev. 2017, 38, 3–45. [Google Scholar] [CrossRef] [PubMed]
- Terazono, H.; Mutoh, T.; Yamaguchi, S.; Kobayashi, M.; Akiyama, M.; Udo, R.; Ohdo, S.; Okamura, H.; Shibata, S. Adrenergic regulation of clock gene expression in mouse liver. Proc. Natl. Acad. Sci. USA 2003, 100, 6795–6800. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.J.L.; Hu, K.; Evoniuk, H.; Kelly, E.E.; Malhotra, A.; Hilton, M.F.; Shea, S.A. Impact of the human circadian system, exercise, and their interaction on cardiovascular function. Proc. Natl. Acad. Sci. USA 2010, 107, 20541–20546. [Google Scholar] [CrossRef] [PubMed]
- Davies, A. Regulation of Beta-Adrenergic Receptors by Steroid Hormones. Annu. Rev. Physiol. 1984, 46, 119–130. [Google Scholar] [CrossRef]
- Cairns, S.P.; Borrani, F. β-Adrenergic modulation of skeletal muscle contraction: Key role of excitation-contraction coupling. J. Physiol. 2015, 593, 4713–4727. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Ma, Y.; Moore, M.; Hemmings, B.A.; Taylor, S.S. Phosphorylation and activation of cAMP-dependent protein kinase by phosphoinositide-dependent protein kinase. Proc. Natl. Acad. Sci. USA 1998, 95, 9849–9854. [Google Scholar] [CrossRef]
- Limbourg, F.P.; Huang, Z.; Plumier, J.-C.; Simoncini, T.; Fujioka, M.; Tuckermann, J.; Schütz, G.; Moskowitz, M.A.; Liao, J.K. Rapid nontranscriptional activation of endothelial nitric oxide synthase mediates increased cerebral blood flow and stroke protection by corticosteroids. J. Clin. Investig. 2002, 110, 1729–1738. [Google Scholar] [CrossRef]
- Fang, Y.; Sathyanarayanan, S.; Sehgal, A. Post-translational regulation of the Drosophila circadian clock requires protein phosphatase 1 (PP1). Genes Dev. 2007, 21, 1506–1518. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt ID | Description | Spot # | Morning | Evening | p |
|---|---|---|---|---|---|
| MYL1/3 | Essential myosin light chain 1/3 | 1 | 924 (498) | 785 (612) | 0.727 |
| 2 | 4426 (1504) | 3968 (1984) | 0.884 | ||
| 3 | 10,942 (3173) | 9788 (4894) | 0.485 | ||
| 4 | 2378 (808) | 2192 (1293) | 0.431 | ||
| MLRV | Regulatory myosin light chain, slow/ventricular | 5 | 2675 (1524) | 2985 (2179) | 0.498 |
| 6 | 4176 (1837) | 3699 (2182) | 0.459 | ||
| 7 | 1940 (950) | 1600 (10,089) | 0.389 | ||
| MLRS | Regulatory myosin light chain, fast/skeletal | 8 | 1557 (1027) | 2206 (1875) | 0.216 |
| 9 | 4675 (1589) | 5214 (3128) | 0.151 | ||
| 10 | 4420 (1149) | 3762 (1768) | 0.467 | ||
| 11 | 1286 (540) | 982 (569) | 0.426 | ||
| HSPB6 | Heat shock protein 20 | 12 | 282 (214) | 0.00 (0.00) | - |
| Spot # | UniProt ID | Protein Name | MOWSE | % Seq | Anova (p) | FC | Morning Mean | Morning SD | Evening Mean | Evening SD |
|---|---|---|---|---|---|---|---|---|---|---|
| 335 | PYGM | Glycogen phosphorylase, muscle form | 1938 | 53 | 0.0155 | 1.43 | 245 | 44 | 350 | 104 |
| 163 | MYPC1 | Myosin-binding protein C, slow-type | 2135 | 42 | 0.0226 | 1.80 | 343 | 86 | 617 | 357 |
| 703 | MYBPH | Myosin-binding protein H | 425 | 23 | 0.0313 | 1.17 | 133 | 28 | 114 | 26 |
| 874 | ENOB | Beta-enolase | 1040 | 55 | 0.0335 | 1.24 | 814 | 199 | 1007 | 174 |
| 1046 | TNNT1 | Troponin T, slow skeletal muscle | 738 | 32 | 0.0433 | 1.22 | 239 | 71 | 196 | 45 |
| 154 | NEBU | Nebulin | 964 | 6 | 0.0435 | 1.48 | 959 | 352 | 648 | 252 |
| 391 | MYH2 | Myosin-2 | 1510 | 18 | 0.0546 | 1.37 | 239 | 56 | 329 | 158 |
| 881 | KCRM | Creatine kinase M-type | 922 | 54 | 0.0570 | 1.17 | 535 | 74 | 625 | 118 |
| 795 | ACTS | Actin, alpha skeletal muscle | 1011 | 54 | 0.0620 | 1.47 | 271 | 114 | 185 | 56 |
| 742 | ACTN2 | Alpha-actinin-2 | 80 | 4 | 0.0727 | 1.25 | 510 | 102 | 408 | 134 |
| 1059 | TNNT3 | Troponin T, fast skeletal muscle | 394 | 24 | 0.0735 | 1.32 | 244 | 84 | 186 | 54 |
| 992 | ALDOA | Fructose-bisphosphate aldolase A | 733 | 58 | 0.0929 | 1.19 | 472 | 126 | 559 | 138 |
| 903 | ACTS | Actin, alpha skeletal muscle | 1009 | 62 | 0.0931 | 1.67 | 16,872 | 10,694 | 10,109 | 2314 |
| 135 | MYPC2 | Myosin-binding protein C, fast-type | 2015 | 45 | 0.0978 | 1.23 | 168 | 54 | 137 | 41 |
| 1357 | KCRM | Creatine kinase M-type | 466 | 32 | 0.0978 | 1.43 | 2542 | 1135 | 1782 | 577 |
| 686 | NEBU | Nebulin | 2181 | 13 | 0.1017 | 1.17 | 336 | 66 | 394 | 69 |
| 386 | MYH2 | Myosin-2 | 1266 | 13 | 0.1048 | 1.27 | 233 | 78 | 295 | 104 |
| 739 | DESM | Desmin | 1884 | 63 | 0.1072 | 1.20 | 1176 | 181 | 979 | 292 |
| 1043 | TNNT1 | Troponin T, slow skeletal muscle | 599 | 33 | 0.1143 | 1.28 | 884 | 293 | 689 | 278 |
| 596 | MYH7 | Myosin-7 | 1229 | 17 | 0.1321 | 1.15 | 259 | 66 | 298 | 49 |
| 392 | MYH2 | Myosin-2 | 709 | 12 | 0.1328 | 1.17 | 109 | 44 | 128 | 36 |
| 390 | MYH2 | Myosin-2 | 1367 | 14 | 0.1481 | 1.28 | 177 | 40 | 227 | 89 |
| 855 | ACTS | Actin, alpha skeletal muscle | 1319 | 65 | 0.1531 | 1.25 | 18,032 | 6216 | 14,461 | 4631 |
| 158 | MYPC1 | Myosin-binding protein C, slow-type | 2460 | 49 | 0.1606 | 1.32 | 547 | 224 | 721 | 297 |
| 387 | MYH2 | Myosin-2 | 915 | 12 | 0.1636 | 1.24 | 173 | 85 | 214 | 71 |
| 685 | KPYM | Pyruvate kinase isozymes M1/M2 | 1329 | 51 | 0.1662 | 1.17 | 398 | 68 | 466 | 112 |
| 731 | DESM | Desmin | 2161 | 72 | 0.1818 | 1.16 | 2919 | 408 | 2527 | 728 |
| 153 | MYPC1 | Myosin-binding protein C, slow-type | 2648 | 51 | 0.1857 | 1.34 | 518 | 218 | 694 | 314 |
| 344 | PYGM | Glycogen phosphorylase, muscle form | 1560 | 43 | 0.1876 | 1.53 | 198 | 48 | 304 | 242 |
| 741 | DESM | Desmin | 1927 | 67 | 0.1927 | 1.17 | 3290 | 778 | 2811 | 997 |
| 544 | ALBU | Serum albumin | 1539 | 56 | 0.2047 | 1.25 | 682 | 203 | 852 | 348 |
| 389 | TNNT1 | Troponin T, slow skeletal muscle | 153 | 23 | 0.2070 | 1.16 | 558 | 103 | 645 | 155 |
| 1065 | TNNT3 | Troponin T, fast skeletal muscle | 421 | 26 | 0.2103 | 1.18 | 542 | 124 | 460 | 117 |
| 164 | MYPC1 | Myosin-binding protein C, slow-type | 2581 | 51 | 0.2105 | 1.25 | 337 | 57 | 423 | 190 |
| 751 | TBA4A | Tubulin alpha-4A | 467 | 29 | 0.2232 | 1.14 | 312 | 60 | 274 | 43 |
| 997 | ALDOA | Fructose-bisphosphate aldolase A | 812 | 60 | 0.2234 | 1.21 | 652 | 188 | 789 | 289 |
| 546 | ALBU | Serum albumin | 1622 | 59 | 0.2244 | 1.34 | 1381 | 468 | 1850 | 766 |
| 812 | ATPA | ATP synthase subunit alpha, mitochondrial | 1275 | 45 | 0.2274 | 1.15 | 688 | 164 | 790 | 219 |
| 150 | MYPC1 | Myosin-binding protein C, slow-type | 2849 | 50 | 0.2300 | 1.30 | 442 | 236 | 575 | 280 |
| 813 | ATPA | ATP synthase subunit alpha, mitochondrial | 963 | 42 | 0.2328 | 1.10 | 755 | 167 | 829 | 176 |
| 1606 | VIME | Vimentin | 1861 | 69 | 0.2342 | 1.35 | 707 | 339 | 955 | 389 |
| 1105 | TPM2 | Tropomyosin beta chain | 567 | 32 | 0.2501 | 1.19 | 454 | 120 | 381 | 80 |
| 1251 | CAH3 | Carbonic anhydrase 3 | 419 | 44 | 0.2622 | 1.14 | 829 | 240 | 944 | 230 |
| 878 | ENOB | Beta-enolase | 1196 | 53 | 0.2624 | 1.19 | 1109 | 283 | 1325 | 358 |
| 616 | CATA | Catalase | 710 | 38 | 0.2974 | 1.77 | 832 | 103 | 1476 | 1963 |
| 941 | KCRM | Creatine kinase M-type | 890 | 54 | 0.3013 | 1.14 | 1295 | 299 | 1480 | 439 |
| 947 | KCRM | Creatine kinase M-type | 1295 | 57 | 0.3075 | 1.14 | 6801 | 1461 | 7786 | 1689 |
| 1057 | ACTC | Actin, alpha cardiac muscle 1 | 52 | 11 | 0.3596 | 1.16 | 329 | 118 | 383 | 185 |
| 505 | HSP7C | Heat shock cognate 71 kDa protein | 1246 | 39 | 0.3731 | 1.19 | 1386 | 475 | 1643 | 850 |
| 500 | HSP71 | Heat shock 70 kDa protein 1A/1B | 1470 | 46 | 0.3816 | 1.02 | 583 | 99 | 570 | 136 |
| 1085 | TPM3 | Tropomyosin alpha-3 chain | 1478 | 72 | 0.3846 | 1.10 | 37,171 | 11,538 | 33,691 | 9083 |
| 1603 | ALBU | Serum albumin | 1625 | 58 | 0.3869 | 1.22 | 714 | 404 | 867 | 335 |
| 1097 | TNNT1 | Troponin T, slow skeletal muscle | 576 | 32 | 0.3914 | 1.20 | 1448 | 656 | 1207 | 609 |
| 553 | ALBU | Serum albumin | 1838 | 60 | 0.3938 | 1.29 | 4881 | 2765 | 6293 | 3338 |
| 902 | ACTS | Actin, alpha skeletal muscle | 962 | 63 | 0.4326 | 1.25 | 8992 | 5539 | 7196 | 2076 |
| 1374 | MYL3 | Myosin light chain 3 | 856 | 77 | 0.4537 | 1.10 | 8098 | 3473 | 7382 | 2866 |
| 1078 | MYOZ1 | Myozenin-1 | 848 | 85 | 0.4556 | 1.00 | 401 | 76 | 402 | 239 |
| 1336 | TPIS | Triosephosphate isomerase | 929 | 76 | 0.4585 | 1.04 | 316 | 48 | 304 | 100 |
| 1103 | KCRM | Creatine kinase M-type | 418 | 29 | 0.4598 | 1.40 | 623 | 607 | 446 | 123 |
| 944 | KCRM | Creatine kinase M-type | 1100 | 53 | 0.4644 | 1.12 | 3853 | 690 | 4310 | 1195 |
| 900 | ACTS | Actin, alpha skeletal muscle | 415 | 39 | 0.4649 | 1.21 | 3869 | 2169 | 3198 | 713 |
| 1272 | MYOZ1 | Myozenin-1 | 154 | 29 | 0.4785 | 1.05 | 355 | 88 | 373 | 47 |
| 532 | PGM1 | Phosphoglucomutase-1 | 876 | 37 | 0.4889 | 1.09 | 446 | 125 | 409 | 80 |
| 106 | MYH2 | Myosin-2 | 3726 | 33 | 0.4924 | 1.59 | 550 | 340 | 874 | 888 |
| 1090 | TNNT3 | Troponin T, fast skeletal muscle | 520 | 32 | 0.5034 | 1.12 | 591 | 178 | 528 | 89 |
| 333 | PYGM | Glycogen phosphorylase, muscle form | 1611 | 47 | 0.5039 | 1.21 | 164 | 33 | 198 | 95 |
| 1503 | KCRM | Creatine kinase M-type | 151 | 13 | 0.5085 | 1.06 | 1063 | 345 | 1007 | 590 |
| 1032 | TNNT3 | Troponin T, fast skeletal muscle | 491 | 27 | 0.5192 | 1.23 | 1032 | 534 | 837 | 258 |
| 943 | KCRM | Creatine kinase M-type | 1126 | 59 | 0.5267 | 1.06 | 2690 | 574 | 2846 | 551 |
| 1056 | TNNT1 | Troponin T, slow skeletal muscle | 687 | 37 | 0.5340 | 1.13 | 8515 | 4319 | 7506 | 4221 |
| 1049 | TNNT1 | Troponin T, slow skeletal muscle | 652 | 36 | 0.5550 | 1.10 | 1792 | 601 | 1630 | 818 |
| 165 | MYPC1 | Myosin-binding protein C, slow-type | 2366 | 51 | 0.5618 | 1.14 | 323 | 69 | 368 | 161 |
| 1266 | TPIS | Triosephosphate isomerase | 932 | 65 | 0.5624 | 1.07 | 1389 | 280 | 1489 | 354 |
| 1132 | MYH2 | Myosin-2 | 616 | 8 | 0.5679 | 1.05 | 1154 | 324 | 1209 | 255 |
| 604 | MYH2 | Myosin-2 | 1104 | 13 | 0.5805 | 1.05 | 199 | 43 | 189 | 41 |
| 121 | MYH2 | Myosin-2 | 4044 | 37 | 0.5910 | 1.51 | 691 | 599 | 1041 | 1196 |
| 455 | KPYM | Pyruvate kinase isozymes M1/M2 | 815 | 36 | 0.6060 | 1.06 | 856 | 158 | 810 | 117 |
| 550 | ALBU | Serum albumin | 1852 | 65 | 0.6109 | 1.23 | 2514 | 1360 | 3093 | 1897 |
| 1074 | G3P | Glyceraldehyde-3-phosphate dehydrogenase | 1068 | 54 | 0.6170 | 1.03 | 3635 | 771 | 3532 | 1105 |
| 925 | ACTS | Actin, alpha skeletal muscle | 1277 | 63 | 0.6537 | 1.05 | 73,798 | 17,469 | 77,544 | 17,257 |
| 123 | MYH2 | Myosin-2 | 3325 | 34 | 0.6580 | 1.31 | 819 | 851 | 1072 | 1290 |
| 798 | ATPB | ATP synthase subunit beta, mitochondrial | 1882 | 68 | 0.6678 | 1.13 | 947 | 356 | 1069 | 547 |
| 1083 | TNNT3 | Troponin T, fast skeletal muscle | 583 | 34 | 0.6867 | 1.12 | 2406 | 775 | 2689 | 1066 |
| 136 | MYPC2 | Myosin-binding protein C, fast-type | 1838 | 41 | 0.6868 | 1.07 | 223 | 72 | 207 | 45 |
| 1073 | TNNT3 | Troponin T, fast skeletal muscle | 482 | 26 | 0.6887 | 1.09 | 1620 | 579 | 1492 | 603 |
| 1048 | TNNT1 | Troponin T, slow skeletal muscle | 738 | 32 | 0.6912 | 1.25 | 3355 | 2869 | 2681 | 1052 |
| 471 | MYH2 | Myosin-2 | 1204 | 15 | 0.6940 | 1.09 | 202 | 91 | 186 | 98 |
| 934 | KCRM | Creatine kinase M-type | 1134 | 54 | 0.6981 | 1.03 | 1253 | 233 | 1222 | 210 |
| 1115 | MYOZ1 | Myozenin-1 | 378 | 30 | 0.7011 | 1.05 | 1462 | 416 | 1393 | 415 |
| 904 | ACTS | Actin, alpha skeletal muscle | 877 | 55 | 0.7143 | 1.01 | 9431 | 1361 | 9383 | 3034 |
| 443 | MYH2 | Myosin-2 | 978 | 13 | 0.7267 | 1.05 | 331 | 103 | 348 | 96 |
| 1381 | KCRM | Creatine kinase M-type | 469 | 28 | 0.7267 | 1.08 | 24,048 | 9478 | 22,334 | 6612 |
| 115 | MYH7 | Myosin-7 | 4288 | 35 | 0.7551 | 1.14 | 2646 | 2960 | 3028 | 3141 |
| 1061 | TNNT3 | Troponin T, fast skeletal muscle | 457 | 36 | 0.7595 | 1.28 | 540 | 382 | 424 | 155 |
| 1044 | ACTC | Actin, alpha cardiac muscle 1 | 342 | 28 | 0.7668 | 1.14 | 2912 | 1480 | 2552 | 917 |
| 1080 | ACTC | Actin, alpha cardiac muscle 1 | 183 | 23 | 0.7707 | 1.06 | 4741 | 1932 | 4479 | 2310 |
| 270 | ACTN2 | Alpha-actinin-2 | 3558 | 69 | 0.7779 | 1.08 | 2145 | 422 | 2326 | 812 |
| 271 | ACTN2 | Alpha-actinin-2 | 3354 | 71 | 0.7866 | 1.04 | 679 | 243 | 706 | 256 |
| 274 | ACTN2 | Alpha-actinin-2 | 3445 | 69 | 0.7899 | 1.07 | 1058 | 239 | 1128 | 390 |
| 187 | MYH7 | Myosin-7 | 3534 | 31 | 0.7939 | 1.55 | 6318 | 8061 | 4082 | 3064 |
| 1051 | TNNT3 | Troponin T, fast skeletal muscle | 507 | 36 | 0.8007 | 1.11 | 1454 | 657 | 1312 | 446 |
| 1108 | MYOZ1 | Myozenin-1 | 708 | 50 | 0.8010 | 1.02 | 3435 | 1569 | 3513 | 1097 |
| 332 | PYGM | Glycogen phosphorylase, muscle form | 1651 | 44 | 0.8203 | 1.01 | 295 | 50 | 299 | 93 |
| 330 | TPM1 | Tropomyosin alpha-1 chain | 947 | 47 | 0.8216 | 1.07 | 1964 | 874 | 1834 | 716 |
| 1100 | TNNT3 | Troponin T, fast skeletal muscle | 474 | 37 | 0.8243 | 1.12 | 4641 | 2707 | 4144 | 1391 |
| 1095 | TNNT3 | Troponin T, fast skeletal muscle | 452 | 28 | 0.8373 | 1.09 | 1477 | 719 | 1355 | 374 |
| 499 | HSP71 | Heat shock 70 kDa protein 1A/1B | 1070 | 39 | 0.8419 | 1.02 | 516 | 114 | 508 | 118 |
| 980 | TNNT1 | Troponin T, slow skeletal muscle | 696 | 35 | 0.8427 | 1.01 | 1131 | 271 | 1146 | 265 |
| 1198 | LDB3 | LIM domain-binding protein 3 | 609 | 17 | 0.8445 | 1.01 | 2437 | 827 | 2415 | 1038 |
| 899 | DESM | Desmin | 198 | 11 | 0.8495 | 1.08 | 1329 | 574 | 1230 | 235 |
| 1129 | VDAC1 | Voltage-dependent anion-selective channel protein 1 | 612 | 59 | 0.8774 | 1.10 | 383 | 206 | 347 | 82 |
| 1552 | HERC1 | Probable E3 ubiquitin-protein ligase HERC1 | 52 | 2 | 0.8852 | 1.06 | 1557 | 547 | 1652 | 1207 |
| 1101 | TNNT3 | Troponin T, fast skeletal muscle | 540 | 33 | 0.9155 | 1.01 | 1068 | 440 | 1077 | 383 |
| 477 | MYH2 | Myosin-2 | 2036 | 26 | 0.9173 | 1.08 | 564 | 398 | 609 | 497 |
| 533 | MYH2 | Myosin-2 | 1084 | 13 | 0.9250 | 1.27 | 365 | 409 | 289 | 165 |
| 1397 | ENOB | Beta-enolase | 266 | 26 | 0.9287 | 1.02 | 12,335 | 3415 | 12,540 | 3761 |
| 535 | MYH2 | Myosin-2 | 1195 | 17 | 0.9394 | 1.23 | 456 | 482 | 370 | 163 |
| 1451 | TNNI2 | Troponin I, fast skeletal muscle | 378 | 50 | 0.9545 | 1.02 | 229 | 127 | 225 | 130 |
| 479 | MYH2 | Myosin-2 | 1359 | 18 | 0.9612 | 1.04 | 543 | 385 | 563 | 481 |
| 275 | ACTN2 | Alpha-actinin-2 | 3512 | 71 | 0.9648 | 1.04 | 925 | 221 | 965 | 355 |
| 534 | MYH2 | Myosin-2 | 1360 | 16 | 0.9683 | 1.14 | 588 | 587 | 515 | 327 |
| 336 | PYGM | Glycogen phosphorylase, muscle form | 1778 | 50 | 0.9720 | 1.02 | 310 | 96 | 305 | 69 |
| 1251 | HBB | Hemoglobin subunit beta | 441 | 60 | 0.9780 | 1.00 | 834 | 137 | 837 | 171 |
| 195 | MYH7 | Myosin-7 | 4307 | 32 | 0.9894 | 1.34 | 2446 | 3633 | 1821 | 1188 |
| 1060 | TNNT3 | Troponin T, fast skeletal muscle | 481 | 26 | 0.9915 | 1.01 | 2012 | 749 | 2001 | 801 |
| 1034 | TPM2 | Tropomyosin beta chain | 1450 | 69 | 0.9963 | 1.01 | 22,483 | 6641 | 22,339 | 5236 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ab Malik, Z.; Bowden Davies, K.A.; Hall, E.C.R.; Barrett, J.; Pullinger, S.A.; Erskine, R.M.; Shepherd, S.O.; Iqbal, Z.; Edwards, B.J.; Burniston, J.G. Diurnal Differences in Human Muscle Isometric Force In Vivo Are Associated with Differential Phosphorylation of Sarcomeric M-Band Proteins. Proteomes 2020, 8, 22. https://doi.org/10.3390/proteomes8030022
Ab Malik Z, Bowden Davies KA, Hall ECR, Barrett J, Pullinger SA, Erskine RM, Shepherd SO, Iqbal Z, Edwards BJ, Burniston JG. Diurnal Differences in Human Muscle Isometric Force In Vivo Are Associated with Differential Phosphorylation of Sarcomeric M-Band Proteins. Proteomes. 2020; 8(3):22. https://doi.org/10.3390/proteomes8030022
Chicago/Turabian StyleAb Malik, Zulezwan, Kelly A. Bowden Davies, Elliott C. R. Hall, Jennifer Barrett, Samuel A. Pullinger, Robert M. Erskine, Sam O. Shepherd, Zafar Iqbal, Ben J. Edwards, and Jatin G. Burniston. 2020. "Diurnal Differences in Human Muscle Isometric Force In Vivo Are Associated with Differential Phosphorylation of Sarcomeric M-Band Proteins" Proteomes 8, no. 3: 22. https://doi.org/10.3390/proteomes8030022
APA StyleAb Malik, Z., Bowden Davies, K. A., Hall, E. C. R., Barrett, J., Pullinger, S. A., Erskine, R. M., Shepherd, S. O., Iqbal, Z., Edwards, B. J., & Burniston, J. G. (2020). Diurnal Differences in Human Muscle Isometric Force In Vivo Are Associated with Differential Phosphorylation of Sarcomeric M-Band Proteins. Proteomes, 8(3), 22. https://doi.org/10.3390/proteomes8030022