Gathering and Analyzing Surface Parameters for Diet Identification Purposes



Abstract
1. Introduction
2. Materials and Methods
2.1. Material
- different tooth microwear textures,
- known diet habits of the studied species, and
- the confidence in the procedure reliability.
2.1.1. Triplet 1 (T1), “Old World Monkeys”
2.1.2. Triplet 2 (T2), “European Ruminants”
2.1.3. Triplet 3 (T3), “African Ruminants”
2.1.4. Quadruplet (Q1), “Cervus”
2.1.5. Triplet 4 (T4), “Three cervids”
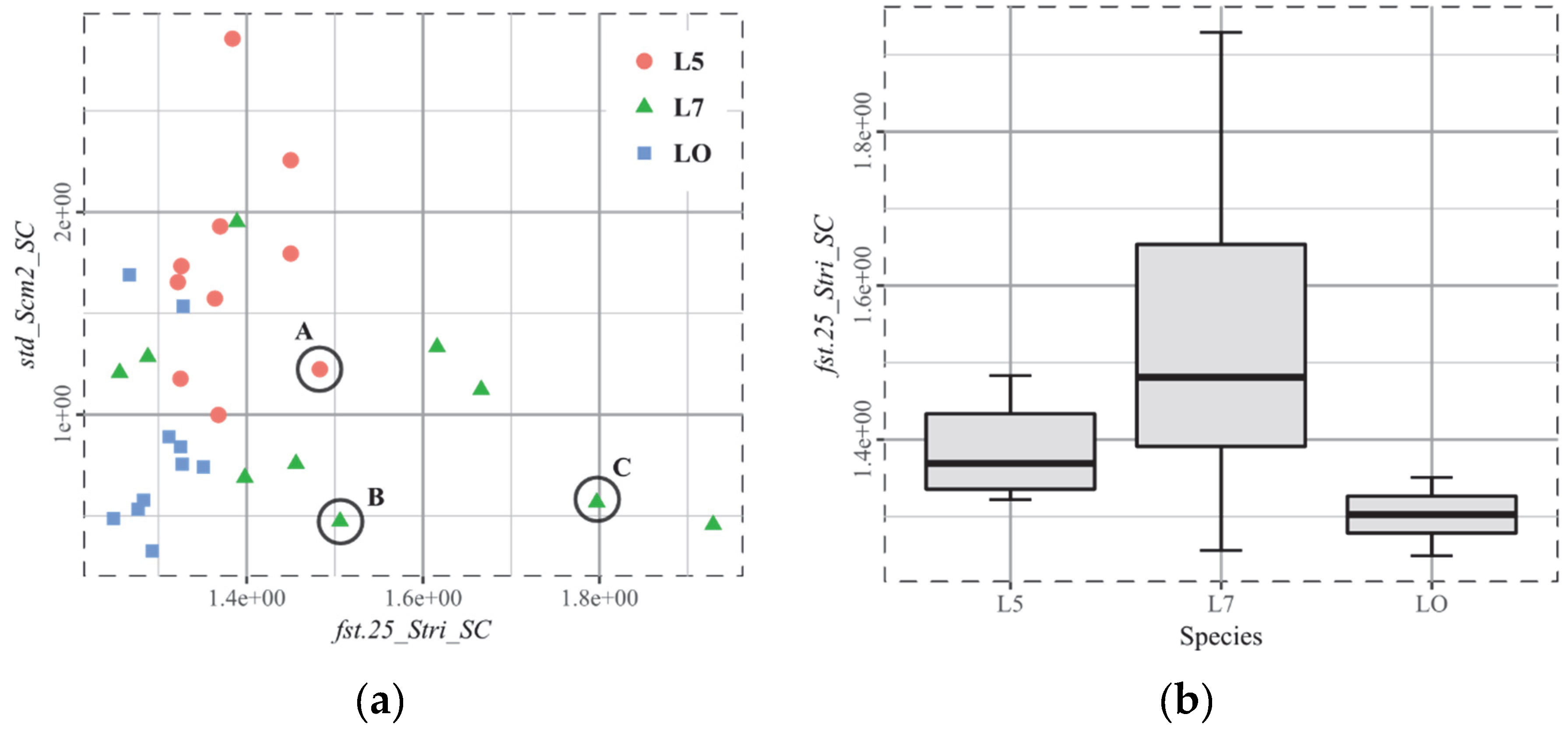
- Although the red deer in southern Spain is highly engaged in grazing and the roe deer from Dourdan in France is a selective browser, many specimens share high anisotropy. The challenge here is to be able to distinguish between the two dietary habits, and thus, to find out which parameters are relevant for that purpose.
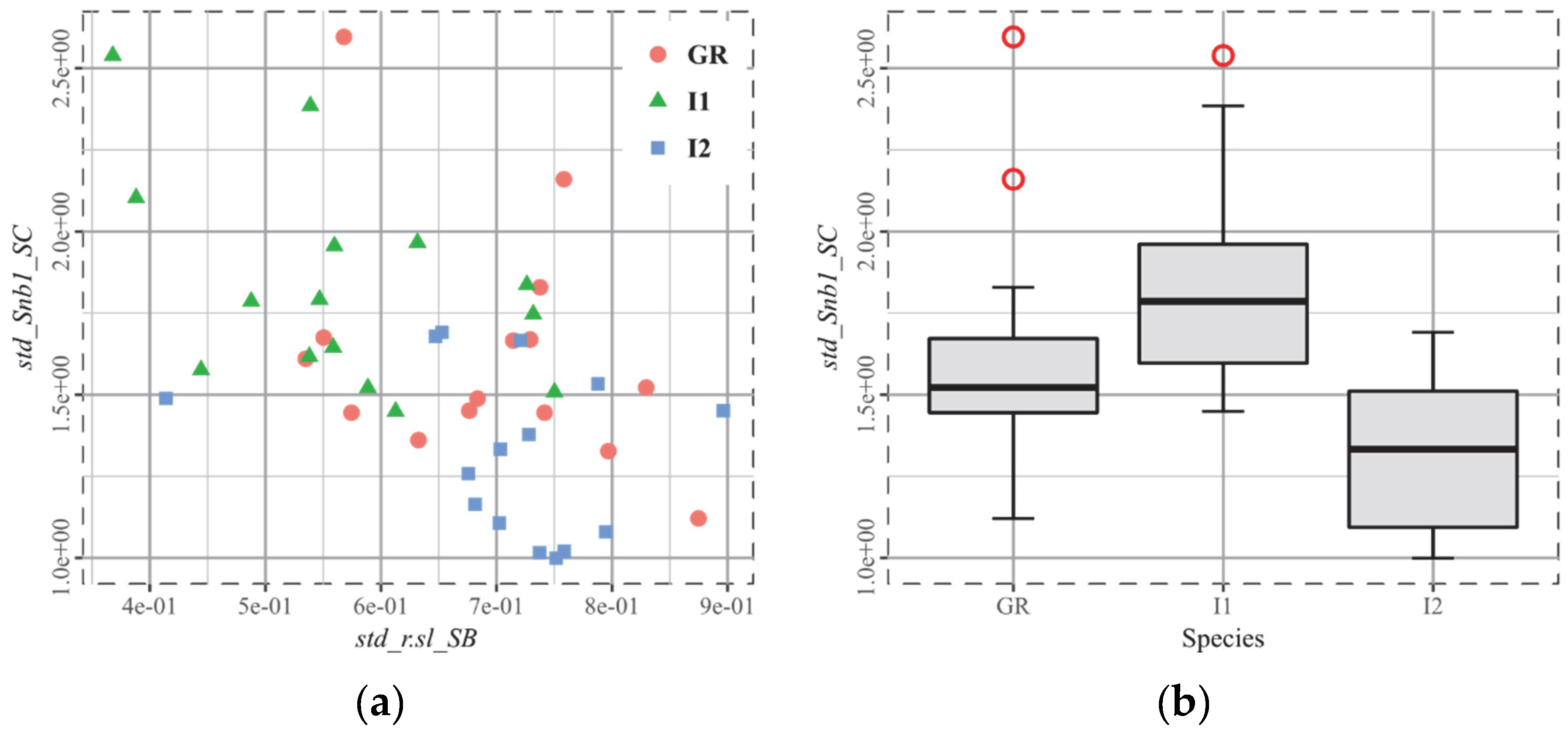
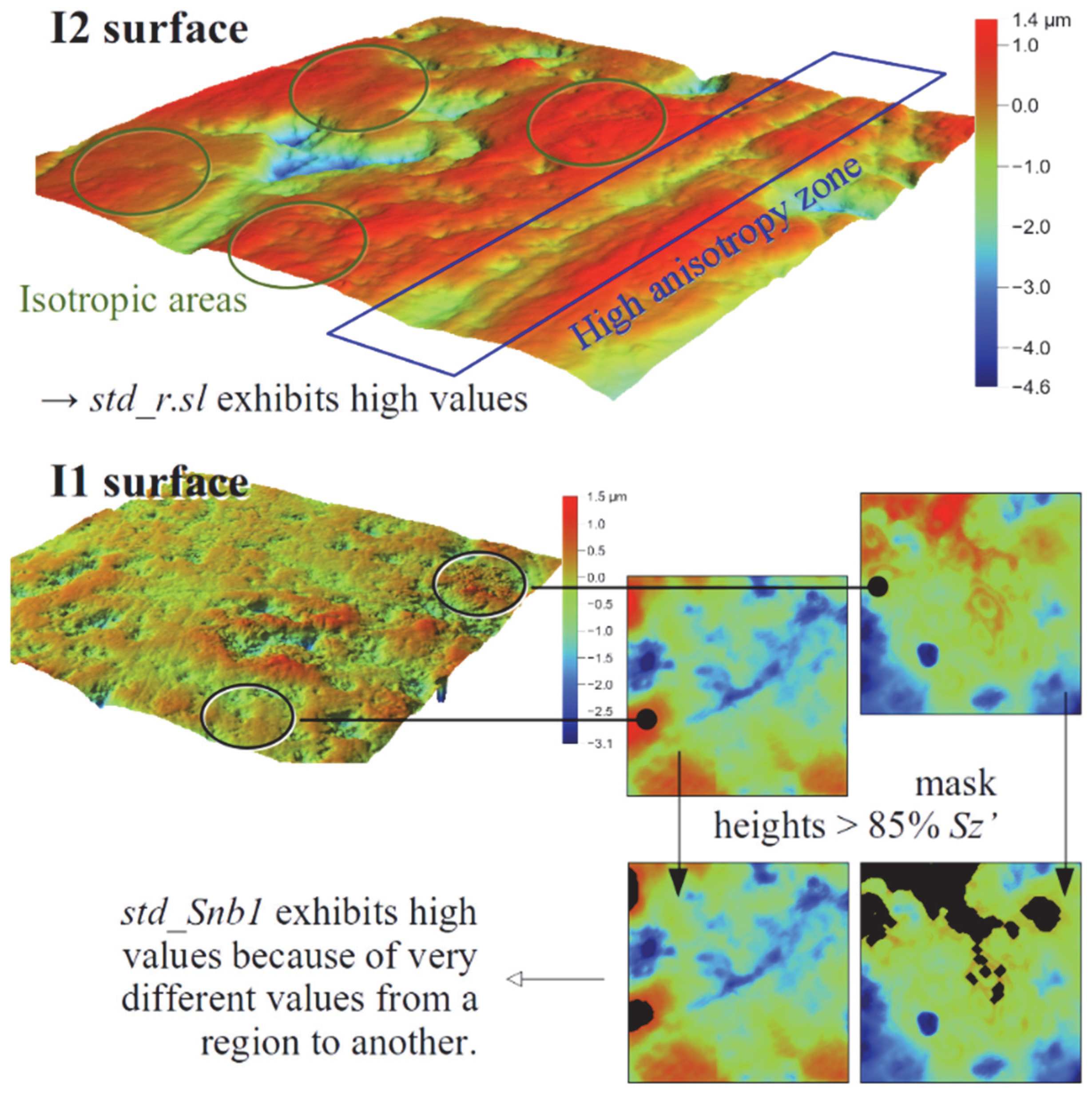
- Roe deer and moose are both browsers. However, they differ significantly in the widely used parameter epLsar (exact proportion length–scale anisotropy of relief, an anisotropy marker). The authors of this study hypothesize that it could be linked to the differences in the amount of ligneous material (woody parts), which is greater for the moose. So, the challenge is to be able to unite them and thus differentiate them from the grazing deer (Cervus elaphus).
2.1.6. Quintet (Q2), “Browse, Grass and Dust” (sheep experiment)
2.1.7. Triplet 5 (T5), “Seeds, Browse, and Grass” (Sheep Experiment)
2.2. Methods
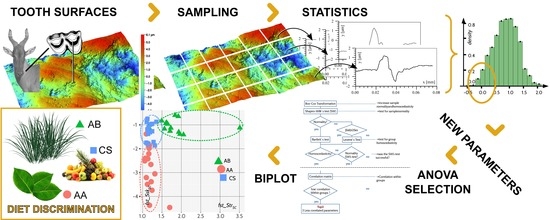
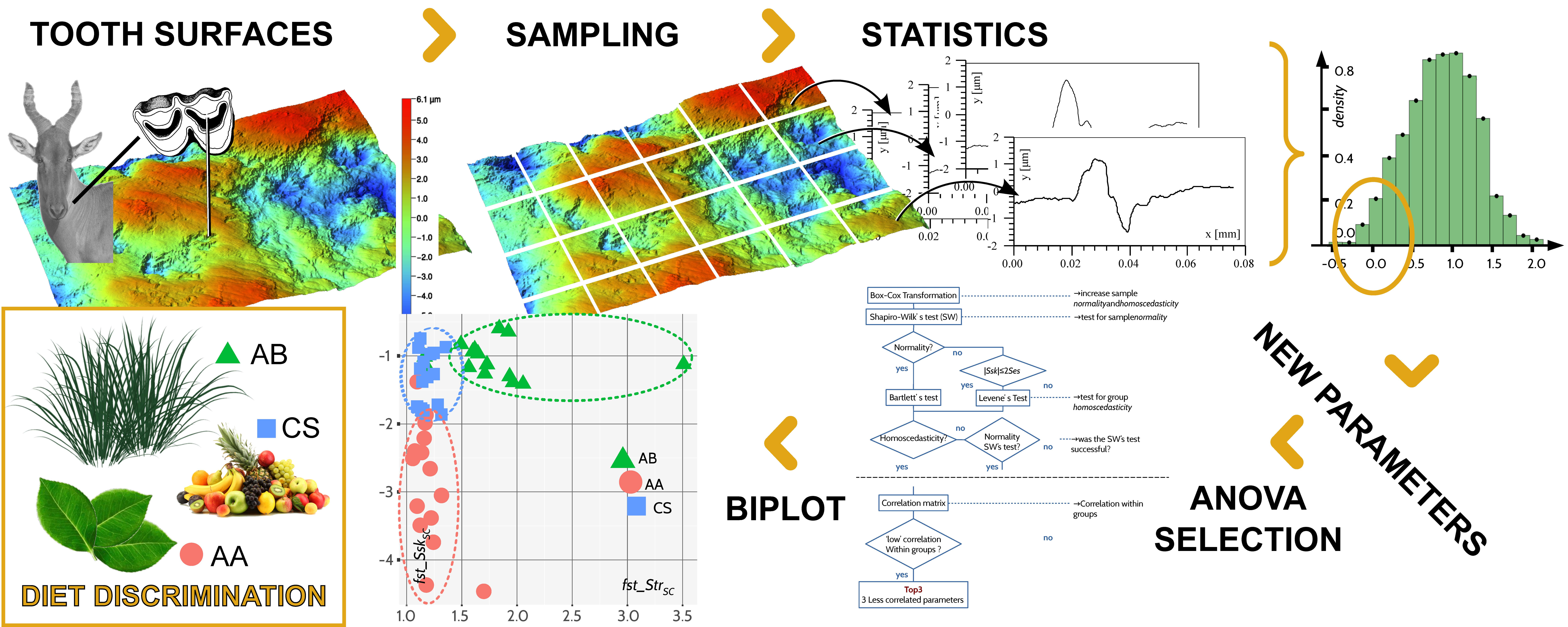
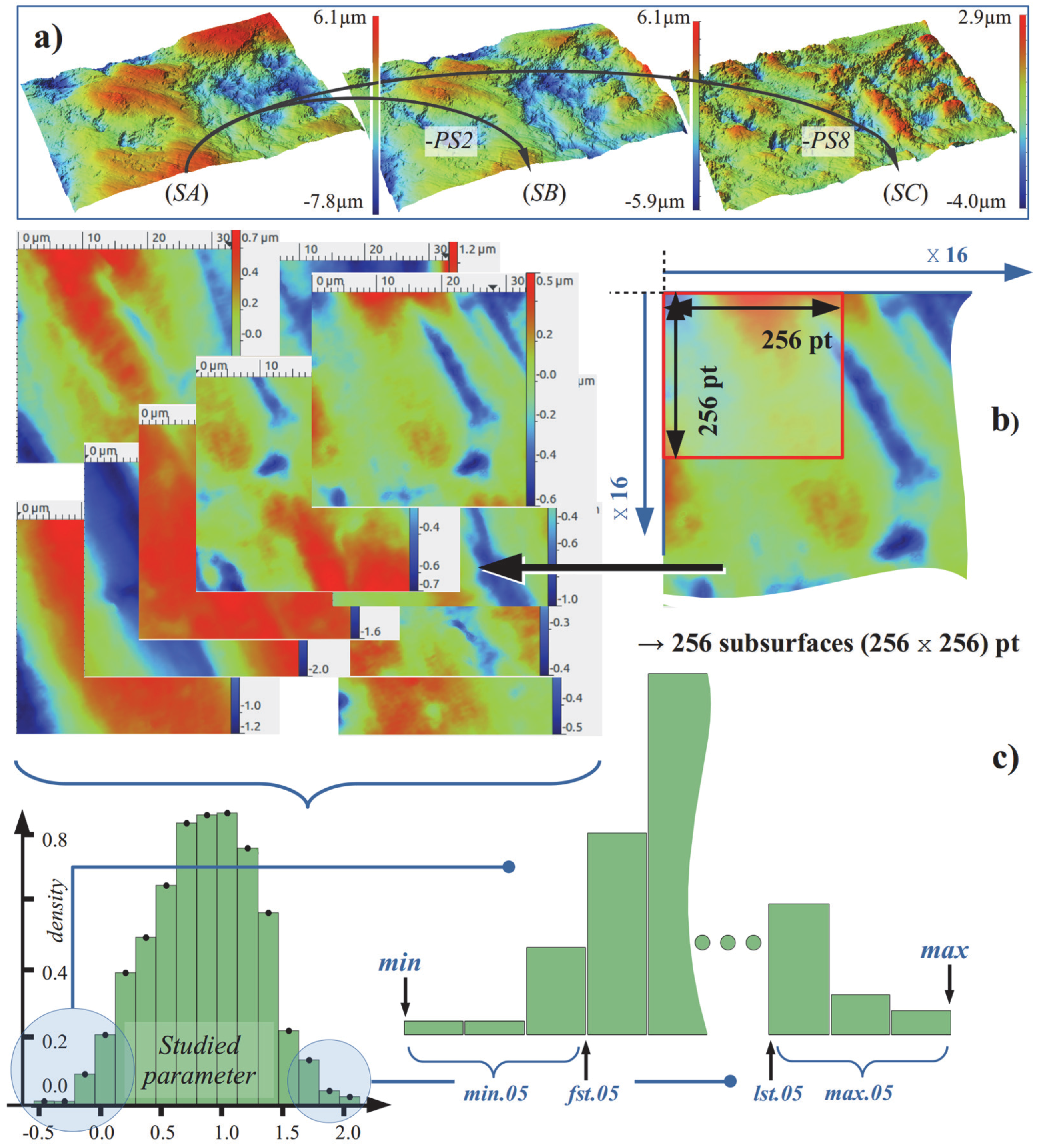
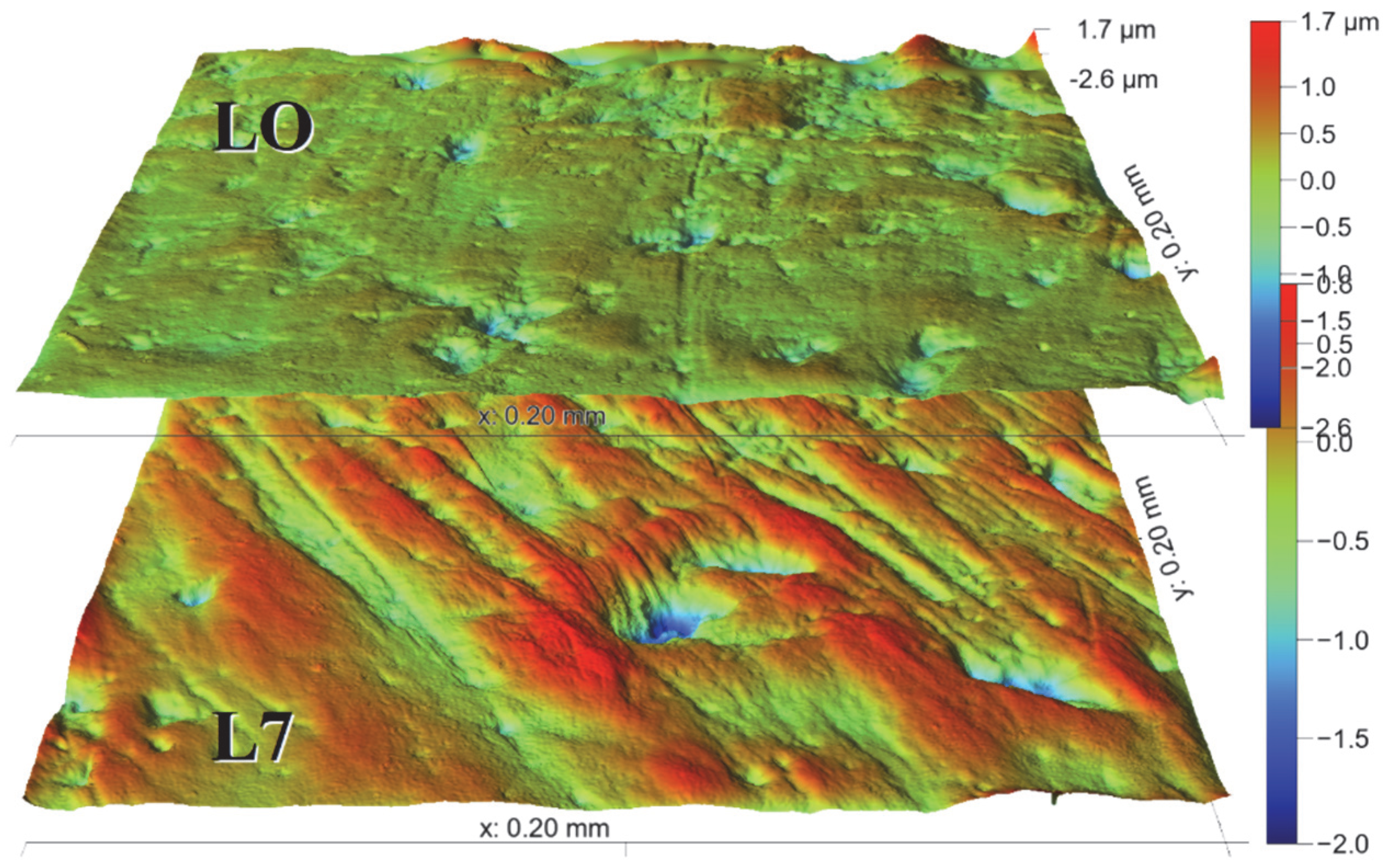
2.2.1. Surface Treatment and Sampling
2.2.2. Height Parameters
2.2.3. Spatial Parameters
2.2.4. Topological Parameters
2.2.5. Fractal Methods, Relative Area Analysis
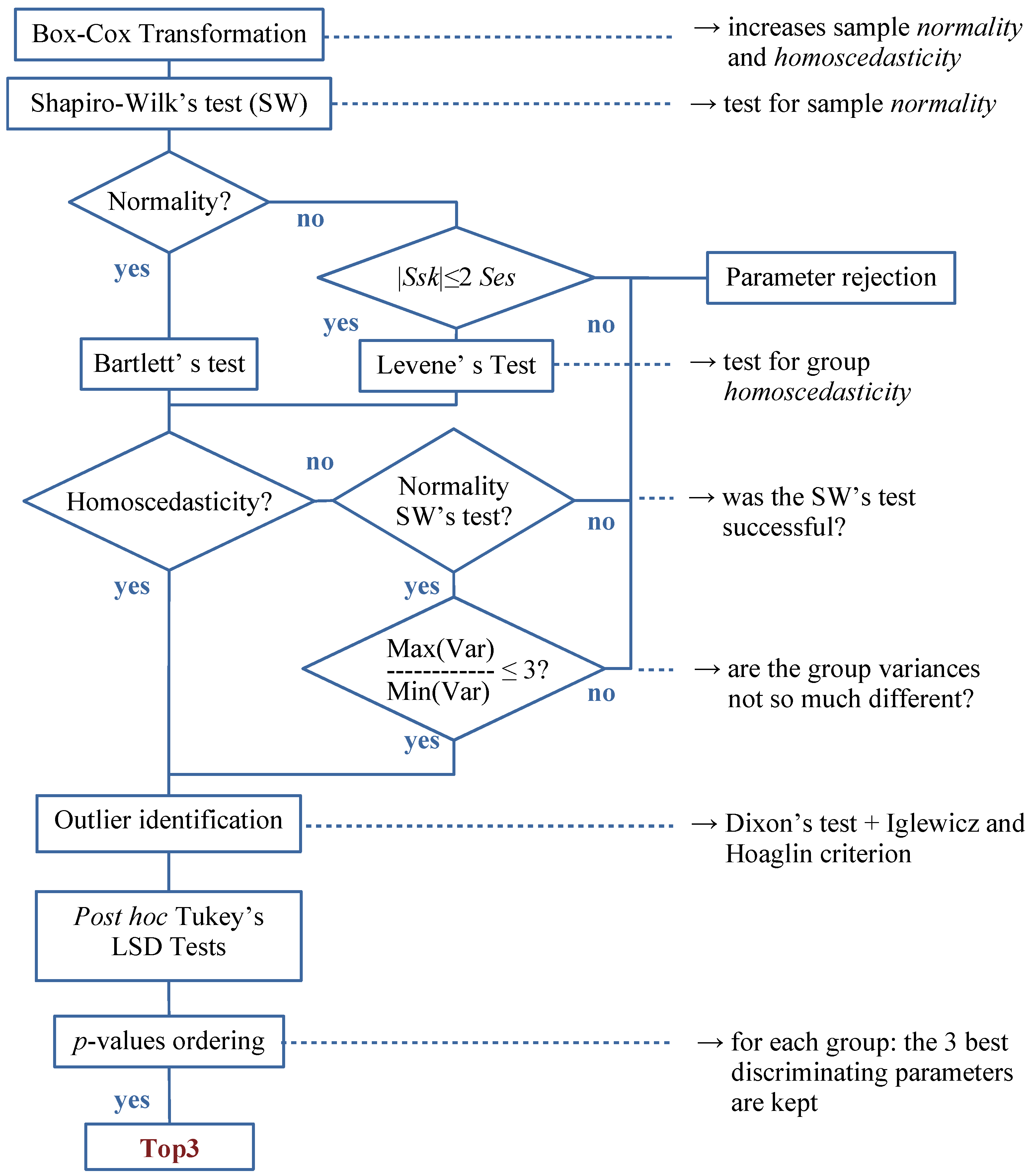
2.2.6. The Automatic Discriminative Procedure
3. Results
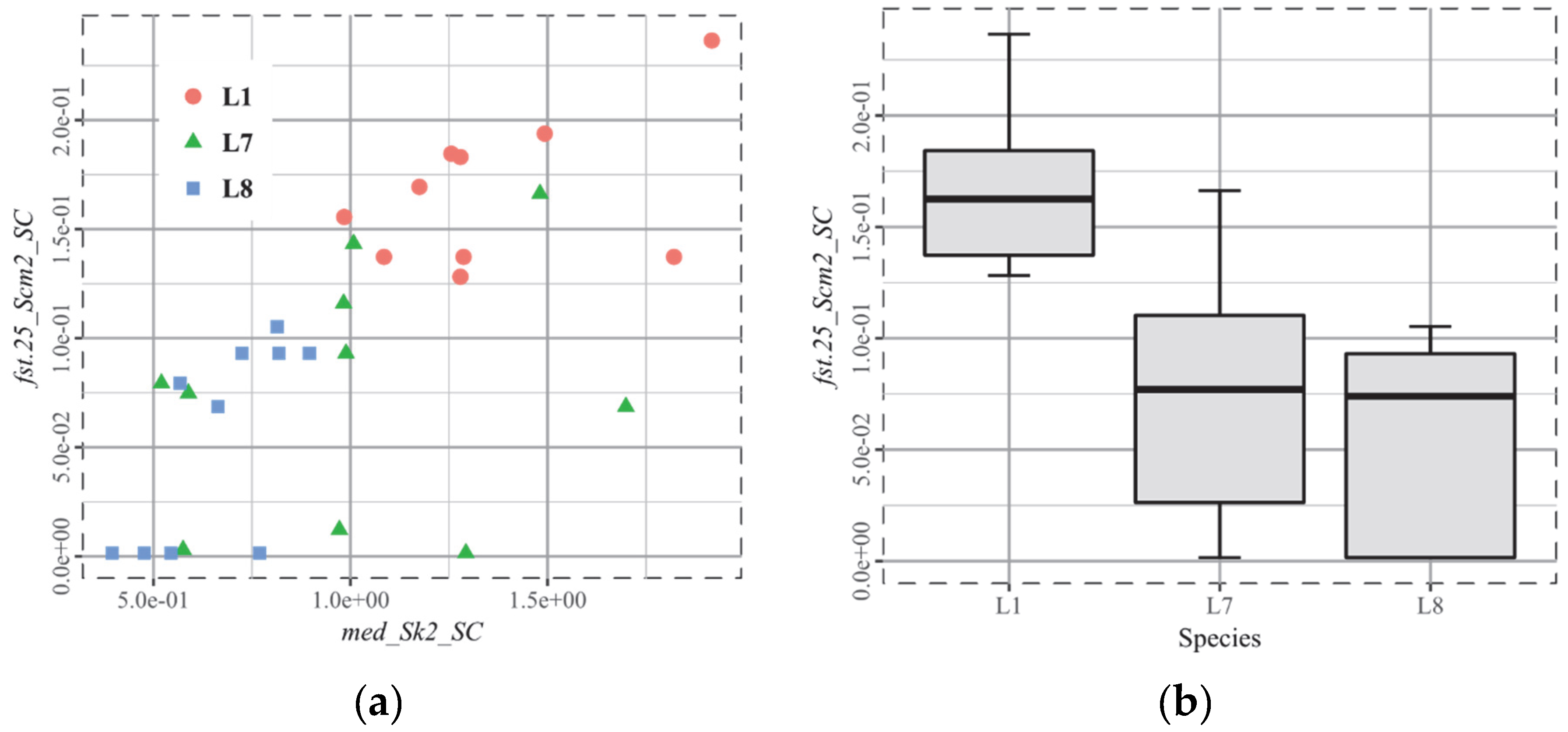
3.1. T1, Old World Monkeys
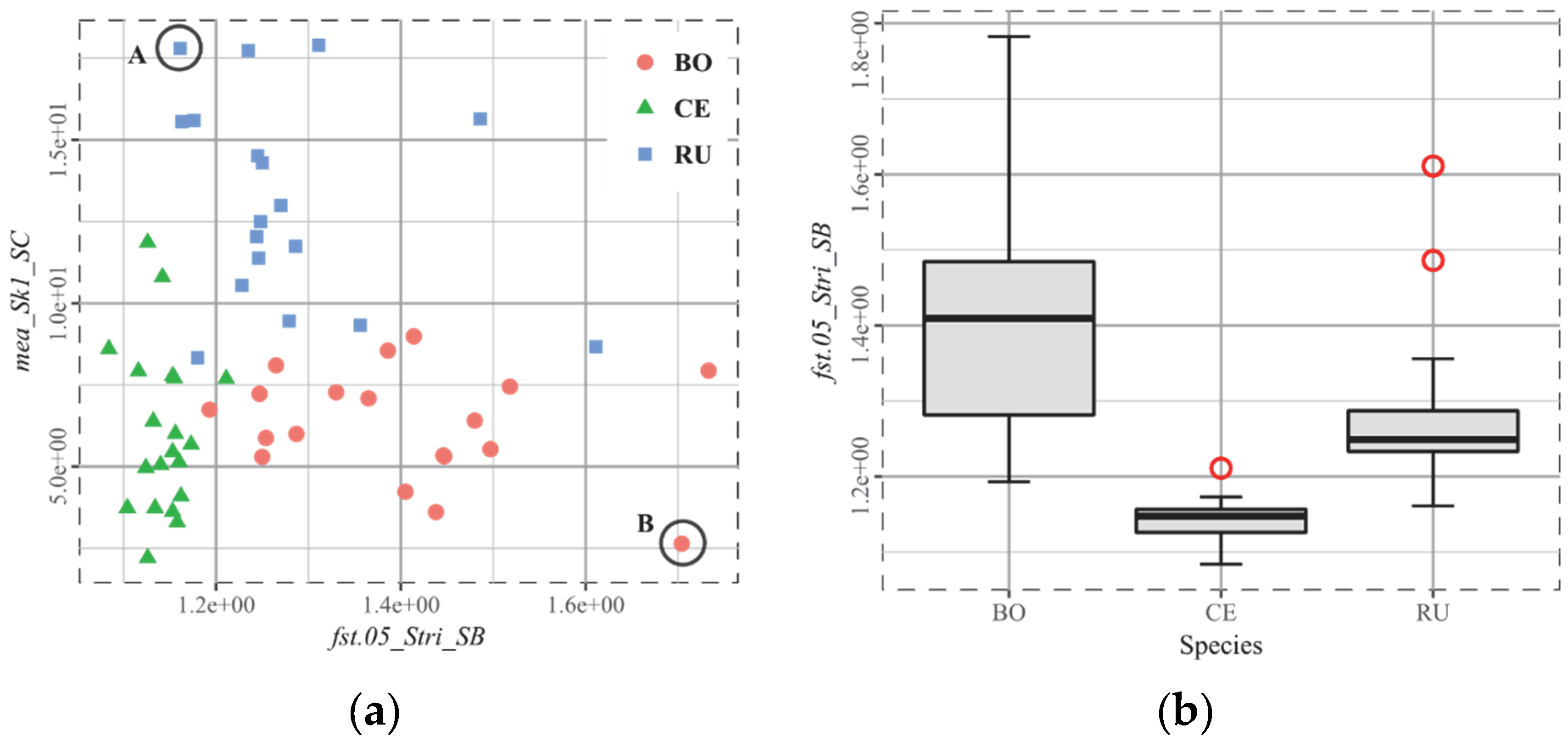
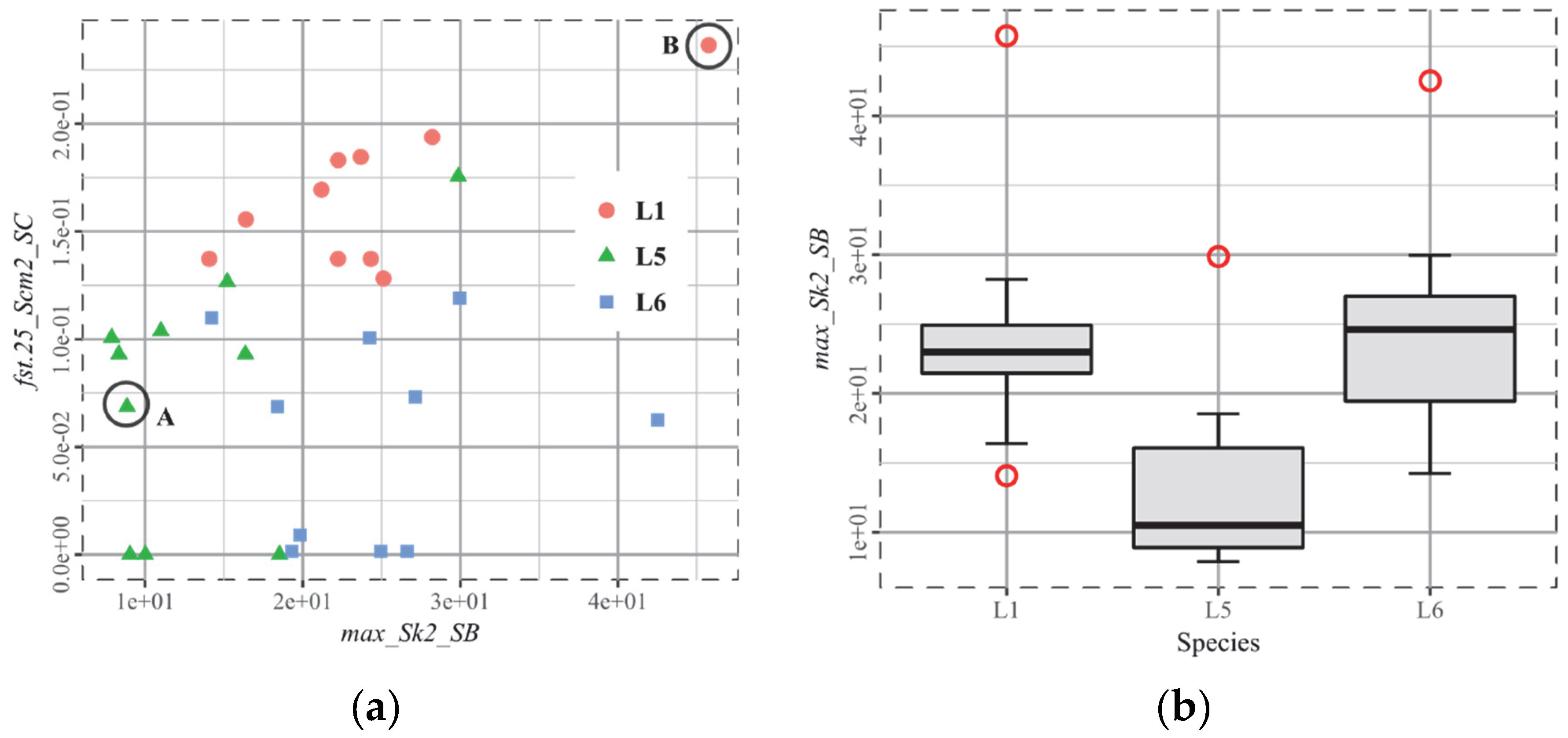
3.2. T2, European Ruminants
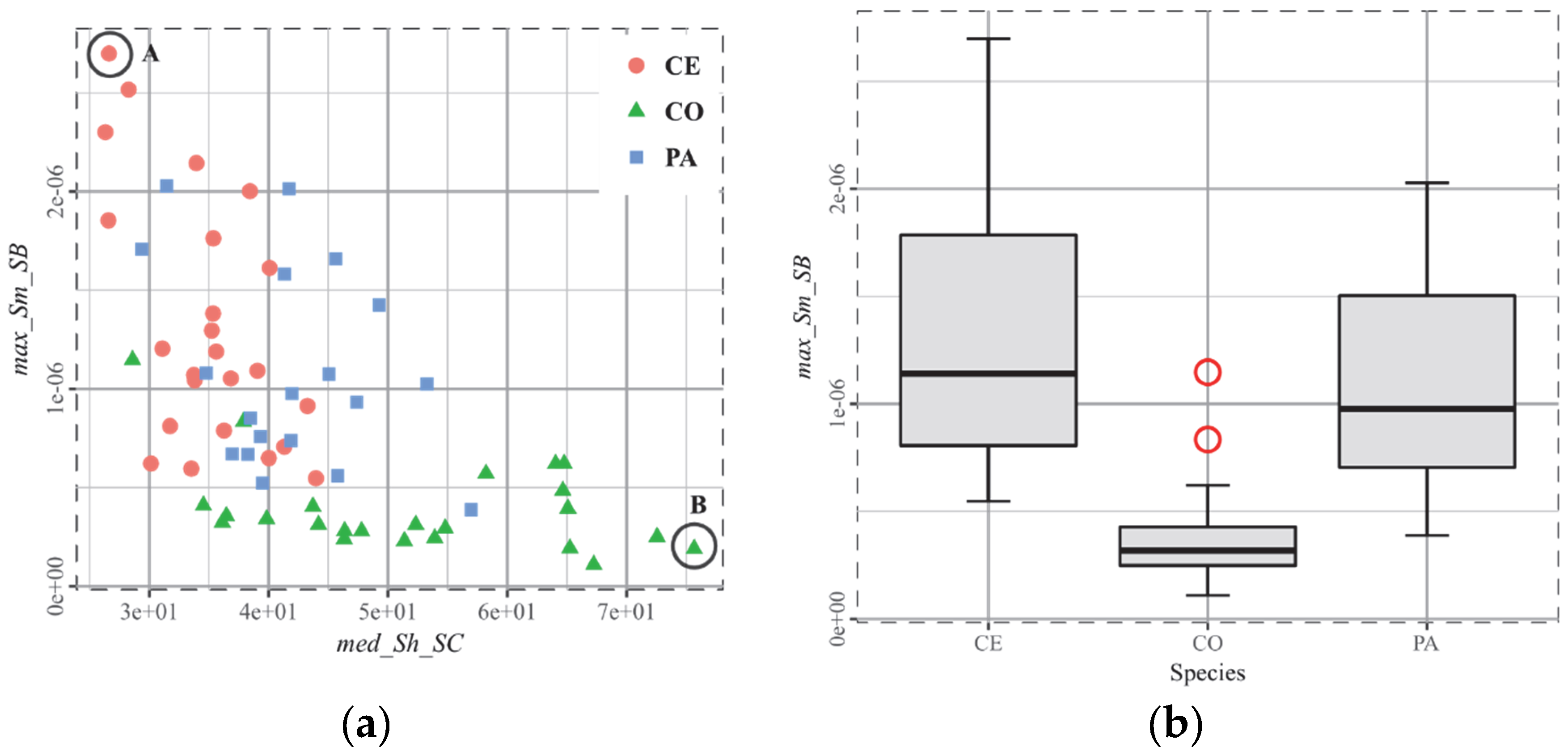
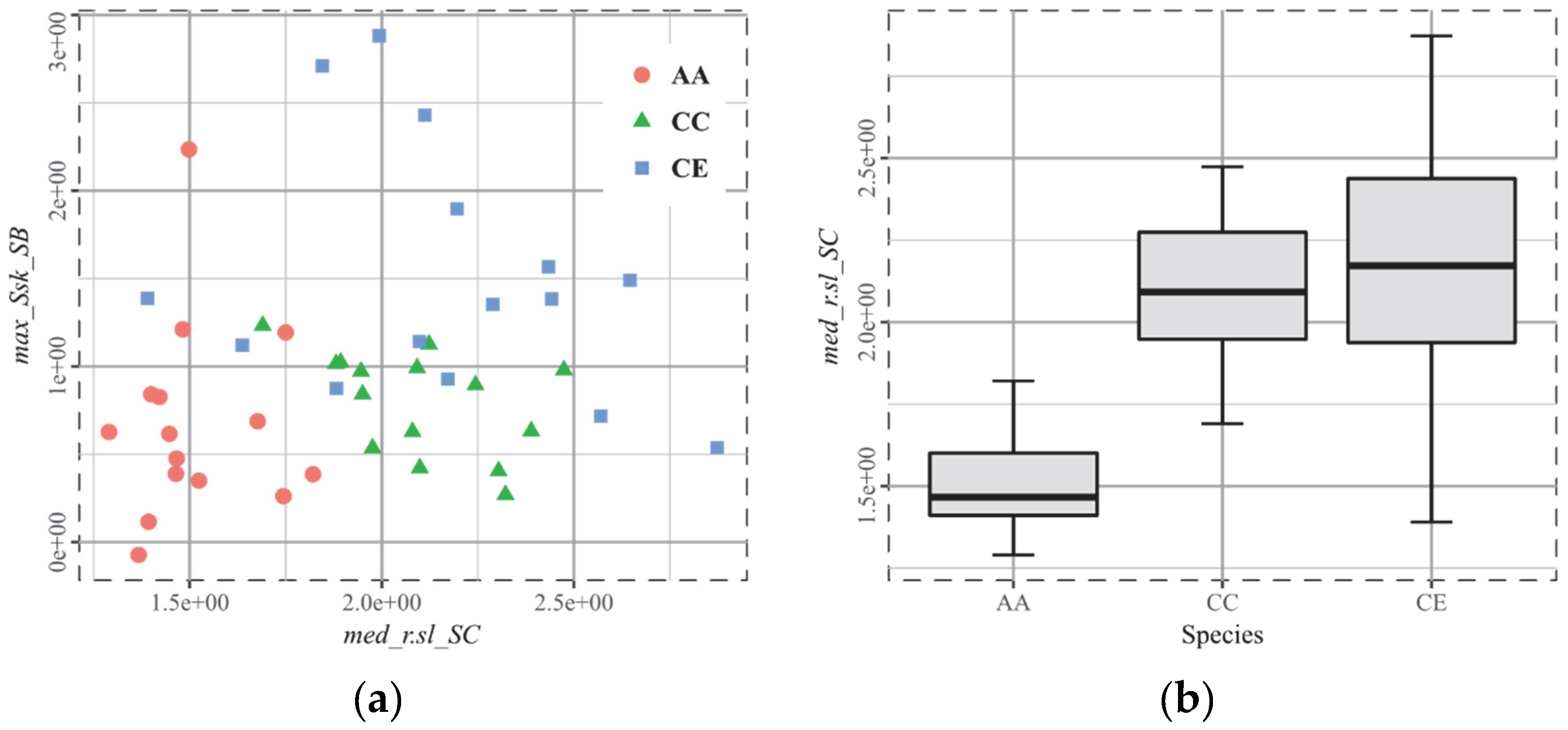
3.3. T3, African Ruminants
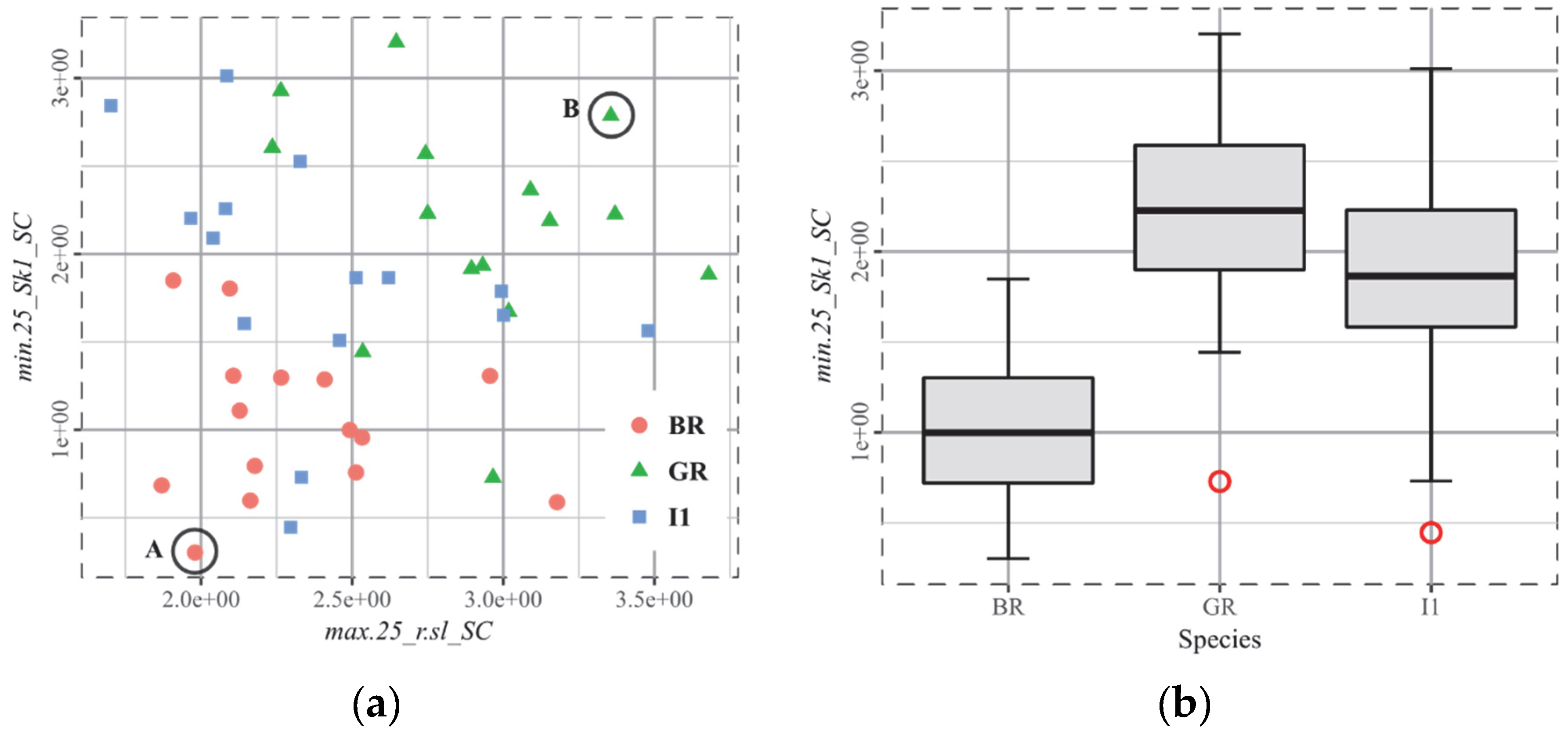
3.4. Q1, Cervus
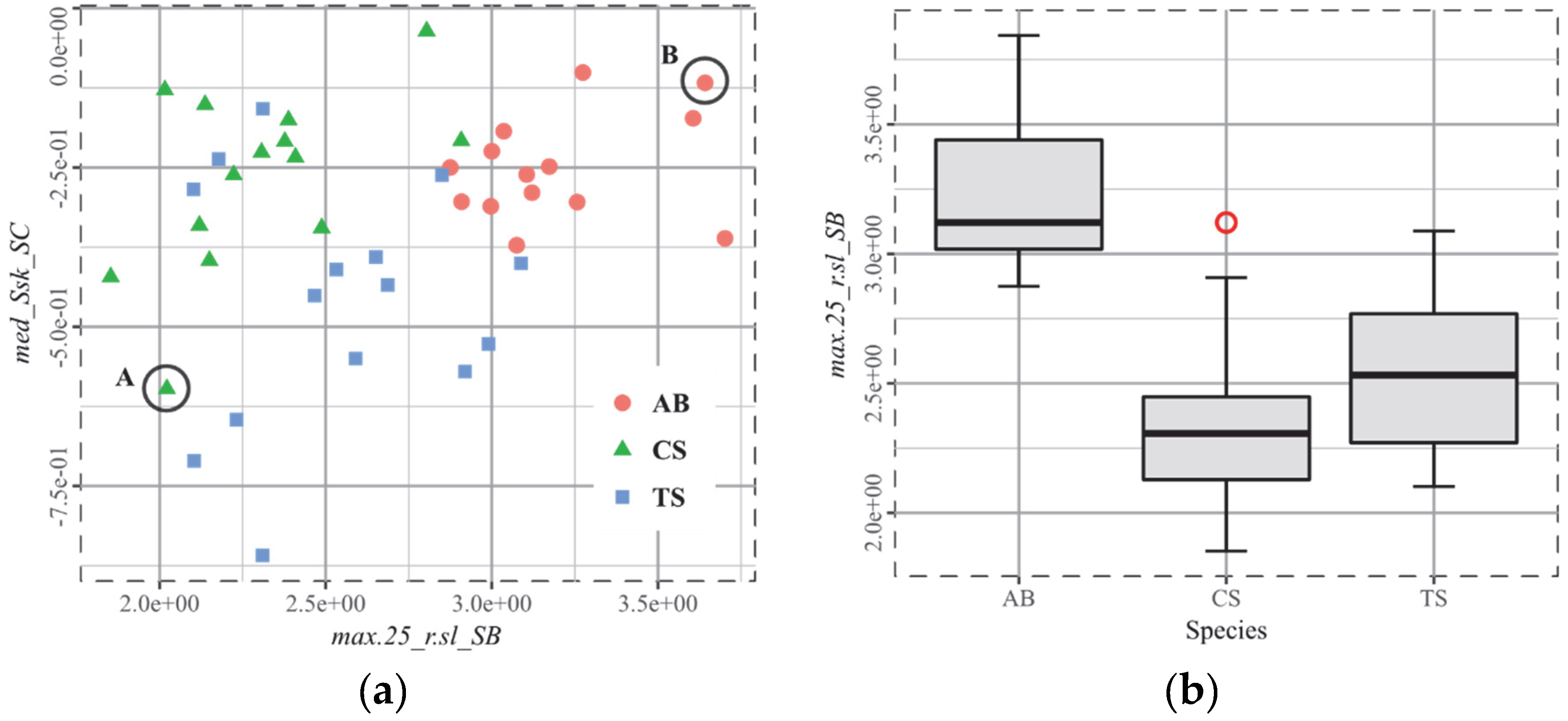
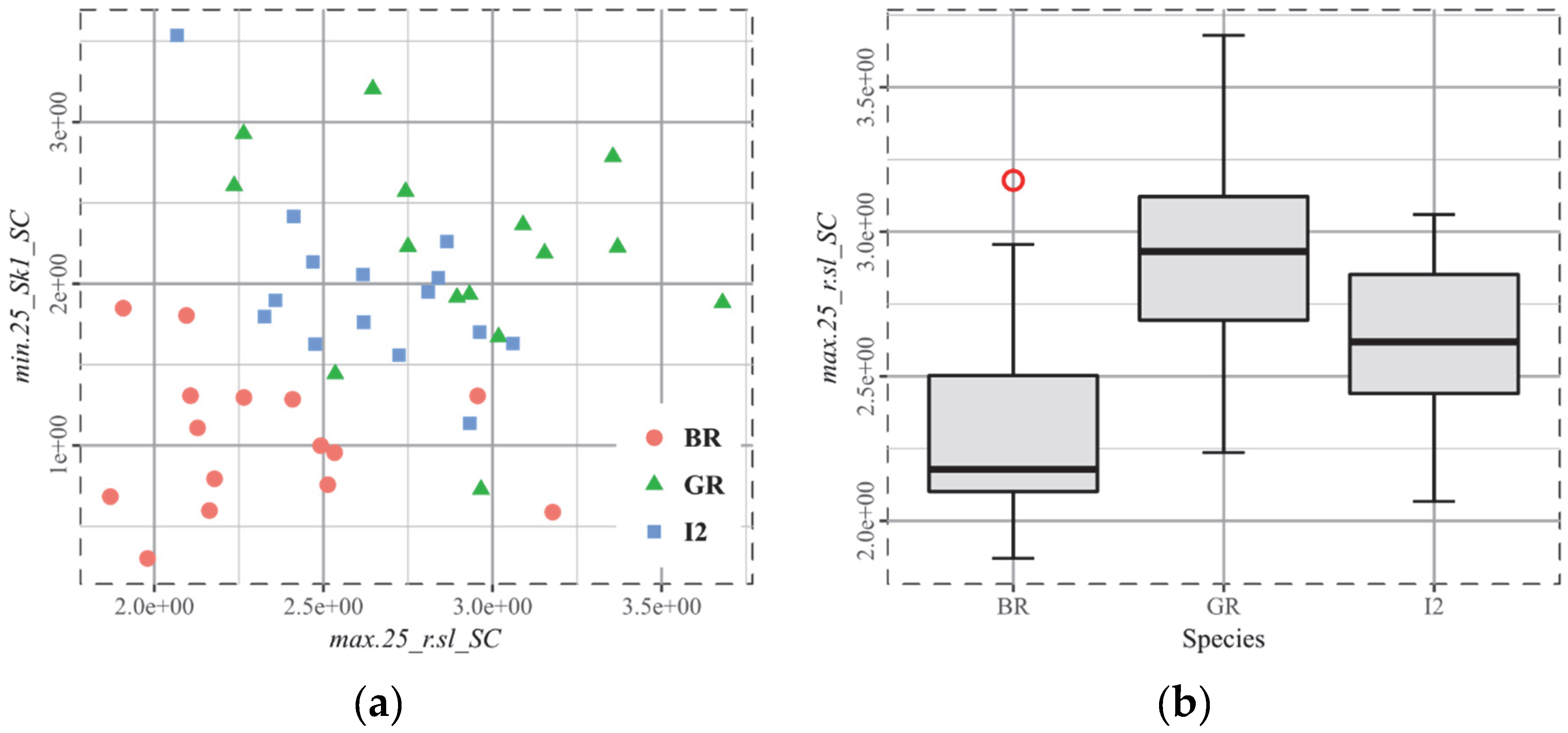
3.5. T4, Cervids
3.6. Q2, Browse, Grass and Dust (Sheep Experiment)
3.7. T5, Seeds, Browse, and Grass (Sheep Experiment)
4. Discussion
4.1. Untangling the Jungle of Parameters
4.2. The Derived Parameters Outclass the Whole-Surface Parameters
- Subtracting a second order polynomial does not alter the wear information: the geometry removed is not related to microwear. Things are different for an eighth order polynomial that can remove microwear patterns, but it also brings out details that are hidden in longer wavelength reliefs.
- When a given parameter, Stri, for instance, applied to the whole surface hardly separates two groups, its derived parameters, fst.25_Stri_SC for instance, are likely to perform a clearer separation. Besides this, the smallest p-values determined for the seven cases are all related to derived parameters. Thus, the quantiles 5%, 25%, 75%, and 95% are considered as “contrast mediums”. However, the standard deviation plays another role, especially when being applied to the texture direction Std. It then quantifies the different anisotropy orientations on a surface.
4.3. The Biplot as the Arbitor
4.4. Beware the Overfitting
4.5. Going Further
- If the final goal is to be able to classify an unknown species based upon tooth surface characteristics, then machine learning will be the first step towards an appropriate answer. Supposing that an Artificial Neuron Network (ANN) is chosen for that task and provided a large training set of surfaces of several thousands, and a hundred parameters, the machine will succeed in classifying a given surface in the right diet group. Actually, the surface will be located across all of the diet groups together with a probability for each. It can be interpreted as a signal amount. For instance, if the classifying procedure results in 70% browser, 25% grazer, and 5% fruit or seed eater, if the training set is large and variate, it can be proposed that the individual fed on 70% smooth grass (or leaves), 25% tough grass, and 5% fruit (or seeds).Going even further, the deep learning is able to identify elementary surface features in its neuron deep layers. These are the elementary signatures of the different diets. Provided that it is possible to associate a given surface to the diet content, with in vivo experiments, for example, and that the training set is large enough, the ANN could propose a typical bolus content for an unknown surface. This second point is impractical for the moment because of the lack of precise information on diets, but the first is possible. It “only” needs the researchers to share a common surface database.The main drawback is that the whole process is a black box, the results are less prone to physical interpretation.
- If the final goal is to find the parameters that accurately separate already identified groups and to understand why this is, further developments are less ambitious and they have been mentioned in the first part of the discussion.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Technique and Application in Dental Anthropology; Irish, J.D., Nelson, G.C., Eds.; Cambridge Studies in Biological and Evolutionary Anthropology; Cambridge University Press: Cambridge, UK, 2008; ISBN 978-0-521-87061-0. [Google Scholar]
- Ungar, P.S. Mammalian dental function and wear: A review. Biosurf. Biotribol. 2015, 1, 25–41. [Google Scholar] [CrossRef]
- DeSantis, L.R.G. Dental microwear textures: Reconstructing diets of fossil mammals. Surf. Topogr. Metrol. Prop. 2016, 4, 023002. [Google Scholar] [CrossRef]
- Mihlbachler, M.C.; Beatty, B.L.; Caldera-Siu, A.; Chan, D.; Lee, R. Error rates and observer bias in dental microwear analysis using light microscopy. Palaeontol. Electron. 2012, 15, 12A. [Google Scholar]
- Mihlbachler, M.C.; Beatty, B.L. Magnification and resolution in dental microwear analysis using light microscopy. Palaeontol. Electron. 2012, 15, 25A. [Google Scholar]
- Merceron, G.; Costeur, L.; Maridet, O.; Ramdarshan, A.; Göhlich, U.B. Multi-proxy approach detects heterogeneous habitats for primates during the Miocene climatic optimum in Central Europe. J. Hum. Evol. 2012, 63, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Ramdarshan, A.; Merceron, G.; Marivaux, L. Spatial and temporal ecological diversity amongst eocene primates of France: Evidence from teeth. Am. J. Phys. Anthropol. 2012, 147, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Semprebon, G.M.; Solounias, N.; Tao, D. Dietary reconstruction of Hezhengia bohlini (Artiodactyla, Bovidae) from the late Miocene Linxia Basin of China using enamel microwear. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 481, 57–63. [Google Scholar] [CrossRef]
- Xafis, A.; Nagel, D.; Bastl, K. Which tooth to sample? A methodological study of the utility of premolar/non-carnassial teeth in the microwear analysis of mammals. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 487, 229–240. [Google Scholar] [CrossRef]
- Percher, A.M.; Romero, A.; Galbany, J.; Akoue, G.N.; Pérez-Pérez, A.; Charpentier, M.J.E. Buccal dental-microwear and dietary ecology in a free-ranging population of mandrills (Mandrillus sphinx) from southern Gabon. PLoS ONE 2017, 12, e0186870. [Google Scholar] [CrossRef] [PubMed]
- Teaford, M.F.; Ungar, P.S.; Taylor, A.B.; Ross, C.F.; Vinyard, C.J. In vivo rates of dental microwear formation in laboratory primates fed different food items. Biosurf. Biotribol. 2017. [Google Scholar] [CrossRef]
- Henton, E.; Martin, L.; Garrard, A.; Jourdan, A.-L.; Thirlwall, M.; Boles, O. Gazelle seasonal mobility in the Jordanian steppe: The use of dental isotopes and microwear as environmental markers, applied to Epipalaeolithic Kharaneh IV. J. Archaeol. Sci. Rep. 2017, 11, 147–158. [Google Scholar] [CrossRef]
- Berlioz, E.; Kostopoulos, D.S.; Blondel, C.; Merceron, G. Feeding ecology of Eucladoceros ctenoides as a proxy to track regional environmental variations in Europe during the early Pleistocene. Comptes Rendus Palevol 2017. [Google Scholar] [CrossRef]
- Green, J.L.; DeSantis, L.R.G.; Smith, G.J. Regional variation in the browsing diet of Pleistocene Mammut americanum (Mammalia, Proboscidea) as recorded by dental microwear textures. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 487, 59–70. [Google Scholar] [CrossRef]
- Smith, G.J.; Desantis, L.R.G. Dietary ecology of Pleistocene mammoths and mastodons as inferred from dental microwear textures. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Janis, C.M.; Goodall, R.H.; Purnell, M.A. An examination of feeding ecology in Pleistocene proboscideans from southern China (Sinomastodon, Stegodon, Elephas), by means of dental microwear texture analysis. Q. Int. 2017, 445, 60–70. [Google Scholar] [CrossRef]
- Mahoney, P.; Schmidt, C.W.; Deter, C.; Remy, A.; Slavin, P.; Johns, S.E.; Miszkiewicz, J.J.; Nystrom, P. Deciduous enamel 3D microwear texture analysis as an indicator of childhood diet in medieval Canterbury, England. J. Archaeol. Sci. 2016, 66, 128–136. [Google Scholar] [CrossRef]
- De Bonis, L.; Abella, J.; Merceron, G.; Begun, D.R. A new late Miocene ailuropodine (Giant Panda) from Rudabánya (North-central Hungary). Geobios 2017, 50, 413–421. [Google Scholar] [CrossRef]
- Calandra, I.; Labonne, G.; Schulz-Kornas, E.; Kaiser, T.M.; Montuire, S. Tooth wear as a means to quantify intra-specific variations in diet and chewing movements. Sci. Rep. 2016, 6, 34037. [Google Scholar] [CrossRef] [PubMed]
- Purnell, M.A.; Goodall, R.H.; Thomson, S.; Matthews, C.J.D. Tooth microwear texture in odontocete whales: Variation with tooth characteristics and implications for dietary analysis. Biosurf. Biotribol. 2017, 3, 184–195. [Google Scholar] [CrossRef]
- Kubo, M.O.; Yamada, E.; Kubo, T.; Kohno, N. Dental microwear texture analysis of extant sika deer with considerations on inter-microscope variability and surface preparation protocols. Biosurf. Biotribol. 2017, 3, 155–165. [Google Scholar] [CrossRef]
- Najjar, D.; Bigerelle, M.; Iost, A. The computer-based bootstrap method as a tool to select a relevant surface roughness parameter. Wear 2003, 254, 450–460. [Google Scholar] [CrossRef]
- Najjar, D.; Bigerelle, M.; Migaud, H.; Iost, A. About the relevance of roughness parameters used for characterizing worn femoral heads. Tribol. Int. 2006, 39, 1527–1537. [Google Scholar] [CrossRef]
- Bigerelle, M.; Najjar, D.; Mathia, T.; Iost, A.; Coorevits, T.; Anselme, K. An expert system to characterise the surfaces morphological properties according to their tribological functionalities: The relevance of a pair of roughness parameters. Tribol. Int. 2013, 59, 190–202. [Google Scholar] [CrossRef]
- Deltombe, R.; Kubiak, K.J.; Bigerelle, M. How to select the most relevant 3D roughness parameters of a surface. Scanning 2014, 36, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Francisco, A.; Blondel, C.; Brunetière, N.; Ramdarshan, A.; Merceron, G. Enamel surface topography analysis for diet discrimination. A methodology to enhance and select discriminative parameters. Surf. Topogr. Metrol. Prop. 2017. [Google Scholar] [CrossRef]
- Merceron, G.; Ramdarshan, A.; Blondel, C.; Boisserie, J.-R.; Brunetiere, N.; Francisco, A.; Gautier, D.; Milhet, X.; Novello, A.; Pret, D. Untangling the environmental from the dietary: Dust does not matter. Proc. Biol. Sci. 2016, 283. [Google Scholar] [CrossRef] [PubMed]
- Merceron, G.; Blondel, C.; Brunetiere, N.; Francisco, A.; Gautier, D.; Ramdarshan, A. Dental microwear and controlled food testing on sheep: The TRIDENT project. Biosurf. Biotribol. 2017, 3, 174–183. [Google Scholar] [CrossRef]
- Ramdarshan, A.; Blondel, C.; Gautier, D.; Surault, J.; Merceron, G. Overcoming sampling issues in dental tribology: Insights from an experimentation on sheep. Palaeontol. Electron. 2017, 20, 1–19. [Google Scholar] [CrossRef]
- Lambert, J.E.; Chapman, C.A.; Wrangham, R.W.; Conklin-Brittain, N.L. Hardness of cercopithecine foods: Implications for the critical function of enamel thickness in exploiting fallback foods. Am. J. Phys. Anthropol. 2004, 125, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Daegling, D.J.; McGraw, W.S.; Ungar, P.S.; Pampush, J.D.; Vick, A.E.; Bitty, E.A. Hard-object feeding in sooty mangabeys (Cercocebus atys) and interpretation of early hominin feeding ecology. PLoS ONE 2011, 6, e23095. [Google Scholar] [CrossRef] [PubMed]
- Maisels, F.; Gautier-Hion, A.; Gautier, J.-P. Diets of two sympatric colobines in Zaire: More evidence on seed-eating in forests on poor soils. Int. J. Primatol. 1994, 15, 681. [Google Scholar] [CrossRef]
- Daegling, D.J.; McGraw, W.S. Feeding, Diet, and Jaw Form in West African Colobus and Procolobus. Int. J. Primatol. 2001, 22, 1033–1055. [Google Scholar] [CrossRef]
- Post, D.G.; Hausfater, G.; McCuskey, S. Feeding Behavior of Yellow Baboons (Papio cynocephalus): Relationship to Age, Gender and Dominance Rank. Folia Primatol. 1980, 34, 170–195. [Google Scholar] [CrossRef] [PubMed]
- Rhine, R.J.; Norton, G.W.; Wynn, G.M.; Wynn, R.D.; Rhine, H.B. Insect and meat eating among infant and adult baboons (Papio cynocephalus) of Mikumi National Park, Tanzania. Am. J. Phys. Anthropol. 1986, 70, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.S.; Teaford, M.F.; Ungar, P.S. Dental microwear texture and anthropoid diets. Am. J. Phys. Anthropol. 2012, 147, 551–579. [Google Scholar] [CrossRef] [PubMed]
- Merceron, G.; Hofman-Kamińska, E.; Kowalczyk, R. 3D dental microwear texture analysis of feeding habits of sympatric ruminants in the Białowieża Primeval Forest, Poland. For. Ecol. Manag. 2014, 328, 262–269. [Google Scholar] [CrossRef]
- Massey, F.; Smith, M.; Lambin, X.; Hartley, S. Are silica defences in grasses driving vole population cycles? Biol. Lett. 2008, 4, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.S.; Ungar, P.S.; Bergstrom, T.S.; Brown, C.A.; Childs, B.E.; Teaford, M.F.; Walker, A. Dental microwear texture analysis: Technical considerations. J. Hum. Evol. 2006, 51, 339–349. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Significance |
|---|---|
| Sa | Arithmetic mean of the absolute of the heights (*) |
| Sp | Absolute of the largest height (*) |
| Sq | Height standard deviation (*) |
| Sv | Absolute of the smallest height (*) |
| Ssk | Height skewness (*) |
| Sku | Height kurtosis (*) |
| Sdar | Relative area (developed area/projected area) |
| Sm | Mean height (0 for the whole surface, but non-zero for its samples) |
| Smd | Median height |
| Parameter | Significance |
|---|---|
| Rmax | Semi-major axis of the fACF ellipsis |
| Sal | Semi-minor axis of the fACF ellipsis (*) |
| Std | Texture direction (*) |
| Stri = Str−1 | Rmax/Sal ratio (*) |
| b.sl | Highest slope of fACF at the distance rs from the origin |
| r.sl | b.sl/s.sl ratio |
| s.sl | Smallest slope of fACF at the distance rs from the origin |
| Parameter | Significance |
|---|---|
| Sk1, Sk2 | Relative area of the surface above h1 and h2 respectively |
| Smc1, Smc2 | Median relative area of the cells with heights exceeding h1 and h2 respectively |
| Snb1, Snb2 | Number of cells with heights exceeding h1 and h2 respectively |
| Sh | Percentage of quasi-horizontal faces (normal within a 4° cone) |
| Sku | Sk1 | Scm1 | Sk2 | Smd | Std |
| 1.47 × 10−4 | 2.65 × 10−4 | 4.04 × 10−4 | 5.13 × 10−4 | 6.79 × 10−4 | 7.75 × 10−4 |
| Ssk | r.sl | Scm2 | Snb2 | Sh | Rmax |
| 1.06 × 10−3 | 1.07 × 10−3 | 1.17 × 10−3 | 1.21 × 10−3 | 1.35 × 10−3 | 2.06 × 10−3 |
| Stri | Sv | Sdar | Asfc | Snb1 | Sp |
| 2.25 × 10−3 | 2.44 × 10−3 | 3.16 × 10−3 | 3.50 × 10−3 | 3.86 × 10−3 | 5.13 × 10−3 |
| Sa | Sal | Sm | Sq | s.sl | b.sl |
| 6.70 × 10−3 | 7.96 × 10−3 | 8.62 × 10−3 | 8.67 × 10−3 | 4.91 × 10−2 | 1.31 × 10−1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francisco, A.; Brunetière, N.; Merceron, G. Gathering and Analyzing Surface Parameters for Diet Identification Purposes. Technologies 2018, 6, 75. https://doi.org/10.3390/technologies6030075
Francisco A, Brunetière N, Merceron G. Gathering and Analyzing Surface Parameters for Diet Identification Purposes. Technologies. 2018; 6(3):75. https://doi.org/10.3390/technologies6030075
Chicago/Turabian StyleFrancisco, Arthur, Noël Brunetière, and Gildas Merceron. 2018. "Gathering and Analyzing Surface Parameters for Diet Identification Purposes" Technologies 6, no. 3: 75. https://doi.org/10.3390/technologies6030075
APA StyleFrancisco, A., Brunetière, N., & Merceron, G. (2018). Gathering and Analyzing Surface Parameters for Diet Identification Purposes. Technologies, 6(3), 75. https://doi.org/10.3390/technologies6030075