
Is Early Bilingual Experience Associated with Greater Fluid Intelligence in Adults?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Materials
2.2.1. Background Questionnaire
2.2.2. Raven Advanced Progressive Matrices—Set I
2.3. Procedure
2.4. Ethical Approval
3. Results
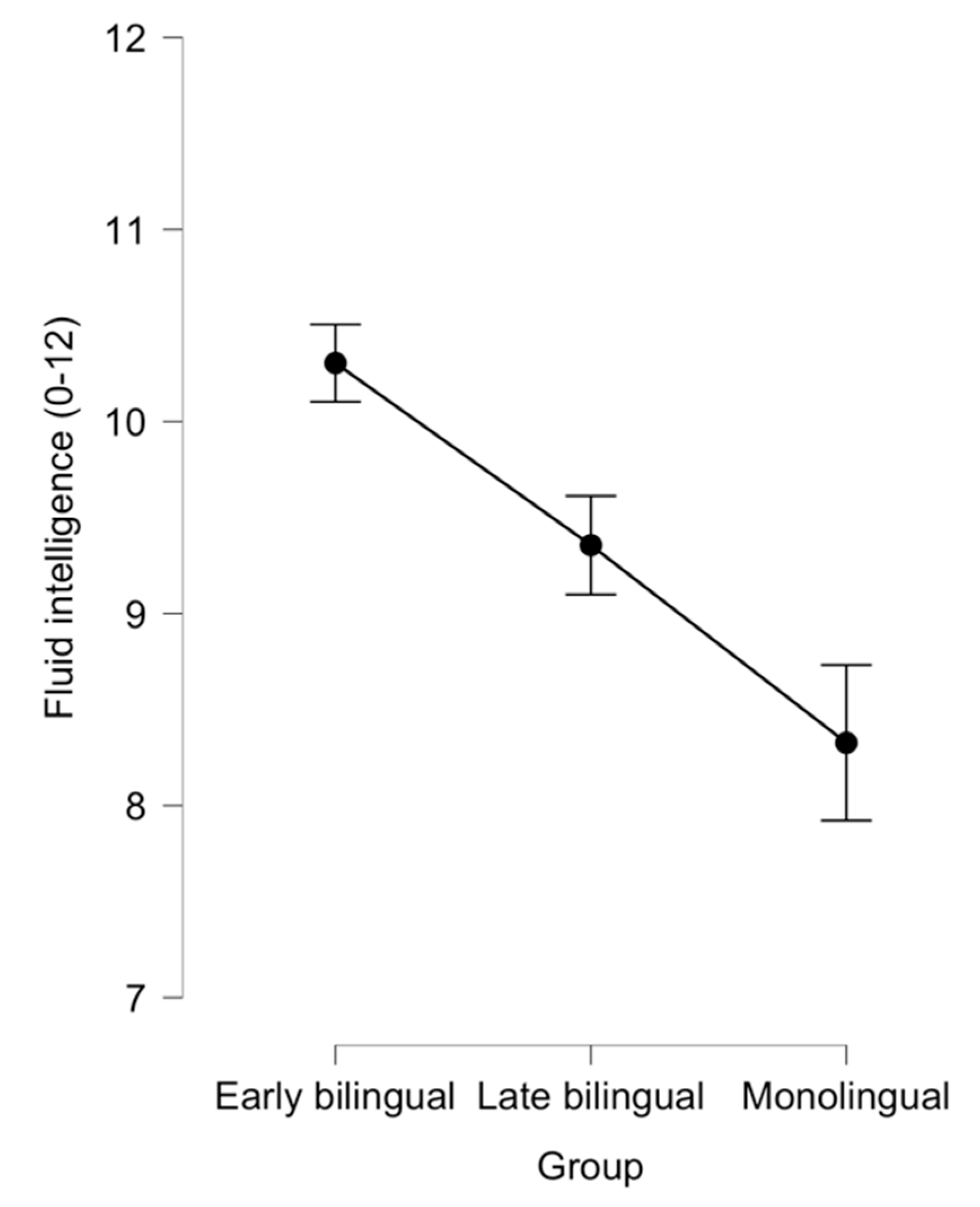
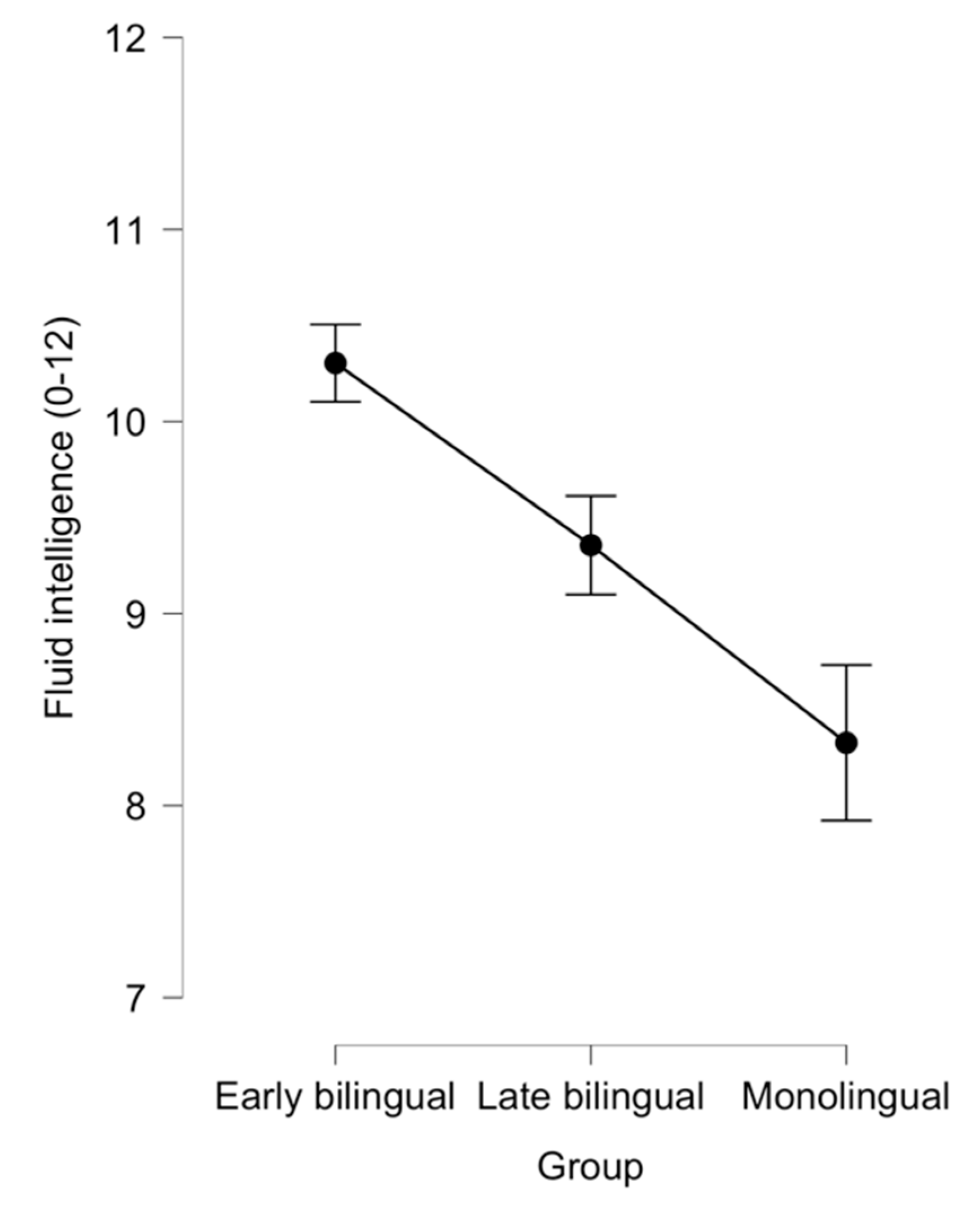
3.1. Comparing Early Bilinguals with Late Bilinguals and Monolinguals
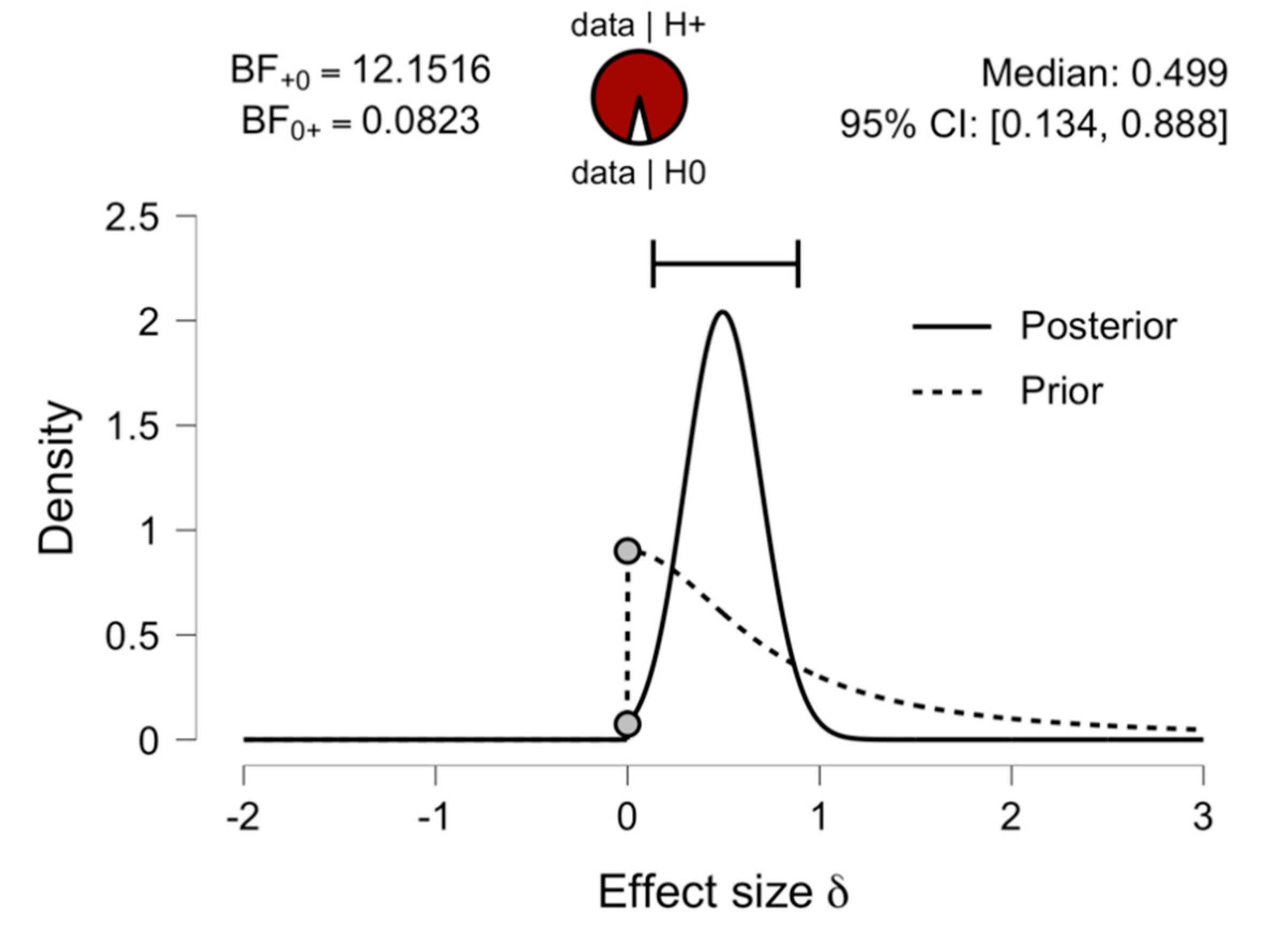
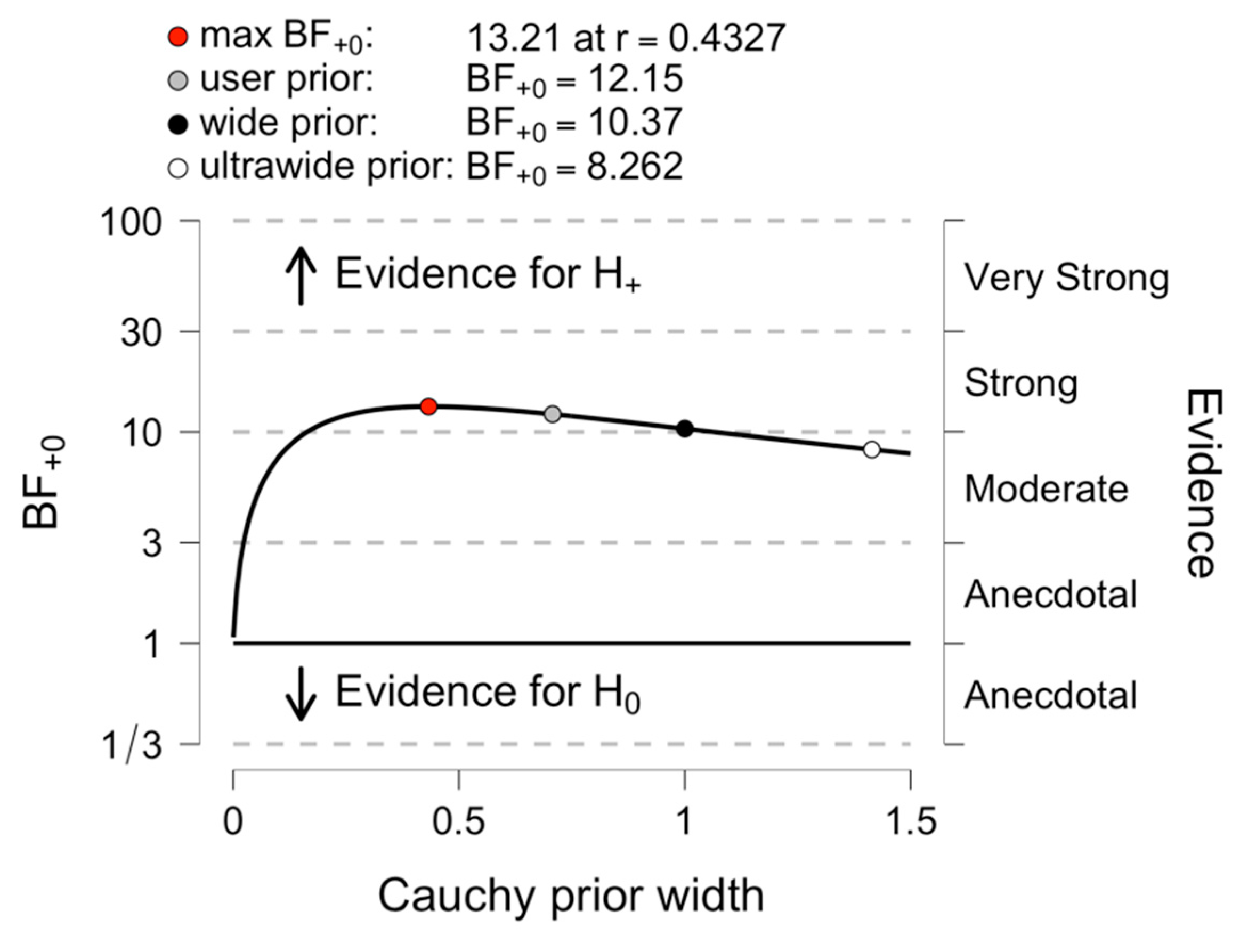
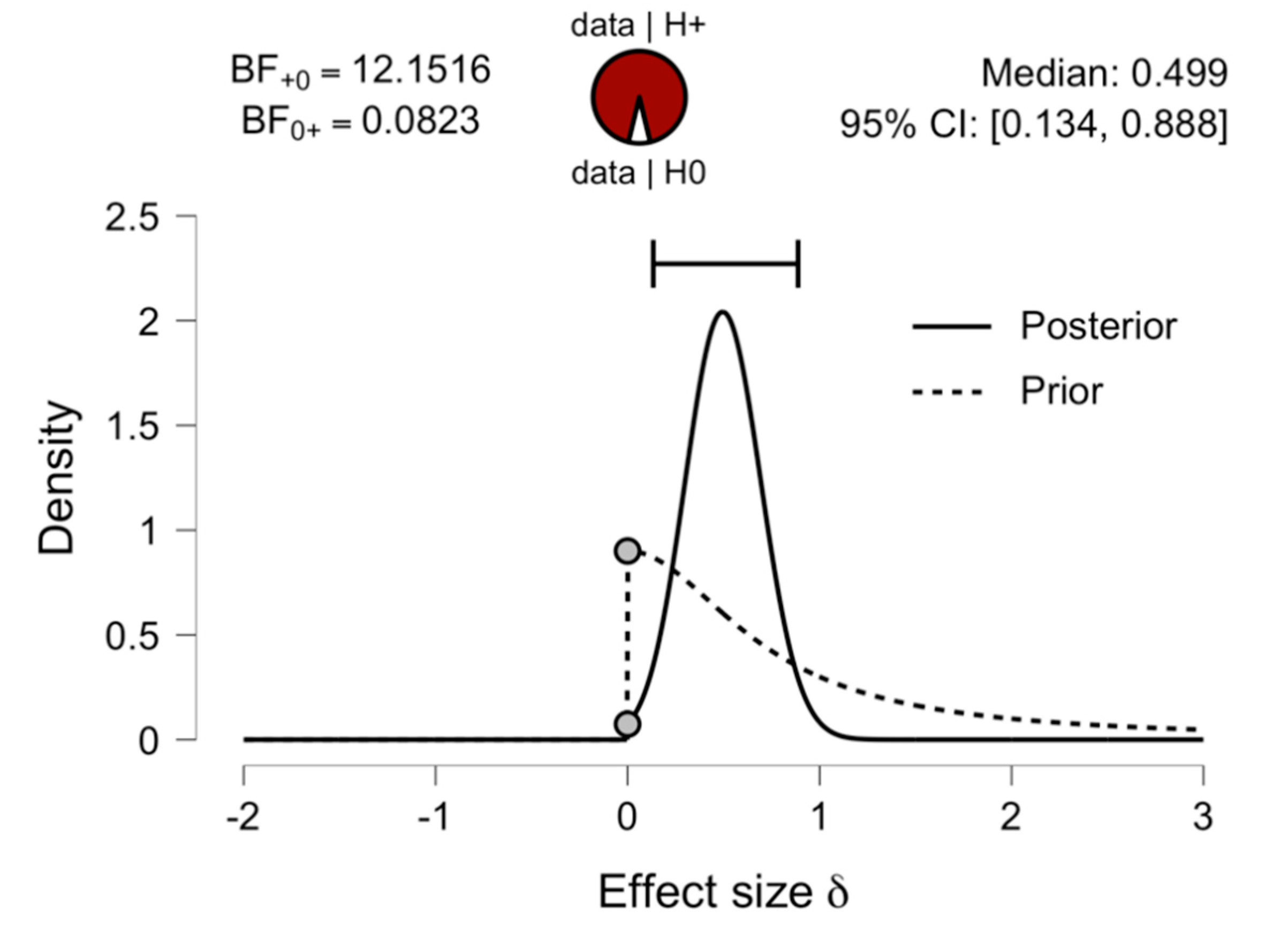
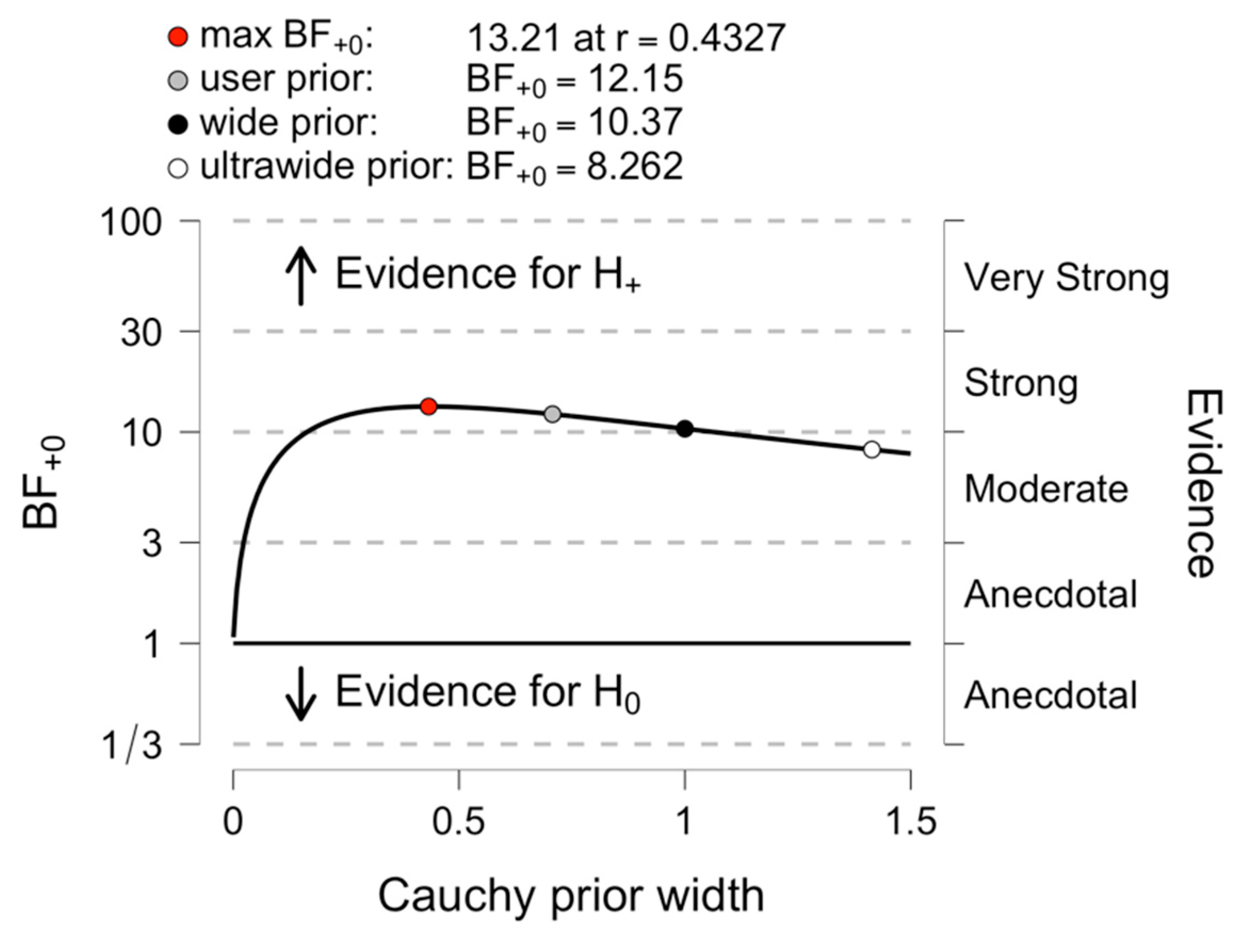
3.2. Comparing Early and Late Bilinguals: A Bayesian Analysis
3.3. Exploratory Analyses
4. Discussion
Limitations and Future Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| 1 | A recent review and meta-analysis of 1194 effect sizes from studies that included 10,937 bilinguals and 12,477 monolinguals between 3 and 17 years of age found no effect of language status (bilingual vs. monolingual) on cognitive ability, after accounting for moderating factors and publication bias (Lowe et al. 2021). The meta-analysis did not compare early bilinguals with late bilinguals. |
| 2 | Although it is possible that a dedicated control system inhibits the activation of words in the non-target language (Green 1998), such a system is not necessary. It is possible that greater activation of task-relevant words attenuates activation of task-irrelevant words. This could be achieved through simple lateral inhibition mechanisms, whereby the synchronous activation of one neural population automatically disrupts the synchronous activation of nearby populations (Buzsaki 2006). These lateral inhibition mechanisms are ubiquitous in the human brain and may help to prevent task-irrelevant interference (Buzsaki 2006). In other words, control processes and attentional processes may not be two independent systems but two aspects of one complex process involving highly diverse, interconnected, and interdependent neurons. |
References
- Antoniou, Mark, and Sarah M. Wright. 2017. Uncovering the mechanisms responsible for why language learning may promote healthy cognitive aging. Frontiers in Psychology 15: 2217. [Google Scholar] [CrossRef] [Green Version]
- Anwyl-Irvine, Alexander L., Jessica Massonnié, Adam Flitton, Natasha Kirkham, and Jo K. Evershed. 2020. Gorilla in our midst: An online behavioral experiment builder. Behavior Research Methods 52: 388–407. [Google Scholar] [CrossRef] [Green Version]
- Baddeley, Alan D., and Graham Hitch. 1974. Working memory. In Psychology of Learning and Motivation. New York: Academic Press, vol. 8, pp. 47–89. [Google Scholar]
- Bail, Amelie, Giovanna Morini, and Rochelle S. Newma. 2015. Look at the gato! Code-switching in speech to toddlers. Journal of Child Language 42: 1073. [Google Scholar] [CrossRef] [Green Version]
- Basten, Ulrike, Kirsten Hilger, and Christian J. Fiebach. 2015. Where smart brains are different: A quantitative meta-analysis of functional and structural brain imaging studies on intelligence. Intelligence 51: 10–27. [Google Scholar] [CrossRef]
- Berken, Jonathan A., Vincent L. Gracco, Jen-Kai Chen, and Denise Klein. 2015. The timing of language learning shapes brain structure associated with articulation. Brain Structure and Function 221: 3591–600. [Google Scholar] [CrossRef]
- Berken, Jonathan A., Xiaoqian Chai, Jen-Kai Chen, Vincent L. Gracco, and Denise Klein. 2016. Effects of early and late bilingualism on resting-state functional connectivity. Journal of Neuroscience 36: 1165–72. [Google Scholar] [CrossRef] [Green Version]
- Borragan, Maria, Clara D. Martin, Angela De Bruin, and Jon Andoni Duñabeitia. 2018. Exploring different types of inhibition during bilingual language production. Frontiers in Psychology 9: 2256. [Google Scholar] [CrossRef]
- Bosch, Laura, and Marta Ramon-Casas. 2011. Variability in vowel production by bilingual speakers: Can input properties hinder the early stabilization of contrastive categories? Journal of Phonetics 39: 514–26. [Google Scholar] [CrossRef]
- Braver, Todd S. 2012. The variable nature of cognitive control: A dual mechanisms framework. Trends in Cognitive Sciences 16: 106–13. [Google Scholar] [CrossRef] [Green Version]
- Braver, Todd S., Jeremy R. Gray, and Gregory C. Burgess. 2007. Explaining the many varieties of working memory variation: Dual mechanisms of cognitive control. Variation in Working Memory 75: 106. [Google Scholar]
- Buzsaki, Gyorgy. 2006. Rhythms of the Brain. Oxford: Oxford University Press. [Google Scholar]
- Byers-Heinlein, Krista, Elizabeth Morin-Lessard, and Casey Lew-Williams. 2017. Bilingual infants control their languages as they listen. Proceedings of the National Academy of Sciences 114: 9032–37. [Google Scholar] [CrossRef] [Green Version]
- Chein, Jason M., and Walter Schneider. 2005. Neuroimaging studies of practice-related change: fMRI and meta-analytic evidence of a domain-general control network for learning. Cognitive Brain Research 25: 607–23. [Google Scholar] [CrossRef]
- Chein, Jason M., and Walter Schneider. 2012. The brain’s learning and control architecture. Current Directions in Psychological Science 21: 78–84. [Google Scholar] [CrossRef]
- Chiesi, Francesca, Matteo Ciancaleoni, Silvia Galli, and Caterina Primi. 2012. Using the Advanced Progressive Matrices (Set I) to assess fluid ability in a short time frame: An item response theory–based analysis. Psychological Assessment 24: 892. [Google Scholar] [CrossRef]
- Cipolotti, Lisa, Barbara Spanò, Colm Healy, Carina Tudor-Sfetea, Edgar Chan, Mark White, Francesca Biondo, John Duncan, Tim Shallice, and Marco Bozzali. 2016. Inhibition processes are dissociable and lateralized in human prefrontal cortex. Neuropsychologia 93: 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cocchi, Luca, Graeme S. Halford, Andrew Zalesky, Ian H. Harding, Brentyn J. Ramm, Tim Cutmore, David HK Shum, and Jason B. Mattingley. 2014. Complexity in relational processing predicts changes in functional brain network dynamics. Cerebral Cortex 24: 2283–96. [Google Scholar] [CrossRef] [Green Version]
- Cole, Michael W., Takuya Ito, and Todd S. Braver. 2015. Lateral prefrontal cortex contributes to fluid intelligence through multinetwork connectivity. Brain Connectivity 5: 497–504. [Google Scholar] [CrossRef] [Green Version]
- Colom, Roberto, Miguel Burgaleta, Francisco J. Román, Sherif Karama, Juan Álvarez-Linera, Francisco J. Abad, Kenia Martínez, Ma Ángeles Quiroga, and Richard J. Haier. 2013. Neuroanatomic overlap between intelligence and cognitive factors: Morphometry methods provide support for the key role of the frontal lobes. Neuroimage 72: 143–52. [Google Scholar] [CrossRef]
- Conway, Andrew R. A., Nelson Cowan, Michael F. Bunting, David J. Therriault, and Scott R. B. Minkoff. 2002. A latent variable analysis of working memory capacity, short-term memory capacity, processing speed, and general fluid intelligence. Intelligence 30: 163–83. [Google Scholar] [CrossRef]
- Corbetta, Maurizio, and Gordon L. Shulman. 2002. Control of goal-directed and stimulus-driven attention in the brain. Nature Reviews Neuroscience 3: 201–15. [Google Scholar] [CrossRef]
- Corbetta, Maurizio, Gaurav Patel, and Gordon L. Shulman. 2008. The reorienting system of the human brain: From environment to theory of mind. Neuron 58: 306–24. [Google Scholar] [CrossRef] [Green Version]
- Cowan, Nelson. 1995. Attention and Memory: An Integrative Framework. Oxford: Oxford University Press. [Google Scholar]
- Cowan, Nelson. 1999. An embedded-processes model of working memory. In Models of Working Memory: Mechanisms of Active Maintenance and Executive Control. Edited by Akira Miyake and Priti Shah. Cambridge: Cambridge University Press, pp. 62–101. [Google Scholar]
- Cowan, Nelson. 2010. The magical mystery four: How is working memory capacity limited, and why? Current Directions in Psychological Science 19: 51–57. [Google Scholar] [CrossRef] [Green Version]
- Cowan, Nelson, Nathanael M. Fristoe, Emily M. Elliott, Ryan P. Brunner, and J. Scott Saults. 2006. Scope of attention, control of attention, and intelligence in children and adults. Memory & Cognition 34: 1754–68. [Google Scholar]
- D’Souza, Dean, and Hana D’Souza. 2016. Bilingual language control mechanisms in anterior cingulate cortex and dorsolateral prefrontal cortex: A developmental perspective. Journal of Neuroscience 36: 5434–36. [Google Scholar] [CrossRef]
- D’Souza, Dean, and Hana D’Souza. 2021. Bilingual adaptations in early development. Trends in Cognitive Sciences 25: 727–29. [Google Scholar] [CrossRef]
- D’Souza, Dean, Dan Brady, Jennifer X. Haensel, and Hana D’Souza. 2020. Is mere exposure enough? The effects of bilingual environments on infant cognitive development. Royal Society Open Science 7: 180191. [Google Scholar] [CrossRef]
- D’Souza, Dean, Dan Brady, Jennifer X. Haensel, and Hana D’Souza. 2021. Early bilingual experience is associated with change detection ability in adults. Scientific Reports 11: 1–9. [Google Scholar] [CrossRef]
- Dal Ben, Rodrigo, Hilary Killam, Sadaf Pour Iliaei, and Krista Byers-Heinlein. 2021. Bilingualism Affects Infant Cognition: Insights from New and Open Data. PsyArXiv, December 7. [Google Scholar] [CrossRef]
- Davis-Kean, Pamela E. 2005. The influence of parent education and family income on child achievement: The indirect role of parental expectations and the home environment. Journal of Family Psychology 19: 294–304. [Google Scholar] [CrossRef] [Green Version]
- Deary, Ian J., Steve Strand, Pauline Smith, and Cres Fernandes. 2007. Intelligence and educational achievement. Intelligence 35: 13–21. [Google Scholar] [CrossRef]
- Donnelly, Seamus, Patricia J. Brooks, and Bruce D. Homer. 2019. Is there a bilingual advantage on interference-control tasks? A multiverse meta-analysis of global reaction time and interference cost. Psychonomic Bulletin & Review 26: 1122–47. [Google Scholar]
- Duncan, John. 1995. Attention, intelligence and the frontal lobes. In The Cognitive Neuroscience. Edited by Michael S. Gazzaniga. Cambridge: MIT Press, pp. 721–33. [Google Scholar]
- Duncan, John. 2010. The multiple-demand (MD) system of the primate brain: Mental programs for intelligent behaviour. Trends in Cognitive Sciences 14: 172–79. [Google Scholar] [CrossRef]
- Duncan, John, and Adrian M. Owen. 2000. Common regions of the human frontal lobe recruited by diverse cognitive demands. Trends in Neurosciences 23: 475–83. [Google Scholar] [CrossRef]
- Duncan, John, Daphne Chylinski, Daniel J. Mitchell, and Apoorva Bhandari. 2017. Complexity and compositionality in fluid intelligence. Proceedings of the National Academy of Sciences 114: 5295–99. [Google Scholar] [CrossRef] [Green Version]
- Engle, Randall W., Michael J. Kane, and Stephen W. Tuholski. 1999a. Individual differences in working memory capacity and what they tell us about controlled attention, general fluid intelligence, and functions of the prefrontal cortex. Models of Working Memory: Mechanisms of Active Maintenance and Executive Control 4: 102–34. [Google Scholar]
- Engle, Randall W., Stephen W. Tuholski, James E. Laughlin, and Andrew RA Conway. 1999b. Working memory, short-term memory, and general fluid intelligence: A latent-variable approach. Journal of Experimental Psychology: General 128: 309. [Google Scholar] [CrossRef]
- Farah, Martha J. 2017. The neuroscience of socioeconomic status: Correlates, causes, and consequences. Neuron 96: 56–71. [Google Scholar] [CrossRef] [Green Version]
- Fedorenko, Evelina, John Duncan, and Nancy Kanwisher. 2013. Broad domain generality in focal regions of frontal and parietal cortex. Proceedings of the National Academy of Sciences 110: 16616–621. [Google Scholar] [CrossRef] [Green Version]
- Fiori, Valentina, Lisa Kunz, Philipp Kuhnke, Paola Marangolo, and Gesa Hartwigsen. 2018. Transcranial direct current stimulation (tDCS) facilitates verb learning by altering effective connectivity in the healthy brain. NeuroImage 181: 550–59. [Google Scholar] [CrossRef]
- Fox, Michael D., Maurizio Corbetta, Abraham Z. Snyder, Justin L. Vincent, and Marcus E. Raichle. 2006. Spontaneous neuronal activity distinguishes human dorsal and ventral attention systems. Proceedings of the National Academy of Sciences 103: 10046–51. [Google Scholar] [CrossRef] [Green Version]
- Friedman, Naomi P., and Akira Miyake. 2004. The relations among inhibition and interference control functions: A latent-variable analysis. Journal of Experimental Psychology: General 133: 101. [Google Scholar] [CrossRef] [Green Version]
- Friedman, Naomi P., Akira Miyake, Susan E. Young, John C. DeFries, Robin P. Corley, and John K. Hewitt. 2008. Individual differences in executive functions are almost entirely genetic in origin. Journal of Experimental Psychology: General 137: 201. [Google Scholar] [CrossRef] [PubMed]
- Friedman, Naomi P., Akira Miyake, JoAnn L. Robinson, and John K. Hewitt. 2011. Developmental trajectories in toddlers’ self-restraint predict individual differences in executive functions 14 years later: A behavioral genetic analysis. Developmental Psychology 47: 1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fries, Pascal. 2005. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends in Cognitive Sciences 9: 474–80. [Google Scholar] [CrossRef] [PubMed]
- Friston, Karl. 2009. The free-energy principle: A rough guide to the brain? Trends in Cognitive Sciences 13: 293–301. [Google Scholar] [CrossRef] [PubMed]
- Friston, Karl. 2010. The free-energy principle: A unified brain theory? Nature Reviews Neuroscience 11: 127–38. [Google Scholar] [CrossRef]
- Gogtay, Nitin, Jay N. Giedd, Leslie Lusk, Kiralee M. Hayashi, Deanna Greenstein, A. Catherine Vaituzis, Tom F. Nugent, David H. Herman, Liv S. Clasen, Arthur W. Toga, and et al. 2004. Dynamic mapping of human cortical development during childhood through early adulthood. Proceedings of the National Academy of Sciences 101: 8174–79. [Google Scholar] [CrossRef] [Green Version]
- Gottfried, Allen W., Adele E. Gottfried, Kay Bathurst, Diana Wright Guerin, and Makeba M. Parramore. 2003. Socioeconomic Status in Children’s Development and Family Environment: Infancy through Adolescence. Mahwah: Lawrence Erlbaum Associates Publishers. [Google Scholar]
- Grady, Cheryl L., Gigi Luk, Fergus I. M. Craik, and Ellen Bialystok. 2015. Brain network activity in monolingual and bilingual older adults. Neuropsychologia 66: 170–81. [Google Scholar] [CrossRef] [Green Version]
- Gray, Jeremy R., Christopher F. Chabris, and Todd S. Braver. 2003. Neural mechanisms of general fluid intelligence. Nature Neuroscience 6: 316–22. [Google Scholar] [CrossRef]
- Green, David W. 1998. Mental control of the bilingual lexico-semantic system. Bilingualism: Language and Cognition 1: 67–81. [Google Scholar] [CrossRef]
- Hartanto, Andree, and Hwajin Yang. 2019. Does early active bilingualism enhance inhibitory control and monitoring? A propensity-matching analysis. Journal of Experimental Psychology: Learning, Memory, and Cognition 45: 360. [Google Scholar] [CrossRef]
- Hull, Rachel, and Jyotsna Vaid. 2006. Laterality and language experience. Laterality 11: 436–64. [Google Scholar] [CrossRef] [PubMed]
- Hull, Rachel, and Jyotsna Vaid. 2007. Bilingual language lateralization: A meta-analytic tale of two hemispheres. Neuropsychologia 45: 1987–2008. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, A. D., and Virginia Valian. 2012. Multilingualism and cognitive control: Simon and flanker task performance in monolingual and multilingual young adults. Paper presented at the 53rd Annual Meeting of the Psychonomic Society, Minneapolis, MN, USA, November 15–18. [Google Scholar]
- Japee, Shruti, Kelsey Holiday, Maureen D. Satyshur, Ikuko Mukai, and Leslie G. Ungerleider. 2015. A role of right middle frontal gyrus in reorienting of attention: A case study. Frontiers in Systems Neuroscience 9: 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffreys, Harrold. 1961. Theory of Probability, 3rd ed. Oxford: Oxford University Press. [Google Scholar]
- Kalashnikova, Marina, Jovana Pejovic, and Manuel Carreiras. 2021. The effects of bilingualism on attentional processes in the first year of life. Developmental Science 24: e13011. [Google Scholar] [CrossRef]
- Kalia, Vrinda, Makeba Parramore Wilbourn, and Kathleen Ghio. 2014. Better early or late? Examining the influence of age of exposure and language proficiency on executive function in early and late bilinguals. Journal of Cognitive Psychology 26: 699–713. [Google Scholar] [CrossRef]
- Kane, Michael J., and Randall W. Engle. 2003. Working-memory capacity and the control of attention: The contributions of goal neglect, response competition, and task set to Stroop interference. Journal of Experimental Psychology: General 132: 47. [Google Scholar] [CrossRef]
- Kapa, Leah L., and John Colombo. 2013. Attentional control in early and later bilingual children. Cognitive Development 28: 233–46. [Google Scholar] [CrossRef] [Green Version]
- Karr, Justin E., Corson N. Areshenkoff, Philippe Rast, Scott M. Hofer, Grant L. Iverson, and Mauricio A. Garcia-Barrera. 2018. The unity and diversity of executive functions: A systematic review and re-analysis of latent variable studies. Psychological Bulletin 144: 1147. [Google Scholar] [CrossRef] [PubMed]
- Kass, Robert E., and Adrian E. Raftery. 1995. Bayes factors. Journal of the American Statistical Association 90: 773–95. [Google Scholar] [CrossRef]
- Kremin, Lena V., Julia Alves, Adriel John Orena, Linda Polka, and Krista Byers-Heinlein. 2021. Code-switching in parents’ everyday speech to bilingual infants. Journal of Child Language, 1–27. [Google Scholar] [CrossRef]
- Kyllonen, Patrick C., and Raymond E. Christal. 1990. Reasoning ability is (little more than) working-memory capacity?! Intelligence 14: 389–433. [Google Scholar] [CrossRef]
- Li, Shifeng, Qiongying Xu, and Ruixue Xia. 2020. Relationship between SES and academic achievement of junior high school students in China: The mediating effect of self-concept. Frontiers in Psychology 10: 2513. [Google Scholar] [CrossRef] [PubMed]
- Linck, Jared A., Peter Osthus, Joel T. Koeth, and Michael F. Bunting. 2014. Working memory and second language comprehension and production: A meta-analysis. Psychonomic Bulletin & Review 21: 861–83. [Google Scholar]
- Liu, Juan, Peng Peng, and Liang Luo. 2020. The relation between family socioeconomic status and academic achievement in China: A meta-analysis. Educational Psychology Review 32: 49–76. [Google Scholar] [CrossRef]
- Lowe, Cassandra J., Isu Cho, Samantha F. Goldsmith, and J. Bruce Morton. 2021. The bilingual advantage in children’s executive functioning is not related to language status: A meta-analytic review. Psychological Science 32: 1115–46. [Google Scholar] [CrossRef] [PubMed]
- Luk, Gigi, Eric De Sa, and Ellen Bialystok. 2011. Is there a relation between onset age of bilingualism and enhancement of cognitive control? Bilingualism: Language and Cognition 14: 588–95. [Google Scholar] [CrossRef]
- Mohades, Seyede Ghazal, Esli Struys, Peter Van Schuerbeek, Katrien Mondt, Piet Van De Craen, and Robert Luypaert. 2012. DTI reveals structural differences in white matter tracts between bilingual and monolingual children. Brain Research 1435: 72–80. [Google Scholar] [CrossRef]
- Neisser, Ulric, Gwyneth Boodoo, Thomas J. Bouchard Jr., A. Wade Boykin, Nathan Brody, Stephen J. Ceci, Diane F. Halpern, John C. Loehlin, Robert Perloff, Robert J. Sternberg, and et al. 1996. Intelligence: Knowns and unknowns. American Psychologist 51: 77. [Google Scholar] [CrossRef]
- Norman, Donald A., and Tim Shallice. 1986. Attention to action: Willed and automatic control of behavior. In Consciousness and Self-Regulation. Edited by Richard J. Davidson, Gary E. Schwartz and David Shapi. Boston: Springer, pp. 1–18. [Google Scholar]
- Paap, Kenneth R., Hunter A. Johnson, and Oliver Sawi. 2014. Are bilingual advantages dependent upon specific tasks or specific bilingual experiences? Journal of Cognitive Psychology 26: 615–39. [Google Scholar] [CrossRef]
- Pelham, Sabra D., and Lise Abrams. 2014. Cognitive advantages and disadvantages in early and late bilinguals. Journal of Experimental Psychology: Learning, Memory, and Cognition 40: 313. [Google Scholar] [CrossRef] [Green Version]
- Raftery, Adrian E. 1995. Bayesian model selection in social research. Sociological Methodology 25: 111–63. [Google Scholar] [CrossRef]
- Raven, John C., John H. Court, and Jean Raven. 1988. Advanced Progressive Matrices. Raven Manual. Oxford: Oxford Psychologists Press. [Google Scholar]
- Raven, Jean, John C. Raven, and John H. Court. 2003. Manual for Raven’s Progressive Matrices and Vocabulary Scales. San Antonio: Harcourt Assessment. [Google Scholar]
- Ruge, Hannes, and Uta Wolfensteller. 2016. Distinct contributions of lateral orbito-frontal cortex, striatum, and fronto-parietal network regions for rule encoding and control of memory-based implementation during instructed reversal learning. NeuroImage 125: 1–12. [Google Scholar] [CrossRef] [PubMed]
- Salthouse, Timothy A. 1996. The processing-speed theory of adult age differences in cognition. Psychological Review 103: 403–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salthouse, Timothy A., Jeffrey E. Pink, and Elliot M. Tucker-Drob. 2008. Contextual analysis of fluid intelligence. Intelligence 36: 464–86. [Google Scholar] [CrossRef] [Green Version]
- Shallice, Timothy, and Paul Burgess. 1996. The domain of supervisory processes and temporal organization of behaviour. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 351: 1405–12. [Google Scholar]
- Shilling, V. M., A. Chetwynd, and P. M. A. Rabbitt. 2002. Individual inconsistency across measures of inhibition: An investigation of the construct validity of inhibition in older adults. Neuropsychologia 40: 605–19. [Google Scholar] [CrossRef]
- Singer, Wolf. 1999. Neuronal synchrony: A versatile code for the definition of relations? Neuron 24: 49–65. [Google Scholar] [CrossRef] [Green Version]
- Singh, Leher, Charlene S. L. Fu, Aishah A. Rahman, Waseem B. Hameed, Shamini Sanmugam, Pratibha Agarwal, Binyan Jiang, Yap Seng Chong, Michael J. Meaney, Anne Rifkin-Graboi, and et al. 2015. Back to basics: A bilingual advantage in infant visual habituation. Child Development 86: 294–302. [Google Scholar] [CrossRef]
- Sirin, Selcuk R. 2005. Socioeconomic status and academic achievement: A meta-analytic review of research. Review of Educational Research 75: 417–53. [Google Scholar] [CrossRef] [Green Version]
- Sliwinska, Magdalena W., Inês R. Violante, Richard J. S. Wise, Robert Leech, Joseph T. Devlin, Fatemeh Geranmayeh, and Adam Hampshire. 2017. Stimulating multiple-demand cortex enhances vocabulary learning. Journal of Neuroscience 37: 7606–18. [Google Scholar] [CrossRef]
- Sliwinska, Magdalena W., Ryan Elson, and David Pitcher. 2021. Stimulating parietal regions of the multiple-demand cortex impairs novel vocabulary learning. Neuropsychologia 162: 108047. [Google Scholar] [CrossRef] [PubMed]
- Stroop, J. Ridley. 1935. Studies of interference in serial verbal reactions. Journal of Experimental Psychology 18: 643. [Google Scholar] [CrossRef]
- Tamnes, Christian K., Ylva Østby, Kristine B. Walhovd, Lars T. Westlye, Paulina Due-Tønnessen, and Anders M. Fjell. 2010. Intellectual abilities and white matter microstructure in development: A diffusion tensor imaging study. Human Brain Mapping 31: 1609–25. [Google Scholar] [CrossRef] [PubMed]
- Tao, Lily, Anna Marzecová, Marcus Taft, Dariusz Asanowicz, and Zofia Wodniecka. 2011. The efficiency of attentional networks in early and late bilinguals: The role of age of acquisition. Frontiers in Psychology 2: 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traub, Roger D., Andrea Bibbig, Fiona EN LeBeau, Eberhard H. Buhl, and Miles A. Whittington. 2004. Cellular mechanisms of neuronal population oscillations in the hippocampus in vitro. Annual Review of Neuroscience 27: 247–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, Lucina Q. 2015. Salience processing and insular cortical function and dysfunction. Nature Reviews Neuroscience 16: 55–61. [Google Scholar] [CrossRef]
- Uhlhaas, Peter J., and Wolf Singer. 2006. Neural synchrony in brain disorders: Relevance for cognitive dysfunctions and pathophysiology. Neuron 52: 155–68. [Google Scholar] [CrossRef] [Green Version]
- Ullén, Fredrik, Lea Forsman, Örjan Blom, Anke Karabanov, and Guy Madison. 2008. Intelligence and variability in a simple timing task share neural substrates in the prefrontal white matter. Journal of Neuroscience 28: 4238–43. [Google Scholar] [CrossRef] [Green Version]
- van Doorn, Johnny, Don van den Bergh, Udo Böhm, Fabian Dablander, Koen Derks, Tim Draws, Alexander Etz, Nathan J. Evans, Quentin F. Gronau, Julia M. Haaf, and et al. 2021. The JASP guidelines for conducting and reporting a Bayesian analysis. Psychonomic Bulletin & Review 28: 813–26. [Google Scholar]
- Von Stumm, Sophie, and Robert Plomin. 2015. Socioeconomic status and the growth of intelligence from infancy through adolescence. Intelligence 48: 30–36. [Google Scholar] [CrossRef] [Green Version]
- Walsh, Vincent, Elisabeth Ashbridge, and Alan Cowey. 1998. Cortical plasticity in perceptual learning demonstrated by transcranial magnetic stimulation. Neuropsychologia 36: 45–49. [Google Scholar] [CrossRef]
- Wendelken, Carter, Emilio Ferrer, Simona Ghetti, Stephen K. Bailey, Laurie Cutting, and Silvia A. Bunge. 2017. Frontoparietal structural connectivity in childhood predicts development of functional connectivity and reasoning ability: A large-scale longitudinal investigation. Journal of Neuroscience 37: 8549–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, P. O. 1982. Some major components in general intelligence. In A Model for Intelligence. Edited by Hans J. Eysenck. Berlin/Heidelberg: Springer, pp. 44–90. [Google Scholar]
- Yow, W. Quin, and Xiaoqian Li. 2015. Balanced bilingualism and early age of second language acquisition as the underlying mechanisms of a bilingual executive control advantage: Why variations in bilingual experiences matter. Frontiers in Psychology 6: 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Age (SD) | % Females | Education (SD) 1 |
|---|---|---|---|---|
| Early bilingual | 49 | 26.71 (4.34) | 49% | 3.83 (1.35) |
| Late bilingual | 64 | 27.97 (8.76) | 55% | 3.27 (1.30) |
| Monolingual | 57 | 31.86 (13.90) | 42% | 3.25 (1.42) |
| All Bilinguals | Early Bilinguals | Late Bilinguals | Sig. | |
|---|---|---|---|---|
| Languages | ||||
| All languages | 3.79 (1.33) | 4.06 (1.21) | 3.58 (1.39) | |
| Conversational 1 | 3.12 (1.05) | 3.41 (1.10) | 2.91 (0.97) | .011 * |
| Advanced 2 | 2.52 (0.96) | 2.94 (0.92) | 2.20 (0.88) | <.001 *** |
| Proficiency 3 | ||||
| L1 proficiency | 4.00 (1.00) | 4.00 (0.00) | 4.00 (1.00) | .382 |
| L2 proficiency | 4.00 (2.00) | 4.00 (2.00) | 3.00 (2.00) | <.001 *** |
| L2 minus L1 | 0.00 (2.00) | 0.00 (2.00) | −1.00 (2.00) | .002 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Souza, D.; Dakhch, Y. Is Early Bilingual Experience Associated with Greater Fluid Intelligence in Adults? Languages 2022, 7, 100. https://doi.org/10.3390/languages7020100
D’Souza D, Dakhch Y. Is Early Bilingual Experience Associated with Greater Fluid Intelligence in Adults? Languages. 2022; 7(2):100. https://doi.org/10.3390/languages7020100
Chicago/Turabian StyleD’Souza, Dean, and Yousra Dakhch. 2022. "Is Early Bilingual Experience Associated with Greater Fluid Intelligence in Adults?" Languages 7, no. 2: 100. https://doi.org/10.3390/languages7020100
APA StyleD’Souza, D., & Dakhch, Y. (2022). Is Early Bilingual Experience Associated with Greater Fluid Intelligence in Adults? Languages, 7(2), 100. https://doi.org/10.3390/languages7020100