
Vocal Learning and Behaviors in Birds and Human Bilinguals: Parallels, Divergences and Directions for Research


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Preliminaries
2.1. The Scope of Inquiry and the Boundaries of Our Discussion
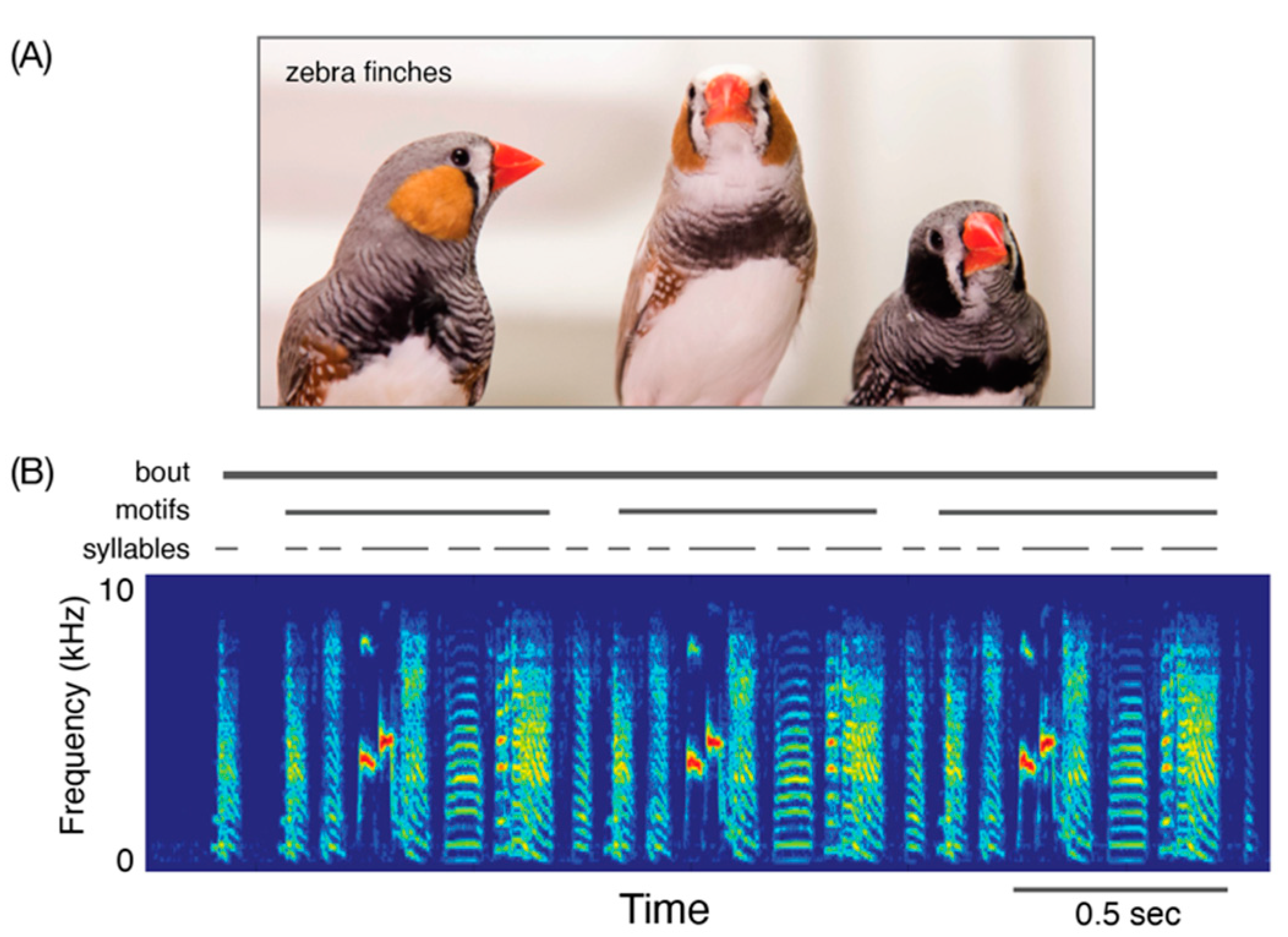
2.2. Songbirds and Birdsongs
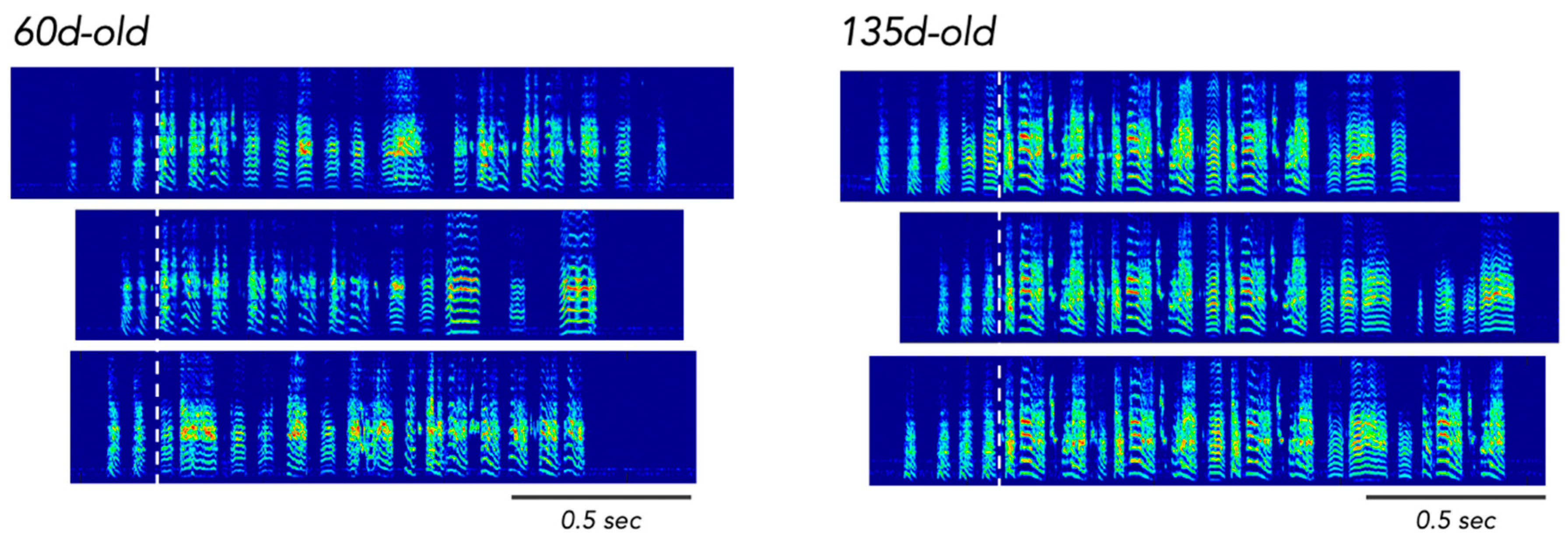
2.3. How Are Songs Learned? The Basics
2.4. Closed-Ended Learning vs. Open-Ended Learning
2.5. Sensory Learning and Sensorimotor Learning
2.6. Concluding Remarks
3. Comparing Late Birdsong Learning and L2 Speech Learning
3.1. Repertoire Size, Timing of Learning, and Variability
3.2. Neural Representation of Speech and Song
3.3. Mechanisms Underlying Age-Dependent Changes in Vocal Learning Abilities
3.4. Concluding Remarks
4. Factors in Variable Outcomes
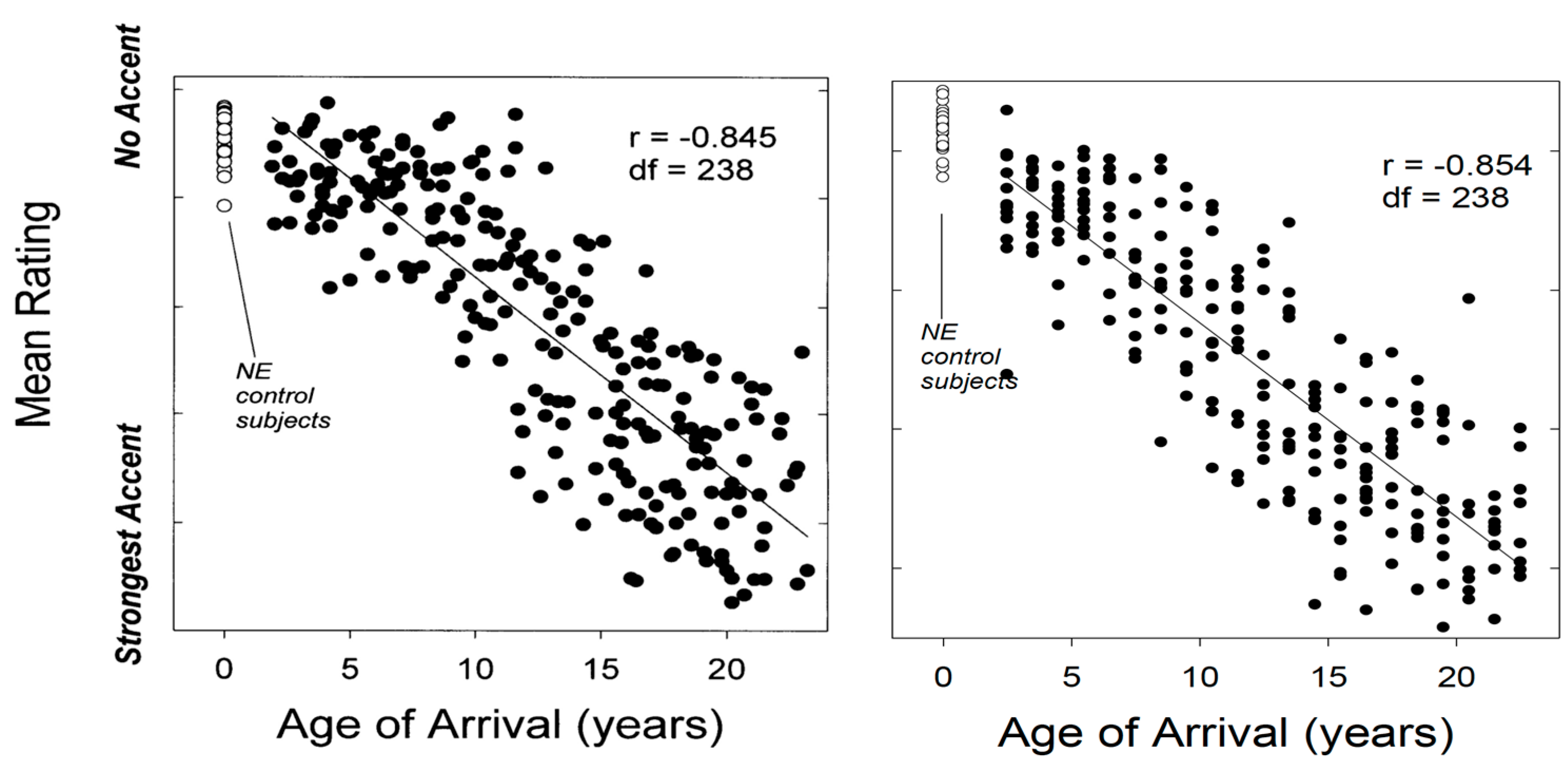
4.1. Age of Learning, Accentedness and Variability
4.2. The Role of Entrenchment
4.3. The Importance of Social Interactions for Vocal Learning
4.4. Concluding Remarks
5. In What Ways Are “Bilingual Birds” Comparable to Bilingual Humans?
5.1. Among Bird Species That Learn an S2, Is There Evidence of Interference from S1 in S2 Production?
5.2. Among Bird Species That Learn an S2, Is There Evidence for S1 Attrition? And If So, Is S1 Attrition Age-Dependent? Relatedly, among Bird Species That Learn an S2, Is There Evidence of Effects of S2 on S1 Production?
5.3. Is There Evidence of “Code-Switching” or “Code-Mixing” in Songbirds?
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
| 1 | Parrots and hummingbirds are other species that learn their vocalizations but will not be the focus here. As an additional clarification, to be amplified later, we note that songbirds, in the same manner as other birds, use their vocalizations during social interactions (e.g., males courting females or defending their territory from other males: Catchpole and Slater 2008; Nowicki and Searcy 2014; Podos and Sung 2020). |
| 2 | However, a recent paper documents evidence of human speech imitation by musk ducks (ten Cate and Fullagar 2021). |
| 3 | In the interest of clarity, we highlight a distinction between songs and calls. Whereas songs consist of multiple, learned vocal elements that are strung together in rapid succession, calls are most often produced as single vocalizations (i.e., calls are generally not concatenated together to form stereotyped sequences of sounds) and often do not require learning (but, for discussions of calls that do require learning: e.g., Elie and Theunissen 2020). In addition, songs and calls are used in different social contexts: songs are used primarily during mating or aggressive interactions, whereas diverse types of calls are used in different contexts including feeding, defense, and bonding contexts (Elie and Theunissen 2020; Marler 2004). |
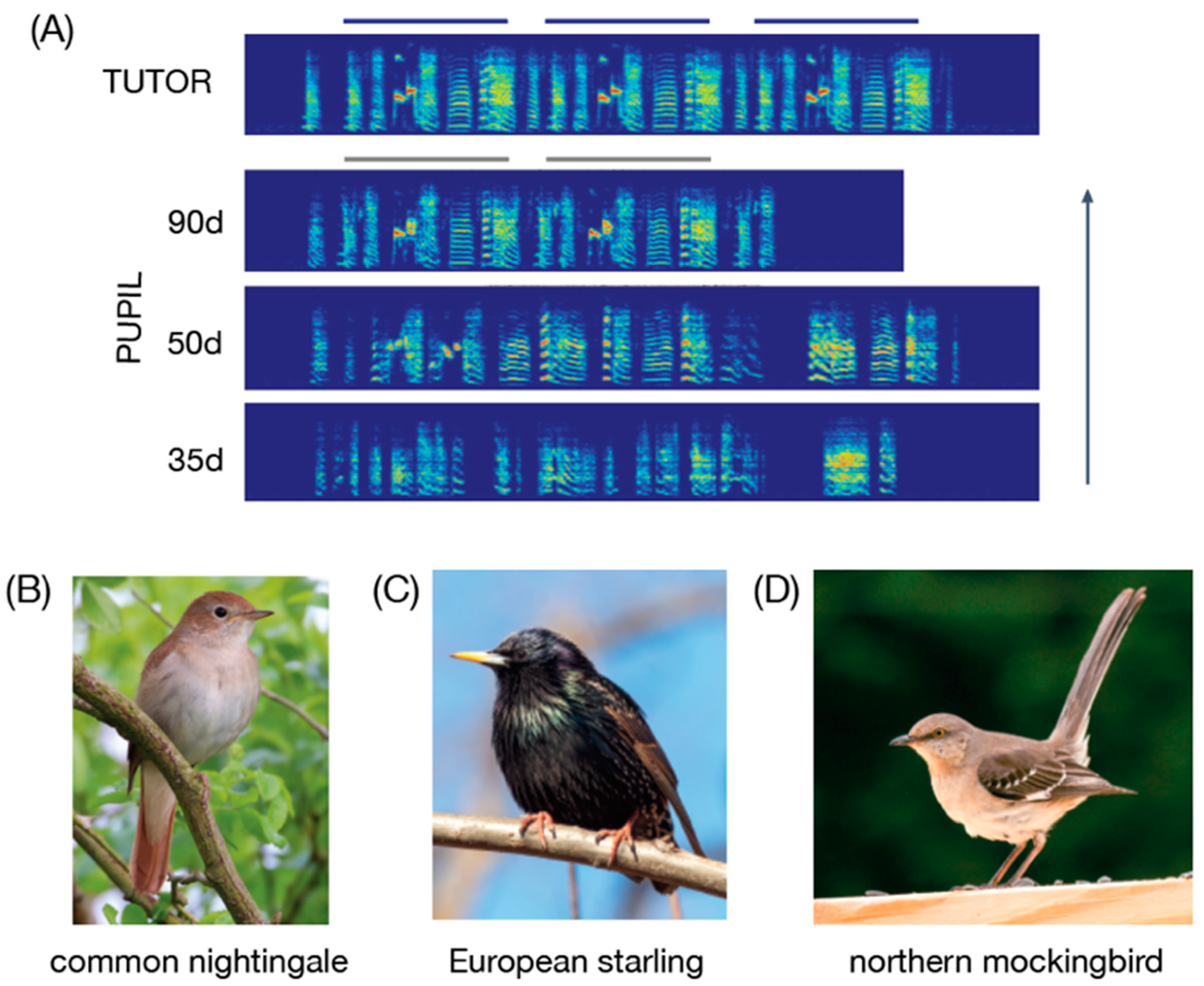
| 4 | Although the terms “tutor” and “pupil” could suggest some degree of active teaching, there are insufficient data to indicate that directed instruction takes place in the context of vocal learning by songbirds; see Caro and Hauser (1992) for their description of teaching. |
| 5 | Although we, similarly to other researchers, define mimicry as the “imitation of all types of non-conspecific sounds: other species, anthropogenic (e.g., dog whistle, chainsaw), and environmental noises (e.g., water drip, leaves rustling)” (Goller and Shizuka 2018), others define mimicry based on the functional consequences of the vocalizations (e.g., whether individuals of the different species respond in the “appropriate” way to the mimic’s imitation of that species vocalization; Dalziell et al. 2015). We align our discussions of imitation fidelity to the degree to which the speech sounds of an L2 speaker acoustically resemble those same sounds used by L1 speakers of that language. That being said, a number of L2 researchers have emphasized the extent to which L2 speech serves the social function of acceptance (e.g., “passing for” a native speaker, Piller 2002), a reminder that function, purpose and intent are not irrelevant in this context. |
| 6 | A video of birds’ imitations of human speech: https://www.youtube.com/watch?v=N5YbWHrnjrg (accessed on 13 December 2021). |
| 7 | The existence and nature of periods for optimal speech acquisition continue to be debated, but generally speaking, it is agreed that humans are able to learn new languages and sounds throughout their lives, that attainment of new language and pronunciation features declines but does not cease over age of learning, and that factors unrelated to neurological maturation, detailed in later sections of this review, can condition the outcomes of L2 speech learning (e.g., Birdsong 2017, 2018; Flege et al. 1999; Flege and Bohn 2021; Werker and Hensch 2015). |
| 8 | An example of how will and identity determine specific features of L2 pronunciation is noted by Walters (2011). By choice, Tunisian women speakers of L2 French pronounce French /r/ as the uvular [ʁ], just as native French speakers do. Tunisian men, on the other hand, choose to pronounce their L2 French /r/ as apical [r]. In this way they deliberately distinguish themselves from Tunisian women and from native speakers of standard French. Importantly, in Tunisian Arabic (the L1 of Tunisian men and women), both /r/ and /ʁ/ are produced, and in fact both are phonemic. Thus, articulating the French-like [ʁ] among Tunisian speakers of L2 French is not a question of having to master a new speech sound, but a matter of assertion of identity. |
| 9 | Estimates of individual or species variation in repertoire size can dramatically vary depending on the unit of measurement: e.g., repertoire size as the number of song types, phrase types (strophes or motifs), syllable types, or note types (Creanza et al. 2016). (As an analogy, one might imagine quantifying the number of words vs. syllables in lexicons). Rather than limit ourselves to a single definition of repertoire size, in relevant contexts we will consider a range of “units” of birdsong and will explicitly indicate the unit of analysis. |
| 10 | This could be likened to discovering that a friend of yours is able to speak a different language, one that they learned while growing up. |
| 11 | Given the emphasis on experimental design for providing compelling evidence of adult vocal learning, it should be mentioned that previous studies would be strengthened with the inclusion of a control group in which birds are not exposed to tutor songs in adulthood. For example, the European starlings that demonstrated vocal changes in response to late tutoring (Chaiken et al. 1994) were exposed to tutor songs during both development (“early tutors”) and in adulthood (“late tutors”), but it is possible that starlings that were exposed to tutor songs only during development could have “improvised” the early tutor songs in a way that caused their songs to coincidentally match late tutor songs. The degree to which this alternative explanation accounts for published observations remains unclear (because the full range of songs used for tutoring is generally not published along with these papers), but it stands as an important control to consider for future experiments. |
| 12 | There are a number of papers describing adult vocal learning in birds raised without exposure to adult song during development (e.g., in zebra finches, canaries, and brown-headed cowbirds: Gobes et al. 2019; Lehongre et al. 2009; Leitner and Catchpole 2007; O’Loghlen and Rothstein 2010; Sakata and Yazaki-Sugiyama 2020). Although orthogonal to our highlighting of parallels between S2 learning and L2 speech acquisition (i.e., acquiring new vocalizations after learning a previous set of vocalizations), these are compelling and useful examples because adult learning in these cases cannot be attributed to developmental song learning. We refer readers interested in this topic to various reviews and primary research articles cited in this paragraph. |
| 13 | A number of studies have found that individual neurons in HVC are “tuned” to multiple song types. For example, adult male swamp sparrows produce 2–5 different song types that consist of distinct sounds. Although various neurons in the HVC of male song sparrows are activated when the bird hears only one of the bird’s song types (i.e., neurons are “tuned” to one song type), many HVC neurons are activated in response to hearing multiple song types that are produced by the bird (Mooney et al. 2001). Similar findings have been observed in white-crowned sparrows (Prather et al. 2010). These findings suggest that there is not a simple one-to-one correspondence between the size of a brain area (or number of neurons in a brain area) and the repertoire of sounds that an individual songbird uses for communication. |
| 14 | Studies of AoA-related morphosyntactic attainment, which are more numerous than those for pronunciation, reveal a similar pattern of decline with greater dispersion at later AoA (e.g., DeKeyser et al. 2010; Chen and Hartshorne 2021; Flege et al. 1999; Hartshorne et al. 2018). |
| 15 | In the area of morphosyntax, L2 effects on the L1 are seen in online processing and in judgments of grammaticality (Kasparian and Steinhauer 2017), and have been attributed by different researchers to changes in representational knowledge and to non-pathological neurocognitive changes traceable to dominance factors (e.g., Steinhauer and Kasparian 2020 and references therein). |
References
- Abutalebi, Jubin. 2008. Neural aspects of second language representation and language control. Acta Psychologica 128: 466–78. [Google Scholar] [CrossRef] [PubMed]
- Airey, David C., and Timothy J. DeVoogd. 2000. Greater song complexity is associated with augmented song system anatomy in zebra finches. Neuroreport 11: 1749–54. [Google Scholar] [CrossRef]
- Airey, David C., Donald E. Kroodsma, and Timothy J. DeVoogd. 2000. Differences in the complexity of song tutoring cause differences in the amount learned and in dendritic spine density in a songbird telencephalic song control nucleus. Neurobiology of Learning and Memory 73: 274–81. [Google Scholar] [CrossRef][Green Version]
- Amin, Noopur, Michael Gastpar, and Frédéric E. Theunissen. 2013. Selective and efficient neural coding of communication signals depends on early acoustic and social environment. PLoS ONE 8: e61417. [Google Scholar] [CrossRef]
- Araki, Makoto, Manesh M. Bandi, and Yoko Yazaki-Sugiyama. 2016. Mind the gap: Neural coding of species identity in birdsong prosody. Science 354: 1282–87. [Google Scholar] [CrossRef] [PubMed]
- Au, Terry Kit-Fong, Leah M. Knightly, Sun-Ah Jun, and Janet S. Oh. 2002. Overhearing a language during childhood. Psychological Science 13: 238–43. [Google Scholar] [CrossRef]
- Ball, Gregory F., and Jacques Balthazart. 2020. Sex differences and similarities in the neural circuit regulating song and other reproductive behaviors in songbirds. Neuroscience & Biobehavioral Reviews 118: 258–69. [Google Scholar] [CrossRef]
- Balmer, Timothy S., Vanessa M. Carels, Jillian L. Frisch, and Teresa A. Nick. 2009. Modulation of perineuronal nets and parvalbumin with developmental song learning. Journal of Neuroscience 29: 12878–85. [Google Scholar] [CrossRef]
- Baptista, Luis F., and Lewis Petrinovich. 1986. Song development in the white-crowned sparrow: Social factors and sex differences. Animal Behaviour 34: 1359–71. [Google Scholar] [CrossRef]
- Barrington, Daines. 1773. Experiments and observations on the singing of birds. Philosophical Transactions of the Royal Society of London 63: 249–91. [Google Scholar]
- Beckers, Gabriël J. L., Brian S. Nelson, and Roderick A. Suthers. 2004. Vocal-tract filtering by lingual articulation in a parrot. Current Biology 14: 1592–97. [Google Scholar] [CrossRef]
- Beecher, Michael D. 2017. Birdsong learning as a social process. Animal Behaviour 124: 233–46. [Google Scholar] [CrossRef]
- Beecher, Michael D., and Eliot A. Brenowitz. 2005. Functional aspects of song learning in songbirds. Trends in Ecology & Evolution 20: 143–49. [Google Scholar] [CrossRef]
- Bergmann, Christopher, Amber Nota, Simone A. Sprenger, and Monika S. Schmid. 2016. L2 immersion causes non-native-like L1 pronunciation in German attriters. Journal of Phonetics 58: 71–86. [Google Scholar] [CrossRef]
- Beurdeley, Marine, Julien Spatazza, Henry H. C. Lee, Sayaka Suglyama, Clémence Bernard, Ariel A. Di Nardo, Takao K. Hensch, and Alain Prochiantz. 2012. Otx2 binding to perineuronal nets persistently regulates plasticity in the mature visual cortex. Journal of Neuroscience 32: 9429–37. [Google Scholar] [CrossRef]
- Birdsong, David. 1999. Introduction: Whys and why nots of the Critical Period Hypothesis for second language acquisition. In Second Language Acquisition and the Critical Period Hypothesis. Edited by David Birdsong. Mahwah: Lawrence Erlbaum, pp. 1–22. [Google Scholar]
- Birdsong, David. 2007. Nativelike pronunciation among late learners of French as a second language. In Language Experience in Second Language Speech Learning, in Honor of James Emil Flege. Edited by Ocke-Schwen Bohn and Murray J. Munro. Amsterdam: John Benjamins, pp. 99–116. [Google Scholar]
- Birdsong, David. 2009. Age and the end state of second language acquisition. In The New Handboook of Second Language Acquisition, 2nd ed. Edited by William C. Ritchie and Tej K. Bhatia. Bingley: Emerald, pp. 401–24. [Google Scholar]
- Birdsong, David. 2014. Dominance and age in bilingualism. Applied Linguistics 35: 374–92. [Google Scholar] [CrossRef]
- Birdsong, David. 2017. Critical periods. In Oxford Bibliographies in Linguistics. Edited by Mark Aronoff. New York: Oxford University Press. [Google Scholar]
- Birdsong, David. 2018. Plasticity, variability and age in second language acquisition and bilingualism. Frontiers in Psychology 9: 81. [Google Scholar] [CrossRef]
- Bolhuis, Johan J., Kazuo Okanoya, and Constance Scharff. 2010. Twitter evolution: Converging mechanisms in birdsong and human speech. Nature Reviews Neuroscience 11: 747–59. [Google Scholar] [CrossRef] [PubMed]
- Bongaerts, Theo. 1999. Ultimate attainment in L2 pronunciation: The case of very advanced late L2 learners. In Second Language Acquisition and the Critical Period Hypothesis. Edited by David Birdsong. Mahwah: Lawrence Erlbaum, pp. 133–59. [Google Scholar]
- Bradbury, Jack W., and Sandra L. Vehrencamp. 2011. Principles of Animal Communication, 2nd ed. Massachusetts: Sinauer Associates. [Google Scholar]
- Brainard, Michael S., and Allison J. Doupe. 2000. Interruption of a basal ganglia-forebrain circuit prevents plasticity of learned vocalizations. Nature 404: 762–66. [Google Scholar] [CrossRef] [PubMed]
- Brainard, Michael S., and Allison J. Doupe. 2002. What songbirds teach us about learning. Nature 417: 351–58. [Google Scholar] [CrossRef]
- Brainard, Michael S., and Allison J. Doupe. 2013. Translating birdsong: Songbirds as a model for basic and applied medical research. Annual Review of Neuroscience 36: 489–517. [Google Scholar] [CrossRef]
- Brenowitz, Eliot A., and Michael D. Beecher. 2005. Song learning in birds: Diversity and plasticity, opportunities and challenges. Trends in Neurosciences 28: 127–32. [Google Scholar] [CrossRef]
- Bullock, Barbara E., and Almeida Jacqueline Toribio, eds. 2009. The Cambridge Handbook of Linguistic Code-Switching. Cambridge: Cambridge University Press. [Google Scholar] [CrossRef]
- Byers, Bruce E., and Donald E. Kroodsma. 2009. Female mate choice and songbird song repertoires. Animal Behaviour 77: 13–22. [Google Scholar] [CrossRef]
- Carey, Daniel, Marc E. Miquel, Bronwen G. Evans, Patti Adank, and Carolyn McGettigan. 2017. Functional brain outcomes of L2 speech learning emerge during sensorimotor transformation. NeuroImage 159: 18–31. [Google Scholar] [CrossRef] [PubMed]
- Caro, Tim M., and Marc D. Hauser. 1992. Is there teaching in nonhuman animals? Quarterly Review of Biology 67: 151–74. [Google Scholar] [CrossRef]
- Carouso-Peck, Samantha, Otilia Menyhart, Timothy J. DeVoogd, and Michael H. Goldstein. 2020. Contingent parental responses are naturally associated with zebra finch song learning. Animal Behaviour 165: 123–32. [Google Scholar] [CrossRef]
- Catchpole, Clive K., and Peter J. B. Slater. 2008. Bird Song: Biological Themes and Variations, 2nd ed. Cambridge: Cambridge University Press. [Google Scholar]
- Chaiken, Marthaleah, Jörg Böhner, and Peter Marler. 1994. Repertoire turnover and the timing of song acquisition in European starlings. Behaviour 128: 25–39. [Google Scholar] [CrossRef]
- Chen, Tony, and Joshua K. Hartshorne. 2021. More evidence from over 1.1 million subjects that the critical period for syntax closes in late adolescence. Cognition 214: 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, Yining, and Jon T. Sakata. 2021. Norephinephrine in the avian auditory cortex enhances developmental song learning. Journal of Neurophysiology 125: 2397–407. [Google Scholar] [CrossRef]
- Chen, Yining, Laura E. Matheson, and Jon T. Sakata. 2016. Mechanisms underlying the social enhancement of vocal learning. Proceedings of the National Academy of Sciences of the United States of America 113: 6641–46. [Google Scholar] [CrossRef]
- Choe, Ha Na, and Erich D. Jarvis. 2021. The role of sex chromosomes and sex hormones in vocal learning systems. Hormones and Behavior 132: 104978. [Google Scholar] [CrossRef] [PubMed]
- Choi, Jiyoun, Mirjam Broersma, and Anne Cutler. 2017. Early phonology revealed by international adoptees’ birth language retention. Proceedings of the National Academy of Sciences of the United States of America 114: 7307–12. [Google Scholar] [CrossRef] [PubMed]
- Clayton, Nicky S. 1989. The effects of cross-fostering on selective song learning in estrildid finches. Behaviour 109: 163–75. [Google Scholar] [CrossRef]
- Cornez, Gilles, Farrah N. Madison, Annemie Van der Linden, Charlotte Cornil, Kathleen M. Yoder, Gregory F. Ball, and Jacques Balthazart. 2017. Perineuronal nets and vocal plasticity in songbirds: A proposed mechanism to explain the difference between closed-ended and open-ended learning. Developmental Neurobiology 77: 975–94. [Google Scholar] [CrossRef]
- Cornez, Gilles, Elisabeth Jonckers, Sita M. Ter Haar, Annemie Van der Linden, Charlotte A. Cornil, and Jacques Balthazart. 2018. Timing of perineuronal net development in the zebra finch song control system correlates with developmental song learning. Proceedings of the Royal Society B: Biological Sciences 285: 20180849. [Google Scholar] [CrossRef] [PubMed]
- Coumel, Marion, Markus Christiner, and Susanne Maria Reiterer. 2019. Second language accent faking ability depends on musical abilities, not on working memory. Frontiers in Psychology 10: 257. [Google Scholar] [CrossRef] [PubMed]
- Creanza, Nicole, Laurel Fogarty, and Marcus W. Feldman. 2016. Cultural niche construction of repertoire size and learning strategies in songbirds. Evolutionary Ecology 30: 285–305. [Google Scholar] [CrossRef]
- Dai, Jennifer B., Yining Chen, and Jon T. Sakata. 2018. EGR-1 expression in catecholamine-synthesizing neurons reflects auditory learning and correlates with responses in auditory processing areas. Neuroscience 379: 415–27. [Google Scholar] [CrossRef] [PubMed]
- Dalziell, Anastasia H., Justin A. Welbergen, Branislav Igic, and Robert D. Magrath. 2015. Avian vocal mimicry: A unified conceptual framework. Biological Reviews of the Cambridge Philosophical Society 90: 643–68. [Google Scholar] [CrossRef] [PubMed]
- de Bruin, Angela, Arthur G. Samuel, and Jon Anoni Duñabeitia. 2018. Voluntary language switching: When and why do bilinguals switch between their languages? Journal of Memory and Language 103: 28–43. [Google Scholar] [CrossRef]
- de Leeuw, Esther, Monika S. Schmid, and Ineke Mennen. 2010. The effects of contact on native language pronunciation in an L2 migrant setting. Bilingualism: Language and Cognition 13: 33–40. [Google Scholar] [CrossRef]
- Dehaene, Stanislas, Emmanuel Dupoux, Jacques Mehler, Laurent Cohen, Eraldo Paulesu, Daniela Perani, Pierre-François van de Moortele, Stéphane Lehéricy, and Denis Le Bihan. 1997. Anatomical variability in the cortical representation of first and second language. Neuroreport 8: 3809–15. [Google Scholar] [CrossRef] [PubMed]
- Dehaene-Lambertz, Ghislaine, Stanislas Dehaene, and Lucie Hertz-Pannier. 2002. Functional neuroimaging of speech perception in infants. Science 298: 2013–15. [Google Scholar] [CrossRef]
- DeKeyser, Robert, Iris Alfi-Shabtay, and Dorit Ravid. 2010. Cross-linguistic evidence for the nature of age effects in second language acquisition. Applied Psycholinguistics 31: 413–38. [Google Scholar] [CrossRef]
- Derégnaucourt, Sébastien, Colline Poirier, Anne Van der Kant, Annemie Van der Linden, and Manfred Gahr. 2013. Comparisons of different methods to train a young zebra finch (Taeniopygia guttata) to learn a song. Journal of Physiology-Paris 107: 210–18. [Google Scholar] [CrossRef] [PubMed]
- Derrickson, Kim C. 1987. Yearly and situational changes in the estimate of repertoire size in Northern Mockingbirds (Mimus polyglottos). The Auk 104: 198–207. [Google Scholar] [CrossRef]
- DeVoogd, Timothy J. 2004. Neural constraints on the complexity of avian song. Brain, Behavior and Evolution 63: 221–32. [Google Scholar] [CrossRef] [PubMed]
- Dhawale, Ashesh K., Maurice A. Smith, and Bence P. Ölveczky. 2017. The role of variability in motor learning. Annual Review of Neuroscience 40: 479–98. [Google Scholar] [CrossRef]
- Díaz, Begoña, Cristina Baus, Carles Escera, Albert Costa, and Núria Sebastián-Gallés. 2008. Brain potentials to native phoneme discrimination reveal the origin of individual differences in learning the sounds of a second language. Proceedings of the National Academy of Sciences of the United States of America 105: 16083–88. [Google Scholar] [CrossRef]
- Díaz, Begoña, Holger Mitterer, Mirjam Broersma, Carles Escera, and Núria Sebastián-Gallés. 2016. Variability in L2 phonemic learning originates from speech-specific capabilities: An MMN study on late bilinguals. Bilingualism: Language and Cognition 19: 955–70. [Google Scholar] [CrossRef]
- Doupe, Allison J. 1997. Song-and order-selective neurons in the songbird anterior forebrain and their emergence during vocal development. Journal of Neuroscience 17: 1147–67. [Google Scholar] [CrossRef] [PubMed]
- Doupe, Allison J., and Patricia K. Kuhl. 1999. Birdsong and human speech: Common themes and mechanisms. Annual Review of Neuroscience 22: 567–631. [Google Scholar] [CrossRef] [PubMed]
- Doya, Kenji, and Terrence J. Sejnowski. 1995. A novel reinforcement model of birdsong vocalization learning. In Advances in Neural Information Processing Systems. Edited by Gerald Tesauro, David S. Touretzky and Todd K. Leen. Cambridge: MIT Press, pp. 101–8. [Google Scholar]
- Eales, Lucy A. 1985. Song learning in zebra finches: Some effects of song model availability on what is learnt and when. Animal Behaviour 33: 1293–300. [Google Scholar] [CrossRef]
- Elemans, Coen P. H., Jeppe Have Rasmussen, Christian T. Herbst, Daniel Normen Düring, Sue Anne Zollinger, Henrik Brumm, Krishna C. Srivastava, Niels Svane, Ming Ding, Ole Naesbye Larsen, and et al. 2015. Universal mechanisms of sound production and control in birds and mammals. Nature Communications 6: 8978. [Google Scholar] [CrossRef] [PubMed]
- Elie, Julie E., and Frédéric Theunissen. 2020. The neuroethology of vocal communication in songbirds: Production and perception of a call repertoire. In The Neuroethology of Birdsong. Edited by Jon T. Sakata, Sarah C. Woolley, Richard R. Fay and Arthur N. Popper. Cham: Springer Nature, pp. 175–209. [Google Scholar] [CrossRef]
- Eriksen, Ane, Tore Slagsvold, and Helene Marie Lampe. 2011. Vocal plasticity–are pied flycatchers, Ficedula hypoleuca, open-ended learners? Ethology 117: 188–98. [Google Scholar] [CrossRef]
- Fee, Michale S., and Jesse H. Goldberg. 2011. A hypothesis for basal ganglia-dependent reinforcement learning in the songbird. Neuroscience 198: 152–70. [Google Scholar] [CrossRef]
- Fehér, Olga, Haibin Wang, Sigal Saar, Partha P. Mitra, and Ofer Tchernichovski. 2009. De novo establishment of wild-type song culture in the zebra finch. Nature 459: 564–68. [Google Scholar] [CrossRef] [PubMed]
- Feng, Gangyi, Yu Li, Shen-Mou Hsu, Patrick C. M. Wong, Tai-Li Chou, and Bharath Chandrasekaran. 2021. Emerging native-similar neural representations underlie non-native speech category learning success. Neurobiology of Language 2: 280–307. [Google Scholar] [CrossRef] [PubMed]
- Fitch, W. Tecumseh, Ludwig Huber, and Thomas Bugnyar. 2010. Social cognition and the evolution of language: Constructing cognitive phylogenies. Neuron 65: 795–814. [Google Scholar] [CrossRef]
- Flege, James Emil. 1998. The role of subject and phonetic variables in second-language learning. In CLS 34: The Panels. Edited by M. Catherine Gruber, Derrick Higgins, Kenneth S. Olson and Tamra Wysocki. Chicago: Chicago Linguistic Society, pp. 213–32. [Google Scholar]
- Flege, James E. 1999. Age of learning and second language speech. In Second Language Acquisition and the Critical Period Hypothesis. Edited by David Birdsong. Mahwah: Lawrence Erlbaum, pp. 101–31. [Google Scholar]
- Flege, James Emil. 2018. A non-critical period for second language. In A Sound Approach to Language Matters: In Honor of Ocke-Schwen Bohn. Edited by Anne Mette Nyvad, Michaela Hejná, Anders Højen, Anna Bothe Jespersen and Mette Hjortshøj Sørensen. Aarhus: Aarhus University Library Scholarly Publishing Services, pp. 501–41. [Google Scholar]
- Flege, James Emil, and Ocke-Schwen Bohn. 2021. The Revised Speech Learning Model (SLM-r). In Second Language Speech Learning: Theoretical and Empirical Progress. Edited by Ratree Wayland. Cambridge: Cambridge University Press, pp. 3–83. [Google Scholar] [CrossRef]
- Flege, James Emil, Murray J. Munro, and Ian R. A. MacKay. 1995. Factors affecting strength of perceived foreign accent in a second language. Journal of the Acoustic Society of America 97: 3125–34. [Google Scholar] [CrossRef] [PubMed]
- Flege, James Emil, Grace H. Yeni-Komshian, and Serena Liu. 1999. Age constraints on second-language acquisition. Journal of Memory and Language 41: 78–104. [Google Scholar] [CrossRef]
- Friederici, Angela. 2011. The brain basis of language processing: From structure to function. Physiological Reviews 91: 1357–92. [Google Scholar] [CrossRef] [PubMed]
- Gammon, David E. 2013. How is model selection determined in a vocal mimic?: Tests of five hypotheses. Behaviour 150: 1375–97. [Google Scholar] [CrossRef]
- Gammon, David E. 2020. Are northern mockingbirds classic open-ended song learners? Ethology 126: 1038–47. [Google Scholar] [CrossRef]
- Garcia-Sierra, Adrian, Maritza Rivera-Gaxiola, Cherie R. Percaccio, Barbara T. Conboy, Harriett Romo, Lindsay Klarman, Sophia Ortiz, and Patricia K. Kuhl. 2011. Bilingual language learning: An ERP study relating early brain responses to speech, language input, and later word recognition. Journal of Phonetics 39: 546–57. [Google Scholar] [CrossRef]
- Gardner, Timothy J., Felix Naef, and Fernando Nottebohm. 2005. Freedom and rules: The acquisition and reprogramming of a bird’s learned song. Science 308: 1046–49. Available online: https://www.science.org/doi/10.1126/science.1108214 (accessed on 13 December 2021). [CrossRef]
- Gaskell, M. Gareth, and Andrew W. Ellis. 2009. Word learning and lexical development across the lifespan. Philosophical Transactions of the Royal Society B: Biological Sciences 364: 3607–15. [Google Scholar] [CrossRef]
- Geberzahn, Nicole, and Henrike Hultsch. 2003. Long–time storage of song types in birds: Evidence from interactive playbacks. Proceedings of the Royal Society of London. Series B: Biological Sciences 270: 1085–90. [Google Scholar] [CrossRef]
- Geberzahn, Nicole, Henrike Hultsch, and Dietmar Todt. 2002. Latent song type memories are accessible through auditory stimulation in a hand-reared songbird. Animal Behaviour 64: 783–90. [Google Scholar] [CrossRef][Green Version]
- Gervain, Judit, and Jacques Mehler. 2010. Speech perception and language acquisition in the first year of life. Annual Review of Psychology 61: 191–218. [Google Scholar] [CrossRef]
- Gobes, Sharon MH, and Johan J. Bolhuis. 2007. Birdsong memory: A neural dissociation between song recognition and production. Current Biology 17: 789–93. [Google Scholar] [CrossRef]
- Gobes, Sharon M. H., Rebecca B. Jennings, and Rie K. Maeda. 2019. The sensitive period for auditory-vocal learning in the zebra finch: Consequences of limited-model availability and multiple-tutor paradigms on song imitation. Behavioural Processes 163: 5–12. [Google Scholar] [CrossRef]
- Goffinet, Jack, Samuel Brudner, Richard Mooney, and John Pearson. 2021. Low-dimensional learned feature spaces quantify individual and group differences in vocal repertoires. eLife 10: e67855. [Google Scholar] [CrossRef]
- Golestani, Narly, and Robert J. Zatorre. 2004. Learning new sounds of speech: Reallocation of neural substrates. NeuroImage 21: 494–506. [Google Scholar] [CrossRef]
- Golestani, Narly, Tomás Paus, and Robert J. Zatorre. 2002. Anatomical correlates of learning novel speech sounds. Neuron 35: 997–1010. [Google Scholar] [CrossRef]
- Golestani, Narly, Nicolas Molko, Stanislas DeHaene, Denis Le Bihan, and Christophe Pallier. 2007. Brain structure predicts the learning of foreign speech sounds. Cerebral Cortex 17: 575–82. [Google Scholar] [CrossRef]
- Goller, Maria, and Daizaburo Shizuka. 2018. Evolutionary origins of vocal mimicry in songbirds. Evolution Letters 2: 417–26. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Marin, Alex, Joseph J. Patton, Adam R. Kampff, Rui M. Costa, and Zachary F. Mainen. 2014. Big behavioral data: Psychology, ethology and the foundations of neuroscience. Nature Neuroscience 17: 1455–62. [Google Scholar] [CrossRef]
- Goodale, Eben, and Sarath W. Kotagama. 2006. Context-dependent vocal mimicry in a passerine bird. Proceedings of the Royal Society B: Biological Sciences 273: 875–80. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, François. 2010. Bilingual: Life and Reality. Cambridge: Harvard University Press. [Google Scholar]
- Guion, Susan G., James E. Flege, and Jonathan D. Loftin. 2000. The effect of L1 use on pronunciation in Quichua-Spanish bilinguals. Journal of Phonetics 28: 27–42. [Google Scholar] [CrossRef]
- Hartshorne, Joshua K., Joshua B. Tenenbaum, and Steven Pinker. 2018. A critical period for second language acquisition: Evidence from 2/3 million English speakers. Cognition 177: 263–77. [Google Scholar] [CrossRef]
- Hausberger, Martine, Peter F. Jenkins, and Jeremy Keene. 1991. Species-specificity and mimicry in bird song: Are they paradoxes? A reevaluation of song mimicry in the European starling. Behaviour 117: 53–81. [Google Scholar] [CrossRef]
- Helekar, Santosh, ed. 2013. Animal Models of Speech and Language Disorders. New York: Springer. [Google Scholar]
- Heredia, Roberto R., and Jeanette Altarriba. 2001. Bilingual language mixing: Why do bilinguals code-switch? Current Directions in Psychological Science 10: 164–68. [Google Scholar] [CrossRef]
- Hernandez, Arturo. 2009. Language switching in the bilingual brain: What’s next? Brain and Language 108: 133–40. [Google Scholar] [CrossRef] [PubMed]
- Hopp, Holger, and Monika S. Schmid. 2013. Perceived foreign accent in first language attrition and second language acquisition: The impact of age of acquisition and bilingualism. Applied Psycholinguistics 34: 361–94. [Google Scholar] [CrossRef]
- Hough, Gerald E. II, Douglas A. Nelson, and Susan F. Volman. 2000. Re-expression of songs deleted during vocal development in white-crowned sparrows, Zonotrichia leucophrys. Animal Behaviour 60: 279–87. [Google Scholar] [CrossRef] [PubMed]
- Houx, Bart B., and Carel ten Cate. 1998. Do contingencies with tutor behaviour influence song learning in zebra finches? Behaviour 135: 599–614. Available online: https://www.jstor.org/stable/4535547 (accessed on 13 December 2021). [CrossRef]
- Hu, Xiaochen, Hermann Ackermann, Jason A. Martin, Michael Erb, Susanne Winkler, and Susanne M. Reiterer. 2012. Language aptitude for pronunciation in advanced second language (L2) learners: Behavioural predictors and neural substrates. Brain and Language 127: 366–76. [Google Scholar] [CrossRef] [PubMed]
- Hyltenstam, Kenneth, and Niclas Abrahamsson. 2003. Maturational constraints in SLA. In The Handbook of Second Language Acquisition. Edited by Catherine J. Doughty and Michael H. Long. Malden: Blackwell, pp. 539–88. [Google Scholar]
- Igic, Branislav, and Robert D. Magrath. 2014. A songbird mimics different heterospecific alarm calls in response to different types of threat. Behavioral Ecology 25: 538–48. [Google Scholar] [CrossRef]
- Ingvalson, Erin M., Casandra Nowicki, Audrey Zong, and Patrick C. M. Wong. 2017. Non-native speech learning in older adults. Frontiers in Psychology 8: 148. [Google Scholar] [CrossRef] [PubMed]
- James, Logan S., and Jon T. Sakata. 2017. Learning biases underlie “universals” in avian vocal sequencing. Current Biology 27: 3676–82. [Google Scholar] [CrossRef]
- James, Logan S., Ronald Davies Jr, Chihiro Mori, Kazuhiro Wada, and Jon T. Sakata. 2020. Manipulations of sensory experiences during development reveal mechanisms underlying vocal learning biases in zebra finches. Developmental Neurobiology 80: 132–46. [Google Scholar] [CrossRef] [PubMed]
- Janik, Vincent M., and Peter J. Slater. 2000. The different roles of social learning in vocal communication. Animal Behaviour 60: 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, Erich D. 2019. Evolution of vocal learning and spoken language. Science 366: 50–54. [Google Scholar] [CrossRef] [PubMed]
- Baylis, Jeffrey R. 1982. Avian vocal mimicry: Its function and evolution. In Acoustic Communication in Birds. Edited by Donald E. Kroodsma, Edward H. Miller and Henri Ouellet. New York: Academic Press, vol. 2, pp. 51–83. [Google Scholar]
- Jenkins, Jane, and Jennifer Setter. 2005. State of the art review article: Pronunciation. Language Teaching 38: 1–17. [Google Scholar] [CrossRef]
- Jones, Alex E., Carel ten Cate, and Peter J. B. Slater. 1996. Early experience and plasticity of song in adult male zebra finches (Taeniopygia guttata). Journal of Comparative Psychology 110: 354–69. [Google Scholar] [CrossRef]
- Karayayla, Tugba, and Monika S. Schmid. 2019. First language attrition as a function of age of onset of bilingualism: First language attainment of Turkish-English bilinguals in the United Kingdom. Language Learning 69: 106–42. [Google Scholar] [CrossRef]
- Kartushina, Natalia, and Ulrich H. Frauenfelder. 2014. On the effects of L2 perception and of individual differences in L1 production on L2 pronunciation. Frontiers in Psychology 5: 1246. [Google Scholar] [CrossRef]
- Kasparian, Kristina, and Karsten Steinhauer. 2017. When the second language takes the lead: Neurocognitive processing changes in the first language of adult attriters. Frontiers in Psychology 8: 389. [Google Scholar] [CrossRef]
- Keller, Georg B., and Richard H. R. Hahnloser. 2009. Neural processing of auditory feedback during vocal practice in a songbird. Nature 457: 187–90. [Google Scholar] [CrossRef] [PubMed]
- Kelley, Laura A., Rebecca L. Coe, Joah R. Madden, and Susan D. Healy. 2008. Vocal mimicry in songbirds. Animal Behaviour 76: 521–28. [Google Scholar] [CrossRef]
- Kipper, Silke, and Sarah Kiefer. 2010. Age-related changes in birds’ singing styles: On fresh tunes and fading voices? Advances in the Study of Behavior 41: 77–118. [Google Scholar] [CrossRef]
- Klein, Denise, Brenda Milner, Robert J. Zatorre, Ernst Meyer, and Alan C. Evans. 1995. The neural substrates underlying word generation: A bilingual functional-imaging study. Proceedings of the National Academy of Sciences of the United States of America 92: 2899–903. [Google Scholar] [CrossRef] [PubMed]
- Köpke, Barbara. 2021. Language attrition: A matter of brain plasticity? Some preliminary thoughts. Language, Interaction and Acquisition 12: 110–32. [Google Scholar] [CrossRef]
- Köpke, Barbara, Monika S. Schmid, Merel Keijzer, and Susan Dostert, eds. 2007. Language Attrition: Theoretical Perspectives. Amsterdam: John Benjamins. [Google Scholar]
- Kornder, Lisa, and Ineke Mennen. 2021. Longitudinal developments in bilingual second language acquisition and first language attrition of speech: The case of Arnold Schwarzenegger. Languages 6: 61. [Google Scholar] [CrossRef]
- Kuhl, Patricia K. 2007. Is speech learning ‘gated’by the social brain? Developmental Science 10: 110–20. [Google Scholar] [CrossRef]
- Lehongre, Katia, Thierry Aubin, and Catherine Del Negro. 2009. Influence of social conditions in song sharing in the adult canary. Animal Cognition 12: 823–32. [Google Scholar] [CrossRef]
- Leitner, Stefan, and Clive K. Catchpole. 2007. Song and brain development in canaries raised under different conditions of acoustic and social isolation over two years. Developmental Neurobiology 67: 1478–87. [Google Scholar] [CrossRef] [PubMed]
- Li, Ping, and Hyeonjeong Jeong. 2020. The social brain of language: Grounding second language learning in social interaction. NPJ Science of Learning 5: 1–9. [Google Scholar] [CrossRef]
- Lipkind, Dina, Gary F. Marcus, Douglas K. Bemis, Kazutoshi Sasahara, Nori Jacoby, Miki Takahasi, Kenta Suzuki, Olga Feher, Primoz Ravbar, Kazuo Okanoya, and et al. 2013. Stepwise acquisition of vocal combinatorial capacity in songbirds and human infants. Nature 498: 104–8. [Google Scholar] [CrossRef]
- Lipkind, Dina, Anja T. Zai, Alexander Hanuschkin, Gary F. Marcus, Ofer Tchernichovski, and Richard H. R. Hahnloser. 2017. Songbirds work around computational complexity by learning song vocabulary independently of sequence. Nature Communications 8: 1247. [Google Scholar] [CrossRef]
- Lipkind, Dina, Andreea Geambasu, and Clara C. Levelt. 2020. The development of structured vocalizations in songbirds and humans: A comparative analysis. Topics in Cognitive Science 12: 894–909. [Google Scholar] [CrossRef] [PubMed]
- Ljubičić, Iva, Julia Hyland Bruno, and Ofer Tchernichovski. 2016. Social influences on song learning. Current Opinion in Behavioral Sciences 7: 101–7. [Google Scholar] [CrossRef]
- London, Sarah E., and David F. Clayton. 2008. Functional identification of sensory mechanisms required for developmental song learning. Nature Neuroscience 11: 579–86. [Google Scholar] [CrossRef]
- Love, Jay, Amanda Hoepfner, and Franz Goller. 2019. Song feature specific analysis of isolate song reveals interspecific variation in learned components. Developmental Neurobiology 79: 350–69. [Google Scholar] [CrossRef]
- Mack, Molly. 1989. Consonant and vowel perception and production: Early English-French bilinguals and English monolinguals. Perceptual Psychophysics 46: 187–200. [Google Scholar] [CrossRef] [PubMed]
- MacWhinney, Brian. 2005. A unified model of language acquisition. In Handbook of Bilingualism: Psycholinguistic Approaches. Edited by Judith F. Kroll and Annette M. B. de Groot. New York: Oxford University Press, pp. 49–67. [Google Scholar]
- Marchman, Virginia A. 1993. Constraints on plasticity in a connectionist model of the English Past tense. Journal of Cognitive Neuroscience 5: 215–34. [Google Scholar] [CrossRef]
- Margoliash, Daniel, and Marc F. Schmidt. 2010. Sleep, off-line processing, and vocal learning. Brain and Language 115: 45–58. [Google Scholar] [CrossRef]
- Margoliash, Daniel, Cynthia Staicer, and Sue A. Inoue. 1994. The process of syllable acquisition in adult indigo buntings (Passerina cyanea). Behaviour 131: 39–64. Available online: https://www.jstor.org/stable/4535227 (accessed on 13 December 2021).
- Marler, Peter. 1970. A comparative approach to vocal learning: Song development in white-crowned sparrows. Journal of Comparative and Physiological Psychology 71: 1–25. [Google Scholar] [CrossRef]
- Marler, Peter. 1997. Three models of song learning: Evidence from behavior. Journal of Neurobiology 33: 501–16. [Google Scholar] [CrossRef]
- Marler, Peter. 2004. Bird calls: Their potential for neurobiology. Annals of the New York Academy of Sciences 1016: 31–44. [Google Scholar] [CrossRef]
- Marler, Peter, and Miwako Tamura. 1962. Song “dialects” in three populations of white-crowned sparrows. Condor 64: 368–77. [Google Scholar] [CrossRef]
- Mayr, Robert, Sacha Price, and Ineke Mennen. 2012. First language attrition in the case of Dutch-English bilinguals: The case of monozygotic twin sisters. Bilingualism: Language and Cognition 15: 687–700. [Google Scholar] [CrossRef]
- Mets, David G., and Michael S. Brainard. 2018. Genetic variation interacts with experience to determine interindividual differences in learned song. Proceedings of the National Academy of Sciences of the United States of America 115: 421–26. [Google Scholar] [CrossRef]
- Mets, David G., and Michael S. Brainard. 2019. Learning is enhanced by tailoring instruction to individual genetic differences. eLife 8: e47216. [Google Scholar] [CrossRef] [PubMed]
- Minagawa-Kawai, Yasuyo, Cristià Alejandrina, and Emmanuel Dupoux. 2011. Cerebral lateralization and early speech acquisition: A developmental scenario. Developmental Cognitive Neuroscience 1: 217–32. [Google Scholar] [CrossRef]
- Mol, Carien, Aoju Chen, René W. J. Kager, and Sita M. ter Haar. 2017. Prosody in birdsong: A review and perspective. Neuroscience & Biobehavioral Reviews 81: 167–80. [Google Scholar] [CrossRef]
- Mooney, Richard, William Hoese, and Stephen Nowicki. 2001. Auditory representation of the vocal repertoire in a songbird with multiple song types. Proceedings of the National Academy of Sciences of the United States of America 98: 12778–83. [Google Scholar] [CrossRef] [PubMed]
- Moore, Jordan M., and Sarah M. N. Woolley. 2019. Emergent tuning for learned vocalizations in auditory cortex. Nature Neuroscience 22: 1469–76. [Google Scholar] [CrossRef] [PubMed]
- Moore, Jordan M., Tamás Székely, József Büki, and Timothy J. DeVoogd. 2011. Motor pathway convergence predicts syllable repertoire size in oscine birds. Proceedings of the National Academy of Sciences of the United States of America 108: 16440–45. [Google Scholar] [CrossRef]
- Moorman, Sanne, Sharon MH Gobes, Maaike Kuijpers, Amber Kerkhofs, Matthijs A. Zandbergen, and Johan J. Bolhuis. 2012. Human-like brain hemispheric dominance in birdsong learning. Proceedings of the National Academy of Sciences 109: 12782–87. [Google Scholar] [CrossRef]
- Mora, Joan C., Youssef Rochdi, and Hanna Kivistö-de-Souza. 2013. Mimicking accented speech as L2 phonological awareness. Language Awareness 23: 57–75. [Google Scholar] [CrossRef]
- Morrison, Robert G., and Fernando Nottebohm. 1993. Role of a telencephalic nucleus in the delayed song learning of socially isolated zebra finches. Journal of Neurobiology 24: 1045–64. [Google Scholar] [CrossRef]
- Moyer, Alene. 2004. Age, Accent and Experience in Second Language Acquisition: An Integrated Approach to Critical Period Inquiry. Clevedon: Multilingual Matters. [Google Scholar]
- Moyer, Alene. 2014. Exceptional outcomes in L2 phonology: The critical factors of learner engagement and self-regulation. Applied Linguistics 35: 418–40. [Google Scholar] [CrossRef]
- Murphy, Karagh, Logan S. James, Jon T. Sakata, and Jonathan F. Prather. 2017. Advantages of comparative studies to understand the neural basis of sensorimotor integration. Journal of Neurophysiology 118: 800–16. [Google Scholar] [CrossRef] [PubMed]
- Murphy, Karagh, Koedi Lawley, Perry Smith, and Jonathan F. Prather. 2020. New insights into the avian song system and neuronal control of learned vocalizations. In The Neuroethology of Birdsong. Edited by Jon T. Sakata, Sarah C. Woolley, Richard R. Fay and Arthur N. Popper. Switzerland: Springer Nature, pp. 65–92. [Google Scholar] [CrossRef]
- Muysken, Pieter. 2000. Bilingual Speech: A Typology of Code-Mixing. Cambridge: Cambridge University Press. [Google Scholar] [CrossRef]
- Myers-Scotton, Carol. 1993. Social Motivations for Codeswitching: Evidence from Africa. Oxford: Clarendon. [Google Scholar]
- Nelson, Douglas A., and Peter Marler. 1994. Selection-based learning in bird song development. Proceedings of the National Academy of Sciences of the United States of America 91: 10498–501. [Google Scholar] [CrossRef] [PubMed]
- Nick, Teresa A., and Masakazu Konishi. 2005a. Neural auditory selectivity develops in parallel with song. Journal of Neurobiology 62: 469–81. [Google Scholar] [CrossRef] [PubMed]
- Nick, Teresa A., and Masakazu Konishi. 2005b. Neural song preference during vocal learning in the zebra finch depends on age and state. Journal of Neurobiology 62: 231–42. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, Stephen, and William A. Searcy. 2014. The evolution of vocal learning. Current Opinion in Neurobiology 28: 48–53. [Google Scholar] [CrossRef] [PubMed]
- Odom, Karan J., Michelle L. Hall, Katharina Riebel, Kevin E. Omland, and Naomi E. Langmore. 2014. Female song is widespread and ancestral in songbirds. Nature Communications 5: 1–6. [Google Scholar] [CrossRef]
- Oh, Janet S., Terry Kit-Fong Au, and Sun-Ah Jun. 2010. Early childhood language memory in the speech perception of international adoptees. Journal of Child Language 37: 1123–32. [Google Scholar] [CrossRef]
- O’Loghlen, Adrian L., and Stephen I. Rothstein. 2010. Delayed sensory learning and development of dialect songs in brown-headed cowbirds, Molothrus ater. Animal Behaviour 79: 299–311. [Google Scholar] [CrossRef]
- Olson, Elizabeth M., Rie K. Maeda, and Sharon M. H. Gobes. 2016. Mirrored patterns of lateralized neuronal activation reflect old and new memories in the avian auditory cortex. Neuroscience 330: 395–402. [Google Scholar] [CrossRef]
- Pallier, Christophe, Stanislas Dehaene, Jean-Baptiste Poline, Denis Le Bihan, A.-M. Argenti, Emmanuel Dupoux, and Jacques Mehler. 2003. Brain imaging of language plasticity in adopted adults: Can a second language replace the first? Cerebral Cortex 13: 155–61. [Google Scholar] [CrossRef]
- Park, Denise C., Thad A. Polk, Joseph A. Mikels, Stephan F. Taylor, and Christy Marshuetz. 2001. Cerebral aging: Integration of brain and behavioral models of cognitive function. Dialogues in Clinical Neuroscience 3: 151–65. [Google Scholar] [CrossRef] [PubMed]
- Paul, Avishek, Helen McLendon, Veronica Rally, Jon T. Sakata, and Sarah C. Woolley. 2021. Behavioral discrimination and time-series phenotyping of birdsong performance. PLoS Computational Biology 17: e1008820. [Google Scholar] [CrossRef]
- Petitto, Laura-Ann, Melody S. Berens, Ioulia Kovelman, M. H. Dubins, Kaja K. Jasinska, and M. Shalinsky. 2012. The “perceptual wedge hypothesis” as the basis for bilingual babies’ phonetic processing advantage: New insights from fNIRS brain imaging. Brain and Language 121: 130–43. [Google Scholar] [CrossRef]
- Petkov, Christopher I., and Erich Jarvis. 2012. Birds, primates, and spoken language origins: Behavioral phenotypes and neurobiological substrates. Frontiers in Evolutionary Neuroscience 4: 12. [Google Scholar] [CrossRef]
- Pfaff, Jeremy A., Liana Zanette, Scott A. MacDougall-Shackleton, and Elizabeth A. MacDougall-Shackleton. 2007. Song repertoire size varies with HVC volume and is indicative of male quality in song sparrows (Melospiza melodia). Proceedings of the Royal Society B: Biological Sciences 274: 2035–40. [Google Scholar] [CrossRef]
- Pierce, Lara J., Denise Klein, Jen-Kai Chen, Audrey Delcenserie, and Fred Genesee. 2014. Mapping the unconscious maintenance of a lost first language. Proceedings of the National Academy of Sciences of the United States of America 111: 17314–19. [Google Scholar] [CrossRef]
- Piller, Ingrid. 2002. Passing for a native speaker: Identity and success in second language learning. Journal of Sociolinguistics 6: 179–206. [Google Scholar] [CrossRef]
- Piske, Thorsten, Ian R. A. MacKay, and James E. Flege. 2001. Factors affecting degree of foreign accent in an L2: A review. Journal of Phonetics 29: 191–215. [Google Scholar] [CrossRef]
- Pizzorusso, Tommaso, Paolo Medini, Nicoletta Berardi, Sabrina Chierzi, James W. Fawcett, and Lamberto Maffei. 2002. Reactivation of ocular dominance plasticity in the adult visual cortex. Science 298: 1248–51. [Google Scholar] [CrossRef]
- Podos, Jeffrey, and Ha-Cheol Sung. 2020. Vocal performance in songbirds: From mechanisms to evolution. In The Neuroethology of Birdsong. Edited by Jon T. Sakata, Sarah C. Woolley, Richard R. Fay and Arthur N. Popper. Cham: Springer Nature, pp. 245–68. [Google Scholar] [CrossRef]
- Poplack, Shana. 1980. “Sometimes I’ll start a sentence in Spanish y termino en español”: Toward a typology of code-switching. Linguistics 18: 581–618. [Google Scholar] [CrossRef]
- Prather, Jonathan F., Susan Peters, Stephen Nowicki, and Richard Mooney. 2010. Persistent representation of juvenile experience in the adult songbird brain. Journal of Neuroscience 30: 10586–98. [Google Scholar] [CrossRef]
- Prather, Jonathan F., Kazuo Okanoya, and Johan J. Bolhuis. 2017. Brains for birds and babies: Neural parallels between birdsong and speech acquisition. Neuroscience & Biobehavioral Reviews 81: 225–37. [Google Scholar] [CrossRef]
- Pytte, Carolyn L., Miles Gerson, Janet Miller, and John R. Kirn. 2007. Increasing stereotypy in adult zebra finch song correlates with a declining rate of adult neurogenesis. Developmental Neurobiology 67: 1699–720. [Google Scholar] [CrossRef] [PubMed]
- Ravbar, Primoz, Dina Lipkind, Lucas C. Parra, and Ofer Tchernichovski. 2012. Vocal exploration is locally regulated during song learning. Journal of Neuroscience 32: 3422–32. [Google Scholar] [CrossRef]
- Reichelt, Amy C., Dominic J. Hare, Timothy J. Bussey, and Lisa M. Saksida. 2019. Perineuronal nets: Plasticity, protection, and therapeutic potential. Trends in Neurosciences 42: 458–70. [Google Scholar] [CrossRef] [PubMed]
- Reiterer, Susanne Maria, Xiaochen Hu, Michael Erb, Guiseppina Rota, Davide Nardo, Wolfgang Grodd, Susanne Winkler, and Hermann Ackermann. 2011. Individual differences in audio-vocal speech imitation aptitude in late bilinguals: Functional neuro-imaging and brain morphology. Frontiers in Psychology 2: 271. [Google Scholar] [CrossRef] [PubMed]
- Reiterer, Susanne M., Xiaochen Hu, T. A. Sumathi, and Nandini C. Singh. 2013. Are you a good mimic? Neuro-acoustic signatures for speech imitation ability. Frontiers in Psychology 4: 782. [Google Scholar] [CrossRef] [PubMed]
- Riebel, Katharina, Karan J. Odom, Naomi E. Langmore, and Michelle L. Hall. 2019. New insights from female bird song: Towards an integrated approach to studying male and female communication roles. Biology Letters 15: 20190059. [Google Scholar] [CrossRef] [PubMed]
- Ripollés, Pablo, Josep Marco-Pallarés, Helena Alicart, Claus Tempelmann, Antonio Rodríguez-Fornells, and Toemme Noesselt. 2016. Intrinsic monitoring of learning success facilitates memory encoding via the activation of the SN/VTA-Hippocampal loop. eLife 5: e17441. [Google Scholar] [CrossRef]
- Roberts, Todd F., and Richard Mooney. 2013. Motor circuits help encode auditory memories of vocal models used to guide vocal learning. Hearing Research 303: 48–57. [Google Scholar] [CrossRef] [PubMed]
- Robinson, Cristina M., and Nicole Creanza. 2019. Species-level repertoire size predicts a correlation between individual song elaboration and reproductive success. Ecology and Evolution 9: 8362–77. [Google Scholar] [CrossRef]
- Robinson, Cristina M., Kate T. Snyder, and Nicole Creanza. 2019. Correlated evolution between repertoire size and song plasticity predicts that sexual selection on song promotes open-ended learning. eLife 8: e44454. [Google Scholar] [CrossRef] [PubMed]
- Rota, Guiseppina, and Susanne Maria Reiterer. 2009. Cognitive aspects of pronunciation talent. In Language Talent and Brain Activity. Edited by Grzegorz Dogil and Susanne Maria Reiterer. Berlin: Mouton de Gruyter, pp. 67–96. [Google Scholar] [CrossRef]
- Sainburg, Tim, Marvin Thielk, and Timothy Q. Gentner. 2020. Finding, visualizing, and quantifying latent structure across diverse animal vocal repertoires. PLoS Computational Biology 16: e1008228. [Google Scholar] [CrossRef]
- Sakata, Jon T., and Michael S. Brainard. 2006. Real-time contributions of auditory feedback to avian vocal motor control. Journal of Neuroscience 26: 9619–28. [Google Scholar] [CrossRef]
- Sakata, Jon T., and Michael S. Brainard. 2008. Online contributions of auditory feedback to neural activity in avian song control circuitry. Journal of Neuroscience 28: 11378–90. [Google Scholar] [CrossRef][Green Version]
- Sakata, Jon T., and Sandra L. Vehrencamp. 2012. Integrating perspectives on vocal performance and consistency. Journal of Experimental Biology 215: 201–9. [Google Scholar] [CrossRef]
- Sakata, Jon T., and Sarah C. Woolley. 2020. Scaling the levels of birdsong analysis. In The Neuroethology of Birdsong. Edited by Jon T. Sakata, Sarah C. Woolley, Richard R. Fay and Arthur N. Popper. Cham: Springer Nature, pp. 1–27. [Google Scholar] [CrossRef]
- Sakata, Jon T., and Yoko Yazaki-Sugiyama. 2020. Neural circuits underlying vocal learning in songbirds. In The Neuroethology of Birdsong. Edited by Jon T. Sakata, Sarah C. Woolley, Richard R. Fay and Arthur N. Popper. Cham: Springer Nature, pp. 29–63. [Google Scholar] [CrossRef]
- Schepens, Job Johannes, Roeland van Hout, and T. Florian Jaeger. 2020. Big data suggest strong constraints of linguistic similarity on adult language learning. Cognition 194: 104056. [Google Scholar] [CrossRef]
- Schmid, Monika S., and Barbara Köpke. 2017. The relevance of first language attrition to theories of bilingual development. Linguistic Approaches to Bilingualism 7: 637–67. [Google Scholar] [CrossRef]
- Schmid, Monika S., Barbara Köpke, Merel Keijzer, and Lina Weilemar, eds. 2004. First Language Attrition: Interdisciplinary Perspectives on Methodological Issues. Amsterdam: John Benjamins. [Google Scholar]
- Seyfarth, Robert M., and Dorothy L. Cheney. 2010. Production, usage, and comprehension in animal vocalizations. Brain and Language 115: 92–100. [Google Scholar] [CrossRef]
- Simmonds, Anna J. 2015. A hypothesis on improving foreign accents by optimizing variability in vocal learning brain circuits. Frontiers in Human Neuroscience 9: 606. [Google Scholar] [CrossRef][Green Version]
- Singleton, David. 1995. Introduction: A critical look at the Critical Period Hypothesis in Second Language Acquisition Research. In The Age Factor in Second Language Acquisition: A Critical Look at the Critical Period Hypothesis. Edited by David Singleton and Zsolt Lengyel. Clevedon: Multilingual Matters, pp. 1–29. [Google Scholar]
- Slevc, L. Robert, and Akira Miyake. 2006. Individual differences in second-language proficiency. Does musical ability matter? Psychological Science 17: 675–81. [Google Scholar] [CrossRef] [PubMed]
- Soha, Jill. 2017. The auditory template hypothesis: A review and comparative perspective. Animal Behaviour 124: 247–54. [Google Scholar] [CrossRef]
- Steinhauer, Karsten, and Kristina Kasparian. 2020. Brain plasticity in adulthood: ERP evidence for L1-attrition in lexicon and morphosyntax after predominant L2 use. Language Learning 70: 171–93. [Google Scholar] [CrossRef]
- Steinhauer, Karsten, Erin J. White, and John E. Drury. 2009. Temporal dynamics of late second language acquisition: Evidence from event-related brain potentials. Second Language Research 25: 13–41. [Google Scholar] [CrossRef]
- Székely, Tamás, Clive K. Catchpole, Albert DeVoogd, Zsuzsa Marchl, and Timothy J. DeVoogd. 1996. Evolutionary changes in a song control area of the brain (HVC) are associated with evolutionary changes in song repertoire among European warblers (Sylviidae). Proceedings of the Royal Society of London. Series B: Biological Sciences 263: 607–10. [Google Scholar] [CrossRef]
- Tachibana, Ryosuke O., Miki Takahasi, Neal A. Hessler, and Kazuo Okanoya. 2017. Maturation-dependent control of vocal temporal plasticity in a songbird. Developmental Neurobiology 77: 995–1006. [Google Scholar] [CrossRef]
- Tagarelli, Kaitlyn M., Kyle F. Shattuck, Peter E. Turkeltaub, and Michael T. Ullman. 2019. Language learning in the adult brain: A neuroanatomical meta-analysis of lexical and grammatical learning. Neuroimage 193: 178–200. [Google Scholar] [CrossRef]
- Takesian, Anne E., and Takao K. Hensch. 2013. Balancing plasticity/stability across brain development. Progress in Brain Research 207: 3–34. [Google Scholar] [CrossRef]
- Tchernichovski, Ofer, Partha P. Mitra, Thierry Lints, and Fernando Nottebohm. 2001. Dynamics of the vocal imitation process: How a zebra finch learns its song. Science 291: 2564–69. [Google Scholar] [CrossRef] [PubMed]
- Tchernichovski, Ofer, Sophie Eisenberg-Edidin, and Erich D. Jarvis. 2021. Balanced imitation sustains song culture in zebra finches. Nature Communications 12: 1–14. [Google Scholar] [CrossRef]
- ten Cate, Carel, and Peter J. Fullagar. 2021. Vocal imitations and production learning by Australian musk ducks (Biziura lobata). Philosophical Transactions of the Royal Society B 376: 20200243. [Google Scholar] [CrossRef]
- Theakston, Anna L. 2017. Entrenchment in first language learning. In Entrenchment and the Psychology of Language Learning: How We Reorganize and Adapt Linguistic Knowledge. Edited by Hans-Jörg Schmid. Washington, DC: American Psychological Association, pp. 315–41. [Google Scholar] [CrossRef]
- Thorpe, William H. 1958. The learning of song patterns by birds, with especial reference to the song of the chaffinch Fringilla coelebs. Ibis 100: 535–70. [Google Scholar] [CrossRef]
- Todt, Dietmar, and Nicole Geberzahn. 2003. Age-dependent effects of song exposure: Song crystallization sets a boundary between fast and delayed vocal imitation. Animal Behaviour 65: 971–79. [Google Scholar] [CrossRef]
- Todt, Dietmar, Henrike Hultsch, and Dietmar Heike. 1979. Conditions affecting song acquisition in nightingales (Luscinia megarhynchos L.). Ethology 51: 23–35. [Google Scholar] [CrossRef]
- Tomasello, Michael. 2005. Constructing a Language: A Usage-Based Theory of Language Acquisition. Cambridge: Harvard University Press. [Google Scholar]
- Tschida, Katherine, and Richard Mooney. 2012. The role of auditory feedback in vocal learning and maintenance. Current Opinion in Neurobiology 22: 320–27. [Google Scholar] [CrossRef] [PubMed]
- Tumer, Evren C., and Michael S. Brainard. 2007. Performance variability enables adaptive plasticity of ‘crystallized’ adult birdsong. Nature 450: 1240–44. [Google Scholar] [CrossRef] [PubMed]
- Vallentin, Daniela, Georg Kosche, Dina Lipkind, and Michael A. Long. 2016. Inhibition protects acquired song segments during vocal learning in zebra finches. Science 351: 267–71. [Google Scholar] [CrossRef]
- Ventureyra, Valerie A. G., Christophe Pallier, and Hi-Yon Yoo. 2004. The loss of first language phonetic perception in adopted Koreans. Journal of Neurolinguistics 17: 79–91. [Google Scholar] [CrossRef]
- Vernes, Sonja C., Buddhamas Pralle Kriengwatana, Veronika C. Beeck, Julia Fischer, Peter L. Tyack, Carel Ten Cate, and Vincent M. Janik. 2021. The multi-dimensional nature of vocal learning. Philosophical Transactions of the Royal Society B 376: 20200236. [Google Scholar] [CrossRef]
- Walters, Keith. 2011. Gendering French in Tunisia: Language ideologies and nationalism. International Journal of The Sociology of Language 211: 83–111. [Google Scholar] [CrossRef]
- Wang, Difei, and James Fawcett. 2012. The perineuronal net and the control of CNS plasticity. Cell and Tissue Research 349: 147–60. [Google Scholar] [CrossRef]
- Warren, Timothy L., Jonathan D. Charlesworth, Evren C. Tumer, and Michael S. Brainard. 2012. Variable sequencing is actively maintained in a well learned motor skill. Journal of Neuroscience 32: 15414–25. [Google Scholar] [CrossRef]
- Watkins, Michael, Andreia S. Rauber, and Barbara O. Baptista. 2009. Recent Research in Second Language Phonetics/Phonology: Perception and Production. Cambridge: Cambridge Scholars Publishing. [Google Scholar]
- Werker, Janet F., and Takao K. Hensch. 2015. Critical periods in speech perception: New directions. Annual Review of Psychology 66: 173–96. [Google Scholar] [CrossRef]
- West, Meredith J., and Andrew P. King. 1988. Female visual displays affect the development of male song in the cowbird. Nature 334: 244–46. [Google Scholar] [CrossRef]
- Wiley, R. Haven. 2000. A new sense of the complexities of bird song. The Auk 117: 861–68. [Google Scholar] [CrossRef]
- Wirthlin, Morgan, Edward F. Chang, Mirjam Knörnschild, Leah A. Krubitzer, Claudio V. Mello, Cory T. Miller, Andreas R. Pfenning, Sonja C. Vernes, Ofer Tchernichovski, and Michael M. Yartsev. 2019. A modular approach to vocal learning: Disentangling the diversity of a complex behavioral trait. Neuron 104: 87–99. [Google Scholar] [CrossRef]
- Wong, Patrick C. M., Kara Morgan-Short, Marc Ettlinger, and Jing Zheng. 2012. Linking neurogenetics and individual differences in language learning: The dopamine hypothesis. Cortex 48: 1091–102. [Google Scholar] [CrossRef]
- Woolley, Sarah C., and Sarah M. N. Woolley. 2020. Integrating form and function in the songbird auditory forebrain. In The Neuroethology of Birdsong. Edited by Jon T. Sakata, Sarah C. Woolley, Richard R. Fay and Arthur N. Popper. Cham: Springer Nature, pp. 127–55. [Google Scholar] [CrossRef]
- Yamaguchi, Ayako. 2001. Sex differences in vocal learning in birds. Nature 411: 257–58. [Google Scholar] [CrossRef]
- Yanagihara, Shin, and Yoko Yazaki-Sugiyama. 2016. Auditory experience-dependent cortical circuit shaping for memory formation in bird song learning. Nature Communications 7: 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yazaki-Sugiyama, Yoko, and Richard Mooney. 2004. Sequential learning from multiple tutors and serial retuning of auditory neurons in a brain area important to birdsong learning. Journal of Neurophysiology 92: 2771–88. [Google Scholar] [CrossRef]
- Yeni-Komshian, Grace H., James E. Flege, and Serena Liu. 2000. Pronunciation proficiency in the first and second languages of Korean-English bilinguals. Bilingualism: Language and Cognition 3: 131–49. [Google Scholar] [CrossRef]
- Yeung, H. Henry, and Janet F. Werker. 2009. Learning words’ sounds before learning how words sound: 9-month-olds use distinct objects as cues to categorize speech information. Cognition 113: 234–43. [Google Scholar] [CrossRef]
- Yu, K., W. E. Wood, and F. E. Theunissen. 2020. High-capacity auditory memory for vocal communication in a social songbird. Science Advances 6: eabe0440. [Google Scholar] [CrossRef]
- Zhang, Xiaopeng, and Chunping Mai. 2018. Effects of entrenchment and preemption in second language learners’ acceptance of English denominal verbs. Applied Psycholinguistics 39: 413–36. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakata, J.T.; Birdsong, D. Vocal Learning and Behaviors in Birds and Human Bilinguals: Parallels, Divergences and Directions for Research. Languages 2022, 7, 5. https://doi.org/10.3390/languages7010005
Sakata JT, Birdsong D. Vocal Learning and Behaviors in Birds and Human Bilinguals: Parallels, Divergences and Directions for Research. Languages. 2022; 7(1):5. https://doi.org/10.3390/languages7010005
Chicago/Turabian StyleSakata, Jon T., and David Birdsong. 2022. "Vocal Learning and Behaviors in Birds and Human Bilinguals: Parallels, Divergences and Directions for Research" Languages 7, no. 1: 5. https://doi.org/10.3390/languages7010005
APA StyleSakata, J. T., & Birdsong, D. (2022). Vocal Learning and Behaviors in Birds and Human Bilinguals: Parallels, Divergences and Directions for Research. Languages, 7(1), 5. https://doi.org/10.3390/languages7010005