
Projected Impacts of Climate and Land Use Change on Endemic Plant Distributions in a Mediterranean Island Hotspot: The Case of Evvia (Aegean, Greece)

 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- (a)
- evaluate species-specific responses to global change drivers, using SDMs and incorporating both climate and land use projections;
- (b)
- identify and project shifts in biodiversity hotspots over time, using a combination of taxonomic and phylogenetic diversity metrics;
- (c)
- provide insights for evidence-based conservation and natural capital management strategies, by analysing projected range changes, fragmentation, and extinction risks;
- (d)
- evaluate the effectiveness of existing protected areas and identify conservation gaps in Evvia under future climate and land use change scenarios, by overlaying projected biodiversity hotspots with the current protected area network;
- (e)
- estimate the current and future extinction risk of the single-island endemic species of Evvia, using SDM projections and IUCN Red List criteria.
2. Materials and Methods
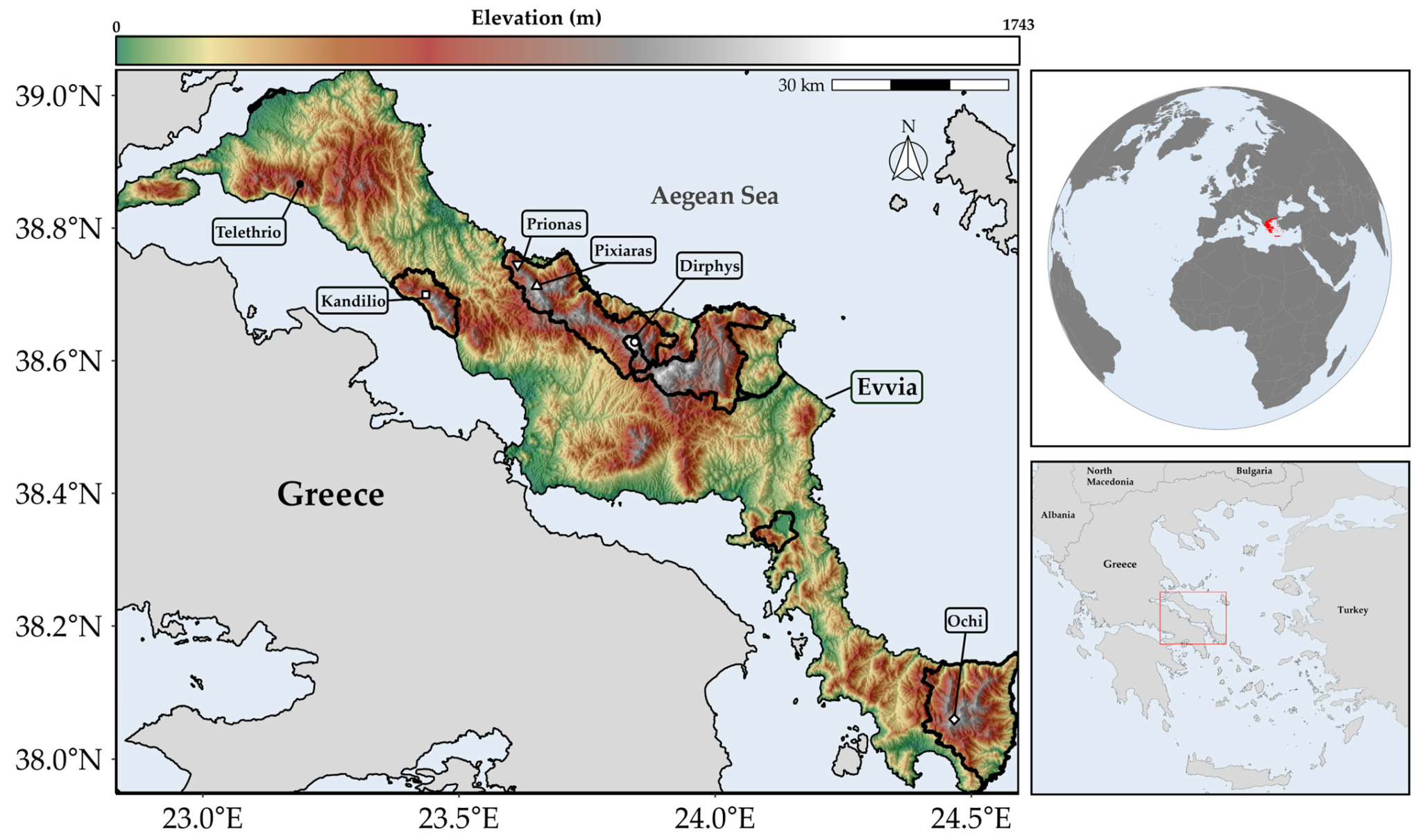
2.1. Species Occurrence Data
2.2. Environmental Data
- 2020s: 2011–2040.
- 2050s: 2041–2070.
- 2080s: 2071–2100.
2.3. Species Distribution Models
- EOO ≤ 1st Quartile: We used the 12.5th percentile of the Shape metric’s extrapolation values as the threshold (most conservative).
- 1st Quartile < EOO ≤ Median (2nd Quartile): We used the 25th percentile of the extrapolation values.
- Median < EOO ≤ 3rd Quartile: We used the 50th percentile of the extrapolation values.
- EOO > 3rd Quartile: We used the 75th percentile of the extrapolation values (least conservative).
ENphylo Modelling
2.4. Biodiversity Hotspot Detection
2.5. Temporal Beta Diversity
2.6. Assessment of Protected Area Effectiveness and Conservation Gaps in Evvia
2.7. Land Use and Land Cover Changes
2.8. Preliminary IUCN Extinction Risk Assessment
2.9. Estimation of the Evolutionarily Distinct and Globally Endangered (EDGE) Index–Current and Future EDGE Spatial Patterns
3. Results
3.1. Species Distribution Models
- (a)
- Occurrence in specific land use categories (forests, grasslands, and shrubs), potential evapotranspiration of the driest quarter, Thornthwaite’s aridity index, count of the number of months with mean temp greater than 10 °C, temperature annual range, and mean diurnal range for the Greek endemics (Table S3; Figure S3), likely reflecting their adaptation to the dry, rocky habitats, and temperature extremes characteristic of the Mediterranean climate and
- (b)
- Occurrence in specific land use categories (barren and grasslands), potential evapotranspiration of the driest quarter, and continentality for the single-island endemics (Table S3; Figure S3), likely reflecting their adaptation to dry, rocky habitats and sensitivity to temperature fluctuations and heat stress in their restricted island ranges
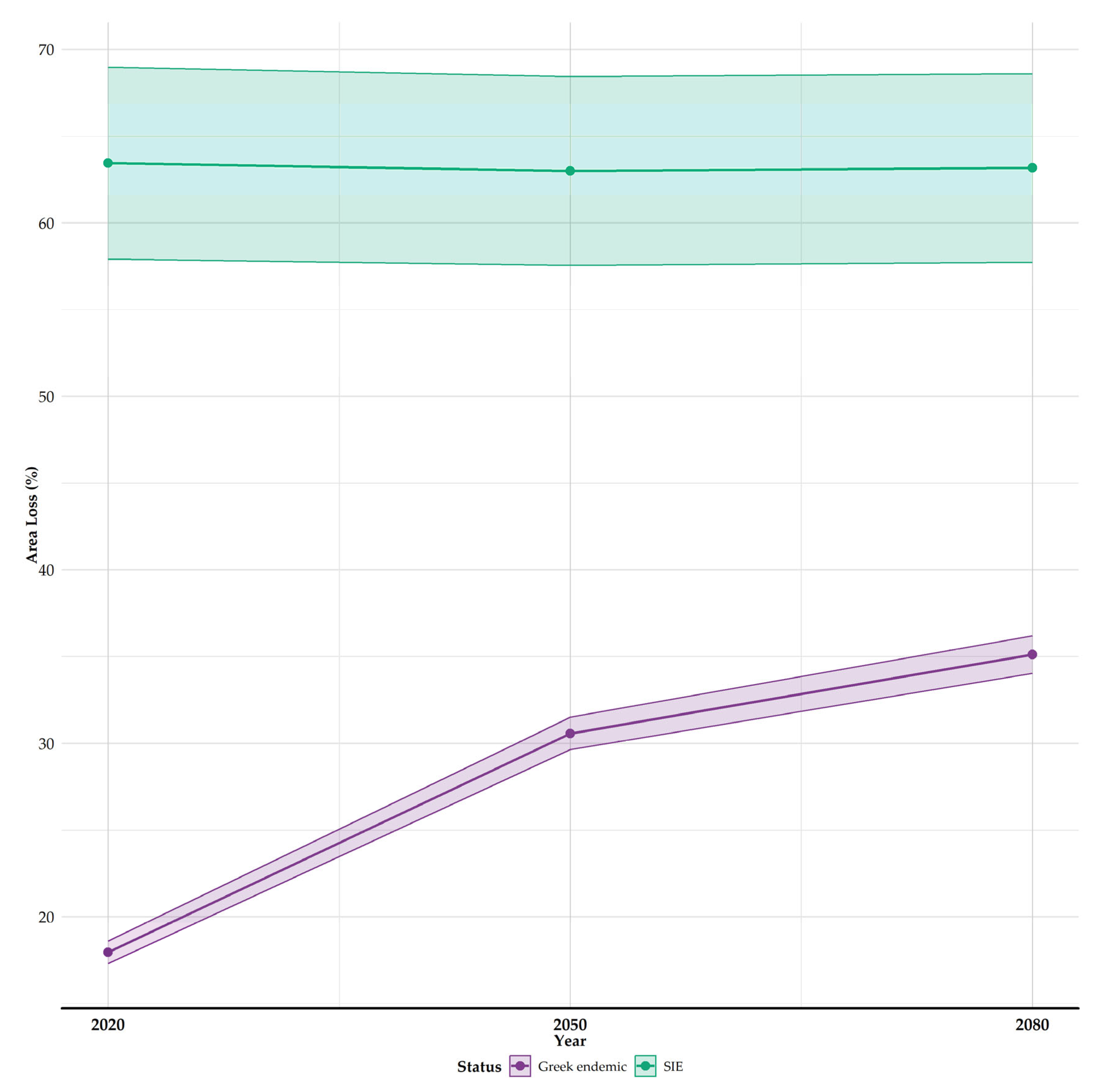
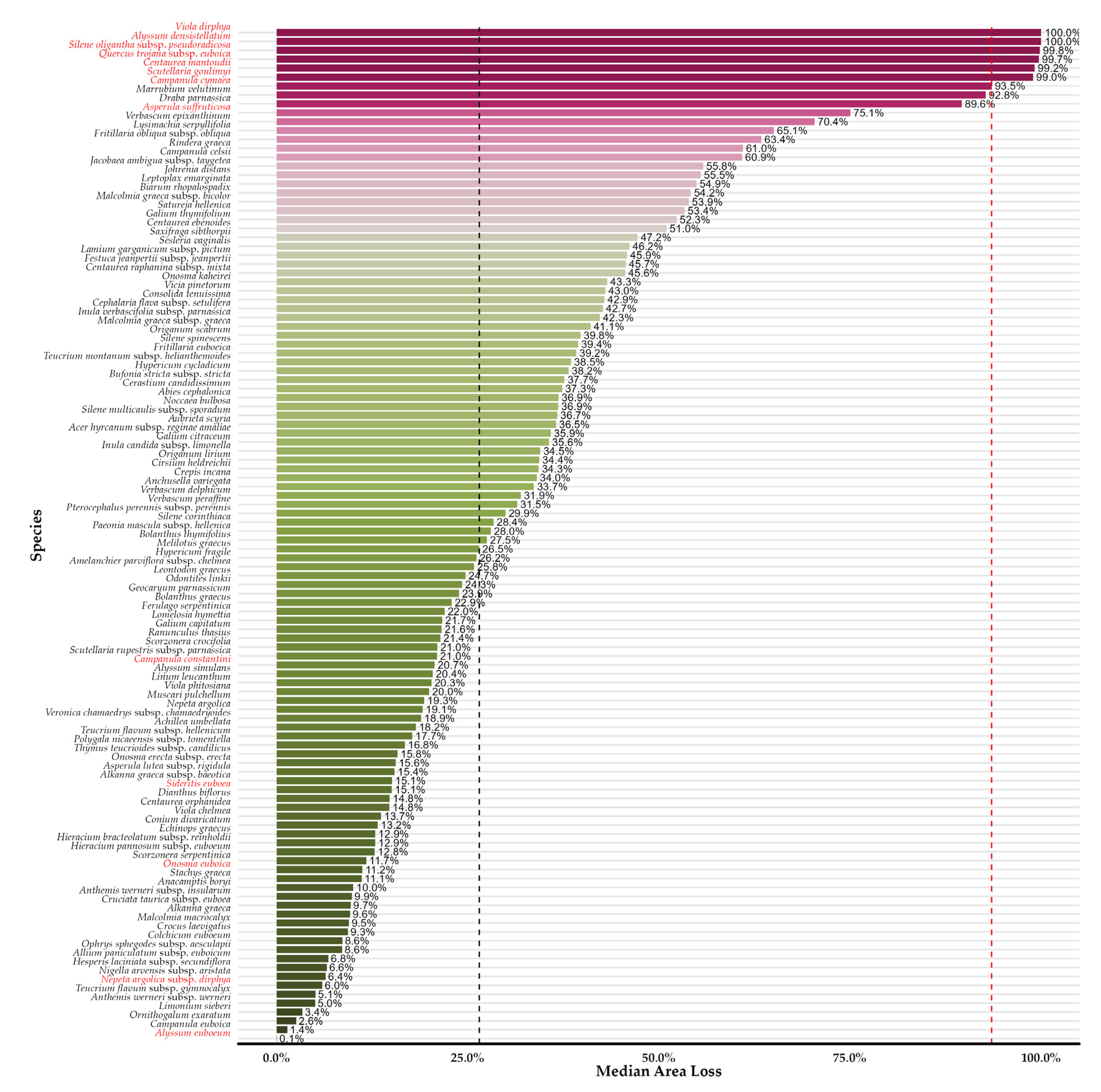
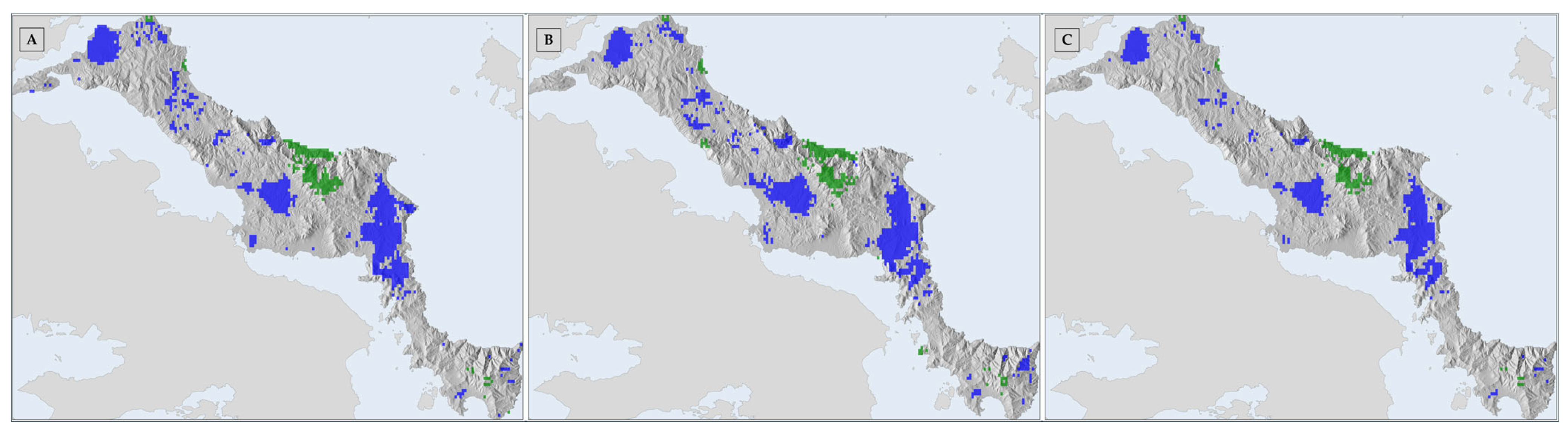
3.2. Habitat Suitability Range Change
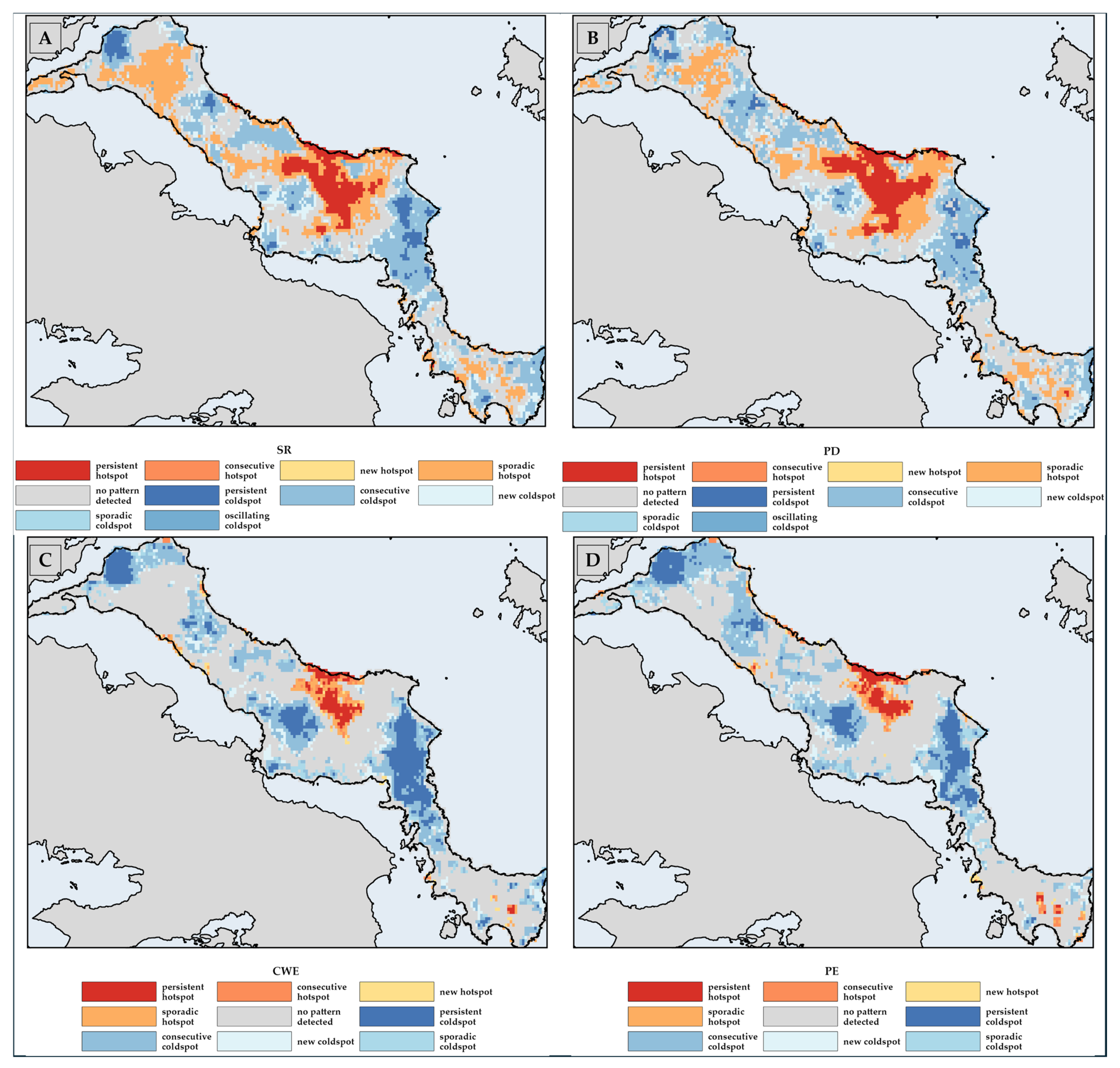
3.3. Biodiversity Hotspots
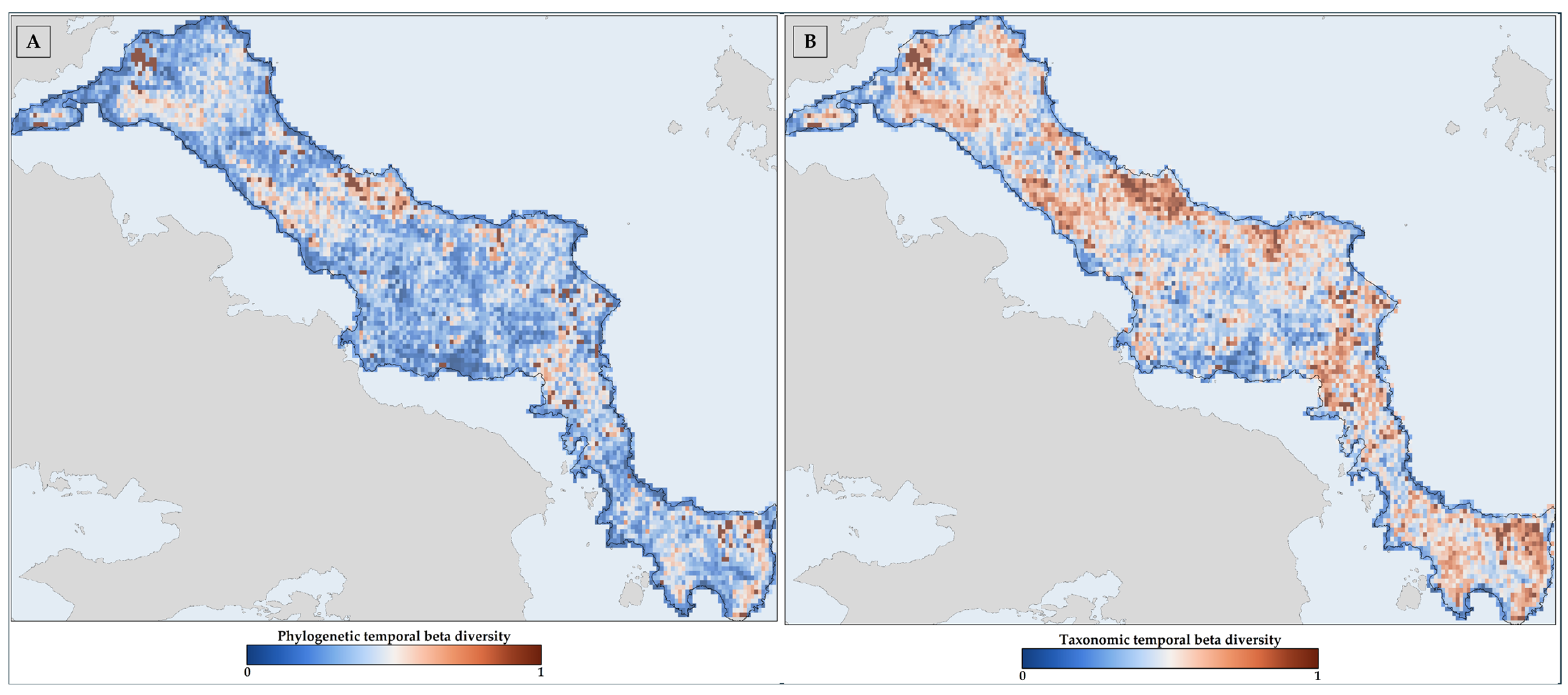
3.4. Temporal Beta Diversity
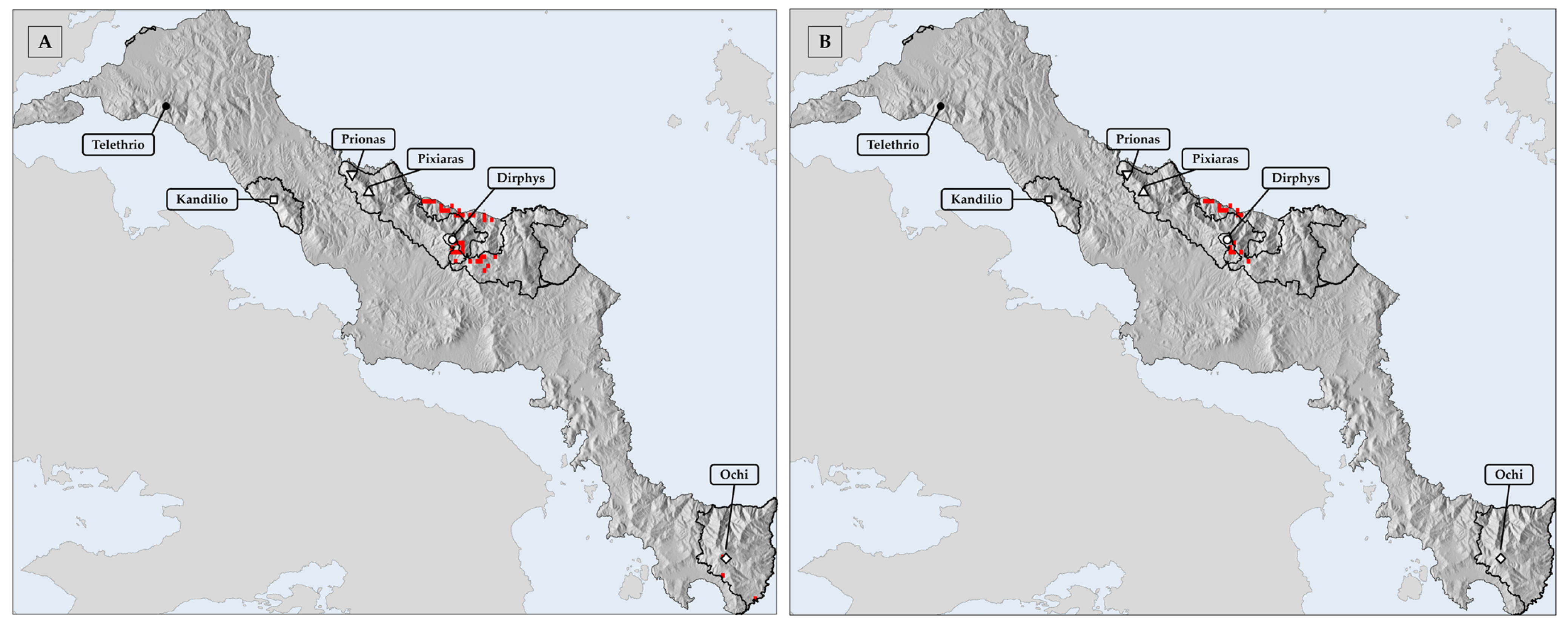
3.5. Assessment of Protected Area Effectiveness and Conservation Gaps in Evvia
3.6. Land Use and Land Cover Changes
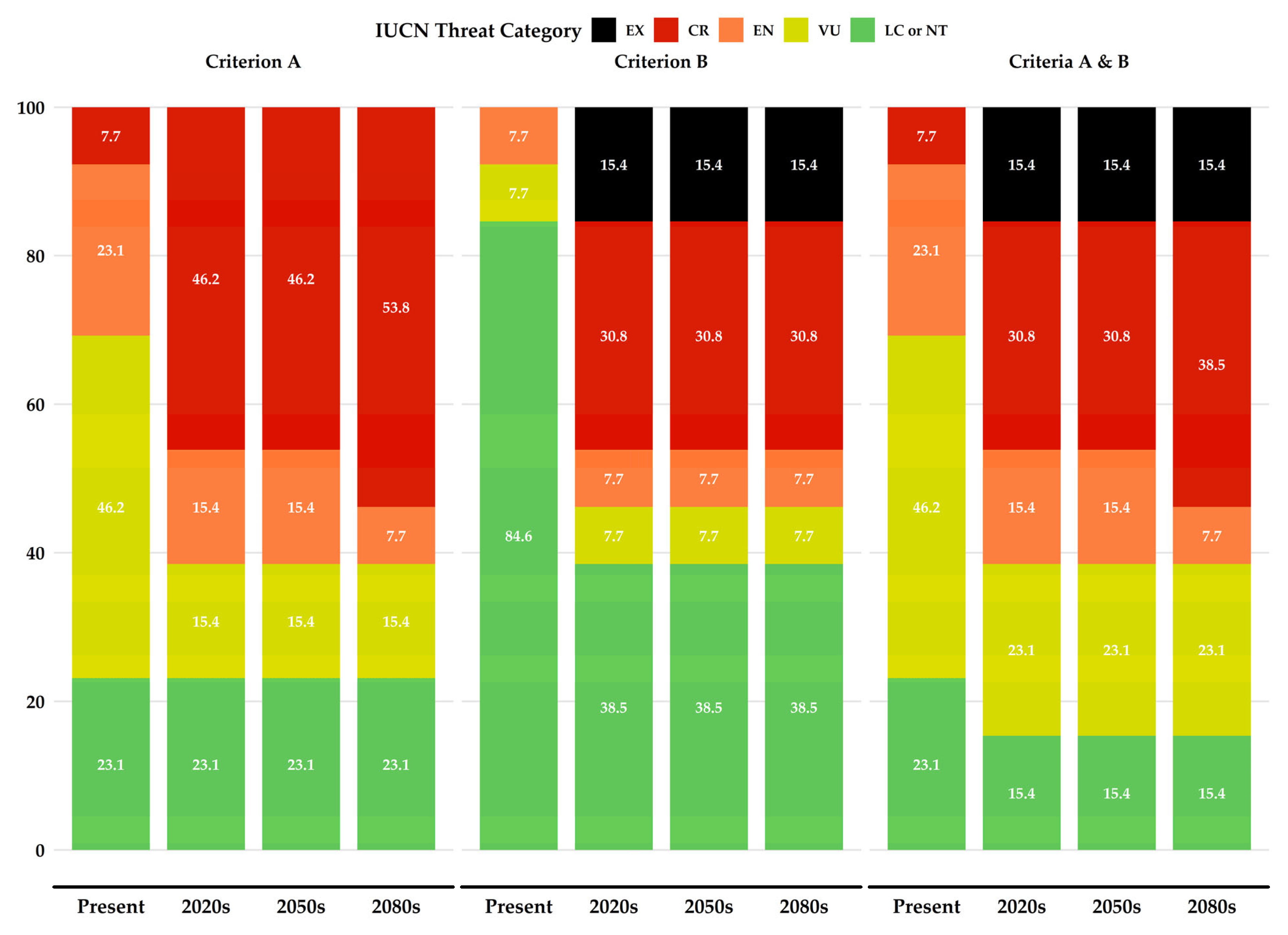
3.7. IUCN Extinction Risk Assessment
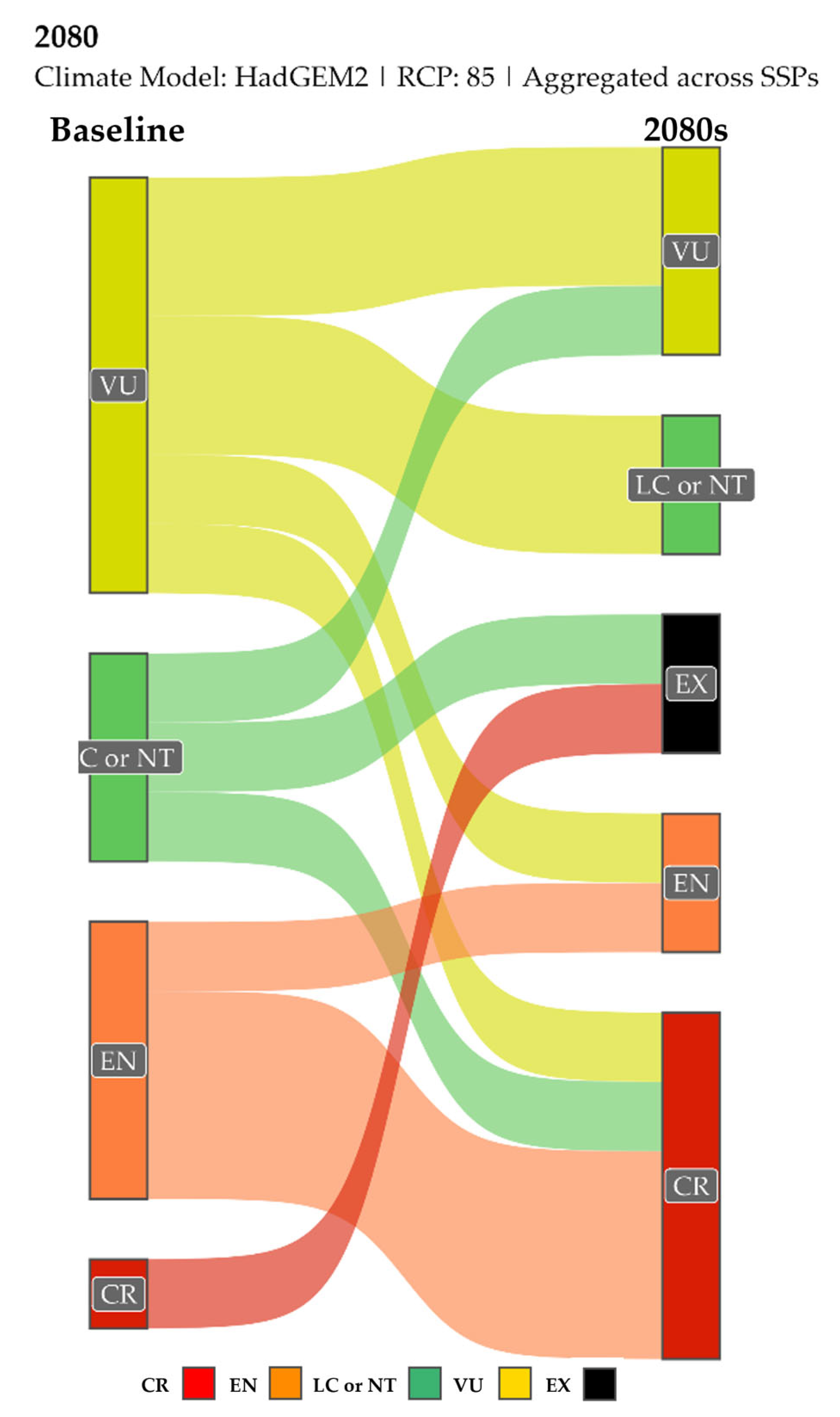
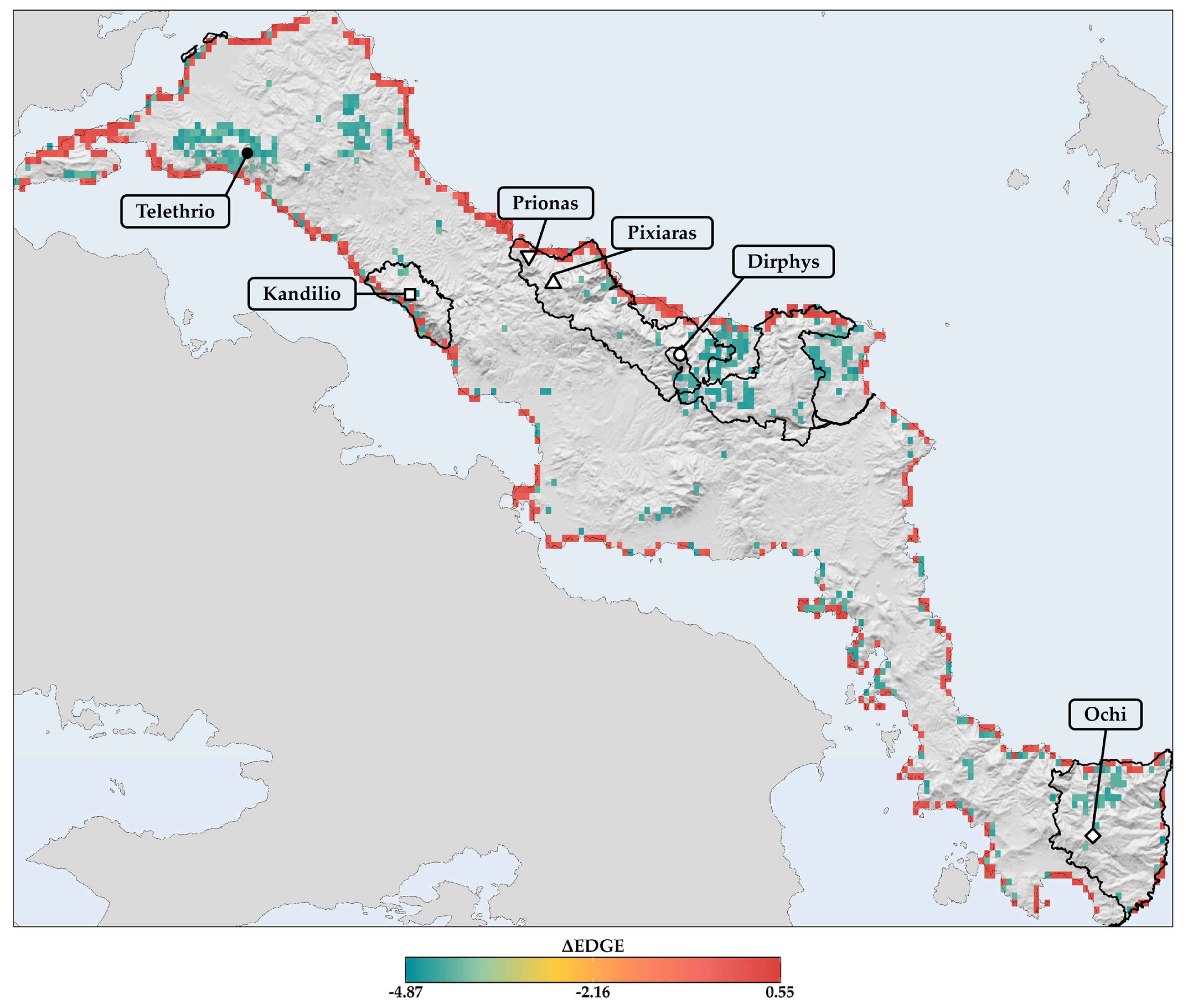
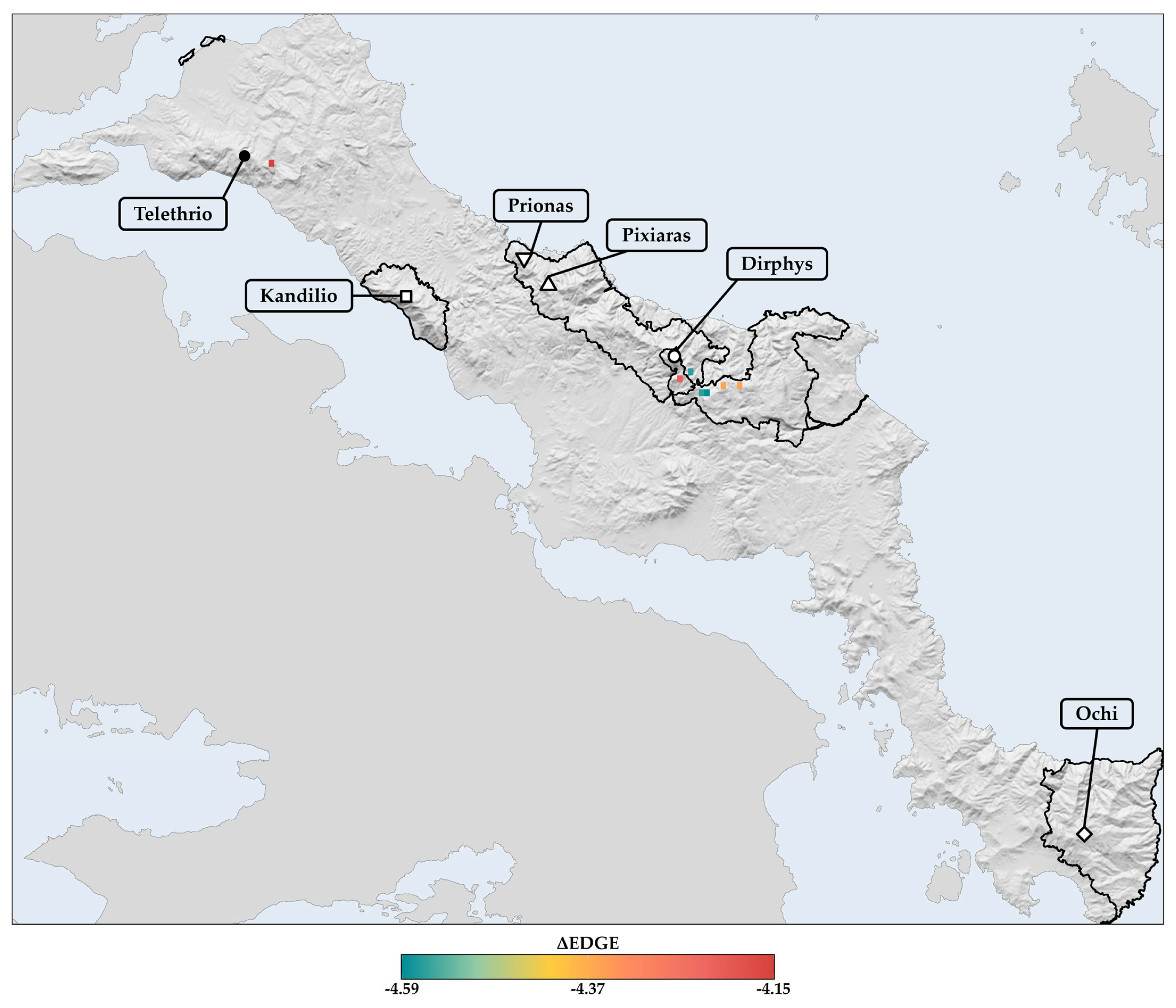
3.8. Estimation of the Evolutionarily Distinct and Globally Endangered (EDGE) Index—Current and Future EDGE Spatial Patterns
4. Discussion
4.1. Species-Specific Responses to Global Change Drivers
Comparative Analysis with Other Island Systems
4.2. Extinction Risk Assessment
4.2.1. Projected Changes in Threat Categories
4.2.2. Comparative Analyses with Other Island Endemics and Island Systems
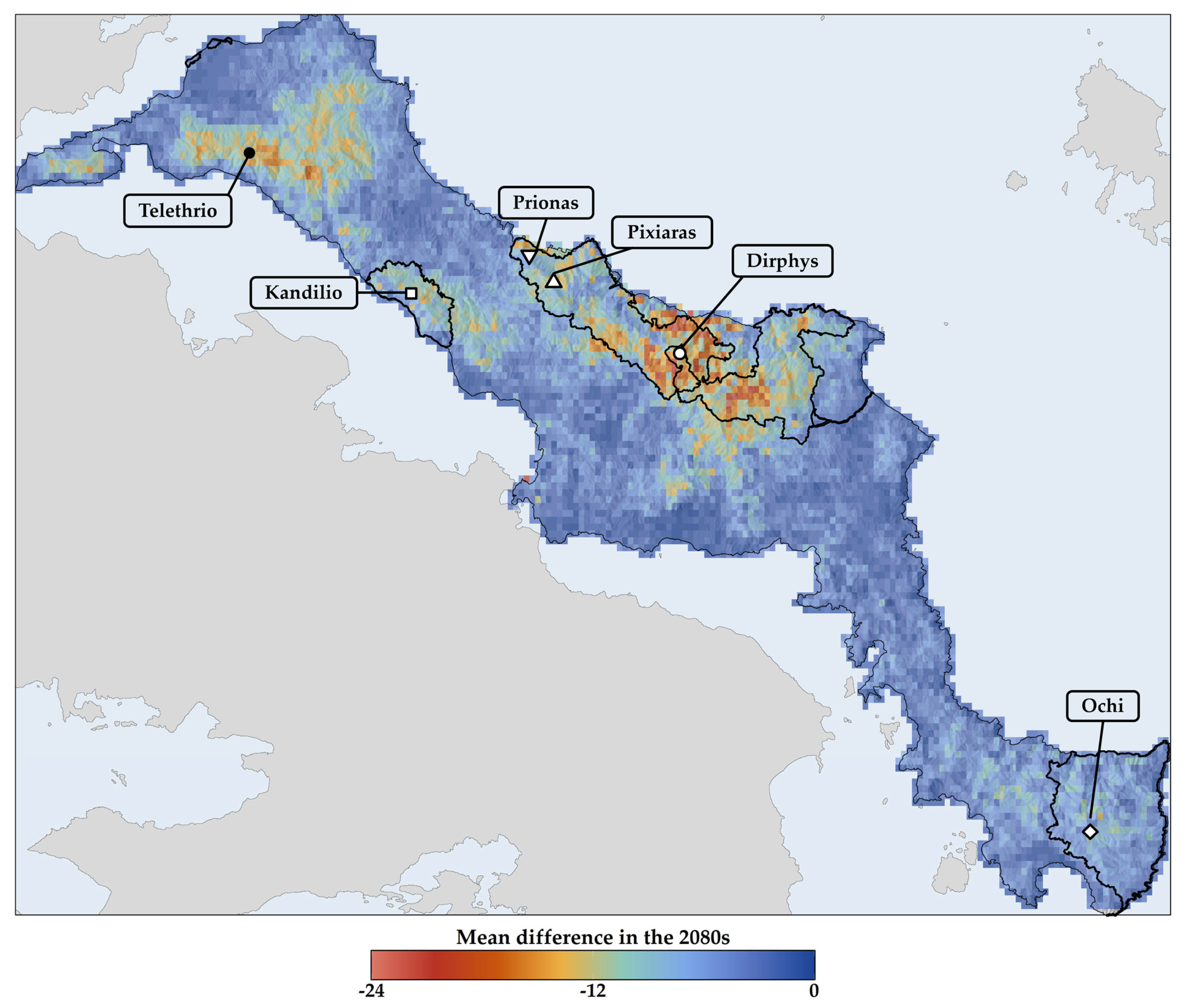
4.3. Shifts in Biodiversity Hotspots
4.3.1. Projected Spatial and Altitudinal Redistributions
4.3.2. Evolutionary Implications and Future Refugia
4.3.3. Conservation Priorities
4.4. Effectiveness of Protected Areas and Conservation Gaps
- (a)
- Identify and prioritise areas projected to serve as future biodiversity hotspots for legal protection and conservation management, contributing to the EU’s target of protecting 30% of land and sea by 2030.
- (b)
- Develop iterative conservation plans that can accommodate shifts in species distributions and ecological requirements over time, ensuring the long-term effectiveness of protected areas.
- (c)
- Engage local communities and stakeholders in conservation planning efforts to ensure that socio-economic considerations are addressed, promoting the integration of biodiversity values and ecosystem services into local planning and development processes.
- (d)
- Strengthen monitoring programmes to track changes in species abundances and distributions, informing adaptive management strategies and contributing to the EU’s goal of improving knowledge, the science base, and technologies relating to biodiversity, relevant ecosystem services and natural capital accounting.
4.5. Management Implications
4.6. Limitations and Uncertainties
5. Conclusions
- Incorporating biotic interactions and species’ adaptive capacities into modelling efforts
- Investigating the possibility for rapid evolution in island endemic plants in response to climate change
- Testing adaptive, long-term conservation strategies, such as flexible protected area designs and adaptive management approaches
- Assessing the implications of changing endemic plant distributions on ecosystem services
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AUC | Area under the Curve |
| CBI | Continuous Boyce Index |
| CGIAR | Consultative Group on International Agricultural Research |
| CMIP6 | Coupled Model Intercomparison Project Phase |
| CORINE | Coordination of information on the environment |
| CR | Critically Endangered |
| CWE | Corrected-weighted endemism |
| ED | Evolutionary Distinct |
| EDGE | Evolutionary Distinct and Globally Endangered |
| EHSA | Emergent Hot Spot Analysis |
| EN | Endangered |
| EU | European Union |
| EX | Extinct |
| GCMs | Global Circulation Models |
| GE | Globally Endangered |
| IUCN | International Union for the Conservation of Nature |
| LC | Least Concern |
| NT | Near Threatened |
| PD | Phylogenetic Diversity |
| PE | Phylogenetic Endemism |
| RCPs | Representative Concentration Pathways |
| SBI | Smoothed Boyce Index |
| SR | Species Richness |
| SSPs | Shared Socioeconomic Pathways |
| TSS | True Skill Statistic |
| VU | Vulnerable |
| UN | United Nations |
| ΔEDGE | Delta Evolutionary Distinct and Globally Endangered |
References
- Watson, R.; Baste, I.; Larigauderie, A.; Leadley, P.; Pascual, U.; Baptiste, B.; Demissew, S.; Dziba, L.; Erpul, G.; Fazel, A. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science—Policy Platform on Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2019; pp. 22–47. [Google Scholar]
- Moreira, H.; Kuipers, K.J.J.; Posthuma, L.; Zijp, M.C.; Hauck, M.; Huijbregts, M.A.J.; Schipper, A.M. Threats of Land Use to the Global Diversity of Vascular Plants. Divers. Distrib. 2023, 29, 688–697. [Google Scholar] [CrossRef]
- Oliver, T.H.; Morecroft, M.D. Interactions between Climate Change and Land Use Change on Biodiversity: Attribution Problems, Risks, and Opportunities. Wiley Interdiscip. Rev. Clim. Change 2014, 5, 317–335. [Google Scholar] [CrossRef]
- Newbold, T.; Oppenheimer, P.; Etard, A.; Williams, J.J. Tropical and Mediterranean Biodiversity Is Disproportionately Sensitive to Land-Use and Climate Change. Nat. Ecol. Evol. 2020, 4, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Poniatowski, D.; Beckmann, C.; Löffler, F.; Münsch, T.; Helbing, F.; Samways, M.J.; Fartmann, T. Relative Impacts of Land-Use and Climate Change on Grasshopper Range Shifts Have Changed over Time. Glob. Ecol. Biogeogr. 2020, 29, 2190–2202. [Google Scholar] [CrossRef]
- Kehoe, R.; Frago, E.; Sanders, D. Cascading Extinctions as a Hidden Driver of Insect Decline. Ecol. Entomol. 2021, 46, 743–756. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; Kreft, H.; Irl, S.D.H.; Norder, S.; Ah-Peng, C.; Borges, P.A.V.; Burns, K.C.; de Nascimento, L.; Meyer, J.-Y.; Montes, E. Scientists’ Warning—The Outstanding Biodiversity of Islands Is in Peril. Glob. Ecol. Conserv. 2021, 31, e01847. [Google Scholar] [CrossRef]
- Médail, F. The Specific Vulnerability of Plant Biodiversity and Vegetation on Mediterranean Islands in the Face of Global Change. Reg. Environ. Change 2017, 17, 1775–1790. [Google Scholar] [CrossRef]
- Lazoglou, G.; Papadopoulos-Zachos, A.; Georgiades, P.; Zittis, G.; Velikou, K.; Manios, E.M.; Anagnostopoulou, C. Identification of Climate Change Hotspots in the Mediterranean. Sci. Rep. 2024, 14, 29817. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean: Insights for Conservation; Oxford University Press: New York, NY, USA, 2020. [Google Scholar]
- Urban, M.C. Climate Change Extinctions. Science 2024, 386, 1123–1128. [Google Scholar] [CrossRef]
- Wiens, J.J.; Zelinka, J. How Many Species Will Earth Lose to Climate Change? Glob. Change Biol. 2024, 30, e17125. [Google Scholar] [CrossRef]
- Asamoah, E.F.; Beaumont, L.J.; Maina, J.M. Climate and Land-Use Changes Reduce the Benefits of Terrestrial Protected Areas. Nat. Clim. Change 2021, 11, 1105–1110. [Google Scholar] [CrossRef]
- Cao, Y.; Tseng, T.-H.; Wang, F.; Jacobson, A.; Yu, L.; Zhao, J.; Carver, S.; Locke, H.; Zhao, Z.; Yang, R. Potential Wilderness Loss Could Undermine the Post-2020 Global Biodiversity Framework. Biol. Conserv. 2022, 275, 109753. [Google Scholar] [CrossRef]
- Cañadas, E.M.; Fenu, G.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within Hotspots: Endemic Plant Richness, Environmental Drivers, and Implications for Conservation. Biol. Conserv. 2014, 170, 282–291. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial Refugia Influence Plant Diversity Patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Hammoud, C.; Kougioumoutzis, K.; Rijsdijk, K.F.; Simaiakis, S.M.; Norder, S.J.; Foufopoulos, J.; Georgopoulou, E.; Van Loon, E.E. Past Connections with the Mainland Structure Patterns of Insular Species Richness in a Continental-shelf Archipelago (Aegean Sea, Greece). Ecol. Evol. 2021, 11, 5441–5458. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Kallimanis, A.; Strid, A.; Dimopoulos, P. Plant Endemism Centres and Biodiversity Hotspots in Greece. Biology 2021, 10, 72. [Google Scholar] [CrossRef]
- Panitsa, M.; Kagiampaki, A.; Kougioumoutzis, K. Plant Diversity and Biogeography of the Aegean Archipelago: A New Synthesis. In Biogeography and Biodiversity of the Aegean: In honour of Prof. Moysis Mylonas; Sfenthourakis, S., Pafilis, P., Parmakelis, A., Poulakakis, N., Triantis, K.A., Eds.; Broken Hill Publishers Ltd.: Nicosia, Cyprus, 2018; pp. 223–244. ISBN 9789925563784. [Google Scholar]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Strid, A.; Dimopoulos, P. Extinction Risk Assessment of the Greek Endemic Flora. Biology 2021, 10, 195. [Google Scholar] [CrossRef]
- Benavides, E.; Sadler, J.; Graham, L.; Matthews, T.J. Species Distribution Models and Island Biogeography: Challenges and Prospects. Glob. Ecol. Conserv. 2024, 51, e02943. [Google Scholar] [CrossRef]
- Hanz, D.M.; Cutts, V.; Barajas-Barbosa, M.P.; Algar, A.; Beierkuhnlein, C.; Collart, F.; Fernández-Palacios, J.M.; Field, R.; Karger, D.N.; Kienle, D.R.; et al. Effects of Climate Change on the Distribution of Plant Species and Plant Functional Strategies on the Canary Islands. Divers. Distrib. 2023, 29, 1157–1171. [Google Scholar] [CrossRef]
- Fortini, L.B.; Price, J.; Jacobi, J.; Vorsino, A.; Burgett, J.; Brinck, K.W.; Amidon, F.; Miller, S.; Koob, G.; Paxton, E.H. A Landscape-Based Assessment of Climate Change Vulnerability for All Native Hawaiian Plants; University of Hawaii: Honolulu, HI, USA, 2013. [Google Scholar]
- Pouteau, R.; Birnbaum, P. Island Biodiversity Hotspots Are Getting Hotter: Vulnerability of Tree Species to Climate Change in New Caledonia. Biol. Conserv. 2016, 201, 111–119. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Valli, A.T.; Georgopoulou, E.; Simaiakis, S.M.; Triantis, K.A.; Trigas, P. Network Biogeography of a Complex Island System: The Aegean Archipelago Revisited. J. Biogeogr. 2017, 44, 651–660. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Papanikolaou, A.; Kokkoris, I.P.; Strid, A.; Dimopoulos, P.; Panitsa, M. Climate Change Impacts and Extinction Risk Assessment of Nepeta Representatives (Lamiaceae) in Greece. Sustainability 2022, 14, 4269. [Google Scholar] [CrossRef]
- Pires, M.B.; Kougioumoutzis, K.; Norder, S.; Dimopoulos, P.; Strid, A.; Panitsa, M. The Future of Plant Diversity within a Mediterranean Endemism Centre: Modelling the Synergistic Effects of Climate and Land-Use Change in Peloponnese, Greece. Sci. Total Environ. 2024, 947, 174622. [Google Scholar]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Trigas, P.; Strid, A.; Dimopoulos, P. Plant Diversity Patterns and Conservation Implications under Climate-Change Scenarios in the Mediterranean: The Case of Crete (Aegean, Greece). Diversity 2020, 12, 270. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kotsakiozi, P.; Stathi, E.; Trigas, P.; Parmakelis, A. Conservation Genetics of Four Critically Endangered Greek Endemic Plants: A Preliminary Assessment. Diversity 2021, 13, 152. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Tsakiri, M.; Kokkoris, I.P.; Trigas, P.; Iatrou, G.; Lamari, F.N.; Tzanoudakis, D.; Koumoutsou, E.; Dimopoulos, P.; Strid, A. Assessing the Vulnerability of Medicinal and Aromatic Plants to Climate and Land-Use Changes in a Mediterranean Biodiversity Hotspot. Land 2024, 13, 133. [Google Scholar] [CrossRef]
- Trigas, P.; Iatrou, G. The Local Endemic Flora of Evvia (W Aegean, Greece). Willdenowia 2006, 36, 257. [Google Scholar] [CrossRef]
- Trigas, P.; Iatrou, G.; Panitsa, M. Vascular Plant Species Diversity, Biogeography and Vulnerability in the Aegean Islands as Exemplified by Evvia Island (W Aegean, Greece). Fresenius Environ. Bull. 2008, 17, 48–57. [Google Scholar]
- Santos, M.J.; Smith, A.B.; Dekker, S.C.; Eppinga, M.B.; Leitão, P.J.; Moreno-Mateos, D.; Morueta-Holme, N.; Ruggeri, M. The Role of Land Use and Land Cover Change in Climate Change Vulnerability Assessments of Biodiversity: A Systematic Review. Landsc. Ecol. 2021, 36, 3367–3382. [Google Scholar] [CrossRef]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular Plants of Greece: An Annotated Checklist. Englera 2013, 1–372. [Google Scholar]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular Plants of Greece: An Annotated Checklist. Supplement. Willdenowia 2016, 46, 301–347. [Google Scholar] [CrossRef]
- Dauby, G.; Stévart, T.; Droissart, V.; Cosiaux, A.; Deblauwe, V.; Simo-Droissart, M.; Sosef, M.S.M.; Lowry, P.P.; Schatz, G.E.; Gereau, R.E.; et al. ConR: An R Package to Assist Large-Scale Multispecies Preliminary Conservation Assessments Using Distribution Data. Ecol. Evol. 2017, 7, 11292–11303. [Google Scholar] [CrossRef] [PubMed]
- Burgman, M.A.; Fox, J.C. Bias in Species Range Estimates from Minimum Convex Polygons: Implications for Conservation and Options for Improved Planning; Cambridge University Press: Cambridge, UK, 2003; Volume 6, pp. 19–28. [Google Scholar]
- Jetz, W.; Sekercioglu, C.H.; Watson, J.E. Ecological Correlates and Conservation Implications of Overestimating Species Geographic Ranges. Conserv. Biol. 2008, 22, 110–119. [Google Scholar] [CrossRef]
- Meyer, L.; Diniz-Filho, J.A.F.; Lohmann, L.G. A Comparison of Hull Methods for Estimating Species Ranges and Richness Maps. Plant Ecol. Divers. 2017, 10, 389–401. [Google Scholar] [CrossRef]
- Zizka, A.; Silvestro, D.; Andermann, T.; Azevedo, J.; Duarte Ritter, C.; Edler, D.; Farooq, H.; Herdean, A.; Ariza, M.; Scharn, R. CoordinateCleaner: Standardized Cleaning of Occurrence Records from Biological Collection Databases. Methods Ecol. Evol. 2019, 10, 744–751. [Google Scholar] [CrossRef]
- Smith, A.B. enmSdm: Tools for Modeling Species Niches and Distributions. R Package Version 0.5.1.5. 2020. Available online: http://github.com/adamlilith/enmSdm (accessed on 6 March 2025).
- Lamboley, Q.; Fourcade, Y. No Optimal Spatial Filtering Distance for Mitigating Sampling Bias in Ecological Niche Models. J. Biogeogr. 2024, 51, 1783–1794. [Google Scholar] [CrossRef]
- Ten Caten, C.; Dallas, T. Thinning Occurrence Points Does Not Improve Species Distribution Model Performance. Ecosphere 2023, 14, e4703. [Google Scholar] [CrossRef]
- Baker, D.J.; Maclean, I.M.; Gaston, K.J. Effective Strategies for Correcting Spatial Sampling Bias in Species Distribution Models without Independent Test Data. Divers. Distrib. 2024, 30, e13802. [Google Scholar] [CrossRef]
- Moua, Y.; Roux, E.; Seyler, F.; Briolant, S. Correcting the Effect of Sampling Bias in Species Distribution Modeling—A New Method in the Case of a Low Number of Presence Data. Ecol. Inform. 2020, 57, 101086. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, X.; Yi, J.; Wang, Y. Bias Correction in Species Distribution Models Based on Geographic and Environmental Characteristics. Ecol. Inform. 2024, 81, 102604. [Google Scholar] [CrossRef]
- Sandel, B.; Merow, C.; Serra-Diaz, J.M.; Svenning, J. Disequilibrium in Plant Distributions: Challenges and Approaches for Species Distribution Models. J. Ecol. 2025, 113, 782–794. [Google Scholar] [CrossRef]
- Steen, V.A.; Tingley, M.W.; Paton, P.W.; Elphick, C.S. Spatial Thinning and Class Balancing: Key Choices Lead to Variation in the Performance of Species Distribution Models with Citizen Science Data. Methods Ecol. Evol. 2021, 12, 216–226. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R Package for Spatial Thinning of Species Occurrence Records for Use in Ecological Niche Models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Robertson, M.P.; Visser, V.; Hui, C. Biogeo: An R Package for Assessing and Improving Data Quality of Occurrence Record Datasets. Ecography 2016, 39, 394–401. [Google Scholar] [CrossRef]
- Moudrý, V.; Bazzichetto, M.; Remelgado, R.; Devillers, R.; Lenoir, J.; Mateo, R.G.; Lembrechts, J.J.; Sillero, N.; Lecours, V.; Cord, A.F. Optimising Occurrence Data in Species Distribution Models: Sample Size, Positional Uncertainty, and Sampling Bias Matter. Ecography 2024, 2024, e07294. [Google Scholar] [CrossRef]
- Soley-Guardia, M.; Alvarado-Serrano, D.F.; Anderson, R.P. Top Ten Hazards to Avoid When Modeling Species Distributions: A Didactic Guide of Assumptions, Problems, and Recommendations. Ecography 2024, 2024, e06852. [Google Scholar] [CrossRef]
- Araújo, M.B.; Anderson, R.P.; Barbosa, A.M.; Beale, C.M.; Dormann, C.F.; Early, R.; Garcia, R.A.; Guisan, A.; Maiorano, L.; Naimi, B.; et al. Standards for Distribution Models in Biodiversity Assessments. Sci. Adv. 2019, 5, eaat4858. [Google Scholar] [CrossRef]
- Sillero, N.; Arenas-Castro, S.; Enriquez-Urzelai, U.; Vale, C.G.; Sousa-Guedes, D.; Martínez-Freiría, F.; Real, R.; Barbosa, A.M. Want to Model a Species Niche? A Step-by-Step Guideline on Correlative Ecological Niche Modelling. Ecol. Model. 2021, 456, 109671. [Google Scholar] [CrossRef]
- Sillero, N.; Barbosa, A.M. Common Mistakes in Ecological Niche Models. Int. J. Geogr. Inf. Sci. 2021, 35, 213–226. [Google Scholar] [CrossRef]
- van Proosdij, A.S.J.; Sosef, M.S.M.; Wieringa, J.J.; Raes, N. Minimum Required Number of Specimen Records to Develop Accurate Species Distribution Models. Ecography 2016, 39, 542–552. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.; Li, J.; Peterson, A.T.; Graham, C.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of Sample Size on the Performance of Species Distribution Models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The Effect of Sample Size and Species Characteristics on Performance of Different Species Distribution Modeling Methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Erickson, K.D.; Smith, A.B. Modeling the Rarest of the Rare: A Comparison between Multi-species Distribution Models, Ensembles of Small Models, and Single-species Models at Extremely Low Sample Sizes. Ecography 2023, 2023, e06500. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Title, P.O.; Bemmels, J.B. ENVIREM: An Expanded Set of Bioclimatic and Topographic Variables Increases Flexibility and Improves Performance of Ecological Niche Modeling. Ecography 2018, 41, 291–307. [Google Scholar] [CrossRef]
- Jarvis, A.; Reuter, H.I.; Nelson, A.; Guevara, E. Hole-Filled SRTM for the Globe Version 4. CGIAR-CSI SRTM 90m Database. 2008. Available online: http://srtm.csi.cgiar.org (accessed on 6 March 2025).
- Hijmans, R.; Philipps, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling. R Package Version 1.1-4. 2017. Available online: https://CRAN.R-project.org/package=dismo (accessed on 6 March 2025).
- Marchi, M.; Castellanos-Acuña, D.; Hamann, A.; Wang, T.; Ray, D.; Menzel, A. ClimateEU, Scale-Free Climate Normals, Historical Time Series, and Future Projections for Europe. Sci. Data 2020, 7, 428. [Google Scholar] [CrossRef]
- Hamann, A.; Wang, T.; Spittlehouse, D.L.; Murdock, T.Q. A Comprehensive, High-Resolution Database of Historical and Projected Climate Surfaces for Western North America. Bull. Am. Meteorol. Soc. 2013, 94, 1307–1309. [Google Scholar] [CrossRef]
- Wang, T.; Hamann, A.; Spittlehouse, D.L.; Murdock, T.Q. ClimateWNA—High-Resolution Spatial Climate Data for Western North America. J. Appl. Meteorol. Climatol. 2012, 51, 16–29. [Google Scholar] [CrossRef]
- Chen, G.; Li, X.; Liu, X. Global Land Projection Based on Plant Functional Types with a 1-Km Resolution under Socio-Climatic Scenarios. Sci. Data 2022, 9, 125. [Google Scholar] [CrossRef]
- Hengl, T.; de Jesus, J.M.; Heuvelink, G.B.M.; Gonzalez, M.R.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global Gridded Soil Information Based on Machine Learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef]
- Ni, M.; Vellend, M. Soil Properties Constrain Predicted Poleward Migration of Plants under Climate Change. New Phytol. 2024, 241, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, F.O.G.; Zuquim, G.; Tuomisto, H.; Moulatlet, G.M.; Balslev, H.; Costa, F.R.C. Beyond Climate Control on Species Range: The Importance of Soil Data to Predict Distribution of Amazonian Plant Species. J. Biogeogr. 2018, 45, 190–200. [Google Scholar] [CrossRef]
- Roe, N.A.; Ducey, M.J.; Lee, T.D.; Fraser, O.L.; Colter, R.A.; Hallett, R.A. Soil Chemical Variables Improve Models of Understorey Plant Species Distributions. J. Biogeogr. 2022, 49, 753–766. [Google Scholar] [CrossRef]
- Gardner, A.S.; Maclean, I.M.; Gaston, K.J. Climatic Predictors of Species Distributions Neglect Biophysiologically Meaningful Variables. Divers. Distrib. 2019, 25, 1318–1333. [Google Scholar] [CrossRef]
- Hijmans, R. Terra: Spatial Data Analysis. R Package Version 1.7-46. 2023. Available online: https://CRAN.R-project.org/package=terra (accessed on 6 March 2025).
- Evans, J.S. spatialEco—R Package Version 1.2-0. 2019. Available online: https://github.com/jeffreyevans/spatialEco (accessed on 6 March 2025).
- McSweeney, C.F.; Jones, R.G.; Lee, R.W.; Rowell, D.P. Selecting CMIP5 GCMs for Downscaling over Multiple Regions. Clim. Dyn. 2015, 44, 3237–3260. [Google Scholar] [CrossRef]
- Parding, K.M.; Dobler, A.; McSweeney, C.F.; Landgren, O.A.; Benestad, R.; Erlandsen, H.B.; Mezghani, A.; Gregow, H.; Räty, O.; Viktor, E. GCMeval—An Interactive Tool for Evaluation and Selection of Climate Model Ensembles. Clim. Serv. 2020, 18, 100167. [Google Scholar] [CrossRef]
- Esser, L.F.; Bailly, D.; Lima, M.R.; Ré, R. chooseGCM: A Toolkit to Select General Circulation Models in R. Glob. Change Biol. 2025, 31, e70008. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, F.; Tseng, T.-H.; Carver, S.; Chen, X.; Zhao, J.; Yu, L.; Li, F.; Zhao, Z.; Yang, R. Identifying Ecosystem Service Value and Potential Loss of Wilderness Areas in China to Support Post-2020 Global Biodiversity Conservation. Sci. Total Environ. 2022, 846, 157348. [Google Scholar] [CrossRef]
- Austin, M.P.; Van Niel, K.P. Improving Species Distribution Models for Climate Change Studies: Variable Selection and Scale. J. Biogeogr. 2011, 38, 1–8. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting Species Distribution: Offering More than Simple Habitat Models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Mod, H.K.; Scherrer, D.; Luoto, M.; Guisan, A. What We Use Is Not What We Know: Environmental Predictors in Plant Distribution Models. J. Veg. Sci. 2016, 27, 1308–1322. [Google Scholar] [CrossRef]
- Scherrer, D.; Massy, S.; Meier, S.; Vittoz, P.; Guisan, A. Assessing and Predicting Shifts in Mountain Forest Composition across 25 Years of Climate Change. Divers. Distrib. 2017, 23, 517–528. [Google Scholar] [CrossRef]
- Dubuis, A.; Giovanettina, S.; Pellissier, L.; Pottier, J.; Vittoz, P.; Guisan, A. Improving the Prediction of Plant Species Distribution and Community Composition by Adding Edaphic to Topo-Climatic Variables. J. Veg. Sci. 2013, 24, 593–606. [Google Scholar] [CrossRef]
- Körner, C.; Kèorner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Lazarina, M.; Kallimanis, A.S.; Dimopoulos, P.; Psaralexi, M.; Michailidou, D.E.; Sgardelis, S.P. Patterns and Drivers of Species Richness and Turnover of Neo-Endemic and Palaeo-Endemic Vascular Plants in a Mediterranean Hotspot: The Case of Crete, Greece. J. Biol. Res. 2019, 26, 12. [Google Scholar] [CrossRef]
- Panitsa, M.; Trigas, P.; Iatrou, G.; Sfenthourakis, S. Factors Affecting Plant Species Richness and Endemism on Land-Bridge Islands—An Example from the East Aegean Archipelago. Acta Oecologica 2010, 36, 431–437. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Tiniakou, A. Ecological Factors Driving Plant Species Diversity in the South Aegean Volcanic Arc and Other Central Aegean Islands. Plant Ecol. Divers. 2015, 8, 173–186. [Google Scholar] [CrossRef]
- Kagiampaki, A.; Triantis, K.; Vardinoyannis, K.; Mylonas, M. Factors Affecting Plant Species Richness and Endemism in the South Aegean (Greece). J. Biol. Res. 2011, 16, 282–295. [Google Scholar]
- Kallimanis, A.S.; Bergmeier, E.; Panitsa, M.; Georghiou, K.; Delipetrou, P.; Dimopoulos, P. Biogeographical Determinants for Total and Endemic Species Richness in a Continental Archipelago. Biodivers. Conserv. 2010, 19, 1225–1235. [Google Scholar] [CrossRef]
- Trigas, P.; Panitsa, M.; Tsiftsis, S. Elevational Gradient of Vascular Plant Species Richness and Endemism in Crete—The Effect of Post-Isolation Mountain Uplift on a Continental Island System. PLoS ONE 2013, 8, e59425. [Google Scholar] [CrossRef]
- Merkenschlager, C.; Bangelesa, F.; Paeth, H.; Hertig, E. Blessing and Curse of Bioclimatic Variables: A Comparison of Different Calculation Schemes and Datasets for Species Distribution Modeling within the Extended Mediterranean Area. Ecol. Evol. 2023, 13, e10553. [Google Scholar] [CrossRef]
- Booth, T.H. Checking Bioclimatic Variables That Combine Temperature and Precipitation Data before Their Use in Species Distribution Models. Austral Ecol. 2022, 47, 1506–1514. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A Review of Methods to Deal with It and a Simulation Study Evaluating Their Performance. Ecography 2013, 36, 027–046. [Google Scholar] [CrossRef]
- Mendes, P.; Velazco, S.J.E.; de Andrade, A.F.A.; Marco, P.D. Dealing with Overprediction in Species Distribution Models: How Adding Distance Constraints Can Improve Model Accuracy. Ecol. Model. 2020, 431, 109180. [Google Scholar] [CrossRef]
- Benito, B. Collinear: R Package for Seamless Multicollinearity Management 2023. R Package Version 1.1.1. Available online: https://doi.org/10.1016/j.ecolmodel.2020.109180 (accessed on 6 March 2025).
- Meyer, H.; Reudenbach, C.; Hengl, T.; Katurji, M.; Nauss, T. Improving Performance of Spatio-Temporal Machine Learning Models Using Forward Feature Selection and Target-Oriented Validation. Environ. Model. Softw. 2018, 101, 1–9. [Google Scholar] [CrossRef]
- Meyer, H.; Reudenbach, C.; Wöllauer, S.; Nauss, T. Importance of Spatial Predictor Variable Selection in Machine Learning Applications–Moving from Data Reproduction to Spatial Prediction. Ecol. Model. 2019, 411, 108815. [Google Scholar] [CrossRef]
- Schratz, P.; Muenchow, J.; Iturritxa, E.; Richter, J.; Brenning, A. Hyperparameter Tuning and Performance Assessment of Statistical and Machine-Learning Algorithms Using Spatial Data. Ecol. Model. 2019, 406, 109–120. [Google Scholar] [CrossRef]
- Meyer, H.; Ludwig, M.; Milà, C.; Linnenbrink, J.; Schumacher, F. The CAST Package for Training and Assessment of Spatial Prediction Models in R. arXiv 2024, arXiv:2404.06978. [Google Scholar]
- Meyer, H.; Milà, C.; Ludwig, M.; Linnenbrink, J.; Schumacher, F. CAST: “caret” Applications for Spatial-Temporal Models. 2024. R Package Version 1.0.2. Available online: https://CRAN.R-project.org/package=CAST (accessed on 6 March 2025).
- Linnenbrink, J.; Milà, C.; Ludwig, M.; Meyer, H. kNNDM: K-Fold Nearest Neighbour Distance Matching Cross-Validation for Map Accuracy Estimation. EGUsphere 2023, 2023, 1–16. [Google Scholar] [CrossRef]
- Mila, C.; Mateu, J.; Pebesma, E.; Meyer, H. Nearest Neighbour Distance Matching Leave-One-Out Cross-Validation for Map Validation. Methods Ecol. Evol. 2022, 13, 1304–1316. [Google Scholar] [CrossRef]
- Ploton, P.; Mortier, F.; Réjou-Méchain, M.; Barbier, N.; Picard, N.; Rossi, V.; Dormann, C.; Cornu, G.; Viennois, G.; Bayol, N. Spatial Validation Reveals Poor Predictive Performance of Large-Scale Ecological Mapping Models. Nat. Commun. 2020, 11, 4540. [Google Scholar] [CrossRef]
- Lomba, A.; Pellissier, L.; Randin, C.; Vicente, J.; Moreira, F.; Honrado, J.; Guisan, A. Overcoming the Rare Species Modelling Paradox: A Novel Hierarchical Framework Applied to an Iberian Endemic Plant. Biol. Conserv. 2010, 143, 2647–2657. [Google Scholar] [CrossRef]
- Mondanaro, A.; Di Febbraro, M.; Castiglione, S.; Melchionna, M.; Serio, C.; Girardi, G.; Belfiore, A.M.; Raia, P. ENphylo: A New Method to Model the Distribution of Extremely Rare Species. Methods Ecol. Evol. 2023, 14, 911–922. [Google Scholar] [CrossRef]
- Mondanaro, A.; Di Febbraro, M.; Castiglione, S.; Belfiore, A.M.; Girardi, G.; Melchionna, M.; Serio, C.; Esposito, A.; Raia, P. Modelling Reveals the Effect of Climate and Land Use Change on Madagascar’s Chameleons Fauna. Commun. Biol. 2024, 7, 889. [Google Scholar] [CrossRef]
- Breiner, F.T.; Guisan, A.; Bergamini, A.; Nobis, M.P. Overcoming Limitations of Modelling Rare Species by Using Ensembles of Small Models. Methods Ecol. Evol. 2015, 6, 1210–1218. [Google Scholar] [CrossRef]
- Breiner, F.T.; Nobis, M.P.; Bergamini, A.; Guisan, A. Optimizing Ensembles of Small Models for Predicting the Distribution of Species with Few Occurrences. Methods Ecol. Evol. 2018, 9, 802–808. [Google Scholar] [CrossRef]
- Breiner, F.T.; Guisan, A.; Nobis, M.P.; Bergamini, A. Including Environmental Niche Information to Improve IUCN Red List Assessments. Divers. Distrib. 2017, 23, 484–495. [Google Scholar] [CrossRef]
- Broennimann, O.; Di Cola, V.; Guisan, A. Ecospat: Spatial Ecology Miscellaneous Methods. R Package Version 3.2. 2021. Available online: https://CRAN.R-project.org/package=ecospat (accessed on 6 March 2025).
- Valavi, R.; Elith, J.; Lahoz-Monfort, J.J.; Guillera-Arroita, G. Modelling Species Presence-only Data with Random Forests. Ecography 2021, 44, 1731–1742. [Google Scholar] [CrossRef]
- Valavi, R.; Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J. Predictive Performance of Presence-only Species Distribution Models: A Benchmark Study with Reproducible Code. Ecol. Monogr. 2022, 92, e01486. [Google Scholar] [CrossRef]
- Curth, A.; Jeffares, A.; van der Schaar, M. Why Do Random Forests Work? Understanding Tree Ensembles as Self-Regularizing Adaptive Smoothers. arXiv 2024, arXiv:2402.01502. [Google Scholar]
- Jimenez-Valverde, A. Prevalence Affects the Evaluation of Discrimination Capacity in Presence-Absence Species Distribution Models. Biodivers. Conserv. 2021, 30, 1331–1340. [Google Scholar] [CrossRef]
- Velazco, S.J.E.; Rose, M.B.; de Andrade, A.F.A.; Minoli, I.; Franklin, J. Flexsdm: An r Package for Supporting a Comprehensive and Flexible Species Distribution Modelling Workflow. Methods Ecol. Evol. 2022, 13, 1661–1669. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting Pseudo-Absences for Species Distribution Models: How, Where and How Many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting Thresholds for the Prediction of Species Occurrence with Presence-Only Data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Broussin, J.; Mouchet, M.; Goberville, E. Generating Pseudo-Absences in the Ecological Space Improves the Biological Relevance of Response Curves in Species Distribution Models. Ecol. Model. 2024, 498, 110865. [Google Scholar] [CrossRef]
- Inman, R.; Franklin, J.; Esque, T.; Nussear, K. Comparing Sample Bias Correction Methods for Species Distribution Modeling Using Virtual Species. Ecosphere 2021, 12, e03422. [Google Scholar] [CrossRef]
- Dubos, N.; Préau, C.; Lenormand, M.; Papuga, G.; Monsarrat, S.; Denelle, P.; Le Louarn, M.; Heremans, S.; May, R.; Roche, P. Assessing the Effect of Sample Bias Correction in Species Distribution Models. Ecol. Indic. 2022, 145, 109487. [Google Scholar] [CrossRef]
- Roberts, D.R.; Bahn, V.; Ciuti, S.; Boyce, M.S.; Elith, J.; Guillera-Arroita, G.; Hauenstein, S.; Lahoz-Monfort, J.J.; Schröder, B.; Thuiller, W.; et al. Cross-Validation Strategies for Data with Temporal, Spatial, Hierarchical, or Phylogenetic Structure. Ecography 2017, 4, 913–929. [Google Scholar] [CrossRef]
- Santini, L.; Benítez-López, A.; Maiorano, L.; Čengić, M.; Huijbregts, M.A.J. Assessing the Reliability of Species Distribution Projections in Climate Change Research. Divers. Distrib. 2021, 27, 1035–1050. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F. Biomod2: Ensemble Platform for Species Distribution Modeling 2016. R Package Version 4.2.4. Available online: https://CRAN.R-project.org/package=biomod2 (accessed on 6 March 2025).
- Collart, F.; Guisan, A. Small to Train, Small to Test: Dealing with Low Sample Size in Model Evaluation. Ecol. Inform. 2023, 75, 102106. [Google Scholar] [CrossRef]
- Collart, F.; Hedenäs, L.; Broennimann, O.; Guisan, A.; Vanderpoorten, A. Intraspecific Differentiation: Implications for Niche and Distribution Modelling. J. Biogeogr. 2021, 48, 415–426. [Google Scholar] [CrossRef]
- Hallman, T.A.; Robinson, W.D. Deciphering Ecology from Statistical Artefacts: Competing Influence of Sample Size, Prevalence and Habitat Specialization on Species Distribution Models and How Small Evaluation Datasets Can Inflate Metrics of Performance. Divers. Distrib. 2020, 26, 315–328. [Google Scholar] [CrossRef]
- Liu, C.; Newell, G.; White, M.; Machunter, J. Improving the Estimation of the Boyce Index Using Statistical Smoothing Methods for Evaluating Species Distribution Models with Presence-only Data. Ecography 2025, 2025, e07218. [Google Scholar] [CrossRef]
- Raes, N.; ter Steege, H. A Null-Model for Significance Testing of Presence-Only Species Distribution Models. Ecography 2007, 30, 727–736. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the Accuracy of Species Distribution Models: Prevalence, Kappa and the True Skill Statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le Lay, G.; Helfer, V.; Randin, C.; Guisan, A. Evaluating the Ability of Habitat Suitability Models to Predict Species Presences. Ecol. Model. 2006, 199, 142–152. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A Review of Methods for the Assessment of Prediction Errors in Conservation Presence/ Absence Models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Sofaer, H.R.; Hoeting, J.A.; Jarnevich, C.S. The Area under the Precision-Recall Curve as a Performance Metric for Rare Binary Events. Methods Ecol. Evol. 2019, 10, 565–577. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Measuring and Comparing the Accuracy of Species Distribution Models with Presence-Absence Data. Ecography 2011, 34, 232–243. [Google Scholar] [CrossRef]
- Konowalik, K.; Nosol, A. Evaluation Metrics and Validation of Presence-Only Species Distribution Models Based on Distributional Maps with Varying Coverage. Sci. Rep. 2021, 11, 1482. [Google Scholar] [CrossRef]
- Arenas-Castro, S.; Regos, A.; Martins, I.; Honrado, J.; Alonso, J. Effects of Input Data Sources on Species Distribution Model Predictions across Species with Different Distributional Ranges. J. Biogeogr. 2022, 49, 1299–1312. [Google Scholar] [CrossRef]
- Hammer, B.; Frasco, M. Metrics: Evaluation Metrics for Machine Learning. R Package Version 0.1.4. 2018. Available online: https://CRAN.R-project.org/package=Metrics (accessed on 6 March 2025).
- Márcia Barbosa, A.; Real, R.; Muñoz, A.R.; Brown, J.A. New Measures for Assessing Model Equilibrium and Prediction Mismatch in Species Distribution Models. Divers. Distrib. 2013, 19, 1333–1338. [Google Scholar] [CrossRef]
- Schwarz, J.; Heider, D. GUESS: Projecting Machine Learning Scores to Well-Calibrated Probability Estimates for Clinical Decision-Making. Bioinformatics 2019, 35, 2458–2465. [Google Scholar] [CrossRef] [PubMed]
- Signorell, A.; Aho, K.; Anderegg, N.; Aragon, T.; Arppe, A.; Baddeley, A.; Bolker, B.; Caeiro, F.; Champely, S.; Chessel, D. DescTools: Tools for Descriptive Statistics. R Package Version 0.99-40. 2021. Available online: https://CRAN.R-project.org/package=DescTools (accessed on 6 March 2025).
- Engler, R.; Randin, C.F.; Thuiller, W.; Dullinger, S.; Zimmermann, N.E.; Araújo, M.B.; Pearman, P.B.; Lay, G.L.; Piedallu, C.; Albert, C.H.; et al. 21st Century Climate Change Threatens Mountain Flora Unequally across Europe. Glob. Change Biol. 2011, 17, 2330–2341. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Ecology, Biodiversity and Conservation; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0-521-70002-3. [Google Scholar]
- González-Irusta, J.M.; González-Porto, M.; Sarralde, R.; Arrese, B.; Almón, B.; Martín-Sosa, P. Comparing Species Distribution Models: A Case Study of Four Deep Sea Urchin Species. Hydrobiologia 2015, 745, 43–57. [Google Scholar] [CrossRef]
- Lahoz-Monfort, J.J.; Guillera-Arroita, G.; Wintle, B.A. Imperfect Detection Impacts the Performance of Species Distribution Models. Glob. Ecol. Biogeogr. 2014, 23, 504–515. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting Thresholds of Occurrence in the Prediction of Species Distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Liu, C.; Newell, G.; White, M. On the Selection of Thresholds for Predicting Species Occurrence with Presence-Only Data. Ecol. Evol. 2016, 6, 337–348. [Google Scholar] [CrossRef]
- Hellegers, M.; van Hinsberg, A.; Lenoir, J.; Dengler, J.; Huijbregts, M.A.; Schipper, A.M. Multiple Threshold-Selection Methods Are Needed to Binarise Species Distribution Model Predictions. Divers. Distrib. 2025, 31, e70019. [Google Scholar] [CrossRef]
- Velazco, S.J.E.; Rose, M.B.; De Marco, P., Jr.; Regan, H.M.; Franklin, J. How Far Can I Extrapolate My Species Distribution Model? Exploring Shape, a Novel Method. Ecography 2024, 2024, e06992. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The Art of Modelling Range-Shifting Species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Tamme, R.; Götzenberger, L.; Zobel, M.; Bullock, J.M.; Hooftman, D.A.P.; Kaasik, A.; Pärtel, M. Predicting Species’ Maximum Dispersal Distances from Simple Plant Traits. Ecology 2014, 95, 505–513. [Google Scholar] [CrossRef] [PubMed]
- McGarigal, K. FRAGSTATS: Spatial Pattern Analysis Program for Categorical Maps; Computer Software Program Produced by the Authors at the University of Massachusetts; University of Massachusetts: Amherst, MA, USA, 2002. [Google Scholar]
- Hesselbarth, M.H.K.; Sciaini, M.; With, K.A.; Wiegand, K.; Nowosad, J. Landscapemetrics: An Open-source R Tool to Calculate Landscape Metrics. Ecography 2019, 42, 1648–1657. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Hausser, J.; Chessel, D.; Perrin, N. Ecological-niche Factor Analysis: How to Compute Habitat-suitability Maps without Absence Data? Ecology 2002, 83, 2027–2036. [Google Scholar] [CrossRef]
- Garland, T., Jr.; Ives, A.R. Using the Past to Predict the Present: Confidence Intervals for Regression Equations in Phylogenetic Comparative Methods. Am. Nat. 2000, 155, 346–364. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Castiglione, S.; Tesone, G.; Piccolo, M.; Melchionna, M.; Mondanaro, A.; Serio, C.; Di Febbraro, M.; Raia, P. A New Method for Testing Evolutionary Rate Variation and Shifts in Phenotypic Evolution. Methods Ecol. Evol. 2018, 9, 974–983. [Google Scholar] [CrossRef]
- Linder, H.P. Plant Diversity and Endemism in Sub-Saharan Tropical Africa. J. Biogeogr. 2001, 28, 169–182. [Google Scholar] [CrossRef]
- Linder, H.P. On Areas of Endemism, with an Example from the African Restionaceae. Syst. Biol. 2001, 50, 892–912. [Google Scholar] [CrossRef]
- Rosauer, D.; Laffan, S.W.; Crisp, M.D.; Donnellan, S.C.; Cook, L.G. Phylogenetic Endemism: A New Approach for Identifying Geographical Concentrations of Evolutionary History. Mol. Ecol. 2009, 18, 4061–4072. [Google Scholar] [CrossRef]
- Daru, B.H.; Karunarathne, P.; Schliep, K. Phyloregion: R Package for Biogeographical Regionalization and Macroecology. Methods Ecol. Evol. 2020, 11, 1483–1491. [Google Scholar] [CrossRef]
- Daru, B.H.; Elliott, T.L.; Park, D.S.; Davies, T.J. Understanding the Processes Underpinning Patterns of Phylogenetic Regionalization. Trends Ecol. Evol. 2017, 32, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Daru, B.H.; Farooq, H.; Antonelli, A.; Faurby, S. Endemism Patterns Are Scale Dependent. Nat. Commun. 2020, 11, 2115. [Google Scholar] [CrossRef] [PubMed]
- Tsirogiannis, C.; Sandel, B. PhyloMeasures: A Package for Computing Phylogenetic Biodiversity Measures and Their Statistical Moments. Ecography 2016, 39, 709–714. [Google Scholar] [CrossRef]
- González-Orozco, C.E.; Pollock, L.J.; Thornhill, A.H.; Mishler, B.D.; Knerr, N.; Laffan, S.W.; Miller, J.T.; Rosauer, D.F.; Faith, D.P.; Nipperess, D.A.; et al. Phylogenetic Approaches Reveal Biodiversity Threats under Climate Change. Nat. Clim. Change 2016, 6, 1110–1114. [Google Scholar] [CrossRef]
- Getis, A.; Ord, J.K. The Analysis of Spatial Association by Use of Distance Statistics. Geogr. Anal. 1992, 4, 189–206. [Google Scholar] [CrossRef]
- Ord, J.K.; Getis, A. Local Spatial Autocorrelation Statistics: Distributional Issues and an Application. Geogr. Anal. 1995, 27, 286–306. [Google Scholar] [CrossRef]
- Parry, J. Sfdep: Spatial Dependence for Simple Features. R Package Version 0.2.3. 2022. Available online: https://cran.r-project.org/web/packages/sfdep/index.html (accessed on 6 March 2025).
- Braithwaite, A.; Li, Q. Transnational Terrorism Hot Spots: Identification and Impact Evaluation. Confl. Manag. Peace Sci. 2007, 24, 281–296. [Google Scholar] [CrossRef]
- Mitchel, A. The ESRI Guide to GIS Analysis, Volume 2: Spartial Measurements and Statistics. ESRI Press: Redlands, CA, USA, 2005; Volume 2. [Google Scholar]
- Nelson, T.A.; Boots, B. Detecting Spatial Hot Spots in Landscape Ecology. Ecography 2008, 31, 556–566. [Google Scholar] [CrossRef]
- Mann, H.B. Nonparametric Tests Against Trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Griffin Publishing: Santa Barbara, CA, USA, 1948. [Google Scholar]
- ESRI. How Emerging Hotspot Analysis Works. Available online: https://pro.arcgis.com/en/pro-app/latest/tool-reference/space-time-pattern-mining/learnmoreemerging.htm (accessed on 10 January 2024).
- Hamed, K.H. Exact Distribution of the Mann-Kendall Trend Test Statistic for Persistent Data. J. Hydrol. 2009, 365, 86–94. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity. Curr. Biol. 2021, 31, R1174–R1177. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.C.; Cardoso, P.; Gomes, P. Determining the Relative Roles of Species Replacement and Species Richness Differences in Generating Beta-diversity Patterns. Glob. Ecol. Biogeogr. 2012, 21, 760–771. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the Turnover and Nestedness Components of Beta Diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Cardoso, P.; Rigal, F.; Carvalho, J.C. BAT—Biodiversity Assessment Tools, an R Package for the Measurement and Estimation of Alpha and Beta Taxon, Phylogenetic and Functional Diversity. Methods Ecol. Evol. 2015, 6, 232–236. [Google Scholar] [CrossRef]
- Mota, F.M.M.; Alves-Ferreira, G.; Talora, D.C.; Heming, N.M. Divraster: An R Package to Calculate Taxonomic, Functional and Phylogenetic Diversity from Rasters. Ecography 2023, 2023, e06905. [Google Scholar] [CrossRef]
- Hanson, J. Wdpar: Interface to the World Database on Protected Areas. R Package Version 1.0.5. J. Open Source Softw. 2020, 7, 4594. [Google Scholar] [CrossRef]
- Pebesma, E. Simple Features for R: Standardized Support for Spatial Vector Data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef]
- Noroozi, J.; Naqinezhad, A.; Talebi, A.; Doostmohammadi, M.; Plutzar, C.; Rumpf, S.B.; Asgarpour, Z.; Schneeweiss, G.M. Hotspots of Vascular Plant Endemism in a Global Biodiversity Hotspot in Southwest Asia Suffer from Significant Conservation Gaps. Biol. Conserv. 2019, 237, 299–307. [Google Scholar] [CrossRef]
- Xu, Y.; Shen, Z.; Ying, L.; Wang, Z.; Huang, J.; Zang, R.; Jiang, Y. Hotspot Analyses Indicate Significant Conservation Gaps for Evergreen Broadleaved Woody Plants in China. Sci. Rep. 2017, 7, 1859. [Google Scholar] [CrossRef]
- Exavier, R.; Zeilhofer, P. OpenLand: Software for Quantitative Analysis and Visualization of Land Use and Cover Change. R J. 2020, 12, 359. [Google Scholar] [CrossRef]
- Stévart, T.; Dauby, G.; Lowry, P.P.; Blach-Overgaard, A.; Droissart, V.; Harris, D.J.; Mackinder, B.A.; Schatz, G.E.; Sonké, B.; Sosef, M.S.M.; et al. A Third of the Tropical African Flora Is Potentially Threatened with Extinction. Sci. Adv. 2019, 5, eaax9444. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, M.; Pickering, C.M. Tourism and Recreation: A Common Threat to IUCN Red-Listed Vascular Plants in Europe. Biodivers. Conserv. 2013, 22, 3027–3044. [Google Scholar] [CrossRef]
- Bergmeier, E.; Strid, A. Regional Diversity, Population Trends and Threat Assessment of the Weeds of Traditional Agriculture in Greece. Bot. J. Linn. Soc. 2014, 175, 607–623. [Google Scholar] [CrossRef]
- García-Vega, D.; Newbold, T. Assessing the Effects of Land Use on Biodiversity in the World’s Drylands and Mediterranean Environments. Biodivers. Conserv. 2020, 29, 393–408. [Google Scholar] [CrossRef]
- Panitsa, M.; Iliadou, E.; Kokkoris, I.; Kallimanis, A.; Patelodimou, C.; Strid, A.; Raus, T.; Bergmeier, E.; Dimopoulos, P. Distribution Patterns of Ruderal Plant Diversity in Greece. Biodivers. Conserv. 2020, 29, 869–891. [Google Scholar] [CrossRef]
- Lughadha, E.N.; Walker, B.E.; Canteiro, C.; Chadburn, H.; Davis, A.P.; Hargreaves, S.; Lucas, E.J.; Schuiteman, A.; Williams, E.; Bachman, S.P.; et al. The Use and Misuse of Herbarium Specimens in Evaluating Plant Extinction Risks. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20170402. [Google Scholar] [CrossRef]
- Zizka, A.; Azevedo, J.; Leme, E.; Neves, B.; da Costa, A.F.; Caceres, D.; Zizka, G. Biogeography and Conservation Status of the Pineapple Family (Bromeliaceae). Divers. Distrib. 2020, 26, 183–195. [Google Scholar] [CrossRef]
- Zizka, A.; Silvestro, D.; Vitt, P.; Knight, T.M. Automated Conservation Assessment of the Orchid Family Using Deep Learning. bioRxiv 2020, 1–25. [Google Scholar] [CrossRef]
- IUCN Standards and Petitions Committee. Guidelines for Using the IUCN Red List Categories and Criteria; Version 15.1, Prepared by the Standards and Petitions Committee; IUCN: Gland, Switzerland, 2022. [Google Scholar]
- Collen, B.; Dulvy, N.K.; Gaston, K.J.; Gärdenfors, U.; Keith, D.A.; Punt, A.E.; Regan, H.M.; Böhm, M.; Hedges, S.; Seddon, M.; et al. Clarifying Misconceptions of Extinction Risk Assessment with the IUCN Red List. Biol. Lett. 2016, 12, 20150843. [Google Scholar] [CrossRef]
- Isaac, N.J.B.; Turvey, S.T.; Collen, B.; Waterman, C.; Baillie, J.E.M. Mammals on the EDGE: Conservation Priorities Based on Threat and Phylogeny. PLoS ONE 2007, 2, e296. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning: Data Mining, Inference, and Prediction; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009; ISBN 0-387-84858-4. [Google Scholar]
- Manes, S.; Costello, M.J.; Beckett, H.; Debnath, A.; Devenish-Nelson, E.; Grey, K.-A.; Jenkins, R.; Khan, T.M.; Kiessling, W.; Krause, C. Endemism Increases Species’ Climate Change Risk in Areas of Global Biodiversity Importance. Biol. Conserv. 2021, 257, 109070. [Google Scholar] [CrossRef]
- Georghiou, K.; Delipetrou, P. Patterns and Traits of the Endemic Plants of Greece. Bot. J. Linn. Soc. 2010, 162, 130–422. [Google Scholar] [CrossRef]
- Sweeney, C.P.; Jarzyna, M.A. Assessing the Synergistic Effects of Land Use and Climate Change on Terrestrial Biodiversity: Are Generalists Always the Winners? Curr. Landsc. Ecol. Rep. 2022, 7, 41–48. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D. Habitat Fragmentation and Its Lasting Impact on Earth’s Ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- Keinath, D.A.; Doak, D.F.; Hodges, K.E.; Prugh, L.R.; Fagan, W.; Sekercioglu, C.H.; Buchart, S.H.; Kauffman, M. A Global Analysis of Traits Predicting Species Sensitivity to Habitat Fragmentation. Glob. Ecol. Biogeogr. 2017, 26, 115–127. [Google Scholar] [CrossRef]
- Brown, K.A.; Parks, K.E.; Bethell, C.A.; Johnson, S.E.; Mulligan, M. Predicting Plant Diversity Patterns in Madagascar: Understanding the Effects of Climate and Land Cover Change in a Biodiversity Hotspot. PLoS ONE 2015, 10, e0122721. [Google Scholar] [CrossRef]
- Montràs-Janer, T.; Suggitt, A.J.; Fox, R.; Jönsson, M.; Martay, B.; Roy, D.B.; Walker, K.J.; Auffret, A.G. Anthropogenic Climate and Land-Use Change Drive Short-and Long-Term Biodiversity Shifts across Taxa. Nat. Ecol. Evol. 2024, 8, 739–751. [Google Scholar] [CrossRef]
- Varela, D.; Romeiras, M.M.; Silva, L. Implications of Climate Change on the Distribution and Conservation of Cabo Verde Endemic Trees. Glob. Ecol. Conserv. 2022, 34, e02025. [Google Scholar] [CrossRef]
- Ogawa-Onishi, Y.; Berry, P.M.; Tanaka, N. Assessing the Potential Impacts of Climate Change and Their Conservation Implications in Japan: A Case Study of Conifers. Biol. Conserv. 2010, 143, 1728–1736. [Google Scholar] [CrossRef]
- La Montagna, D.; Attorre, F.; Hamdiah, S.; Maděra, P.; Malatesta, L.; Vahalík, P.; Van Damme, K.; De Sanctis, M. Climate Change Effects on the Potential Distribution of the Endemic Commiphora Species (Burseraceae) on the Island of Socotra. Front. For. Glob. Change 2023, 6, 1183858. [Google Scholar] [CrossRef]
- Upson, R.; Williams, J.J.; Wilkinson, T.P.; Clubbe, C.P.; Maclean, I.M.; McAdam, J.H.; Moat, J.F. Potential Impacts of Climate Change on Native Plant Distributions in the Falkland Islands. PLoS ONE 2016, 11, e0167026. [Google Scholar] [CrossRef] [PubMed]
- Schrader, J.; Weigelt, P.; Cai, L.; Westoby, M.; Fernández-Palacios, J.M.; Cabezas, F.J.; Plunkett, G.M.; Ranker, T.A.; Triantis, K.A.; Trigas, P. Islands Are Key for Protecting the World’s Plant Endemism. Nature 2024, 634, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Auffret, A.G.; Nenzén, H.; Polaina, E. Underprediction of Extirpation and Colonisation Following Climate and Land-use Change Using Species Distribution Models. Divers. Distrib. 2024, 30, e13834. [Google Scholar] [CrossRef]
- Cronk, Q. Plant Extinctions Take Time. Science 2016, 353, 446–447. [Google Scholar] [CrossRef]
- Plue, J.; Van Calster, H.; Auestad, I.; Basto, S.; Bekker, R.M.; Bruun, H.H.; Chevalier, R.; Decocq, G.; Grandin, U.; Hermy, M. Buffering Effects of Soil Seed Banks on Plant Community Composition in Response to Land Use and Climate. Glob. Ecol. Biogeogr. 2021, 30, 128–139. [Google Scholar] [CrossRef]
- Corlett, R.T. The Ecology of Plant Extinctions. Trends Ecol. Evol. 2024, 40, 286–295. [Google Scholar] [CrossRef]
- Mendes, S.B.; Olesen, J.M.; Memmott, J.; Costa, J.M.; Timóteo, S.; Dengucho, A.L.; Craveiro, L.; Heleno, R. Evidence of a European Seed Dispersal Crisis. Science 2024, 386, 206–211. [Google Scholar] [CrossRef]
- Artamendi, M.; Martin, P.A.; Bartomeus, I.; Magrach, A. Loss of Pollinator Diversity Consistently Reduces Reproductive Success for Wild and Cultivated Plants. Nat. Ecol. Evol. 2025, 9, 296–313. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kaloveloni, A.; Petanidou, T. Assessing Climate Change Impacts on Island Bees: The Aegean Archipelago. Biology 2022, 11, 552. [Google Scholar] [CrossRef]
- Lughadha, E.N.; Bachman, S.P.; Leão, T.C.C.; Forest, F.; Halley, J.M.; Moat, J.; Acedo, C.; Bacon, K.L.; Brewer, R.F.A.; Gâteblé, G.; et al. Extinction Risk and Threats to Plants and Fungi. Plants People Planet 2020, 2, 389–408. [Google Scholar] [CrossRef]
- Attorre, F.; Abeli, T.; Bacchetta, G.; Farcomeni, A.; Fenu, G.; Sanctis, M.D.; Gargano, D.; Peruzzi, L.; Montagnani, C.; Rossi, G.; et al. How to Include the Impact of Climate Change in the Extinction Risk Assessment of Policy Plant Species? J. Nat. Conserv. 2018, 44, 43–49. [Google Scholar] [CrossRef]
- Román-Palacios, C.; Wiens, J.J. Recent Responses to Climate Change Reveal the Drivers of Species Extinction and Survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef] [PubMed]
- Theodoridis, S.; Hickler, T.; Nogues-Bravo, D.; Ploch, S.; Mishra, B.; Thines, M. Satellite-Observed Mountain Greening Predicts Genomic Erosion in a Grassland Medicinal Herb over Half a Century. Curr. Biol. 2025. [Google Scholar] [CrossRef] [PubMed]
- Sarrou, E.; Doukidou, L.; Avramidou, E.V.; Martens, S.; Angeli, A.; Stagiopoulou, R.; Fyllas, N.M.; Tourvas, N.; Abraham, E.; Maloupa, E. Chemodiversity Is Closely Linked to Genetic and Environmental Diversity: Insights into the Endangered Populations of the Local Endemic Plant Sideritis Euboea Heldr. of Evia Island (Greece). J. Appl. Res. Med. Aromat. Plants 2022, 31, 100426. [Google Scholar] [CrossRef]
- Valli, A.-T.; Papaioannou, C.; Liveri, E.; Papasotiropoulos, V.; Trigas, P. Conservation Biology of Three Threatened Limonium Species Endemic to Zakynthos Island (Ionian Islands, Greece). Oryx 2024, 58, 587–599. [Google Scholar] [CrossRef]
- Valli, A.-T.; Koumandou, V.L.; Iatrou, G.; Andreou, M.; Papasotiropoulos, V.; Trigas, P. Conservation Biology of Threatened Mediterranean Chasmophytes: The Case of Asperula Naufraga Endemic to Zakynthos Island (Ionian Islands, Greece). PLoS ONE 2021, 16, e0246706. [Google Scholar] [CrossRef]
- Augustinos, A.; Sotirakis, K.; Trigas, P.; Kalpoutzakis, E.; Papasotiropoulos, V. Genetic Variation in Three Closely Related Minuartia (Caryophyllaceae) Species Endemic to Greece: Implications for Conservation Management. Folia Geobot. 2014, 49, 603–621. [Google Scholar] [CrossRef]
- Liveri, E.; Passa, K.; Papasotiropoulos, V. The Contribution of Genetic and Genomic Tools in Diversity Conservation: The Case of Endemic Plants of Greece. J. Zool. Bot. Gard. 2024, 5, 276–293. [Google Scholar] [CrossRef]
- Rahbek, C.; Borregaard, M.K.; Colwell, R.K.; Dalsgaard, B.; Holt, B.G.; Morueta-Holme, N.; Nogues-Bravo, D.; Whittaker, R.J.; Fjeldså, J. Humboldt’s Enigma: What Causes Global Patterns of Mountain Biodiversity? Science 2019, 365, 1108–1113. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Field, R.; Grytnes, J.A.; Trigas, P.; Ah-Peng, C.; Attorre, F.; Birks, H.J.B.; Borges, P.A.V.; Cardoso, P.; Chou, C.H.; et al. Topography-Driven Isolation, Speciation and a Global Increase of Endemism with Elevation. Glob. Ecol. Biogeogr. 2016, 25, 1097–1107. [Google Scholar] [CrossRef]
- Antonelli, A.; Kissling, W.D.; Flantua, S.G.A.; Bermúdez, M.A.; Mulch, A.; Muellner-Riehl, A.N.; Kreft, H.; Linder, H.P.; Badgley, C.; Fjeldså, J.; et al. Geological and Climatic Influences on Mountain Biodiversity. Nat. Geosci. 2018, 11, 718–725. [Google Scholar] [CrossRef]
- Perrigo, A.; Hoorn, C.; Antonelli, A. Why Mountains Matter for Biodiversity. J. Biogeogr. 2020, 47, 315–325. [Google Scholar] [CrossRef]
- Panitsa, M.; Kontopanou, A. Diversity of Chasmophytes in the Vascular Flora of Greece: Floristic Analysis and Phytogeographical Patterns. Bot. Serbica 2017, 41, 199–211. [Google Scholar]
- Kontopanou, A.; Panitsa, M. Habitat Islands on the Aegean Islands (Greece): Elevational Gradient of Chasmophytic Diversity, Endemism, Phytogeographical Patterns and Need for Monitoring and Conservation. Diversity 2020, 12, 33. [Google Scholar] [CrossRef]
- Panitsa, M.; Kokkoris, I.P.; Kougioumoutzis, K.; Kontopanou, A.; Bazos, I.; Strid, A.; Dimopoulos, P. Linking Taxonomic, Phylogenetic and Functional Plant Diversity with Ecosystem Services of Cliffs and Screes in Greece. Plants 2021, 10, 992. [Google Scholar] [CrossRef]
- Médail, F.; Baumel, A. Using Phylogeography to Define Conservation Priorities: The Case of Narrow Endemic Plants in the Mediterranean Basin Hotspot. Biol. Conserv. 2018, 224, 258–266. [Google Scholar] [CrossRef]
- Kokkoris, I.P.; Skuras, D.; Maniatis, Y.; Dimopoulos, P. Natura 2000 Public Awareness in EU: A Prerequisite for Successful Conservation Policy. Land Use Policy 2023, 125, 106482. [Google Scholar] [CrossRef]
- Rubenstein, M.A.; Weiskopf, S.R.; Bertrand, R.; Carter, S.L.; Comte, L.; Eaton, M.J.; Johnson, C.G.; Lenoir, J.; Lynch, A.J.; Miller, B.W. Climate Change and the Global Redistribution of Biodiversity: Substantial Variation in Empirical Support for Expected Range Shifts. Environ. Evid. 2023, 12, 7. [Google Scholar] [CrossRef]
- Lamprecht, A.; Pauli, H.; Fernández Calzado, M.R.; Lorite, J.; Molero Mesa, J.; Steinbauer, K.; Winkler, M. Changes in Plant Diversity in a Water-Limited and Isolated High-Mountain Range (Sierra Nevada, Spain). Alp. Bot. 2021, 131, 27–39. [Google Scholar] [CrossRef]
- Urban, M.C. Escalator to Extinction. Proc. Natl. Acad. Sci. USA 2018, 115, 11871–11873. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated Increase in Plant Species Richness on Mountain Summits Is Linked to Warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, J.; Gégout, J.; Guisan, A.; Vittoz, P.; Wohlgemuth, T.; Zimmermann, N.E.; Dullinger, S.; Pauli, H.; Willner, W.; Svenning, J. Going against the Flow: Potential Mechanisms for Unexpected Downslope Range Shifts in a Warming Climate. Ecography 2010, 33, 295–303. [Google Scholar] [CrossRef]
- Daru, B.H.; Davies, T.J.; Willis, C.G.; Meineke, E.K.; Ronk, A.; Zobel, M.; Pärtel, M.; Antonelli, A.; Davis, C.C. Widespread Homogenization of Plant Communities in the Anthropocene. Nat. Commun. 2021, 12, 6983. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B.D.; Kent, J. Biodiversity Hotspots for Conservation Priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Guo, W.-Y.; Serra-Diaz, J.M.; Eiserhardt, W.L.; Maitner, B.S.; Merow, C.; Violle, C.; Pound, M.J.; Sun, M.; Slik, F.; Blach-Overgaard, A. Climate Change and Land Use Threaten Global Hotspots of Phylogenetic Endemism for Trees. Nat. Commun. 2023, 14, 6950. [Google Scholar] [CrossRef]
- Minev-Benzecry, S.; Daru, B.H. Climate Change Alters the Future of Natural Floristic Regions of Deep Evolutionary Origins. Nat. Commun. 2024, 15, 9474. [Google Scholar] [CrossRef]
- Múgica, A.; Miranda, H.; García, M. Survival Patterns and Population Stability of Cliff Plants Suggest High Resistance to Climatic Variability. Basic Appl. Ecol. 2024, 80, 128–134. [Google Scholar] [CrossRef]
- Snogerup, S. Evolutionary and Plant Geographical Aspects of Chasmophytic Communities; Botanical Society of Edinburgh: Edinburgh, UK, 1971. [Google Scholar]
- Greuter, W. The Origins and Evolution of Islands Flora as Exemplified by the Aegean Archipelago: 87-106 in BRAMWELL; Academic Press: Cambridge, MA, USA, 1979. [Google Scholar]
- Greuter, W. The Relict Element of the Flora of Crete and its Evolutionary Significance. In Taxonomy, Phylogeography and Evolution; Valentine, D.H., Ed.; Academic Press: Cambridge, MA, USA, 1972; pp. 161–177. [Google Scholar]
- Aguirre-Liguori, J.A.; Ramírez-Barahona, S.; Gaut, B.S. The Evolutionary Genomics of Species’ Responses to Climate Change. Nat. Ecol. Evol. 2021, 5, 1350–1360. [Google Scholar] [CrossRef]
- Jaros, U.; Tribsch, A.; Comes, H.P. Diversification in Continental Island Archipelagos: New Evidence on the Roles of Fragmentation, Colonization and Gene Flow on the Genetic Divergence of Aegean Nigella (Ranunculaceae). Ann. Bot. 2018, 121, 241–254. [Google Scholar] [CrossRef]
- Strid, A. Studies in the Aegean Flora. XVI. Biosystematics of the Nigella Arvensis Complex. With Special Reference to the Problem of Non-Adaptive Radiation. Opera Bot. 1970, 28, 1–169. [Google Scholar]
- Von Bothmer, R. Studies in the Aegean Flora. XXI. Biosystematic Studies in the Allium Ampeloprasum Complex; CABI: Wallingford, UK, 1974. [Google Scholar]
- Braz Pires, M.; Kougioumoutzis, K.; Norder, S.; Dimopoulos, P.; Strid, A.; Panitsa, M. Current Greek Protected Areas Fail to Fully Capture Shifting Endemism Hotspots Under Future Climate and Land-Use Change. 2024. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=5014808 (accessed on 6 March 2025).
- Cui, D.; Frazier, A.; Liang, S.; Roehrdanz, P.; Hurtt, G.; Zhu, Z.; Wang, D. Projected Climate Zone Shifts Could Undermine the Effectiveness of Global Protected Areas for Biodiversity Conservation by the Mid-to-Late Century. Glob. Environ. Change Adv. 2025, 5, 100017. [Google Scholar] [CrossRef]
- Dobrowski, S.Z.; Littlefield, C.E.; Lyons, D.S.; Hollenberg, C.; Carroll, C.; Parks, S.A.; Abatzoglou, J.T.; Hegewisch, K.; Gage, J. Protected-Area Targets Could Be Undermined by Climate Change-Driven Shifts in Ecoregions and Biomes. Commun. Earth Environ. 2021, 2, 198. [Google Scholar] [CrossRef]
- Wang, D.; de Knegt, H.J.; Hof, A.R. The Effectiveness of a Large Protected Area to Conserve a Global Endemism Hotspot May Vanish in the Face of Climate and Land-Use Changes. Front. Ecol. Evol. 2022, 10, 984842. [Google Scholar] [CrossRef]
- Reside, A.E.; Butt, N.; Adams, V.M. Adapting Systematic Conservation Planning for Climate Change. Biodivers. Conserv. 2018, 27, 1–29. [Google Scholar] [CrossRef]
- Spiliopoulou, K.; Dimitrakopoulos, P.G.; Brooks, T.M.; Kelaidi, G.; Paragamian, K.; Kati, V.; Oikonomou, A.; Vavylis, D.; Trigas, P.; Lymberakis, P. The Natura 2000 Network and the Ranges of Threatened Species in Greece. Biodivers. Conserv. 2021, 30, 945–961. [Google Scholar] [CrossRef]
- Spiliopoulou, K.; Brooks, T.M.; Dimitrakopoulos, P.G.; Oikonomou, A.; Karavatsou, F.; Stoumboudi, M.T.; Triantis, K.A. Protected Areas and the Ranges of Threatened Species: Towards the EU Biodiversity Strategy 2030. Biol. Conserv. 2023, 284, 110166. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Koufaki, T.; Sazeides, C.I.; Spyroglou, G.; Theodorou, K. Potential Impacts of Climate Change on the Habitat Suitability of the Dominant Tree Species in Greece. Plants 2022, 11, 1616. [Google Scholar] [CrossRef]
- Arianoutsou, M.; Christopoulou, A.; Kazanis, D.; Tountas, T.; Ganou, E.; Bazos, I.; Kokkoris, Y. Effects of Fire on High Altitude Coniferous Forests of Greece. In Proceedings of the VI International Forest Fire Research Conference, Coimbra, Portugal, 15–18 November 2010. [Google Scholar]
- Sirami, C.; Caplat, P.; Popy, S.; Clamens, A.; Arlettaz, R.; Jiguet, F.; Brotons, L.; Martin, J.L. Impacts of Global Change on Species Distributions: Obstacles and Solutions to Integrate Climate and Land Use. Glob. Ecol. Biogeogr. 2017, 26, 385–394. [Google Scholar] [CrossRef]
- Martin, Y.; Dyck, H.V.; Dendoncker, N.; Titeux, N. Testing Instead of Assuming the Importance of Land Use Change Scenarios to Model Species Distributions under Climate Change. Glob. Ecol. Biogeogr. 2013, 22, 1204–1216. [Google Scholar] [CrossRef]
- Brown, J.L.; Carnaval, A.C. A Tale of Two Niches: Methods, Concepts, and Evolution. Front. Biogeogr. 2019, 11, e44158. [Google Scholar] [CrossRef]
- Qiao, H.; Feng, X.; Escobar, L.E.; Peterson, A.T.; Soberón, J.; Zhu, G.; Papeş, M. An Evaluation of Transferability of Ecological Niche Models. Ecography 2019, 42, 521–534. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kougioumoutzis, K.; Kokkoris, I.P.; Trigas, P.; Strid, A.; Dimopoulos, P. Projected Impacts of Climate and Land Use Change on Endemic Plant Distributions in a Mediterranean Island Hotspot: The Case of Evvia (Aegean, Greece). Climate 2025, 13, 100. https://doi.org/10.3390/cli13050100
Kougioumoutzis K, Kokkoris IP, Trigas P, Strid A, Dimopoulos P. Projected Impacts of Climate and Land Use Change on Endemic Plant Distributions in a Mediterranean Island Hotspot: The Case of Evvia (Aegean, Greece). Climate. 2025; 13(5):100. https://doi.org/10.3390/cli13050100
Chicago/Turabian StyleKougioumoutzis, Konstantinos, Ioannis P. Kokkoris, Panayiotis Trigas, Arne Strid, and Panayotis Dimopoulos. 2025. "Projected Impacts of Climate and Land Use Change on Endemic Plant Distributions in a Mediterranean Island Hotspot: The Case of Evvia (Aegean, Greece)" Climate 13, no. 5: 100. https://doi.org/10.3390/cli13050100
APA StyleKougioumoutzis, K., Kokkoris, I. P., Trigas, P., Strid, A., & Dimopoulos, P. (2025). Projected Impacts of Climate and Land Use Change on Endemic Plant Distributions in a Mediterranean Island Hotspot: The Case of Evvia (Aegean, Greece). Climate, 13(5), 100. https://doi.org/10.3390/cli13050100