Phylogenetic and Comparative Analyses of Complete Chloroplast Genomes of Chinese Viburnum and Sambucus (Adoxaceae)


College of Plant Protection, Henan Agricultural University, Zhengzhou 450002, China
*
Author to whom correspondence should be addressed.
Plants 2020, 9(9), 1143; https://doi.org/10.3390/plants9091143
Submission received: 8 July 2020
/
Revised: 23 August 2020
/
Accepted: 1 September 2020
/
Published: 3 September 2020
(This article belongs to the Special Issue Germplasm Diversity for Sustainability and Crop Improvement)
Abstract
:Phylogenetic analyses of complete chloroplast genome sequences have yielded significant improvements in our understanding of relationships in the woody flowering genus Viburnum (Adoxaceae, Dipsacales); however, these relationships were evaluated focusing only on Viburnum species within Central and South America and Southeast Asia. By contrast, despite being a hotspot of Viburnum diversity, phylogenetic relationships of Viburnum species in China are less well known. Here, we characterized the complete chloroplast (cp) genomes of 21 Viburnum species endemic to China, as well as three Sambucus species. These 24 plastomes were highly conserved in genomic structure, gene order and content, also when compared with other Adoxaceae. The identified repeat sequences, simple sequence repeats (SSRs) and highly variable plastid regions will provide potentially valuable genetic resources for further population genetics and phylogeographic studies on Viburnum and Sambucus. Consistent with previous combined phylogenetic analyses of 113 Viburnum species, our phylogenomic analyses based on the complete cp genome sequence dataset confirmed the sister relationship between Viburnum and the Sambucus-Adoxa-Tetradoxa-Sinadoxa group, the monophyly of four recognized sections in Flora of China (i.e., Viburnum sect. Tinus, Viburnum sect. Solenotinus, Viburnum sect. Viburnum and Viburnum sect. Pseudotinus) and the nonmonophyly of Viburnum sect. Odontotinus and Viburnum sect. Megalotinus. Additionally, our study confirmed the sister relationships between the clade Valvatotinus and Viburnum sect. Pseudotinus, as well as between Viburnum sect. Opulus and the Odontotinus-Megalotinus group. Overall, our results clearly document the power of the complete cp genomes in improving phylogenetic resolution, and will contribute to a better understanding of plastome evolution in Chinese Adoxaceae.
1. Introduction
The eudicot family Adoxaceae (Dipsacales) sensu APG IV contains three small herbaceous genera (less than 10 species) (i.e., Adoxa, Sinadoxa, and Tetradoxa) and two larger genera (i.e., Viburnum and Sambucus) [1]. The woody flowering taxon Viburnum, with approximately 200 species of shrubs and small trees [2], is the largest genus within Adoxaceae, and is of great interest to the horticultural community, since more than 70 of these species (and a variety of artificial hybrids) have been brought into cultivation [3]. Although widely distributed in the Northern Hemisphere, Viburnum has major centers of species diversity in eastern Asia and Central and South America [4,5,6], with significant extensions into the montane forests of Southeast Asia [7] and South America [8]. Sambucus is a relatively small genus occurring mostly in the north temperate zone, comprising about 10 species of small trees, shrubs and perennial herbs [5,9,10], of which many species are cultivated ornamentally, and several produce edible fruits (https://www.britannica.com/plant/Dipsacales). In addition, several species are commonly used in folk medicine (e.g., S. adnata, S. javanica and S. nigra) [11]. Within Adoxaceae, analyses of complete cp genome sequences suggested that Sambucus and Viburnum were the most closely related [12]; more specifically, Viburnum was likely to be the sister group of Sambucus plus Adoxa and its relatives. Although both genera have important horticultural value, limited molecular markers were available for the application, breeding and conservation of these species in the context of population genetics and phylogenetic studies.
Based on various morphological characteristics (e.g., endocarp shape, inflorescence form, leaf morphology, the presence or absence of naked buds and of sterile flowers around the margins of the inflorescences), Viburnum has been subdivided by several researchers, most commonly into ten groups formally recognized as sections [5,13]. Over the past decade, great advances have been made in understanding Viburnum phylogeny [14,15,16,17,18]. The number of species sampled in phylogenetic studies has increased from 40 to 90, representing all major clades within the genus. Additionally, sampling has increased from four to ten genes, thus affording better phylogenetic resolution. These phylogenetic studies have uniformly and strongly supported earlier recognized sections and subsections, while encountering difficulties resolving the relationships with confidence based on limited parsimony informative sites, in particularly with recent divergences within groups of closely related species [19]. Nonetheless, a recent study of 22 species provided us, for the first time, with comparatively high-resolution data of nearly all of the deepest branching events within Viburnum in light of next-generation sequencing of whole plastid genomes [19]. This study demonstrated a reliable framework within which to assess the power of complete cp genome markers and methods to discriminate Viburnum species in Central and South America (16 species) and Southeast Asia (6 species). By contrast, China is considered to be one of the hotspots of Viburnum plant taxa diversity; a total of 8 sections and c. 73 species have been found in this region [2]. Nevertheless, the phylogenetic relationships of Viburnum species there have received much less attention.
In the present study, we reported whole-plastome sequence data for 21 species of Viburnum, covering all of the eight currently diagnosed sections in Flora of China, as well as for three species of Sambucus. The main goals of this study were to: (1) characterize and compare the cp genomes of Viburnum species belonging to all the eight sections occurring in China and related taxa in order to gain insights into their evolutionary patterns; (2) examine the phylogenetic relationships of the main clades of Chinese Adoxaceae, with a particular focus on the generic status of Viburnum; and (3) screen and identify repeat sequences, simple sequence repeats (SSRs) and mutational hotspot regions for future species identification and phylogeographic studies of the two genera.
2. Results and Discussion
2.1. Chloroplast Genome Assembly and Features
With the Illumina HiSeq 2500 system (San Diego, CA, USA), we sequenced the plastomes of 21 species of Viburnum and 3 species of Sambucus. Of these samples, through de novo assembly, the maximum number of assembled contigs ranged from 61,001 (V. odoratissimum) to 388,130 (V. melanocarpum), with N50 contigs varying from 285 to 399 bp. Average sequencing depth ranged from about 268× (S. adnata) to 517× (V. melanocarpum) (Table S1). Subsequently, through reference-based assembly, a total of 165–209 contigs were successfully mapped to the reference plastomes. Among these, three to eight long contigs (>10 kb) that were found to be significantly homologous to the reference genome were combined to generate each chloroplast genome, with no gaps found. The four junctions between IRs and SSC/LSC in each species were initially determined on the basis of these long contigs, and then verified by PCR-based sequencing. The results showed that the assembly sequences were totally identical with the PCR amplified fragments, demonstrating the high quality of our assembly. Finally, we obtained 24 whole chloroplast genome sequences without gaps after de novo and reference-guided assembly, and submitted them to GenBank with accession numbers MT507585–MT507605 for Viburnum and MT457821–MT457823 for Sambucus (Table S1).
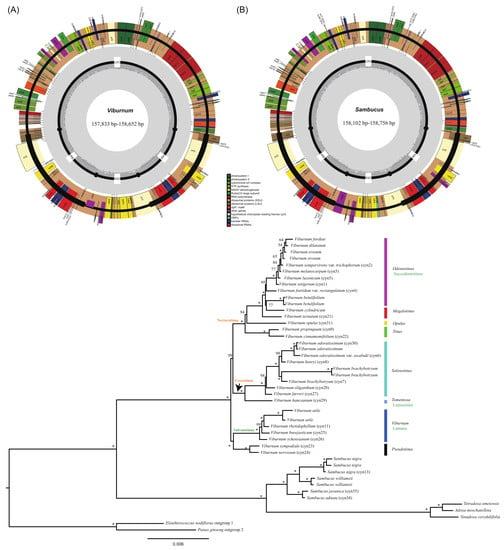
The complete cp genomes of the 21 Viburnum species were determined to be 157,833–158,652 bp in size, and the three Sambucus species ranged from 158,102 bp (S. nigra) to 158,756 bp (S. adnata) (Table 1). Akin to most land plant species, all of these plastomes exhibited a typical quadripartite structure, including a pair of IR regions (26,272–26,564 bp) separating the LSC region (86,430–87,892 bp) and the SSC region (17,674–18,978 bp). The overall GC content in the whole genome sequences was practically identical among these plastomes (38.0–38.2%). The 21 Viburnum cp genomes encoded the same 130 functional genes, consisting of 85 protein-coding genes, 37 transfer RNA (tRNA) genes and 8 ribosomal RNA (rRNA) genes. The 3 Sambucus cp genomes encoded identical sets of 132 genes, with 84 protein-coding genes, 40 tRNA genes and 8 rRNA genes (Table 1). Notably, five genes (i.e., trnM-CAU, trnT-GGU, trnP-GGG, orf188 and lhbA) and three genes (i.e., psbZ, ndhH and rpl22) were only present in Sambucus and Viburnum, respectively. For both genera, 15 genes possessed a single intron (nine protein-coding genes and six tRNA genes), while 3 (ycf3, clpP and rps12) contained two introns, and a total of 17 genes were duplicated in the IR regions (Table 1). In particular, the rps12 was a transspliced gene, with the first exon located in the LSC region, and the second and third in the IR regions. We also found that the ycf1 gene at the SSC and IRa junction was present as a pseudogene in 16 Viburnum species (Table 2), due to the incomplete gene duplication, as shown in previous reports [20,21]. In addition, there were some exceptions where non-ATG codons were translated as Met and identified as start codons, such as GCT for psbL, GTG for rps19 and CTG for ndhD, which has also been observed in many other angiosperms, for instance, Betula platyphylla [22] and Punica granatum [23].
The newly obtained whole plastome sequences for 21 Viburnum species, plus three Sambucus taxa, vary only slightly in size (157,833–158,756 bp) (Table 1), and are greatly similar in overall structure, gene content and arrangement (Figure 1) compared with most other reported Adoxaceae cp genomes [12,24,25].
2.2. Expansion and Contraction of the Inverted Repeat Regions
The IR/single copy (SC) region junctions were analyzed across the 21 Viburnum and 3 Sambucus cp genomes (Figure 2). The trnN-GUU/ndhF and rpl2/trnH-GUG genes were detected around the IRb/SSC and IRa/LSC junction regions, respectively. The LSC/IRb junction was found to reside within the rps19 gene, and the SSC/IRa junction was located in the ycf1 gene. Although the boundaries of these genomic regions were highly conserved, we still observed minor differences between the two genera. At the LSC/IRb junction, except for V. rhytidophyllum, the IRb regions expanded by 32 bp and 116 bp toward the rps19 gene of the remaining Viburnum species and Sambucus species, respectively. The ndhF gene crossed over the IRb/SSC junction in V. cinnamomifolium and overlapped with the IRb region by 135 bp. It was located at the SSC region in all other Viburnum and Sambucus species, and the whole length varied from 2187 bp to 2250 bp. Notably, the ndhF gene was found to be inverted in all Adoxaceae [26], possibly due to an early stage of the IR expansion followed by a contraction of the boundary. As for the ycf1 gene, there were 4147–4334 bp sequences located at SSC in Viburnum and uniformly 4574 bp in Sambucus, while the fragments in IRa ranged from 1364 bp to 1547 bp in Viburnum, and from 1115 bp to 1126 bp in Sambucus. The rpl2 gene was invariable within species in both Viburnum (1490 bp) and Sambucus (1498 bp). In addition, all the trnH-GUG genes within the Adoxaceae species studied here had an equal length of 75 bp except for V. oliganthum (78 bp). Similar IR/SC boundary structures shared among Adoxaceae species have also been reported in previous plastome studies [12,26].
2.3. Sequence Divergence Analysis
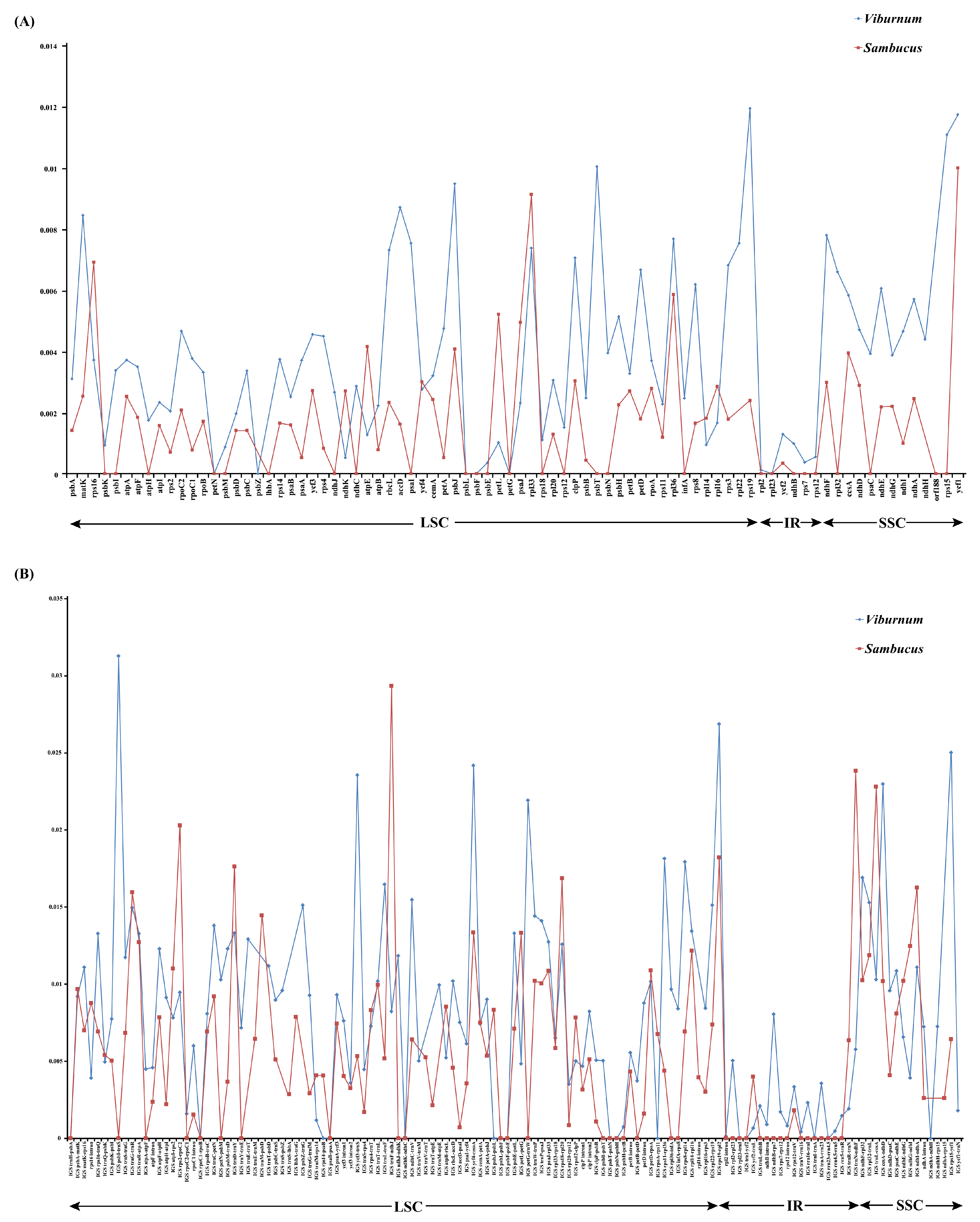
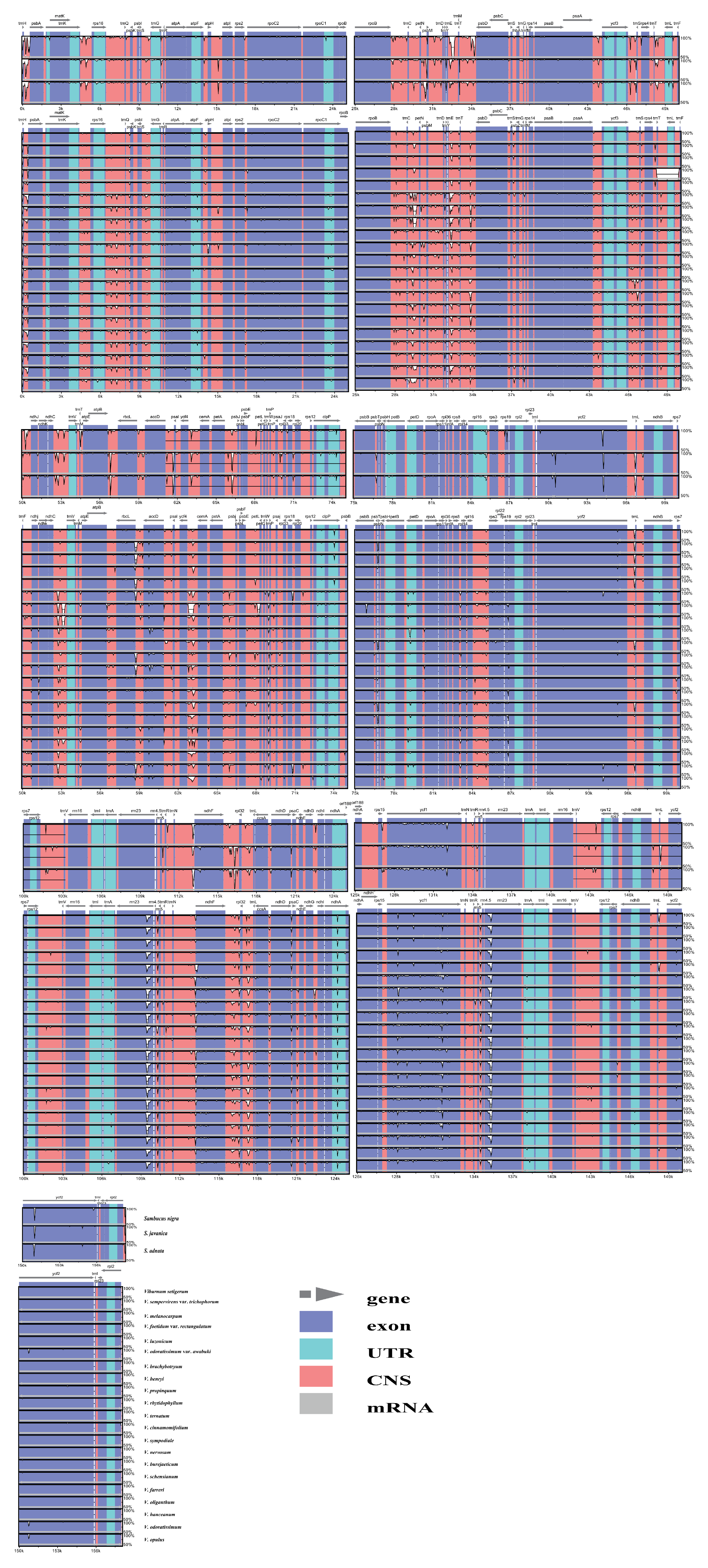
To analyze the level of comprehensive sequence divergence, the 21 Viburnum and 3 Sambucus cp genome sequences were compared and plotted using the mVISTA program (See Appendix A, Figure A1). Based on the overall sequence identity, similar to most of the angiosperms, our results indicated that the LSC and SSC regions were more divergent and variable than the two IR regions [27]. In addition, the cp genomes among species in both genera showed few differences. We calculated Pi values for 213 regions in total [including 82 CDSs, 117 IGSs (intergenic spacers) and 14 introns; Figure 3]. The mean Pi values of the coding regions were 0.00418 and 0.00255, respectively, for Viburnum and Sambucus, i.e., higher than the noncoding regions (Viburnum: 0.0092; Sambucus: 0.00785), as found in the majority of angiosperms [28]. Among coding regions, the Pi values for each region ranged from 0.00012 (rpl2) to 0.01193 (rps19) in Viburnum, among which 10 had high values (Pi > 0.007; Table 3). In contrast, within Sambucus, the Pi values varied from 0.00034 (ycf2) to 0.00999 (ycf1), and the 10 most variable regions had Pi values > 0.002 (Table 3). For the 81 noncoding regions, the Pi values ranged from 0.00014 (rpl2 intron) to 0.03129 (psbI-trnS) in Viburnum, and 0.00072 (accD-psaI) to 0.02934 (trnF-ndhJ) in Sambucus. The 10 most variable regions in both genera had Pi values > 0.01 (Table 3).
The chloroplast DNA region has already been used to explore the phylogenetic structure and phylogeographic patterns at different taxonomic levels. For instance, hypervariable regions of cpDNA (e.g., matK, ndhF, rbcL, petB-petD, rpl32-trnL, trnC-ycf6, trnH-psbA, trnK intron and trnS-trnG) were used to infer phylogenetic relationships for several studies with Viburnum [17,18,19]. Despite increased levels of confidence being revealed in most of the early branches, the relationships within clades of closely related species were still poorly resolved. Most regions used in these studies are today considered low to intermediately variable regions with low Pi values (Figure 3). Additionally, only rpl32-trnL is among the most informative regions of the plastome for most groups (Table 3). Thus, additional phylogenetically informative markers should be included to enhance the phylogenetic resolution in low-level phylogenetic or phylogeographic studies.
2.4. Characterization of Repeat Sequences and SSR Polymorphisms
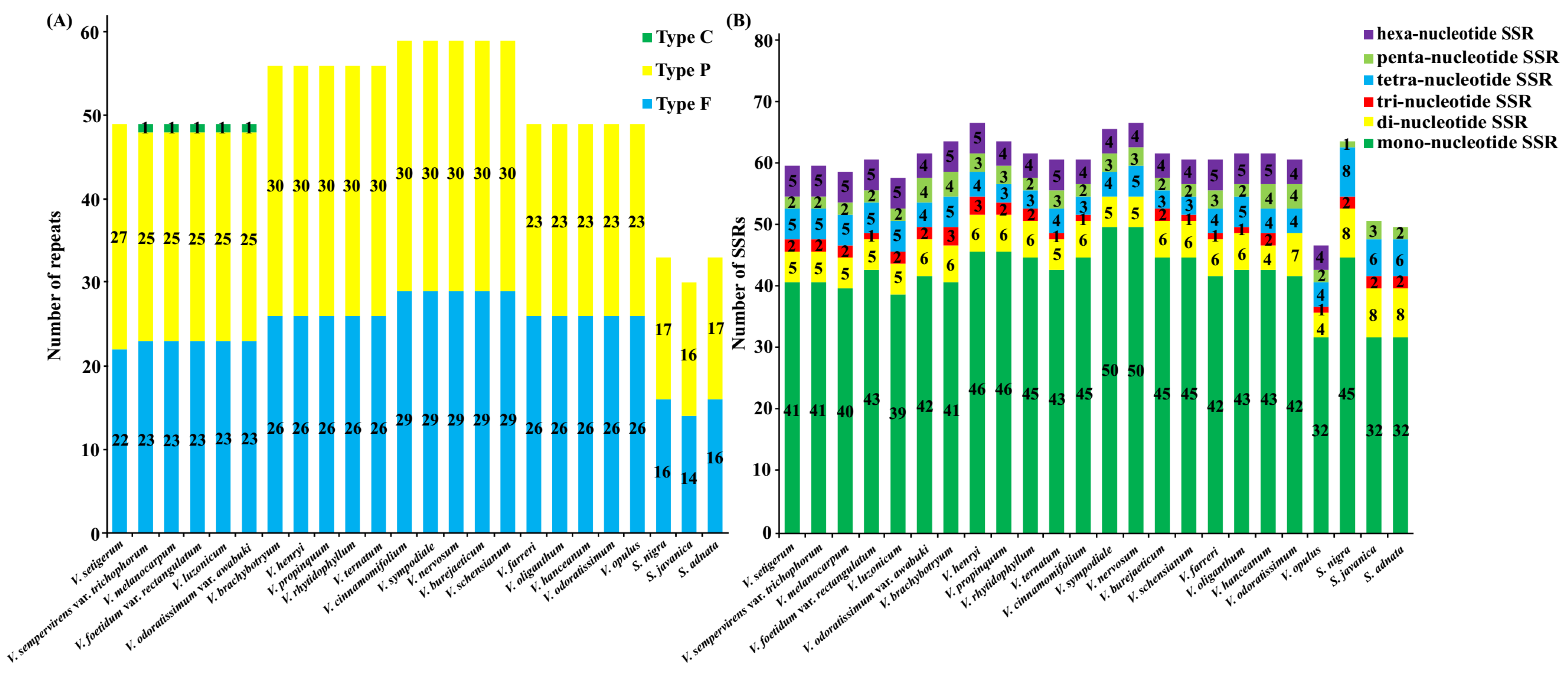
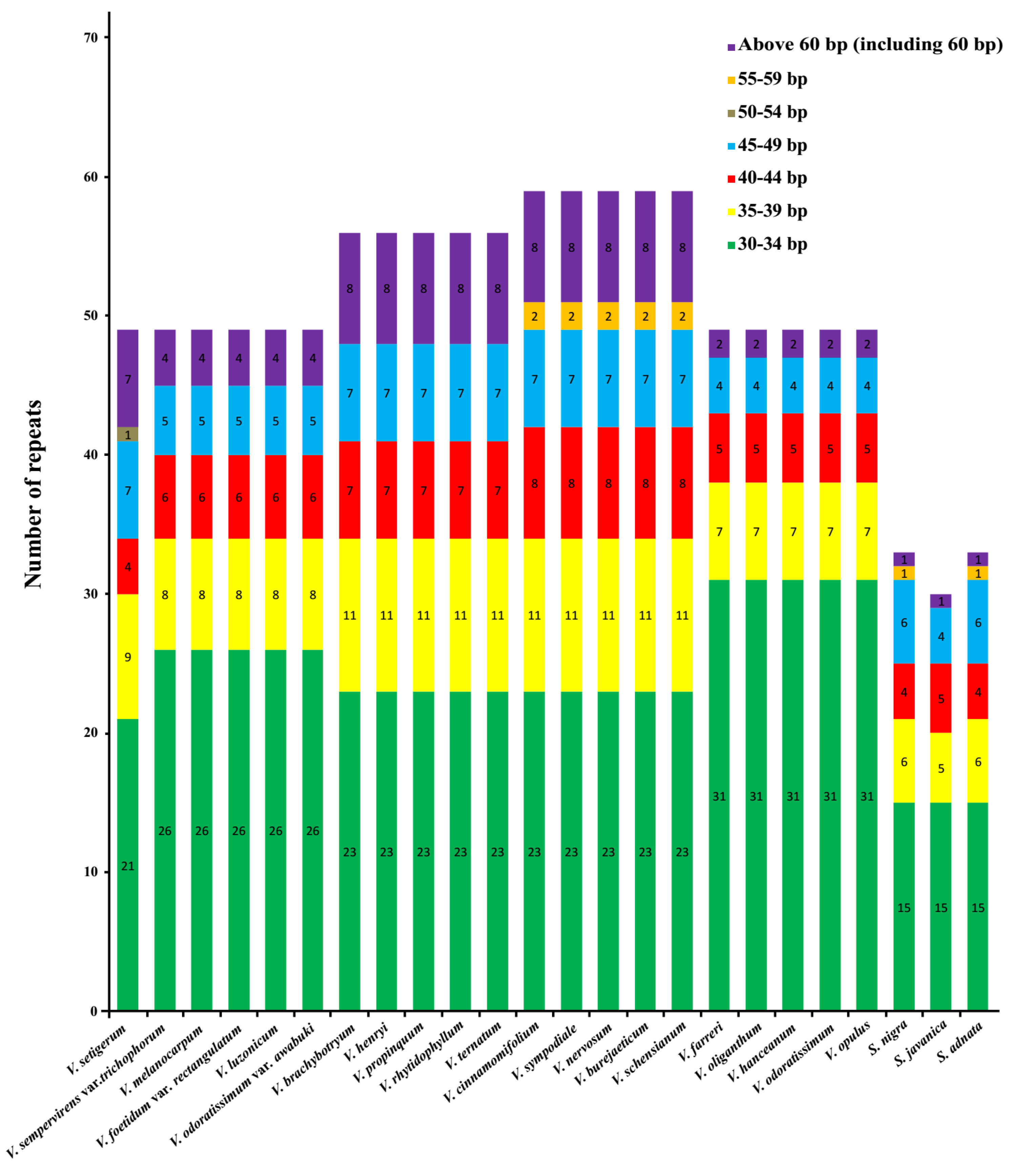
The distribution of repetitive sequences in the cp genomes of the two genera was quite similar: the palindromic repeats were the most abundant repeat category in 16 of 21 Viburnum species and three Sambucus species, followed by forward repeats. The complement repeat was detected and occurred once only in V. sempervirens var. trichophorum, V. melanocarpum, V. foetidum var. rectangulatum, V. luzonicum, and V. odoratissimum var. awabuki (Figure 4A). On the whole, the number of both total repeats and each category of repeats (i.e., palindromic and forward repeats) in the 21 Viburnum species was much higher than that in the three Sambucus species. In all 24 plastomes, most of these repeats exhibited lengths between 30 and 59 bp, and only a minority showed long repeats, i.e., more than 60 bp in size (See Appendix A, Figure A2). In addition, the repeats were more frequently distributed in gene regions or intergenic spacer regions than in intron regions within the family Adoxaceae (See Appendix A, Figure A3). These repeat motifs have promoted the rearrangement of the cp genomes and increased the genetic diversity of populations [29], and usually are useful markers in phylogenetic analyses [30,31].
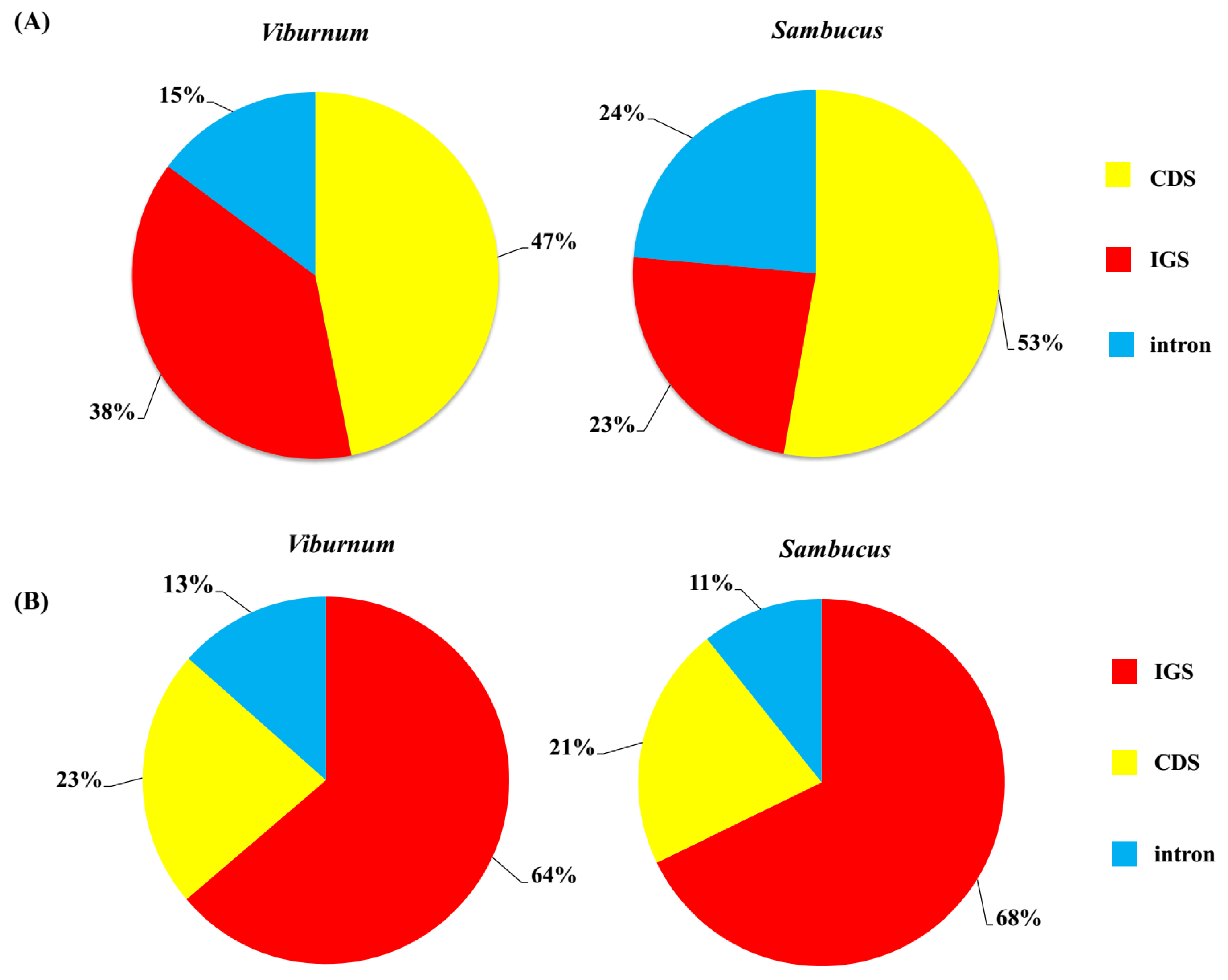
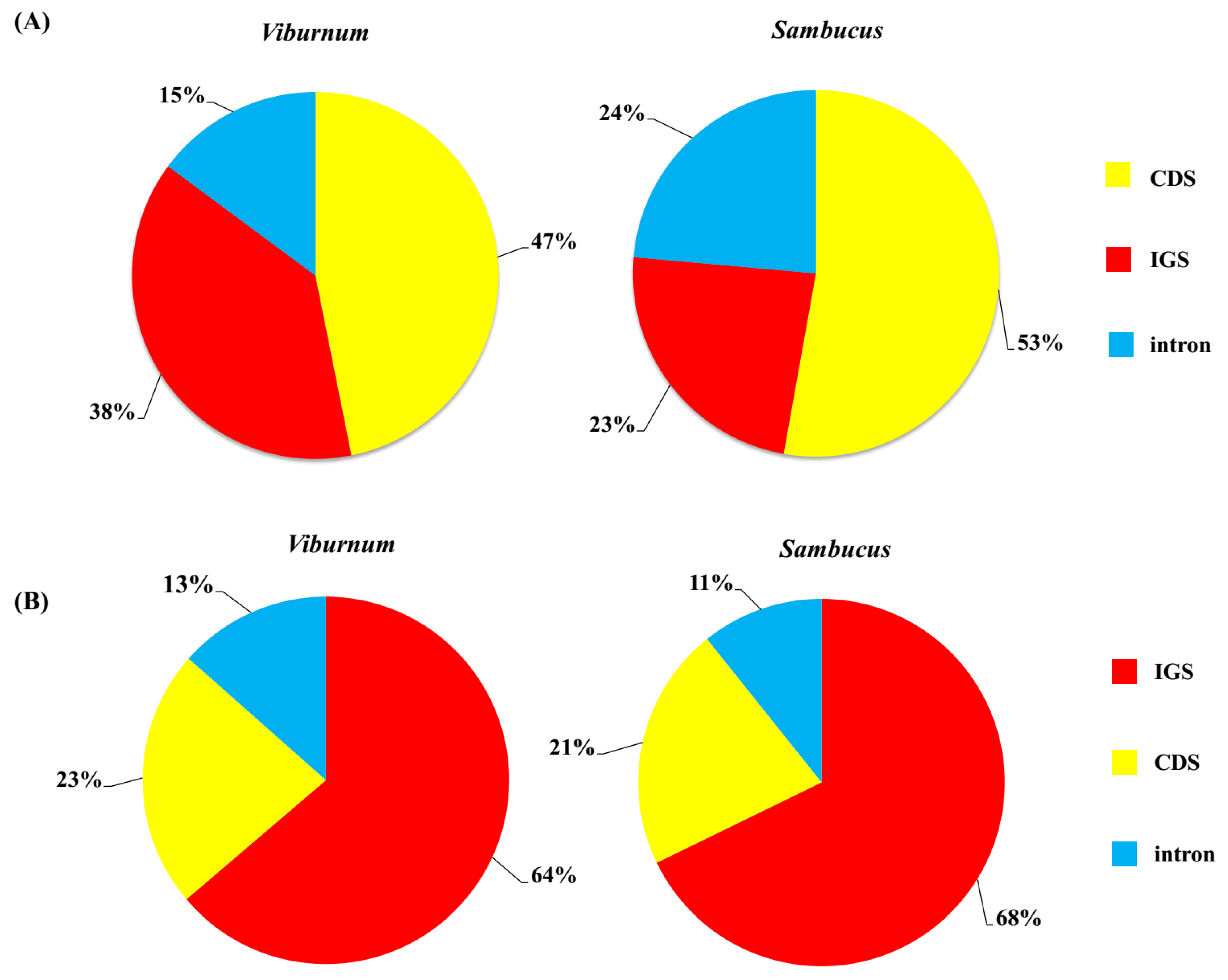
SSRs are abundantly distributed throughout the cp genome and have been widely used in species authentication and population genetics [32,33]. We found similar numbers and distribution pattern of SSR motifs among 21 and 3 accessions, respectively, in Viburnum and Sambucus (Figure 4B). The number of SSRs per plastome ranged from 47 (V. opulus) to 67 (V. henryi and V. nervosum) in Viburnum, and from 50 (S. adnata) to 64 (S. nigra) in Sambucus, with 119 SSRs being shared between all Viburnum plastomes and 149 in those of Sambucus. All the five kinds of SSRs (i.e., mono-, di-, tri-, tetra- and penta- nucleotide repeats) were detected in the 24 plastomes. By contrast, hexanucleotide repeats were only present in the 21 Viburnum species. Overall, mononucleotide SSR loci (A or T) were by far the most frequent type observed in both genera, potentially as a result of the bias toward A and T of cp genomes [34,35]. For both genera, SSRs were mainly situated in IGS (Viburnum: 63.77%; Sambucus: 67.79%; See Appendix A, Figure A3), and were also found in introns (Viburnum: 13.47%; Sambucus: 10.74%) and CDSs (Viburnum: 22.76%; Sambucus: 21.48%). These repeats will serve as useful resources for marker development for future studies on population genetics and phylogeography in Adoxaceae.
2.5. Phylogenetic Relationships
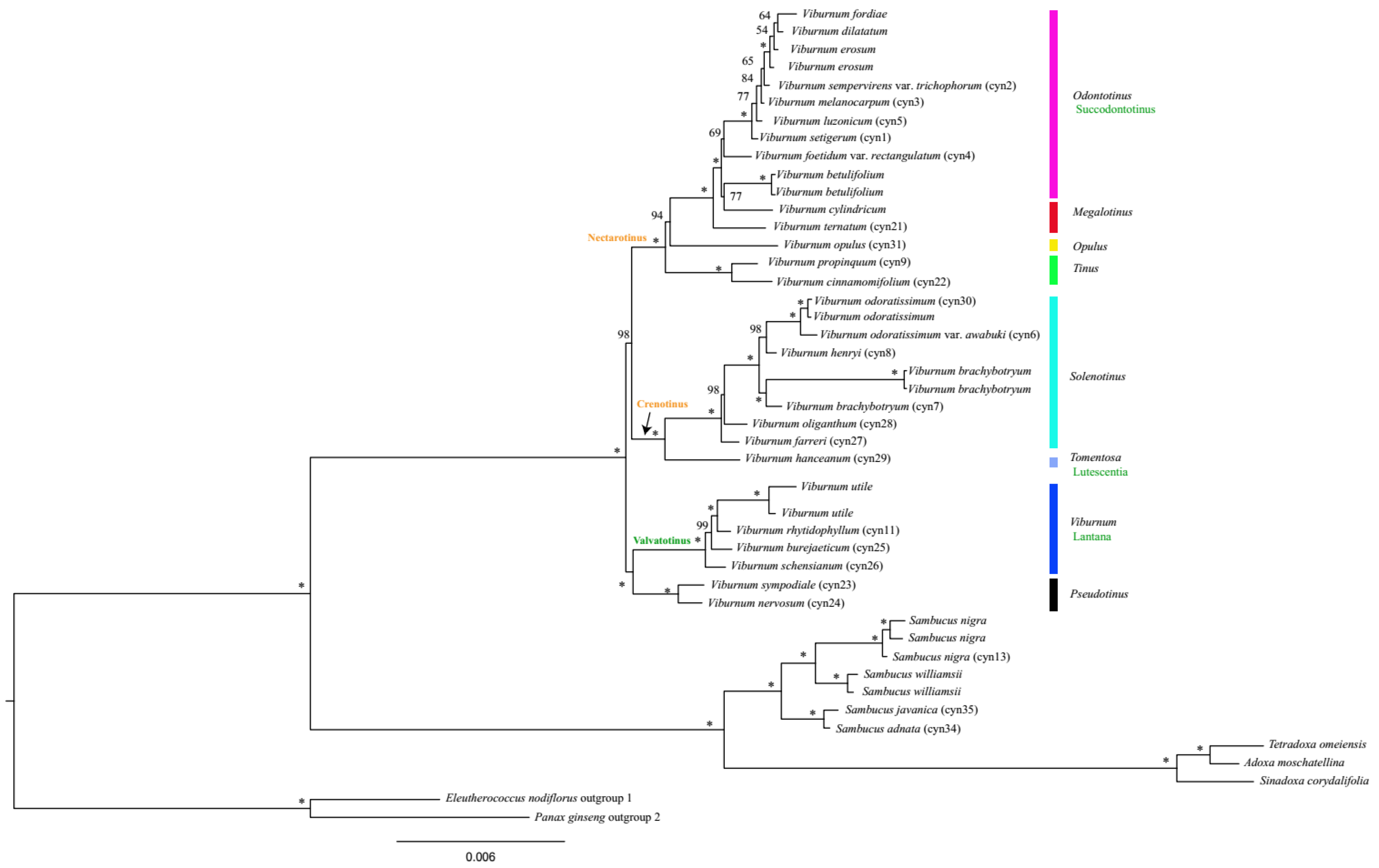
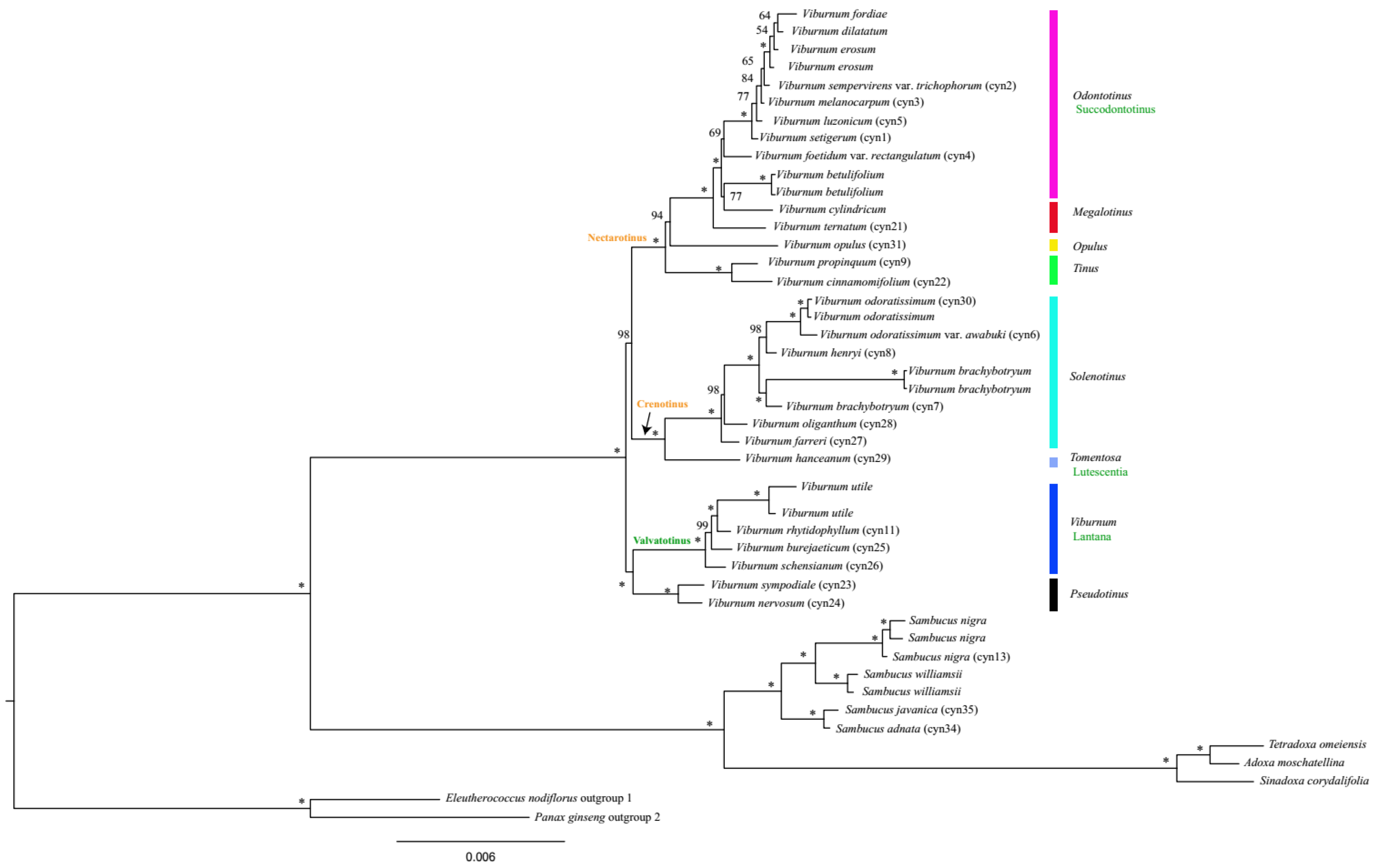
Based on the complete cp genome sequence dataset, two major clades were revealed, comprising a large clade and a small clade with 100% bootstrap support (Figure 5). The small clade included the genera Sambucus, Adoxa, Tetradoxa, and Sinadoxa, within which samples of Sambucus formed a monophyletic clade (bootstrap percentage, BS = 100%) and were sister to the Adoxa-Tetradoxa-Sinadoxa group. The large clade containing all Viburnum species was found to be monophyletic (BS = 100%) as well. Many relationships within this genus were well resolved, and the topology was almost identical to that of Clement et al. [19]. Thus, some clade names used here were taken from their study. Relationships at the base of the Viburnum clade were best represented by a dichotomy that included a group containing the Valvatotinus clade (represented here by Viburnum sect. Viburnum) and Viburnum sect. Pseudotinus, and a group containing all remaining Viburnum (Figure 5) [16,17]. In previous studies, the position of Viburnum sect. Pseudotinus was unstable. In some analyses, it (represented by V. cordifolium, V. furcatum and V. lantanoides) was sister to the clade with the remainder of Viburnum [15,16]; in other analyses, it (represented by V. furcatum, V. lantanoides, V. nervosum and V. sympodiale) appeared as sister to the Valvatotinus clade but with weak support [17,19]. However, in the present study, the sister relationship between the Valvatotinus clade and Viburnum sect. Pseudotinus was strongly supported (BS = 100%; Figure 5).
Two sister clades were clearly indicated (each 100%) within the clade that comprises all remaining Viburnum. The first clade Crenotinus, characterized by curving (crenate) leaf teeth [19], contained Viburnum sect. Tomentosa (represented here by V. hanceanum) and Viburnum sect. Solenotinus. Within the Crenotinus clade, our analysis confirmed the monophyly of the Solenotinus radiation (BS = 100%) and also the sister relationship between this section and Viburnum sect. Tomentosa. The second clade was Nectarotinus [19], which is characterized by extrafloral nectaries, containing the four traditionally recognized sections Viburnum sect. Odontotinus, Viburnum sect. Megalotinus, Viburnum sect. Tinus, and Viburnum sect. Opulus (represented here by V. opulus). Within this clade, consistent with the findings of Clement et al. [19], our analysis provided strong support for the placement of the monophyletic section Viburnum sect. Tinus as sister to the rest of the species (BS = 100%). One important difference between this result and that of Clement et al. [19] concerned the placement of Viburnum sect. Opulus. In line with previous studies [17,18], Viburnum sect. Opulus was recovered as sister to the clade containing sections Viburnum sect. Odontotinus and Viburnum sect. Megalotinus with confidence (BS = 94%). However, there was little support for this position based on the results of Clement et al. [19]. The two remaining sections, i.e., Viburnum sect. Odontotinus and Viburnum sect. Megalotinus, were clearly not monophyletic. This result was expected based on previous analyses [6,15,16,17,18,19,36]. The mostly red-fruited group of Viburnum sect. Odontotinus, namely Succodontotinus [16], was closely related to V. cylindricum of Viburnum sect. Megalotinus. V. ternatum (Viburnum sect. Megalotinus) was revealed to be sister to the polytomy consisting of the clade Succodontotinus plus V. cylindricum (BS = 100%).
In summary, four of the eight traditionally recognized sections in Flora of China were found to be monophyletic (i.e., Viburnum sect. Tinus, Viburnum sect. Solenotinus, Viburnum sect. Viburnum and Viburnum sect. Pseudotinus). The sections Viburnum sect. Odontotinus and Viburnum sect. Megalotinus were recovered as nonmonophyletic, which has been repeatedly shown in various molecular and morphological analyses [6,15,16,17,18,19,36]. Only a single representative was included in our analyses, for sections Viburnum sect. Tomentosa and Viburnum sect. Opulus. Additional sampling will be required to evaluate the monophyly of these groups. In addition, many relationships within our 45-species plastid tree were confidently resolved and the topology was identical to that of Clement et al. [19], with the exception of the relationships between the Valvatotinus clade and Viburnum sect. Pseudotinus and the position of V. opulus (Viburnum sect. Opulus). In the first case, there was strong support for the clade Valvatotinus being sister to Viburnum sect. Pseudotinus (100% bootstrap value; Figure 5). In the other case, as expected, V. opulus of Viburnum sect. Opulus was found to be sister to the clade comprising sections of Viburnum sect. Odontotinus and Viburnum sect. Megalotinus, which generally maintained its previously determined position in relation to Viburnum sect. Megalotinus and Viburnum sect. Odontotinus.
3. Materials and Methods
3.1. Sample Collection, Sequencing and Assembly
Fresh leaves from 21 species of Viburnum, representing all of the 8 sections recognized in Flora of China, together with 3 species of Sambucus, were sampled in China (Table S1) and dried in silica gel. The voucher specimens were deposited in College of Plant Protection, Henan Agricultural University (Table S1). Total genomic DNA of the 24 species was extracted and then sequenced on an Illumina Hiseq2500 Platform at Jinweizhi Biotechnology Institute (Suzhou, China).
We used a combination of de novo and reference-guided methods to assemble these plastomes [37]. Firstly, for each Viburnum and Sambucus species, raw paired-end reads were trimmed to remove low-quality reads with a Phred value < 20 using CLC Genomics Workbench v10.1.1 (CLC Bio, Aarhus, Denmark; http://www.clcbio.com). Secondly, the remaining clean reads were assembled into contigs on the CLC assembler with the following settings: bubble size, 98; minimum contig length, 250 bp; mismatch cost, 2; deletion and insertion costs, 3; length fraction, 0.9; and similarity fraction, 0.8. Thirdly, due to the fact that the original sequences represented a mixture of both nuclear and organellar DNA, to filter the plastid-like ones, all contigs of Viburnum and Sambucus were aligned to the reference genomes Viburnum betulifolium (GenBank accession number: NC_037951) and Sambucus williamsii (GenBank accession number: NC_033878), respectively, using BLAST (http://blast.ncbi.nlm.nih.gov/). Then, the filtered contigs longer than 10 kb were oriented and realigned with the reference genomes for constructing the draft chloroplast genome of each species with GENEIOUS V11.01 software (http: //www.geneious.com). Finally, the ordered contigs were remapped to the draft genome to generate the complete chloroplast genome sequences. To validate the assembly, PCR amplifications and Sanger sequencing were performed to confirm the four junction regions between IRs and LSC/SSC with primers developed from assembled sequences flanking the junction regions (Table S2).
3.2. Whole Chloroplast Genome Annotation and Comparison
The whole chloroplast genomes were annotated using GENEIOUS V11.01 and DOGMA [38]. The start/stop codons and intron/exon boundaries of genes were checked and adjusted manually according to the reference genomes. In addition, the tRNA boundaries were further verified by tRNAscan-SE v1.21 [39] with default settings. Online program OrganellarGenome DRAW (https://chlorobox.mpimp-golm.mpg.de/OGDraw.html) [40] was used to draw the gene maps of Viburnum and Sambucus cp genomes. Finally, the 24 annotated plastome sequences were deposited in GenBank.
Chloroplast genome comparisons across the 21 Viburnum and 3 Sambucus species were conducted on the mVISTA tool (genome.lbl.gov/vista/index.shtml) [41] using Shuffle-LAGAN mode, with the annotations of V. betulifolium and S. williamsii serving as references, respectively. In order to identify the variant hotspot regions for Viburnum and Sambucus, the sequence alignments of their respective plastomes were subjected to a sliding window analysis in DNASP v5.10 [42] to estimate the nucleotide variability (Pi) for all the protein coding and noncoding regions (i.e., IGSs and introns).
3.3. Identification of Repeat Sequences and SSRs
The whole cp genomes of Viburnum and Sambucus were aligned in GENEIOUS v11.1.4 using MAFFT multiple aligner v7 [43], respectively. Then, chloroplast SSR loci (i.e., mono-, di-, tri-, tetra-, penta- and hexa- nucleotide repeats) were identified using Perl script MISA (http://pgrc.ipk-gatersleben.de/misa/misa.html) with minimal repeat numbers of 10, 5, 4, 3, 3 and 3 for mononucleotide, dinucleotide, trinucleotide, tetranucleotide, pentanucleotide and hexanucleotide repeats, respectively. Moreover, the program REPUTER [44] was used to estimate the number and position of repeat elements, including direct (forward), inverted (palindromic), complement and reverse repeats. The constraints to all the four repeat types in REPUTER were 1) a minimum repeat size of 30 bp; and 2) 90% higher sequence identity with a hamming distance of 3 (i.e., the maximum length of the gap size between repeats equals 3 bp).
3.4. Phylogenetic Analysis
We used 45 cp genomes to infer the phylogenetic relationships among Adoxaceae species, including 24 newly obtained plastomes, 19 plastomes downloaded from the GenBank (i.e., 6 plastomes of Viburnum sect. Odontotinus, 1 plastome of Viburnum sect. Megalotinus, 3 plastomes of Viburnum sect. Solenotinus, 2 plastomes of Viburnum sect. Viburnum, plus 7 representatives of Sambucus, Adoxa, Sinadoxa, and Tetradoxa) and two outgroups, Panax ginseng and Eleutherococcus nodiflorus (Table S3). The phylogenetic analysis was performed with a maximum-likelihood (ML) method based on the complete cp genome sequence dataset. Chloroplast sequences of these 45 species were aligned together using MAFFT with default settings. ML analysis was conducted in RAXML-HPC [45] on the CIPRES cluster (http://www.phylo.org/), with a GTR+G+I substitution model selected by jModelTest v2.1.7 [46] and an unpartitioned strategy.
4. Conclusions
This work presents a major advance in understanding Chinese Adoxaceae phylogenetics and plastome evolution with a particular focus on the genus Viburnum. The comparison of the plastomes among each species of Viburnum and Sambucus, and with those of other members of Adoxaceae, revealed high similarities with respect to genomic structure, gene order and content. Repeat sequences, SSRs and highly variable regions were identified with the purpose of developing potential molecular markers for future studies on the population genetics, phylogeny and phylogeography of Viburnum and Sambucus. Our phylogenomic analysis, based on the complete cp genome sequence dataset, strongly supported the relationships within Adoxaceae and Viburnum revealed by previous plastid phylogenomic investigations [12,19,25,26]. Viburnum was shown to be a sister to the Sambucus-Adoxa-Tetradoxa-Sinadoxa group. Within Viburnum, the monophyly of four traditionally recognized sections in Flora of China (i.e., Viburnum sect. Tinus, Viburnum sect. Solenotinus, Viburnum sect. Viburnum and Viburnum sect. Pseudotinus) was strongly supported. The nonmonophyly of sections Viburnum sect. Odontotinus and Viburnum sect. Megalotinus was repeatedly demonstrated. Additionally, our analyses confirmed the sister relationships between the clade Valvatotinus and Viburnum sect. Pseudotinus, as well as between Viburnum sect. Opulus and the Odontotinus-Megalotinus group. Overall, our results clearly exhibited the power of the complete cp genomes to improve phylogenetic resolution, and will contribute to a better understanding of plastome evolution in Chinese Adoxaceae.
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/2223-7747/9/9/1143/s1. Table S1. Sampling, assembly and voucher information for the 24 Adoxaceae species in the present study. Table S2. Information of the specific primer pairs used to verify the four junctions between IRs and SSC/LSC for Viburnum and Sambucus, respectively. Table S3. Summary of the GenBank accession numbers and genome sizes for previously published Adoxaceae species and the outgroups used in the present study.
Author Contributions
Y.C. conceived of the idea. Y.C., Y.L. and H.R. contributed to the sampling. Y.C., C.W. and H.R. analyzed the data. The manuscript was written by Y.C. and H.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 31800179).
Acknowledgments
The authors thank Tian-lei Xie from Central South University of Forestry and Technology, Jia-mei Li from Henan Agricultural University, and Min Chen from Zhongshan Botanical Garden for collecting plant materials in China; Shen-yi Wang (University of Wisconsin-Madison, USA) for insightful comments on an earlier version of this manuscript.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Appendix A
Figure A1.
Sequence identity plots among 24 Adoxaceae chloroplast genomes.
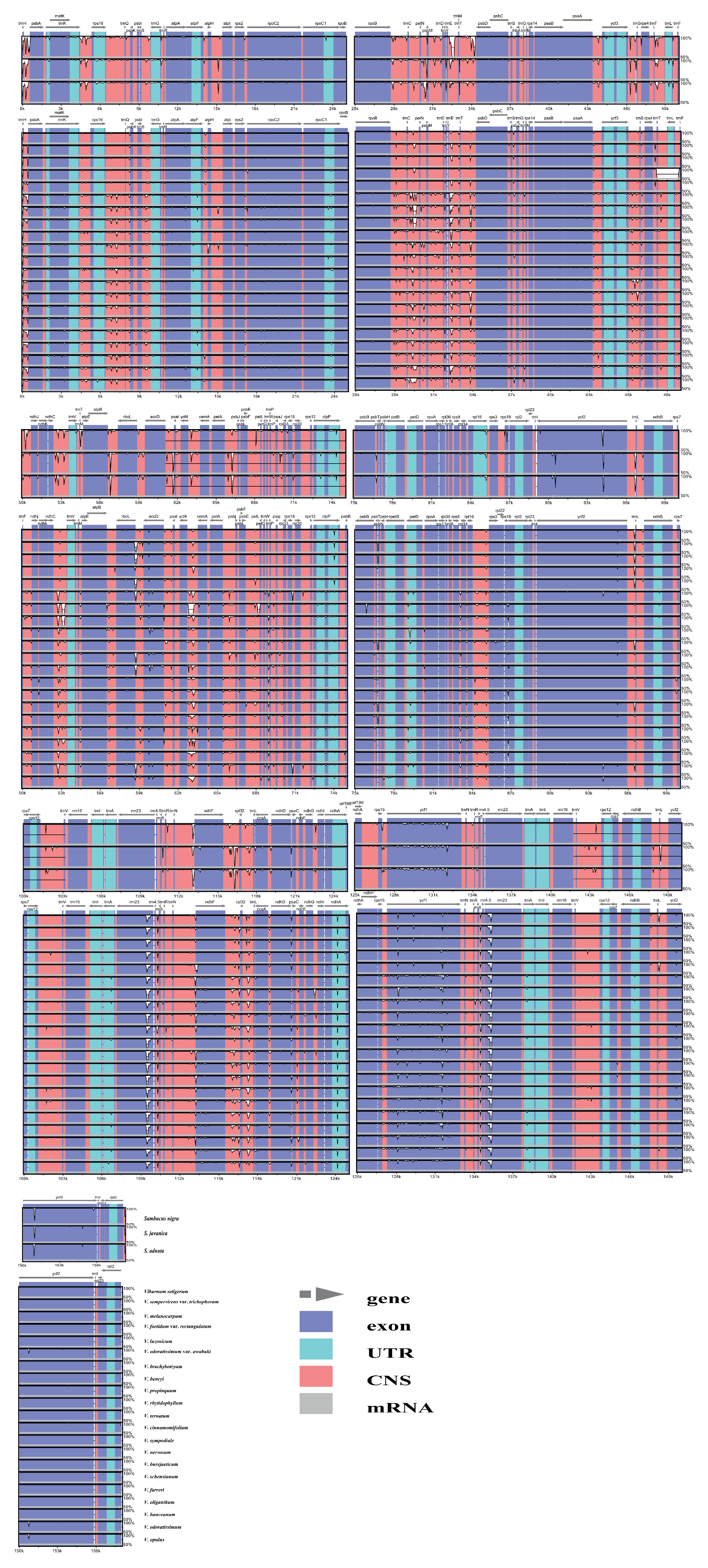
Figure A2.
Compositions of the repeats in 24 Adoxaceae species. Repeats with different lengths are indicated in different colors.
Figure A2.
Compositions of the repeats in 24 Adoxaceae species. Repeats with different lengths are indicated in different colors.
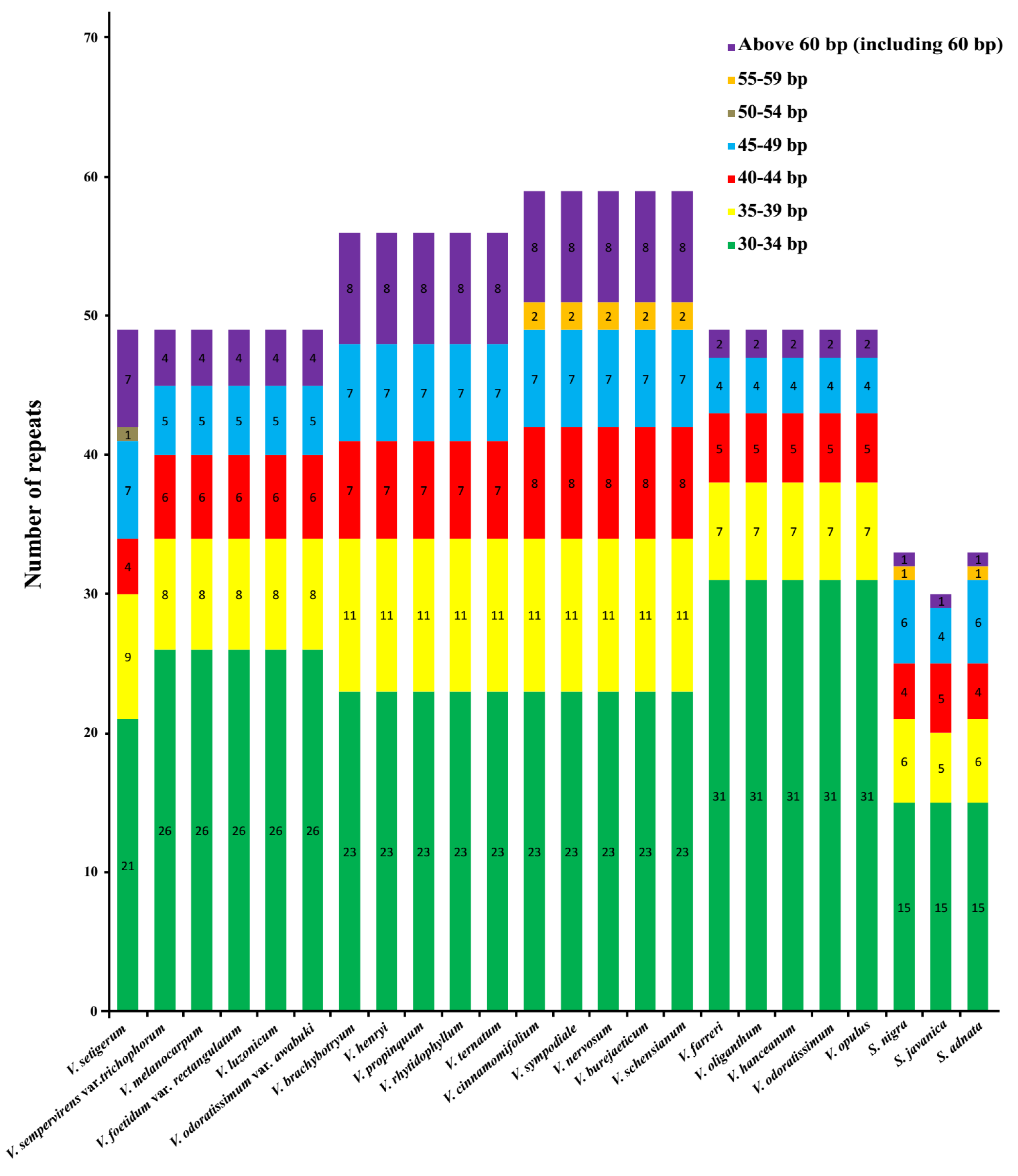
Figure A3.
(A) The distribution frequency of repeats in chloroplast genomes. (B) Distribution of SSRs sites in chloroplast genomes. IGS: intergenic spacer region; CDS: protein-coding sequences.
Figure A3.
(A) The distribution frequency of repeats in chloroplast genomes. (B) Distribution of SSRs sites in chloroplast genomes. IGS: intergenic spacer region; CDS: protein-coding sequences.
References
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.E.; Malecot, V. Viburnum. In Flora of China, Lentibulariaceae Through Dipsacaceae; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2011; Volume 19, pp. 570–611. [Google Scholar]
- Dirr, M.A. Viburnums: Flowering Shrubs for Every Season; Timber Press, Inc.: Portland, OR, USA, 2007. [Google Scholar]
- Rehder, A. The Viburnums of Eastern Asia. In Trees and Shrubs; Part, II; Sargent, C.S., Ed.; Houghton Mifflin: Boston, MA, USA, 1908; Volume II, pp. 105–116. [Google Scholar]
- Nicolson, D.H.; Hara, H. A Revision of Caprifoliaceae of Japan with Reference to Allied Plants in other Districts and the Adoxaceae; Academia Scientific Books, Inc.: Tokyo, Japan, 1983. [Google Scholar]
- Donoghue, M. A preliminary analysis of phylogenetic relationships in Viburnum (Caprifoliaceae s.1.). Syst. Bot. 1983, 8, 45–58. [Google Scholar] [CrossRef]
- Kern, J.H. The genus Viburnum (Caprifoliaceae) in Malaysia. Reinwardtia 1951, 1, 107–170. [Google Scholar]
- Killip, E.P.; Smith, A.C. The south American species of Viburnum. Bull. Torrey Bot. Club 1931, 57, 245–258. [Google Scholar] [CrossRef]
- Donoghue, M. The Phylogenetic Relationships of Viburnum. In Advances in Cladistics; Platnick, N., Funk, V., Eds.; Columbia University Press: New York, NY, USA, 1983; Volume 2, pp. 143–166. [Google Scholar]
- Eriksson, T.; Donoghue, M.J. Phylogenetic relationships of Sambucus and Adoxa (Adoxoideae, Adoxaceae) based on nuclear ribosomal ITS sequences and preliminary morphological data. Syst. Bot. 1997, 22, 555–573. [Google Scholar] [CrossRef]
- Cao, M.Y.; Gao, A.; Gong, J.; Xia, L.; Jia, X.; Li, N.; Hou, X.Y.; Lu, F.; Ni, S.F. Overview of Pharmaceutical Research on Sambucus L. Ningxia J. Agric. For. Sci. Tech. 2011, 52, 59–60. [Google Scholar]
- Fan, W.B.; Wu, Y.; Yang, J.; Shahzad, K.; Li, Z.H. Comparative chloroplast genomics of Dipsacales species: Insights into sequence variation, adaptive evolution, and phylogenetic relationships. Front. Plant Sci. 2018, 9, 689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oersted, A.S. Til belysning af slaegten Viburnum. Vidensk. Medd. Naturhist. Foren. Kjobenhavn 1861, 13, 267–305. [Google Scholar]
- Donoghue, M.J.; Baldwin, B.G.; Li, J.; Winkworth, R.C. Viburnum phylogeny based on chloroplast trnK intron and nuclear ribosomal ITS DNA sequences. Syst. Bot. 2004, 29, 188–198. [Google Scholar] [CrossRef]
- Winkworth, R.C.; Donoghue, M.J. Viburnum phylogeny: Evidence from the duplicated nuclear gene GBSSI. Mol. Phylogenet. Evol. 2004, 33, 109–126. [Google Scholar] [CrossRef]
- Winkworth, R.C.; Donoghue, M.J. Viburnum phylogeny based on combined molecular data: Implications for taxonomy and biogeography. Am. J. Bot. 2005, 92, 653–666. [Google Scholar] [CrossRef] [PubMed]
- Clement, W.L.; Donoghue, M.J. Dissolution of Viburnum section Megalotinus (Adoxaceae) of Southeast Asia and its implications for morphological evolution and biogeography. Int. J. Plant Sci. 2011, 172, 559–573. [Google Scholar] [CrossRef] [Green Version]
- Clement, W.L.; Donoghue, M.J. Barcoding success as a function of phylogenetic relatedness in Viburnum, a clade of woody angiosperms. BMC Evol. Biol. 2012, 12, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, W.L.; Arakaki, M.; Sweeney, P.W.; Edwards, E.J.; Donoghue, M.J. A chloroplast tree for Viburnum (Adoxaceae) and its implications for phylogenetic classification and character evolution. Am. J. Bot. 2014, 101, 1029–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.B.; Yang, S.X.; Li, H.T.; Yang, J.; Li, D.Z. Comparative Chloroplast Genomes of Camellia Species. PLoS ONE 2013, 8, e73053. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Yao, X.; Tan, Y.H.; Gan, Y.; Corlett, R.T. Complete chloroplast genome sequence of the avocado: Gene organization, comparative analysis, and phylogenetic relationships with other Lauraceae. Can. J. For. Res. 2016, 46, 1293–1301. [Google Scholar] [CrossRef]
- Wang, S.; Yang, C.; Zhao, X.; Chen, S.; Qu, G. Complete chloroplast genome sequence of Betula platyphylla: Gene organization, RNA editing, and comparative and phylogenetic analyses. BMC Genom. 2018, 19, 950. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.; Zhao, X.; Zhou, J.; Huo, Y.; Ding, Y.; Yuan, Z. The complete chloroplast genomes of Punica granatum and a comparison with other species in Lythraceae. Int. J. Mol. Sci. 2019, 20, 2886. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.B.; Han, E.K.; Choi, H.J.; Lee, J.H. The complete chloroplast genome sequence of Viburnum japonicum (Adoxaceae), an evergreen broad-leaved shrub. Mitochondrial DNA Part B Resour. 2018, 3, 458–459. [Google Scholar] [CrossRef] [Green Version]
- Xiang, C.L.; Dong, H.J.; Landrein, S.; Zhao, F.; Yu, W.B.; Soltis, D.E.; Soltis, P.S.; Backlund, A.; Wang, H.F.; Li, D.Z.; et al. Revisiting the phylogeny of Dipsacales: New insights from phylogenomic analyses of complete plastomic sequences. J. Syst. Evol. 2020, 58, 103–117. [Google Scholar] [CrossRef]
- Wang, H.X.; Liu, H.; Moore, M.J.; Landrein, S.; Liu, B.; Zhu, Z.X.; Wang, H.F. Plastid phylogenomic insights into the evolution of the Caprifoliaceae s.l. (Dipsacales). Mol. Phylogenet. Evol. 2020, 142, 106641. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.H.; Tang, P.; Li, Z.Z.; Li, D.W.; Liu, Y.F.; Huang, H.W. The first complete chloroplast genome sequences in Actinidiaceae: Genome structure and comparative analysis. PLoS ONE 2015, 10, e0129347. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Park, K.T.; Park, S. The chloroplast genome of Symplocarpus renifolius: A comparison of chloroplast genome structure in Araceae. Genes 2017, 8, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Liu, J.; Luo, L.; Wei, X.; Zhang, J.; Qi, Y.; Zhang, B.; Liu, H.; Xiao, P. Complete chloroplast genome sequences of Schisandra chinensis: Genome structure, comparative analysis, and phylogenetic relationship of basal angiosperms. Sci. China Life Sci. 2017, 60, 1286–1290. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T. Chloroplast evolution: Secondary symbiogenesis and multiple losses. Curr. Biol. 2002, 12, R62–R64. [Google Scholar] [CrossRef] [Green Version]
- Nie, X.; Lv, S.; Zhang, Y.; Du, X.; Wang, L.; Biradar, S.S.; Tan, X.; Wan, F.; Weining, S. Complete chloroplast genome sequence of a major invasive species, crofton weed (Ageratina adenophora). PLoS ONE 2012, 7, e36869. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.H.; Zhang, J.J.; Yao, X.H.; Huang, H.W. Chloroplast microsatellite markers in Liriodendron tulipifera (Magnoliaceae) and cross-species amplification in L. chinense. Am. J. Bot. 2011, 98, 123–126. [Google Scholar] [CrossRef]
- Dong, W.; Xu, C.; Cheng, T.; Lin, K.; Zhou, S. Sequencing angiosperm plastid genomes made easy: A complete set of universal primers and a case study on the phylogeny of Saxifragales. Genome Biol. Evol. 2013, 5, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Li, Q.; Hu, Z.; Li, X.; Chen, S. The complete Amomum kravanh chloroplast genome sequence and phylogenetic analysis of the Commelinids. Molecules 2017, 22, 1875. [Google Scholar] [CrossRef] [Green Version]
- Ye, W.Q.; Yap, Z.Y.; Li, P.; Comes, H.P.; Qiu, Y.X. Plastome organization, genome-based phylogeny and evolution of plastid genes in Podophylloideae (Berberidaceae). Mol. Phylogenet. Evol. 2018, 127, 978–987. [Google Scholar] [CrossRef]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F.; Campbell, C.S.; Donoghue, M.J. The ITS region of nuclear ribosomal DNA: A valuable source of evidence on angiosperm phylogeny. Ann. Mo. Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- Cronn, R.; Liston, A.; Parks, M.; Gernandt, D.S.; Shen, R.; Mockler, T. Multiplex sequencing of plant chloroplast genomes using Solexa sequencing-by-synthesis technology. Nucleic Acids Res. 2008, 36, e122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res. 2005, 33, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Kahlau, S.; Bock, R. OrganellarGenomeDRAW-a suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Res. 2013, 41, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. Reputer: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Chloroplast genome maps for (A) 21 Viburnum species and (B) 3 Sambucus species.
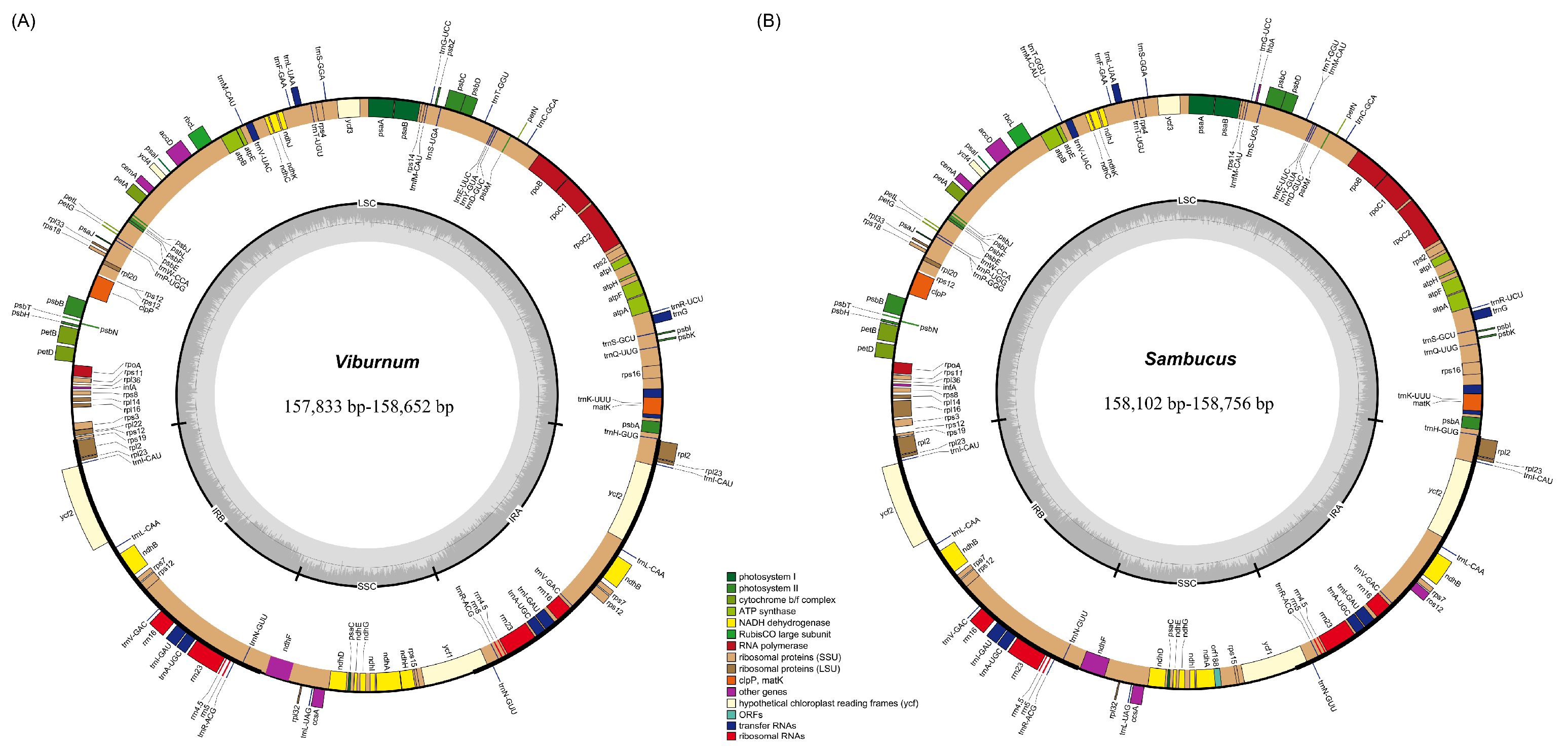
Figure 2.
Comparison of the junctions between IRs and SSC/LSC regions for the 24 Adoxaceae species.

Figure 3.
Percentages of variable characteristics in homologous regions among the chloroplast genomes of 24 Adoxaceae species. (A) Pi values among CDSs. (B) Pi values of intergenic spacer (IGS) regions and introns.
Figure 3.
Percentages of variable characteristics in homologous regions among the chloroplast genomes of 24 Adoxaceae species. (A) Pi values among CDSs. (B) Pi values of intergenic spacer (IGS) regions and introns.
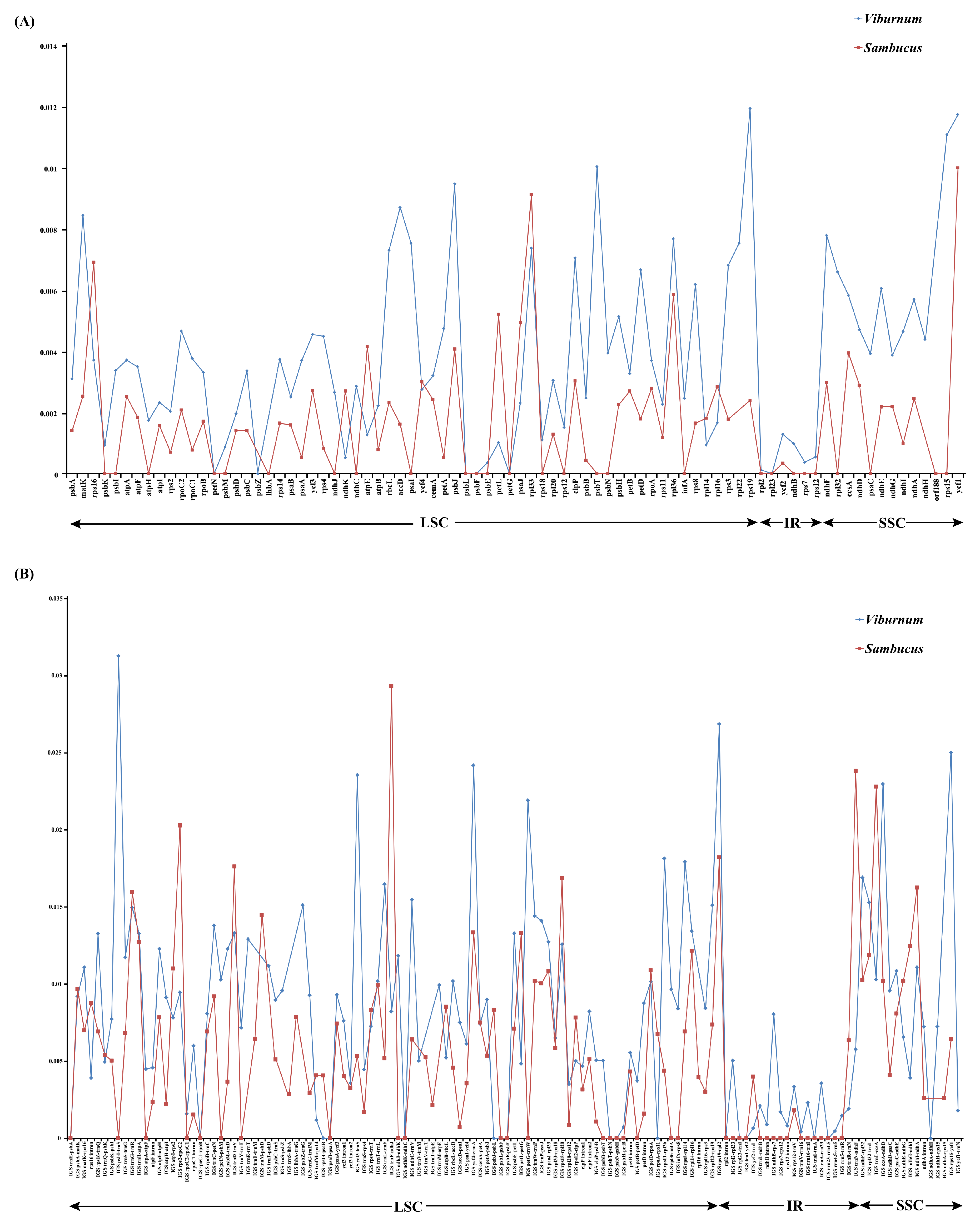
Figure 4.
The distribution of repeats and SSRs in the chloroplast genomes of 24 Adoxaceae species. (A) Frequency of repeat types in 24 Adoxaceae species. F, P and C indicate the forward, palindrome and complement repeat types, respectively. (B) Compositions of the SSR in 24 Adoxaceae species. Different SSR motifs are shown in different colors.
Figure 4.
The distribution of repeats and SSRs in the chloroplast genomes of 24 Adoxaceae species. (A) Frequency of repeat types in 24 Adoxaceae species. F, P and C indicate the forward, palindrome and complement repeat types, respectively. (B) Compositions of the SSR in 24 Adoxaceae species. Different SSR motifs are shown in different colors.
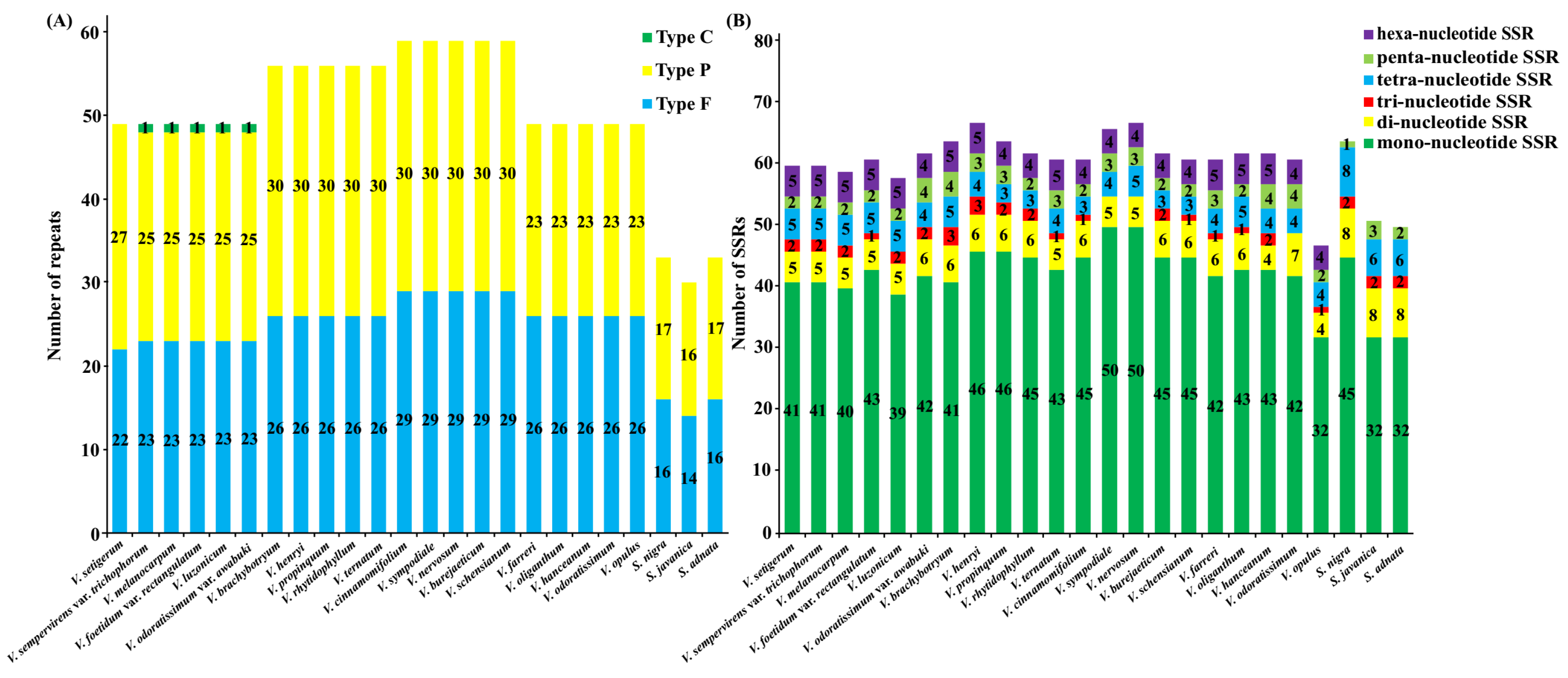
Figure 5.
Phylogenetic relationships among Viburnum and Sambucus species inferred from the complete cp genome sequence dataset based on maximum likelihood (ML) method. Numbers above the nodes represent ML bootstrap values, and “*” indicates 100% support values in ML. Clade names (in the right) marked in black represent previously published names under the ICN (International Code of Nomenclature) that are here converted to phylogenetic names. Those in green (in the right and along a branch) represent names proposed by Winkworth and Donoghue [16] and Clement and Donoghue [17]. Clade names in orange (along a branch) represent names proposed by Clement et al. [19].
Figure 5.
Phylogenetic relationships among Viburnum and Sambucus species inferred from the complete cp genome sequence dataset based on maximum likelihood (ML) method. Numbers above the nodes represent ML bootstrap values, and “*” indicates 100% support values in ML. Clade names (in the right) marked in black represent previously published names under the ICN (International Code of Nomenclature) that are here converted to phylogenetic names. Those in green (in the right and along a branch) represent names proposed by Winkworth and Donoghue [16] and Clement and Donoghue [17]. Clade names in orange (along a branch) represent names proposed by Clement et al. [19].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the main characteristics of Adoxaceae plastomes.
| Species | Genome Size (bp) | LSC Length (bp) | SSC Length (bp) | IR Length (bp) | Total GC Content (%) | Number of Genes | |||
|---|---|---|---|---|---|---|---|---|---|
| Total | CDS | rRNAs | tRNAs | ||||||
| V. setigerum | 158,306 | 86,763 | 18,539 | 26,502 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. sempervirens var. trichophorum | 158,184 | 86,710 | 18,472 | 26,501 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. melanocarpum | 158,196 | 86,695 | 18,497 | 26,502 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. foetidum var. rectangulatum | 158,230 | 86,835 | 18,431 | 26,482 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. luzonicum | 158,652 | 87,892 | 17,674 | 26,543 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. odoratissimum var. awabuki | 158,126 | 86,718 | 18,438 | 26,485 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. brachybotryum | 157,833 | 86,809 | 18,268 | 26,378 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. henryi | 157,862 | 86,430 | 18,452 | 26,490 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. propinquum | 157,987 | 86,839 | 18,350 | 26,399 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. rhytidophyllum | 158,520 | 87,054 | 18,338 | 26,564 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. ternatum | 158,344 | 87,109 | 18,407 | 26,414 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. cinnamomifolium | 158,347 | 87,210 | 18,347 | 26,395 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. sympodiale | 158,238 | 87,118 | 18,330 | 26,395 | 38.0% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. nervosum | 157,890 | 86,715 | 18,341 | 26,417 | 38.0% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. burejaeticum | 157,913 | 86,669 | 18,274 | 26,485 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. schensianum | 157,924 | 86,681 | 18,289 | 26,477 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. farreri | 158,046 | 86,809 | 18,401 | 26,418 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. oliganthum | 158,309 | 87,038 | 18,453 | 26,409 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. hanceanum | 158,195 | 86,815 | 18,436 | 26,472 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. odoratissimum | 158,020 | 86,653 | 18,419 | 26,474 | 38.1% | 130 | 85 (6) | 8 (4) | 37 (7) |
| V. opulus | 158,520 | 87,114 | 18,456 | 26,475 | 38.2% | 130 | 85 (6) | 8 (4) | 37 (7) |
| S. nigra | 158,102 | 86,518 | 18,978 | 26,303 | 38.0% | 132 | 84 (6) | 8 (4) | 40 (7) |
| S. javanica | 158,624 | 87,226 | 18,854 | 26,272 | 38.0% | 132 | 84 (6) | 8 (4) | 40 (7) |
| S. adnata | 158,756 | 87,328 | 18,862 | 26,283 | 38.0% | 132 | 84 (6) | 8 (4) | 40 (7) |
Numbers in brackets indicate the numbers of genes duplicated in the IR regions.
Table 2.
Gene composition in the 24 Adoxaceae chloroplast genomes.
| Gene Group | Gene Name |
|---|---|
| Ribosomal RNAs | rrn16(×2), rrn23(×2), rrn4.5(×2), rrn5(×2) |
| Transfer RNAs | trnH-GUG, trnK-UUUa, trnQ-UUG, trnS-GCU, trnGa, trnR-UCU |
| trnC-GCA, trnD-GUC, trnY-GUA, trnE-UUC, trnT-GGU | |
| trnS-UGA, trnG-UCC, trnfM-CAU, trnS-GGA, trnT-UGU | |
| trnL-UAAa, trnF-GAA, trnV-UACa, trnM-CAU, trnW-CCA | |
| trnP-UGG, trnl-CAU(×2), trnL-CAA(×2), trnV-GAC(×2) | |
| trnl-GAUa(×2), trnA-UGCa(×2), trnR-ACG(×2), trnN-GUU(×2) | |
| trnL-UAG, trnM-CAUx, trnT-GGUx, trnP-GGGx | |
| Photosystem I | psaB, psaA, psal, psaJ, psaC |
| Photosystem II | psbA, psbK, psbl, psbM, psbD, psbC, psbZz, psbB, psbT, |
| psbL, psbF, psbE, psbH, psbN, psbJ | |
| Cytochrome | petN, petA, petL, petG, petBa, petDa |
| ATP synthase | atpA, atpFa, atpH, atpl, atpE, atpB |
| Rubisco | rbcL |
| NADH dehydrogenease | ndhJ, ndhK, ndhC, ndhBa(×2), ndhD, ndhE, ndhG |
| ndhl, ndhAa, ndhHz | |
| Ribosomal proteins (large units) | rpl33, rpl20, rpl36, rpl14, rpl16, rpl16a, rpl22z, rpl2a(×2), |
| rpl23(×2), rpl32 | |
| Ribosomal proteins (small units) | rps16a, rps2, rps14, rps4, rps18, rps12b (×2), rps11, rps8, |
| rps7(×2), rps15, rps3, rps19 | |
| RNA polymerase | rpoC2, rpoC1a, rpoB, rpoA |
| Miscellaneous proteins & ATP-dependent protease subunit P | matK, clpPb |
| Other genes | accD, cemA, infA, ccsA, orf188x, lhbAx |
| Hypothetical proteins & Conserved reading frame | ycf3b, ycf4, ycf2(×2), ycf1Ψ |
a Indicates the genes containing a single intron. b Indicates the genes containing two introns. x Indicates the gene is present only in Sambucus. z Indicates the gene is present only in Viburnum. (×2) indicates genes duplicated in the IR regions; pseudogene is represented by Ψ. ycf1 is a pseudogene only in the following 16 species: V. setigerum, V. foetidum var. rectangulatum, V. luzonicum, V. odoratissimum var. awabuki, V. brachybotryum, V. rhytidophyllum, V. sympodiale, V. nervosum, V. burejaeticum, V. schensianum, V. farreri, V. oliganthum, V. hanceanum, S. nigra, S. javanica, S. adnata.
Table 3.
Pi values of the ten most variable coding and noncoding regions in Viburnum and Sambucus.
| Viburnum | Pi | Sambucus | Pi |
|---|---|---|---|
| noncoding regions | |||
| rps15-ycf1 | 0.02502 | trnF-ndhJ | 0.02934 |
| ycf4-cemA | 0.02419 | trnN-ndhF | 0.02384 |
| ycf3-trnS | 0.02356 | rps2-rpoC2 | 0.02029 |
| ccsA-ndhD | 0.02298 | rps18-rpl20 | 0.01687 |
| rps8-rpl14 | 0.01794 | trnG-trnR | 0.01596 |
| ndhF-rpl32 | 0.01691 | trnM-psbD | 0.01446 |
| trnL-trnF | 0.01647 | ycf4-cemA | 0.01335 |
| ndhC-trnV | 0.01548 | ndhG-ndhI | 0.01247 |
| rpl32-trnL | 0.01529 | rpl32-trnL | 0.01188 |
| psbZ-trnG | 0.01512 | atpI-rps2 | 0.01101 |
| coding regions | |||
| rps19 | 0.01193 | ycf1 | 0.00999 |
| ycf1 | 0.01173 | rpl33 | 0.00912 |
| rps15 | 0.01107 | rps16 | 0.00691 |
| accD | 0.00870 | atpE | 0.00415 |
| matK | 0.00844 | ccsA | 0.00394 |
| ndhF | 0.00779 | clpP | 0.00303 |
| rpl22 | 0.00753 | ycf4 | 0.00300 |
| rpl33 | 0.00737 | ndhF | 0.00298 |
| rbcL | 0.00730 | ndhD | 0.00288 |
| clpP | 0.00705 | rpl16 | 0.00285 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ran, H.; Liu, Y.; Wu, C.; Cao, Y. Phylogenetic and Comparative Analyses of Complete Chloroplast Genomes of Chinese Viburnum and Sambucus (Adoxaceae). Plants 2020, 9, 1143. https://doi.org/10.3390/plants9091143
AMA Style
Ran H, Liu Y, Wu C, Cao Y. Phylogenetic and Comparative Analyses of Complete Chloroplast Genomes of Chinese Viburnum and Sambucus (Adoxaceae). Plants. 2020; 9(9):1143. https://doi.org/10.3390/plants9091143
Chicago/Turabian StyleRan, Hang, Yanyan Liu, Cui Wu, and Yanan Cao. 2020. "Phylogenetic and Comparative Analyses of Complete Chloroplast Genomes of Chinese Viburnum and Sambucus (Adoxaceae)" Plants 9, no. 9: 1143. https://doi.org/10.3390/plants9091143
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.