Time of In Vitro Anther Culture May Moderate Action of Copper and Silver Ions that Affect the Relationship between DNA Methylation Change and the Yield of Barley Green Regenerants


Abstract
:1. Introduction
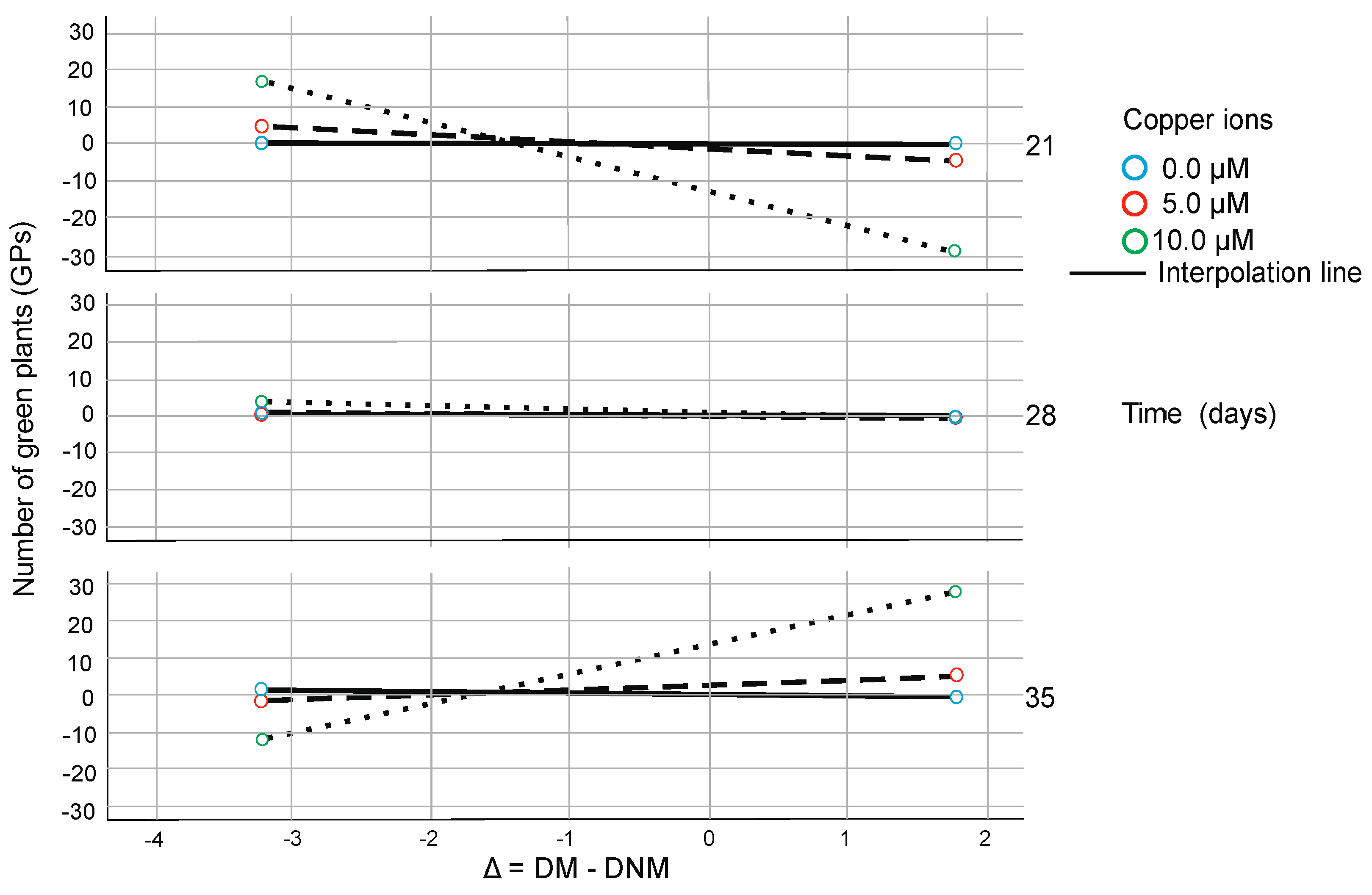
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material
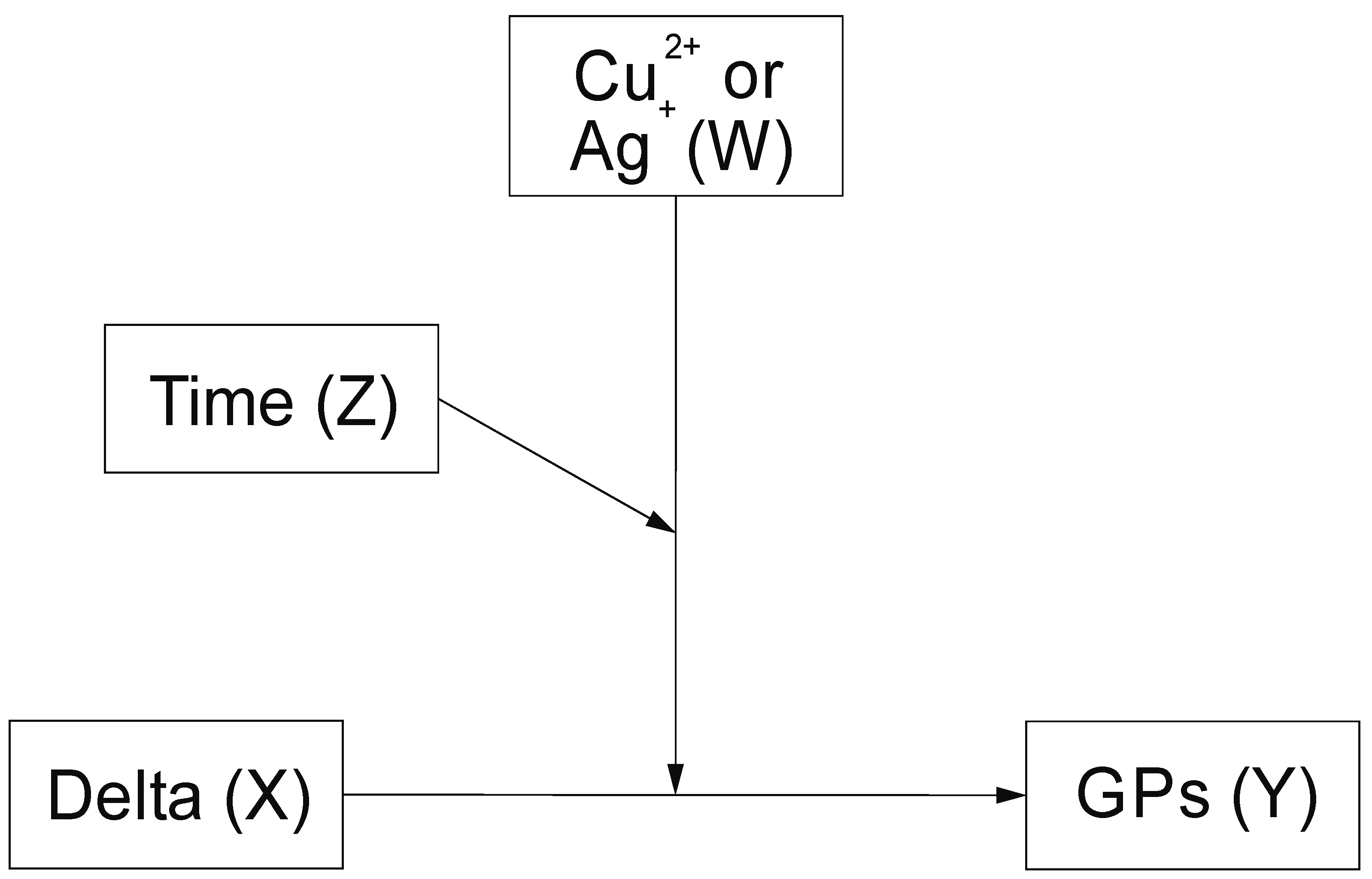
4.2. Moderated Moderation Analysis
4.3. Analysis of Variance
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Delta | change between DM and DNM, Delta = DM - DNM |
| DArT | Diversity Arrays Technology |
| DArTseqMet | Diversity Arrays Technology Methylation Analysis |
| DM | demethylation |
| DNM | de novo methylation |
| GPs | number of green plants regenerated per 100 anthers |
| IUPAC | International Union of Pure and Applied Chemistry |
| metAFLP | methylation sensitive Amplified Fragment Length Polymorphism |
| MSAP | Methylation Sensitive Amplification Polymorphism |
| NGS | Next Generation Sequencing |
| ROS | reactive oxygen species |
| TCIV | tissue culture-induced variation |
References
- Pěnčík, A.; Turečková, V.; Paulišić, S.; Rolčík, J.; Strnad, M.; Mihaljević, S. Ammonium regulates embryogenic potential in Cucurbita pepo through pH-mediated changes in endogenous auxin and abscisic acid. Plant Cell Tissue Organ Cult. (PCTOR) 2015, 122, 89–100. [Google Scholar] [CrossRef]
- Bustillo-Avendaño, E.; Ibáñez, S.; Sanz, O.; Sousa Barros, J.A.; Gude, I.; Perianez-Rodriguez, J.; Micol, J.L.; Del Pozo, J.C.; Moreno-Risueno, M.A.; Pérez-Pérez, J.M. Regulation of Hormonal Control, Cell Reprogramming, and Patterning during De Novo Root Organogenesis. Plant Physiol. 2018, 176, 1709–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyosue, T.; Takano, K.; Kamada, H.; Harada, H. Induction of somatic embryogenesis in carrot by heavy metal ions. Can. J. Bot. 1990, 68, 2301–2303. [Google Scholar] [CrossRef]
- Ikeda-Iwai, M.; Umehara, M.; Satoh, S.; Kamada, H. Stress-induced somatic embryogenesis in vegetative tissues of Arabidopsis thaliana. Plant J. 2003, 34, 107–114. [Google Scholar] [CrossRef]
- Mansilla, N.; Racca, S.; Gras, D.E.; Gonzalez, D.H.; Welchen, E. The Complexity of Mitochondrial Complex IV: An Update of Cytochrome c Oxidase Biogenesis in Plants. Int. J. Mol. Sci. 2018, 19, 662. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.L.C.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14 (Suppl. 1), S131–S151. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Dugardeyn, J.; Zhang, C.; Takenaka, M.; Kühn, K.; Craddock, C.; Smalle, J.; Karampelias, M.; Denecke, J.; Peters, J.; et al. SLO2, a mitochondrial pentatricopeptide repeat protein affecting several RNA editing sites, is required for energy metabolism. Plant J. 2012, 71, 836–849. [Google Scholar] [CrossRef] [Green Version]
- Geisler, D.A.; Päpke, C.; Obata, T.; Nunes-Nesi, A.; Matthes, A.; Schneitz, K.; Maximova, E.; Araújo, W.L.; Fernie, A.R.; Persson, S. Downregulation of the δ-subunit reduces mitochondrial ATP synthase levels, alters respiration, and restricts growth and gametophyte development in Arabidopsis. Plant Cell 2012, 24, 2792–2811. [Google Scholar] [CrossRef] [Green Version]
- Møller, I.M. PLANT MITOCHONDRIA AND OXIDATIVE STRESS: Electron Transport, NADPH Turnover, and Metabolism of Reactive Oxygen Species. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 561–591. [Google Scholar] [CrossRef] [Green Version]
- Aruoma, O.I.; Halliwell, B.; Gajewski, E.; Dizdaroglu, M. Copper-ion-dependent damage to the bases in DNA in the presence of hydrogen peroxide. Biochem. J. 1991, 273 Pt 3, 601–604. [Google Scholar] [CrossRef] [Green Version]
- Drouin, R.; Rodriguez, H.; Gao, S.W.; Gebreyes, Z.; O’Connor, T.R.; Holmquist, G.P.; Akman, S.A. Cupric ion/ascorbate/hydrogen peroxide-induced DNA damage: DNA-bound copper ion primarily induces base modifications. Free Radic. Biol. Med. 1996, 21, 261–273. [Google Scholar] [CrossRef]
- Moreno, L.I.; McCord, B.R. Understanding metal inhibition: The effect of copper (Cu2+) on DNA containing samples. Forensic Chem. 2017, 4, 89–95. [Google Scholar] [CrossRef]
- Sundaravadivel, E.; Reddy, G.R.; Manoj, D.; Rajendran, S.; Kandaswamy, M.; Janakiraman, M. DNA binding and cleavage studies of copper(II) complex containing N2O2 Schiff base ligand. Inorg. Chim. Acta 2018, 482, 170–178. [Google Scholar] [CrossRef]
- Arnesano, F.; Banci, L.; Bertini, I.; Bonvin, A.M.J.J. A Docking Approach to the Study of Copper Trafficking Proteins: Interaction between Metallochaperones and Soluble Domains of Copper ATPases. Structure 2004, 12, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Molphy, Z.; Montagner, D.; Bhat, S.S.; Slator, C.; Long, C.; Erxleben, A.; Kellett, A. A phosphate-targeted dinuclear Cu(II) complex combining major groove binding and oxidative DNA cleavage. Nucleic Acids Res. 2018, 46, 9918–9931. [Google Scholar] [CrossRef]
- Bush, D.S. Calcium Regulation in Plant Cells and its Role in Signaling. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 95–122. [Google Scholar] [CrossRef]
- Kumar, V.; Parvatam, G.; Ravishankar, G.A. AgNO3—A potential regulator of ethylene activity and plant growth modulator. Electron. J. Biotechnol. 2009, 12. [Google Scholar] [CrossRef] [Green Version]
- Testillano, P.S. Microspore embryogenesis: Targeting the determinant factors of stress-induced cell reprogramming for crop improvement. J. Exp. Bot. 2019, 70, 2965–2978. [Google Scholar] [CrossRef]
- El-Tantawy, A.A.; Solís, M.T.; Risueño, M.C.; Testillano, P.S. Changes in DNA Methylation Levels and Nuclear Distribution Patterns after Microspore Reprogramming to Embryogenesis in Barley. Cytogenet. Genome Res. 2014, 143, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Machczyńska, J.; Zimny, J.; Bednarek, P. Tissue culture-induced genetic and epigenetic variation in triticale (× Triticosecale spp. Wittmack ex A. Camus 1927) regenerants. Plant Mol. Biol. 2015, 89, 279–292. [Google Scholar] [CrossRef] [Green Version]
- Baranek, M.; Cechova, J.; Kovacs, T.; Eichmeier, A.; Wang, S.; Raddova, J.; Necas, T.; Ye, X. Use of Combined MSAP and NGS Techniques to Identify Differentially Methylated Regions in Somaclones: A Case Study of Two Stable Somatic Wheat Mutants. PLoS ONE 2016, 11, e0165749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbroucke, H.; Mournet, P.; Vignes, H.; Chaïr, H.; Malapa, R.; Duval, M.F.; Lebot, V. Somaclonal variants of taro (Colocasia esculenta Schott) and yam (Dioscorea alata L.) are incorporated into farmers’ varietal portfolios in Vanuatu. Genet. Resour. Crop. Evol. 2016, 63, 495–511. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Lee, T.v.d.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bednarek, P.T.; Orłowska, R.; Koebner, R.M.D.; Zimny, J. Quantification of the tissue-culture induced variation in barley (Hordeum vulgare L.). BMC Plant Biol. 2007, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machczyńska, J.; Orłowska, R.; Zimny, J.; Bednarek, P.T. Extended metAFLP approach in studies of the tissue culture induced variation (TCIV) in case of triticale. Mol. Breed. 2014, 34, 845–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity arrays: A solid state technology for sequence information independent genotyping. Nucleic Acids Res. 2001, 29, E25. [Google Scholar] [CrossRef] [Green Version]
- Sansaloni, C.; Petroli, C.; Jaccoud, D.; Carling, J.; Detering, F.; Grattapaglia, D.; Kilian, A. Diversity Arrays Technology (DArT) and next-generation sequencing combined: Genome-wide, high throughput, highly informative genotyping for molecular breeding of Eucalyptus. BMC Proc. 2011, 5, P54. [Google Scholar] [CrossRef] [Green Version]
- Bednarek, P.T.; Orłowska, R.; Niedziela, A. A relative quantitative Methylation-Sensitive Amplified Polymorphism (MSAP) method for the analysis of abiotic stress. BMC Plant Biol. 2017, 17, 79. [Google Scholar] [CrossRef] [Green Version]
- Hayes, A.F. Introduction to Mediation, Moderation, and Conditional Process Analysis. A Regression Bases Approach; A Division of Guilford Publications, Inc.: New York, NY, USA, 2018. [Google Scholar]
- MacKinnon, D.P.; Luecken, L.J. How and for whom? Mediation and moderation in health psychology. Health Psychol. 2008, 27, S99–S100. [Google Scholar] [CrossRef] [Green Version]
- Holroyd, K.A.; Labus, J.S.; Carlson, B. Moderation and mediation in the psychological and drug treatment of chronic tension-type headache: The role of disorder severity and psychiatric comorbidity. Pain 2009, 143, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Namazi, M.; Namazi, N.-R. Conceptual Analysis of Moderator and Mediator Variables in Business Research. Procedia Econ. Financ. 2016, 36, 540–554. [Google Scholar] [CrossRef] [Green Version]
- Bednarek, P.T.; Orłowska, R. CG Demethylation Leads to Sequence Mutations in an Anther Culture of Barley Due to the Presence of Cu, Ag Ions in the Medium and Culture Time. Int. J. Mol. Sci. 2020, 21, 4401. [Google Scholar] [CrossRef] [PubMed]
- Orłowska, R.; Pachota, K.A.; Machczyńska, J.; Niedziela, A.; Makowska, K.; Zimny, J.; Bednarek, P.T. Improvement of anther cultures conditions using the Taguchi method in three cereal crops. Electron. J. Biotechnol. 2020, 43, 8–15. [Google Scholar] [CrossRef]
- Bednarek, P.T.; Orłowska, R. Plant tissue culture environment as a switch-key of (epi)genetic changes. Plant Celltissue Organ Cult. (PCTOR) 2020, 140, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez López, C.M.; Wetten, A.C.; Wilkinson, M.J. Progressive erosion of genetic and epigenetic variation in callus-derived cocoa (Theobroma cacao) plants. New Phytol. 2010, 186, 856–868. [Google Scholar] [CrossRef] [PubMed]
- Rival, A.; Ilbert, P.; Labeyrie, A.; Torres, E.; Doulbeau, S.; Personne, A.; Dussert, S.; Beule’, T.; Durand-Gasselin, T.; Tregear, J.W.; et al. Variations in genomic DNA methylation during the long-term in vitro proliferation of oil palm embryogenic suspension cultures. Plant Cell Rep. 2013, 32, 359–368. [Google Scholar] [CrossRef]
- Bordallo, P.N.; Silva, D.H.; Maria, J.; Cruz, C.D.; Fontes, E.P. Somaclonal variation on in vitro callus culture potato cultivars. Hortic. Bras. 2004, 22, 300–304. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.Z.; Xu, C.G.; Saghai Maroof, M.A.; Zhang, Q. Patterns of cytosine methylation in an elite rice hybrid and its parental lines, detected by a methylation-sensitive amplifcation polymorphism technique. Mol Gen. Genet. 1999, 261, 439–446. [Google Scholar] [CrossRef]
- Tenhola-Roininen, T.; Immonen, S.; Tanhuanpää, P. Rye doubled haploids as a research and breeding tool—A practical point of view. Plant Breed. 2006, 125, 584–590. [Google Scholar] [CrossRef]
- Ślusarkiewicz-Jarzina, A.; Pudelska, H.; Wozna, J.; Pniewski, T. Improved production of doubled haploids of winter and spring triticale hybrids via combination of colchicine treatments on anthers and regenerated plants. J. Appl. Genet. 2017, 58, 287–295. [Google Scholar] [CrossRef] [Green Version]
- El-Hennawy, M.A.; Abdalla, A.F.; Shafey, S.A.; Al-Ashkar, I.M. Production of doubled haploid wheat lines (Triticum aestivum L.) using anther culture technique. Ann. Agric. Sci. 2011, 56, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Ohnoutkova, L.; Vlcko, T.; Ayalew, M. Barley anther culture. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2019; Volume 1900, pp. 37–52. [Google Scholar]
- Machczyńska, J.; Orłowska, R.; Mańkowski, D.R.; Zimny, J.; Bednarek, P.T. DNA methylation changes in triticale due to in vitro culture plant regeneration and consecutive reproduction. Plant Cell Tissue Organ Cult. 2014, 119, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Orłowska, R.; Machczyńska, J.; Oleszczuk, S.; Zimny, J.; Bednarek, P.T. DNA methylation changes and TE activity induced in tissue cultures of barley (Hordeum vulgare L.). J. Biol. Res. (Thessalon. Greece) 2016, 23, 19. [Google Scholar] [CrossRef] [Green Version]
- Fiuk, A.; Bednarek, P.T.; Rybczyński, J.J. Flow Cytometry, HPLC-RP, and metAFLP Analyses to Assess Genetic Variability in Somatic Embryo-Derived Plantlets of Gentiana pannonica Scop. Plant Mol. Biol. Rep. 2010, 28, 413–420. [Google Scholar] [CrossRef]
- Coronel, C.J.; González, A.I.; Ruiz, M.L.; Polanco, C. Analysis of somaclonal variation in transgenic and regenerated plants of Arabidopsis thaliana using methylation related metAFLP and TMD markers. Plant Cell Rep. 2018, 37, 137–152. [Google Scholar] [CrossRef]
- Han, Z.; Crisp, P.A.; Stelpflug, S.; Kaeppler, S.M.; Li, Q.; Springer, N.M. Heritable Epigenomic Changes to the Maize Methylome Resulting from Tissue Culture. Genetics 2018, 209, 983–995. [Google Scholar] [CrossRef]
- Orłowska, R.; Bednarek, P.T. Precise evaluation of tissue culture-induced variation during optimisation of in vitro regeneration regime in barley. Plant Mol. Biol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Festa, R.A.; Thiele, D.J. Copper: An Essential Metal in Biology. Curr. Biol. 2011, 8, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Klíma, P.; Laňková, M.; Vandenbussche, F.; Van Der Straeten, D.; Petrášek, J. Silver ions increase plasma membrane permeability through modulation of intracellular calcium levels in tobacco BY-2 cells. Plant Cell Rep. 2018, 37, 809–818. [Google Scholar] [CrossRef]
- Makowska, K.; Oleszczuk, S. Albinism in barley androgenesis. Plant Cell Rep. 2014, 33, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Jacquard, C.; Nolin, F.; Hecart, C.; Grauda, D.; Rashal, I.; Dhondt-Cordelier, S.; Sangwan, R.S.; Devaux, P.; Mazeyrat-Gourbeyre, F.; Clement, C. Microspore embryogenesis and programmed cell death in barley: Effects of copper on albinism in recalcitrant cultivars. Plant Cell Rep. 2009, 28, 1329–1339. [Google Scholar] [CrossRef]
- Fei, S.; Read, P.E.; Riordan, T.P. Improvement of embryogenic callus induction and shoot regeneration of buffalograss by silver nitrate. Plant Cell Tissue Organ Cult. 2000, 60, 197–203. [Google Scholar] [CrossRef]
- Nuutila, A.M.; Hämäläinen, J.; Mannonen, L. Optimization of media nitrogen and copper concentrations for regeneration of green plants from polyembryogenic cultures of barley (Hordeum ulgare L.). Plant Sci. 2000, 151, 85–92. [Google Scholar] [CrossRef]
- Haque, M.; Siddique, A.B.; Shahinul Islam, S.M. Effect of silver nitrate and amino acids on high frequency plants regeneration in barley (Hordeum vulgare L.). Plant Tissue Cult. Biotechnol. 2015, 25, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009, 36, 409–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, R.A.; Fincher, G.B. (1,3;1,4)-β-D-Glucans in Cell Walls of the Poaceae, Lower Plants, and Fungi: A Tale of Two Linkages. Mol. Plant 2009, 2, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Burton, R.A.; Collins, H.M.; Kibble, N.A.J.; Smith, J.A.; Shirley, N.J.; Jobling, S.A.; Henderson, M.; Singh, R.R.; Pettolino, F.; Wilson, S.M.; et al. Over-expression of specific HvCslF cellulose synthase-like genes in transgenic barley increases the levels of cell wall (1,3;1,4)-β-d-glucans and alters their fine structure. Plant Biotechnol. J. 2011, 9, 117–135. [Google Scholar] [CrossRef]
- Oleszczuk, S.; Tyrka, M.; Zimny, J. The origin of clones among androgenic regenerants of hexaploid triticale. Euphytica 2014, 198, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Beyer, E.M. Silver ion: A potent anti-ethylene agent in cucumber and tomato. HortScience 1976, 11, 175–196. [Google Scholar]
- Costa, C.S.; Ronconi, J.V.V.; Daufenbach, J.F.; Gonçalves, C.L.; Rezin, G.T.; Streck, E.L.; da Silva Paula, M.M. In vitro effects of silver nanoparticles on the mitochondrial respiratory chain. Mol. Cell. Biochem. 2010, 342, 51–56. [Google Scholar] [CrossRef]
- Romani, R.J. DNA methylation levels and ethylene production in senescent, suspension-cultured pear fruit cells: Implications for epigenetic control? Physiol. Plant. 1998, 103, 534–540. [Google Scholar] [CrossRef]
- Shrout, P.E.; Bolger, N. Mediation in experimental and nonexperimental studies: New procedures and recommendations. Psychol. Methods 2002, 7, 422–445. [Google Scholar] [CrossRef] [PubMed]
- Preacher, K.J.; Hayes, A.F. SPSS and SAS procedures for estimating indirect effects in simple mediation models. Behav. Res. Methods Instrum. Comput. 2004, 36, 717–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, B.M.; Rodriguez, F.I.; Bleecker, A.B.; Patterson, S.E. The effects of Group 11 transition metals, including gold, on ethylene binding to the ETR1 receptor and growth of Arabidopsis thaliana. FEBS Lett. 2007, 581, 5105–5109. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Regenerant | Trial | DNM (%) | DM (%) | Delta (%) | GPs |
|---|---|---|---|---|---|
| 1 | M1 | 28.153 | 32.179 | 4.026 | 0.64 |
| 2 | M1 | 28.188 | 32.289 | 4.101 | 0.64 |
| 3 | M1 | 28.155 | 32.247 | 4.092 | 0.64 |
| 4 | M1 | 28.135 | 32.172 | 4.037 | 0.64 |
| 5 | M1 | 28.102 | 32.209 | 4.107 | 0.64 |
| 6 | M2 | 31.094 | 29.429 | −1.665 | 0.67 |
| 7 | M2 | 31.095 | 29.583 | −1.512 | 0.67 |
| 8 | M2 | 31.005 | 29.457 | −1.548 | 0.67 |
| 9 | M3 | 30.915 | 29.288 | −1.627 | 1.09 |
| 10 | M3 | 30.855 | 29.301 | −1.554 | 1.09 |
| 11 | M3 | 30.876 | 29.332 | −1.544 | 1.09 |
| 12 | M3 | 30.915 | 29.276 | −1.639 | 1.09 |
| 13 | M3 | 30.920 | 29.309 | −1.611 | 1.09 |
| 14 | M4 | 30.421 | 30.532 | 0.111 | 0.45 |
| 15 | M4 | 30.457 | 30.591 | 0.134 | 0.45 |
| 16 | M5 | 6.897 | 7.834 | 0.937 | 0.1 |
| 17 | M5 | 10.804 | 11.344 | 0.540 | 0.1 |
| 18 | M5 | 6.825 | 7.609 | 0.784 | 0.1 |
| 19 | M5 | 11.043 | 11.170 | 0.127 | 0.1 |
| 20 | M6 | 31.524 | 28.674 | −2.850 | 2.12 |
| 21 | M6 | 31.295 | 28.650 | −2.645 | 2.12 |
| 22 | M6 | 31.505 | 28.567 | −2.938 | 2.12 |
| 23 | M6 | 31.517 | 28.674 | −2.843 | 2.12 |
| 24 | M6 | 31.355 | 28.815 | −2.540 | 2.12 |
| 25 | M7 | 29.909 | 29.972 | 0.063 | 2.91 |
| 26 | M7 | 29.999 | 30.106 | 0.107 | 2.91 |
| 27 | M7 | 29.979 | 29.991 | 0.012 | 2.91 |
| 28 | M8 | 31.260 | 29.659 | −1.601 | 1.77 |
| 29 | M8 | 31.275 | 29.784 | −1.491 | 1.77 |
| 30 | M8 | 31.249 | 29.786 | −1.463 | 1.77 |
| 31 | M9 | 32.197 | 28.127 | −4.07 | 0.54 |
| 32 | M9 | 32.265 | 28.277 | −3.988 | 0.54 |
| 33 | M9 | 32.234 | 28.225 | −4.009 | 0.54 |
| 34 | M9 | 32.195 | 28.308 | −3.887 | 0.54 |
| 35 | M9 | 32.192 | 28.286 | −3.906 | 0.54 |
| M | 28.196 | 27.402 | 0.794 | 1.123 | |
| SD | 7.17 | 6.67 | 2.47 | 0.85 | |
| Predictors | Statistics | |||
|---|---|---|---|---|
| β | SE | 95% LLCI | 95% ULCI | |
| Delta = DM-DNM | 3.06 ***(0.079 ***) | 0.12(0.19) | 2.812(0.323) | 3.310(1.096) |
| Cu2+ (Ag+) | −0.93 ***(−0.87 ***) | 0.05(0.06) | −1.031(−0.987) | −0.837(−0.753) |
| Delta × Cu2+ (Ag+) | −0.86 ***(−0.606 ***) | 0.04(0.04) | −0.950(−0.689) | −0.771(−0523) |
| Time | −0.21 ***(0.017) | 0.01(0.01) | −0.229(−0.007) | −0.192(0.040) |
| Delta × Time | −0.15 ***(−0.028 ***) | 0.01(0.00) | −0.157(−0.044) | −0.136(−0.12) |
| Cu2+ (Ag+) × Time | 0.04 ***(0.031 ***) | 0.01(0.00) | 0.038(0.027) | 0.044(0.035) |
| Delta × Cu2+ (Ag+)+ × Time | 0.03 ***(0.021 ***) | 0.00(0.00) | 0.032(0.018) | 0.038(0.024) |
| Model Summary for Cu2+ (Ag+) | R2 = 0.985, F (7,27) = 249.55, MSE = 0.014, p < 0.001 (R2 = 0.924, F (7,27) = 46.99, MSE = 0.014, p < 0.001) | |||
| Test of highest order unconditional interaction (X × W × Z: Delta × Cu2+ (Ag+) × Time) | F(1,27) = 527.15, R2chng = 0.297, p < 0.001 (F(1,27) = 219.76, R2chng = 0.617, p < 0.0001) *** p < 0.001 | |||
| Days | DM 1 | DNM 1 |
|---|---|---|
| 21 | 30.29 a | 30.13 a |
| 28 | 29.08 ab | 31.52 a |
| 35 | 22.88 b | 23.33 b |
| Regenerant | Trial | Cu2+ (µM) | Ag+ (µM) | Time (Days) |
|---|---|---|---|---|
| 1–5 | M1 | 0.1 | 0 | 21 |
| 6–8 | M2 | 0.1 | 10 | 28 |
| 9–13 | M3 | 0.1 | 60 | 35 |
| 14–15 | M4 | 5 | 60 | 28 |
| 16–19 | M5 | 5 | 0 | 35 |
| 20–24 | M6 | 5 | 10 | 21 |
| 25–27 | M7 | 10 | 10 | 35 |
| 28–30 | M8 | 10 | 60 | 21 |
| 31–35 | M9 | 10 | 0 | 28 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bednarek, P.T.; Orłowska, R. Time of In Vitro Anther Culture May Moderate Action of Copper and Silver Ions that Affect the Relationship between DNA Methylation Change and the Yield of Barley Green Regenerants. Plants 2020, 9, 1064. https://doi.org/10.3390/plants9091064
Bednarek PT, Orłowska R. Time of In Vitro Anther Culture May Moderate Action of Copper and Silver Ions that Affect the Relationship between DNA Methylation Change and the Yield of Barley Green Regenerants. Plants. 2020; 9(9):1064. https://doi.org/10.3390/plants9091064
Chicago/Turabian StyleBednarek, Piotr T., and Renata Orłowska. 2020. "Time of In Vitro Anther Culture May Moderate Action of Copper and Silver Ions that Affect the Relationship between DNA Methylation Change and the Yield of Barley Green Regenerants" Plants 9, no. 9: 1064. https://doi.org/10.3390/plants9091064
APA StyleBednarek, P. T., & Orłowska, R. (2020). Time of In Vitro Anther Culture May Moderate Action of Copper and Silver Ions that Affect the Relationship between DNA Methylation Change and the Yield of Barley Green Regenerants. Plants, 9(9), 1064. https://doi.org/10.3390/plants9091064