Chlorophyll a Fluorescence Transient and 2-Dimensional Electrophoresis Analyses Reveal Response Characteristics of Photosynthesis to Heat Stress in Malus. ‘Prairifire’


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Treatments
2.3. Measurements of Photosynthetic Gas Exchange Parameters
2.4. Extraction and Quantification of Proteins in Leaves of M. ‘Prairifire’
2.5. 2-DE Analysis
2.6. Protein Identification and Database Search
2.7. Protein Annotation and Interaction
2.8. Statistical Analysis
3. Results
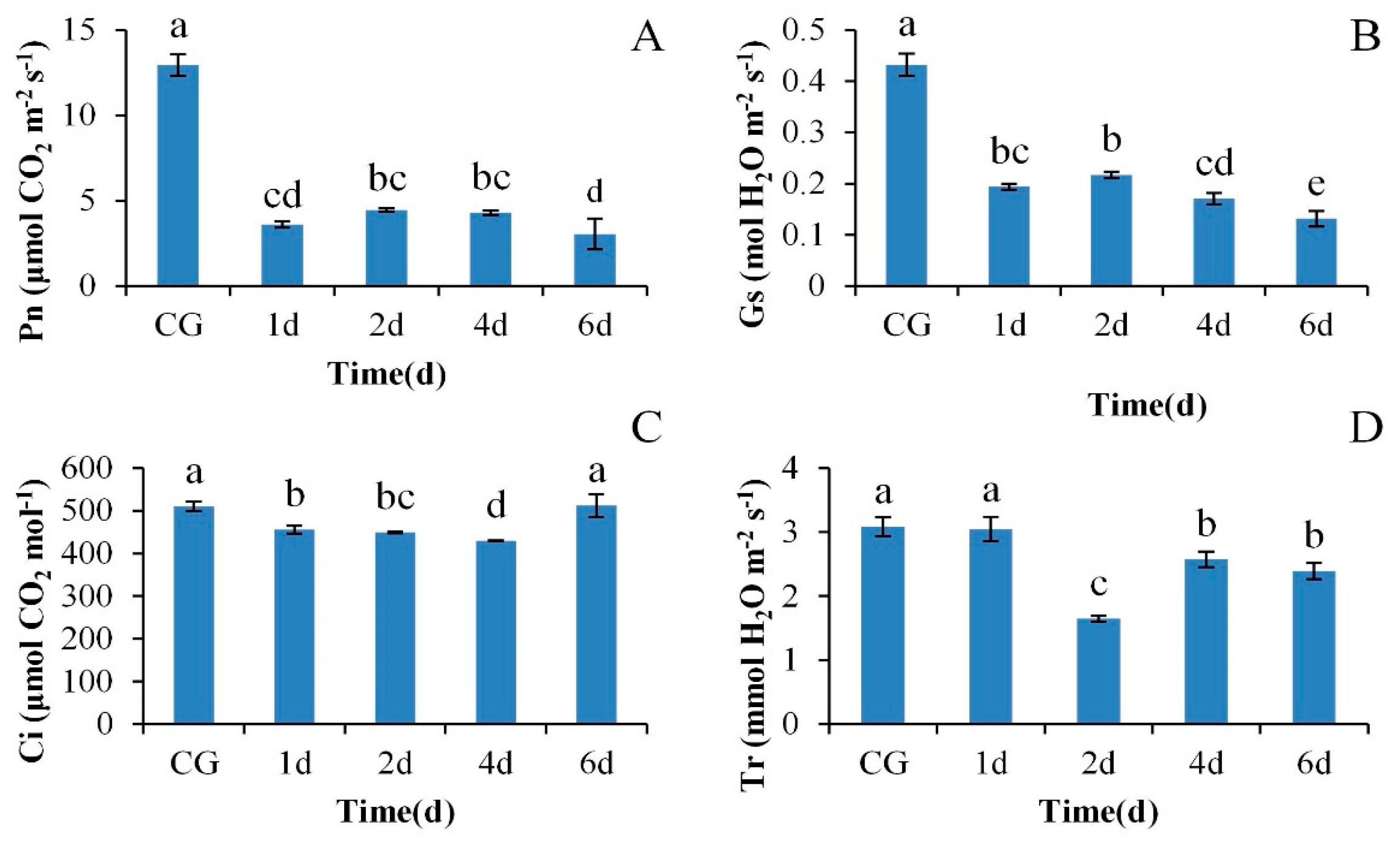
3.1. The Response of Photosynthetic Gas Exchange Parameters of M. ‘Prairifire’ Leaves Exposed to High Temperature Stress
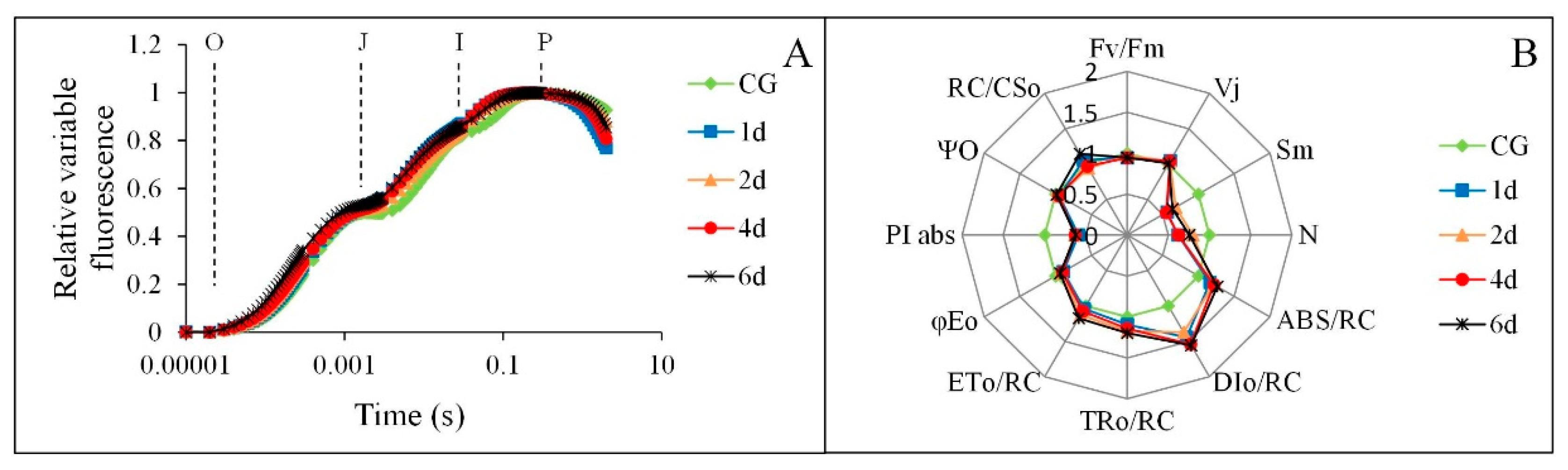
3.2. The Response of OJIP Curves of M. ‘Prairifire’ to Heat Stress
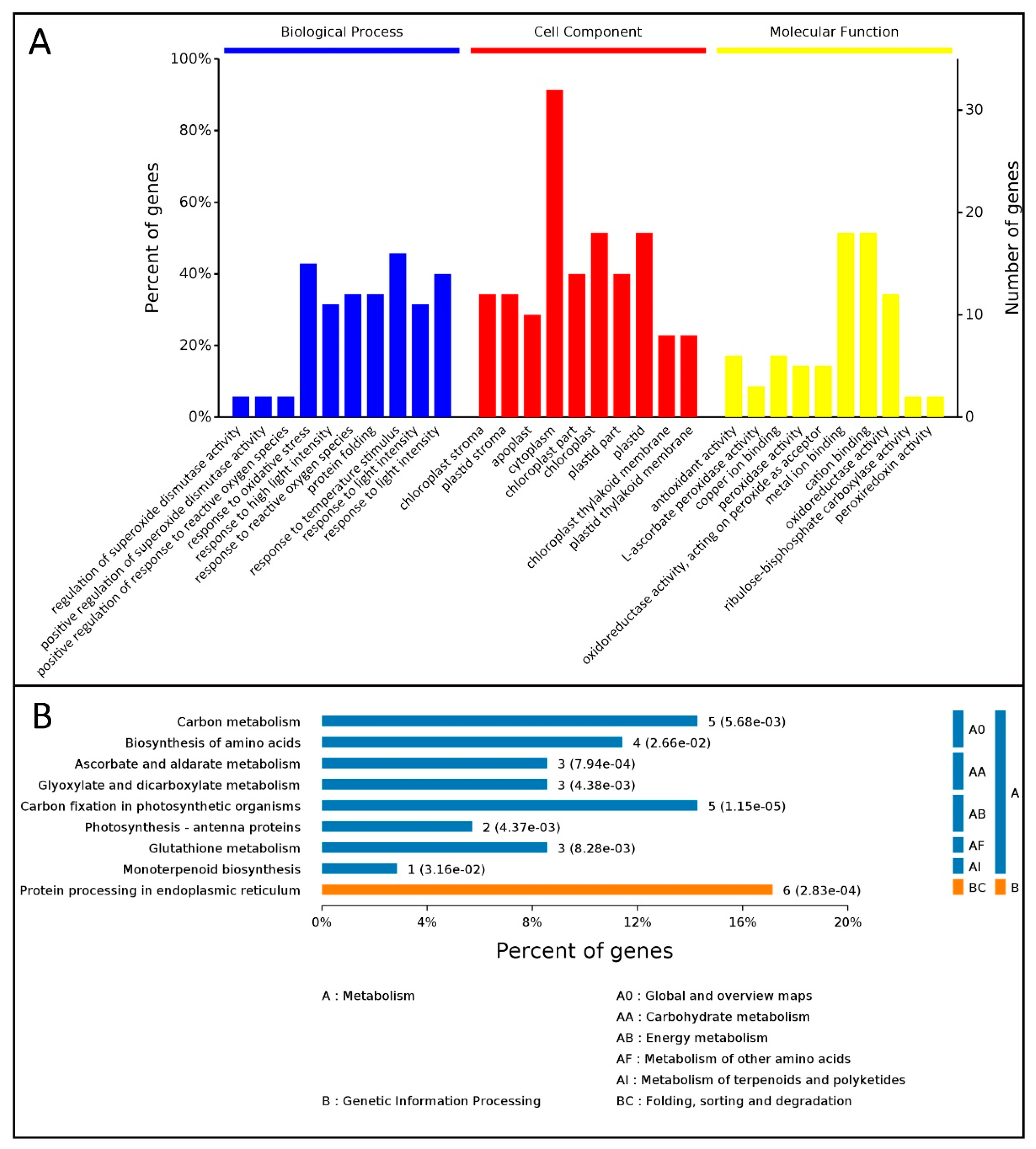
3.3. Identification of DEPs in Leaves of M. ‘Prairifire’ between CG and Shock Treatment for 48 h
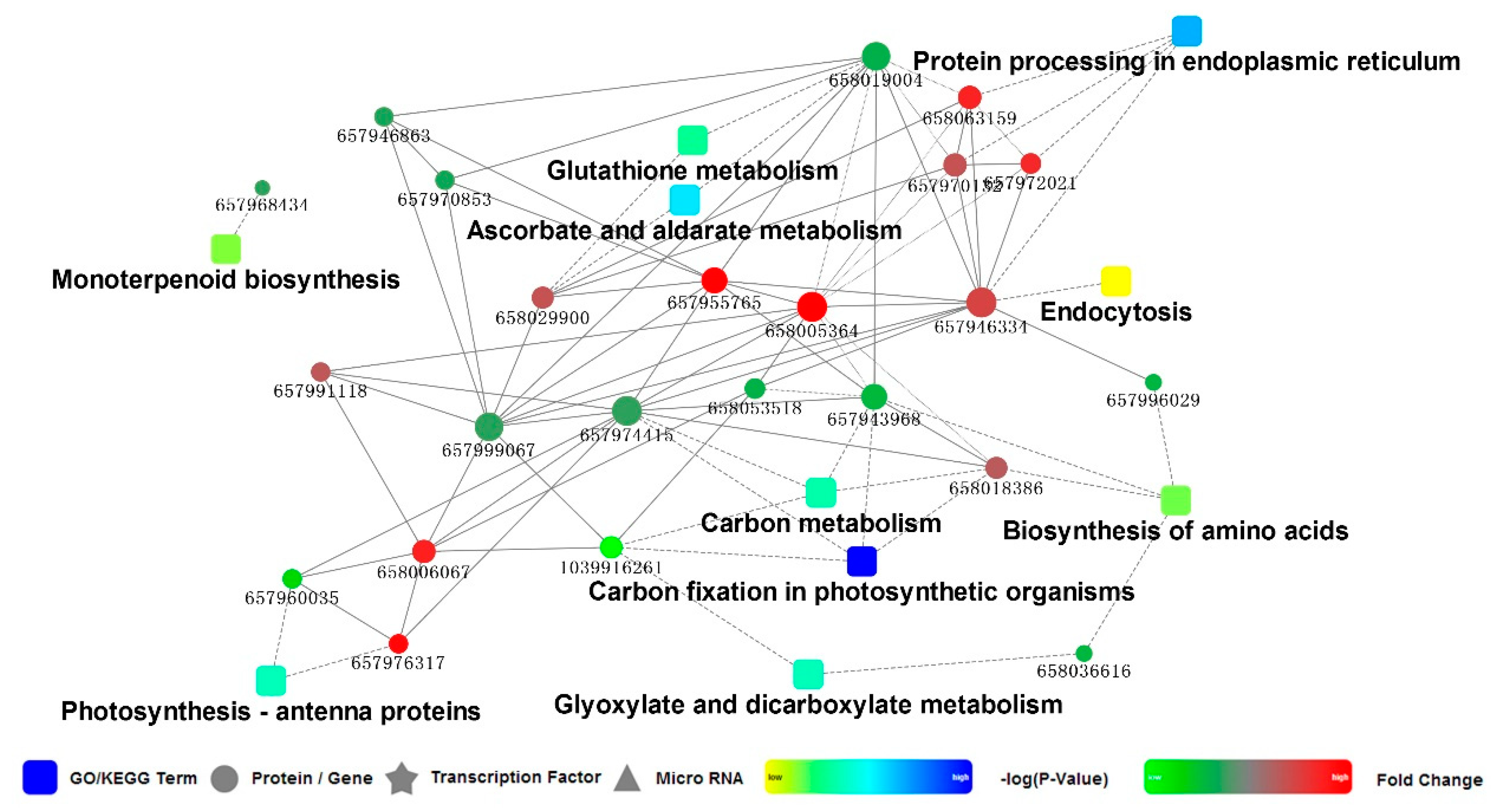
3.4. PPI Network for the DEPs in Leaves of M. ‘Prairifire’ between CG and Heat Shock Treatment for 48 h
3.5. Differential Expression of Photosynthesis-Related Proteins in Leaves of M. ‘Prairifire’ between CG and Heat Shock Treatment for 48 h
4. Discussion
4.1. Electron Transport Chain and Related Proteins Involved in the Light Reaction
4.2. Proteins Involved in the Dark Reaction and Regulatory Mechanisms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, S.; Zhu, S.; Chen, L.; Liu, F.; Zou, M.; Wang, C. Comparative response of two wucai (Brassica campestris L.) genotypes to heat stress on antioxidative system and cell ultrastructure in root. Acta Physiol. Plant 2016, 38, 223. [Google Scholar] [CrossRef]
- Zhang, M.; Shan, Y.J.; Kochian, L.; Strasser, R.J.; Chen, G. Photochemical properties in flag leaves of a super-high-yielding hybrid rice and a traditional hybrid rice (Oryza sativa L.) probed by chlorophyll a fluorescence transient. Photosynth. Res. 2015, 126, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Bechtold, U.; Field, B. Molecular mechanisms controlling plant growth during abiotic stress. J. Exp. Bot. 2018, 69, 2753–2758. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Sun, X.; Amombo, E.; Zhu, Q.; Zhao, Z.; Chen, L.; Xu, Q.; Fu, J. High correlation between thermotolerance and photosystem II activity in tall fescue. Photosynth. Res. 2014, 122, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.S.; Semedo, J.; Ramalho, J.C.; Lidon, F.C. Bread and durum wheat under heat stress: A comparative study on the photosynthetic performance. J. Agron. Crop Sci. 2011, 197, 50–56. [Google Scholar] [CrossRef]
- Yan, K.; Chen, P.; Shao, H.; Zhang, L.; Xu, G. Effects of short-term high temperature on photosynthesis and photosystem II performance in sorghum. J. Agron. Crop Sci. 2011, 197, 400–408. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence a Signature of Photosynthesis; Papageorgiou, G., Govindjee, C., Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 321–362. [Google Scholar]
- Chen, S.; Yang, J.; Zhang, M.; Strasser, R.J.; Qiang, S. Classification and characteristics of heat tolerance in Ageratina adenophora populations using fast chlorophyll a fluorescence rise OJIP. Environ. Exp. Bot. 2016, 122, 126–140. [Google Scholar] [CrossRef]
- Chen, S.; Xu, X.; Dai, X.; Yang, C.; Qiang, S. Identification of tenuazonic acid as a novel type of natural photosystem II inhibitor binding in QB-site of Chlamydomonas reinhardtii. BBA Bioenerg. 2007, 1767, 306–318. [Google Scholar] [CrossRef]
- Zhang, M.; Li, G.; Huang, W.; Bi, T.; Chen, G.; Tang, Z.; Su, W.; Sun, W. Proteomic study of Carissa spinarum in response to combined heat and drought stress. Proteomics 2010, 10, 3117–3129. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, J.; Liu, Q.; Ben, C.; Todd, C.D.; Shi, J.; Yang, Y.; Hu, X. Comparative proteomic analysis of the thermotolerant plant Portulaca oleracea acclimation to combined high temperature and humidity stress. J. Proteome Res. 2012, 11, 3605. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, N.; Donnart, T.; Nouri, M.Z.; Komatsu, S. Tissue-specific defense and thermo-adaptive mechanisms of soybean seedlings under heat stress revealed by proteomic approach. J. Proteome Res. 2010, 9, 4189–4204. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.; Hjernø, K.; Larsen, M.; Wingsle, G.; Larsen, P.; Fey, S.; Roepstorff, P.; Salomé Pais, M. Proteome profiling of Populus euphratica Oliv. upon heat stress. Ann. Bot. Lond. 2006, 98, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Rocco, M.; Arena, S.; Renzone, G.; Scippa, G.S.; Lomaglio, T.; Verrillo, F.; Scaloni, A.; Marra, M. Proteomic analysis of temperature stress-responsive proteins in Arabidopsis thaliana rosette leaves. Mol. Biosyst. 2013, 9, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.X.; Zhao, M.M.; Fan, J.J.; Zhou, T.; Chen, Y.X.; Cao, F.L. Study on relationship between pollen exine ornamentation pattern and germplasm evolution in flowering crabapple. Sci. Rep. UK 2017, 7, 39759. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, M.; Wang, M. Phenolic compounds and antioxidant activities of flowers, leaves and fruits of five crabapple cultivars (Malus Mill. species). Sci. Hortic Amst. 2018, 235, 460–467. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Bu, Y.F.; Zhang, X.; Yao, Y. Factor analysis of MYB gene expression and flavonoid affecting petal color in three crabapple cultivars. Front. Plant Sci. 2017, 8, 137. [Google Scholar] [CrossRef]
- Liu, C.F.; Zhang, W.X.; Sun, L.; Ruan, J.Y.; Wang, T.; Cao, F.L. Effects of high temperature on photosynthesis and growth of crabapple seedling. J. Nanjing Forest. Univ. (Nat. Sci. Ed.) 2013, 4, 17–22. [Google Scholar]
- Wan, H.; Zhang, J.; Song, T.; Tian, J.; Yao, Y. Promotion of flavonoid biosynthesis in leaves and calli of ornamental crabapple (Malus sp.) by high carbon to nitrogen ratios. Front. Plant Sci. 2015, 6, 673. [Google Scholar] [CrossRef]
- Zou, M.; Yuan, L.; Zhu, S.; Liu, S.; Ge, J.; Wang, C. Effects of heat stress on photosynthetic characteristics and chloroplast ultrastructure of a heat-sensitive and heat-tolerant cultivar of wucai (Brassica campestris L.). Acta Physiol. Plant. 2017, 39, 30. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Sade, D.; Sade, N.; Shriki, O.; Lerner, S.; Gebremedhin, A.; Karavani, A. Water balance, hormone homeostasis, and sugar signaling are all involved in tomato resistance to tomato yellow leaf curl virus. Plant Physiol. 2014, 4, 1684–1697. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.T.; Ma, L.; Duan, W.; Wang, B.C.; Li, J.H.; Xu, H.G.; Yan, X.Q.; Yan, B.F.; Li, S.H.; Wang, L.J. Differential proteomic analysis of grapevine leaves by iTRAQ reveals responses to heat stress and subsequent recovery. BMC Plant Biol. 2014, 14, 110. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; Chen, P.; Shao, H.; Shao, C.; Zhao, S.; Brestic, M. Dissection of photosynthetic electron transport process in sweet sorghum under heat stress. PLoS ONE 2013, 8, e62100. [Google Scholar] [CrossRef] [PubMed]
- Roose, J.L.; Wegener, K.M.; Pakrasi, H.B. The extrinsic proteins of photosystem II. Photosynth. Res. 2007, 92, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Markou, G.; Depraetere, O.; Muylaert, K. Effect of ammonia on the photosynthetic activity of Arthrospira and Chlorella: A study on chlorophyll fluorescence and electron transport. Algal Res. 2016, 16, 449–457. [Google Scholar] [CrossRef]
- Stirbet, A. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Jedmowski, C.; Brüggemann, W. Imaging of fast chlorophyll fluorescence induction curve (OJIP) parameters, applied in a screening study with wild barley (Hordeum spontaneum) genotypes under heat stress. J. Photochem. Photobiol. B 2015, 151, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Chen, H.; Li, X.J.; Yang, M.F.; Liu, G.S.; Shen, S.H. A comparative proteomic analysis of rice seedlings under various high-temperature stresses. BBA Proteins Proteom. 2009, 1794, 1625–1634. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Kocsy, G.; Kobrehel, K.; Szalai, G.; Duviau, M.; Buzás, Z.; Galiba, G. Abiotic stress-induced changes in glutathione and thioredoxin h, levels in maize. Environ. Exp. Bot. 2004, 52, 101–112. [Google Scholar] [CrossRef]
- Salekdeh, G.H.; Siopongco, J.; Wade, L.J.; Ghareyazie, B.; Bennett, J. Proteomic analysis of rice leaves during drought stress and recovery. Proteomics 2002, 2, 1131–1145. [Google Scholar] [CrossRef]
- Yoshioka, M.; Yamamoto, Y. Quality control of Photosystem II: Where and how does the degradation of the D1 protein by FtsH proteases start under light stress? -Facts and hypotheses. J. Photochem. Photobiol. B 2011, 104, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Ahsan, N.; Lee, S.H.; Kang, K.Y.; Bahk, J.D.; Lee, I.J.; Lee, B.H. A proteomic approach in analyzing heat-responsive proteins in rice leaves. Proteomics 2007, 7, 3369–3383. [Google Scholar] [CrossRef] [PubMed]
- Bernacchia, G.; Schwall, G.; Lottspeich, F.; Salamini, F.; Bartels, D. The transketolase gene family of the resurrection plant Craterostigma plantagineum: Differential expression during the rehydration phase. EMBO J. 1995, 14, 610. [Google Scholar] [CrossRef]
- Bi, H.; Dong, X.; Wu, G.; Wang, M.; Ai, X. Decreased TK activity alters growth, yield and tolerance to low temperature and low light intensity in transgenic cucumber plants. Plant Cell Rep. 2015, 34, 345–354. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, H. Differential accumulation of proteins in leaves and roots associated with heat tolerance in two Kentucky bluegrass genotypes differing in heat tolerance. Acta Physiol. Plant. 2016, 38, 213. [Google Scholar] [CrossRef]
- Weber, A.P.M. Synthesis, export and partitioning of the end products of photosynthesis. In The Structure and Function of Plastids; Wise, R.R., Hoober, J.K., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 23, pp. 273–292. [Google Scholar]
- Kim, K.H.; Alam, I.; Kim, Y.G.; Sharmin, S.A.; Lee, K.W.; Lee, S.H.; Lee, B.H. Overexpression of a chloroplast-localized small heat shock protein OsHSP26 confers enhanced tolerance against oxidative and heat stresses in tall fescue. Biotechnol. Lett. 2012, 34, 371–377. [Google Scholar] [CrossRef]
- Tsai, Y.C.C.; Mueller-Cajar, O.; Saschenbrecker, S.; Hartl, F.U.; Hayer-Hartl, M. Chaperonin cofactors, Cpn10 and Cpn20, of green algae and plants function as hetero-oligomeric ring complexes. J. Biol. Chem. 2012, 287, 20471–20481. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Protein Name | gi Number | UniProt ID | Fold Change |
|---|---|---|---|---|
| 1 | PREDICTED: thioredoxin H-type | gi|657970853 | P29448 | 0.445 |
| 2 | PREDICTED: triosephosphate isomerase, cytosolic-like | gi|657943968 | P48491 | 0.292 |
| 3 | PREDICTED: 2-Cys peroxiredoxin BAS1, chloroplastic-like | gi|657999067 | Q9C5R8 | 0.486 |
| 4 | PREDICTED: ATP-dependent zinc metalloprotease FTSH 2, chloroplastic | gi|658053518 | O80860 | 0.344 |
| 5 | PREDICTED: chlorophyll a-b binding protein of LHCII type 1 | gi|657960035 | Q39142 | 0.143 |
| 6 | PREDICTED: ribulose bisphosphate carboxylase large chain | gi|1039916261 | O03042 | 0.059 |
| 7 | ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit, partial (plastid) | gi|817992125 | O03042 | 0.243 |
| 8 | PREDICTED: glyceraldehyde-3-phosphate dehydrogenase A, chloroplastic | gi|657974415 | Q9LPW0 | 0.498 |
| 9 | PREDICTED: photosystem I chlorophyll a/b-binding protein 3-1, chloroplastic-like | gi|657976317 | Q9SY97 | 8.559 |
| 10 | PREDICTED: small heat shock protein, chloroplastic | gi|657970132 | P31170 | 2.356 |
| 11 | PREDICTED: 20 kDa chaperonin, chloroplastic-like | gi|658005364 | O65282 | 11.158 |
| 12 | PREDICTED: 20 kDa chaperonin, chloroplastic-like | gi|657947332 | O65282 | 2.415 |
| 13 | PREDICTED: transketolase, chloroplastic, partial | gi|658018386 | F4IW47 | 2 |
| 14 | PREDICTED: oxygen-evolving enhancer protein 1, chloroplastic | gi|658006067 | Q9S841 | 5.688 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Luo, S.; Ma, Y.; Li, L.; Xie, Y.; Zhang, W. Chlorophyll a Fluorescence Transient and 2-Dimensional Electrophoresis Analyses Reveal Response Characteristics of Photosynthesis to Heat Stress in Malus. ‘Prairifire’. Plants 2020, 9, 1040. https://doi.org/10.3390/plants9081040
Wang T, Luo S, Ma Y, Li L, Xie Y, Zhang W. Chlorophyll a Fluorescence Transient and 2-Dimensional Electrophoresis Analyses Reveal Response Characteristics of Photosynthesis to Heat Stress in Malus. ‘Prairifire’. Plants. 2020; 9(8):1040. https://doi.org/10.3390/plants9081040
Chicago/Turabian StyleWang, Tao, Siqian Luo, Yingli Ma, Lingyu Li, Yinfeng Xie, and Wangxiang Zhang. 2020. "Chlorophyll a Fluorescence Transient and 2-Dimensional Electrophoresis Analyses Reveal Response Characteristics of Photosynthesis to Heat Stress in Malus. ‘Prairifire’" Plants 9, no. 8: 1040. https://doi.org/10.3390/plants9081040
APA StyleWang, T., Luo, S., Ma, Y., Li, L., Xie, Y., & Zhang, W. (2020). Chlorophyll a Fluorescence Transient and 2-Dimensional Electrophoresis Analyses Reveal Response Characteristics of Photosynthesis to Heat Stress in Malus. ‘Prairifire’. Plants, 9(8), 1040. https://doi.org/10.3390/plants9081040