Genome-Wide Identification and Functional Characterization of the Heat Shock Factor Family in Eggplant (Solanum melongena L.) under Abiotic Stress Conditions

Abstract
1. Introduction
2. Results
2.1. Identification, Classification, and Characterization of the Hsf Gene Family in Eggplant
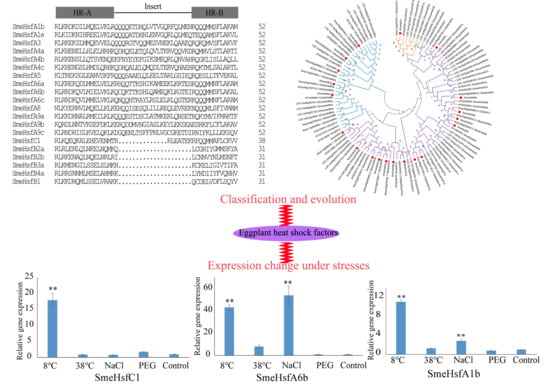
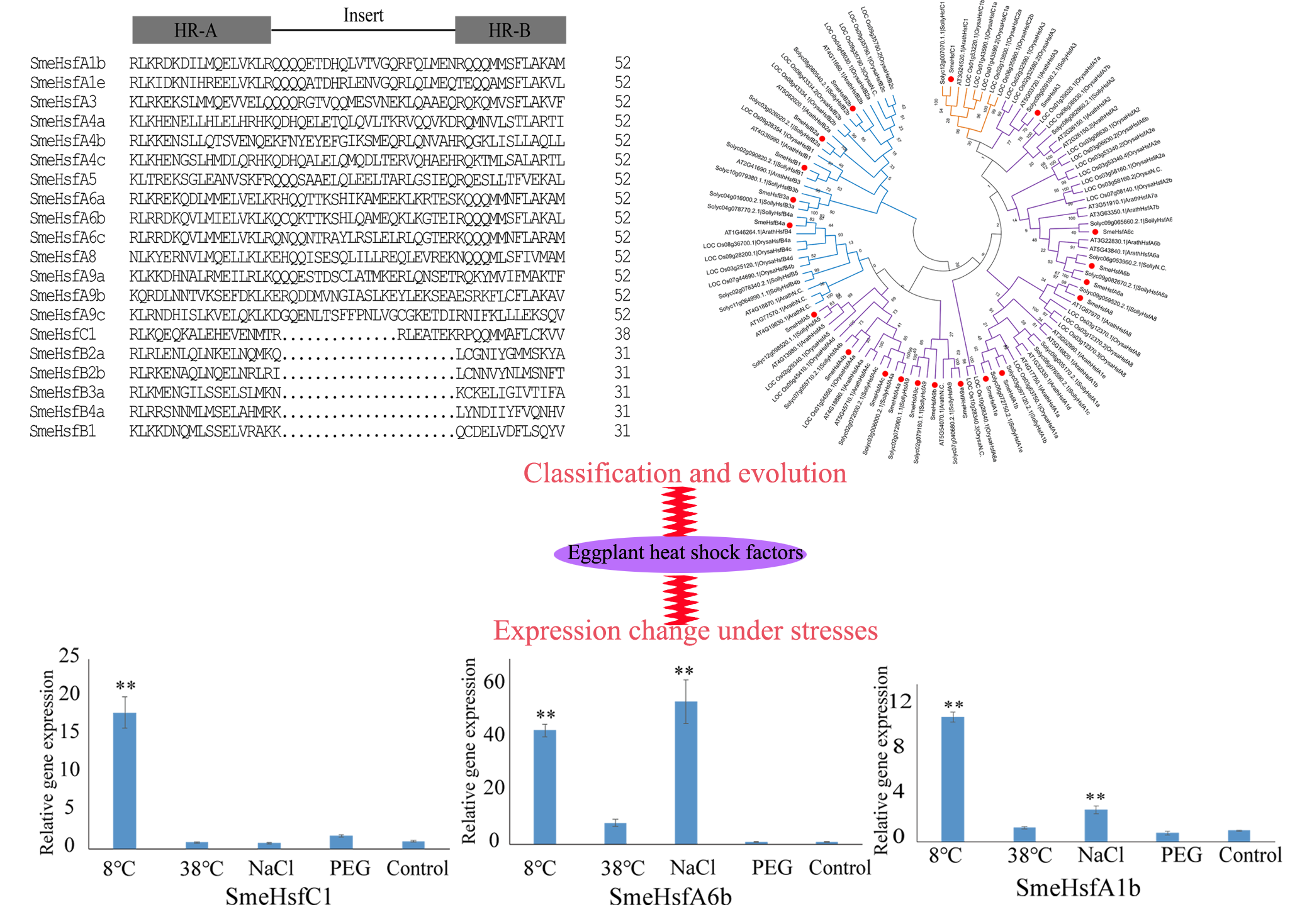
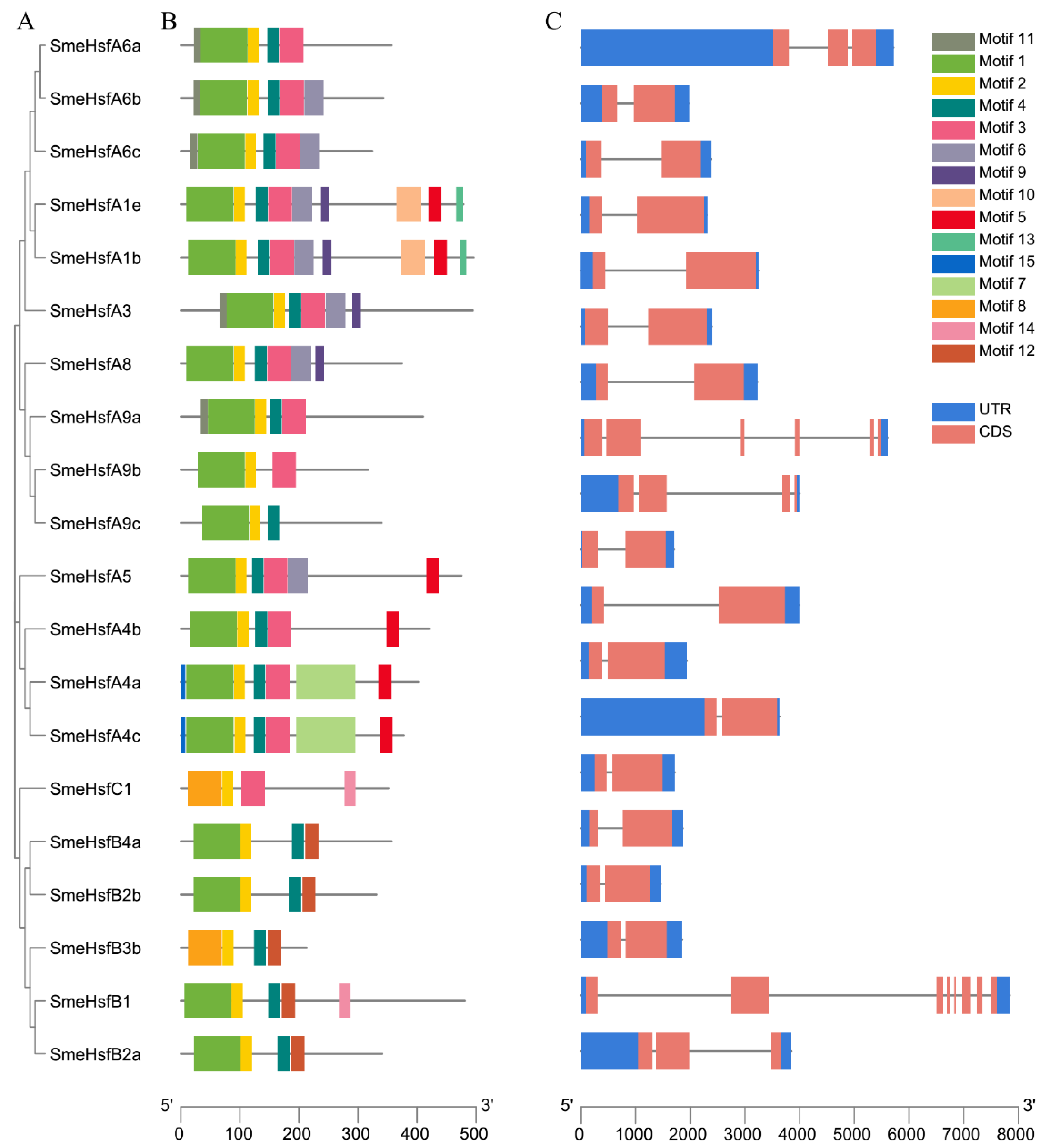
2.2. Conserved Domains and Structural Analysis of SmeHsfs
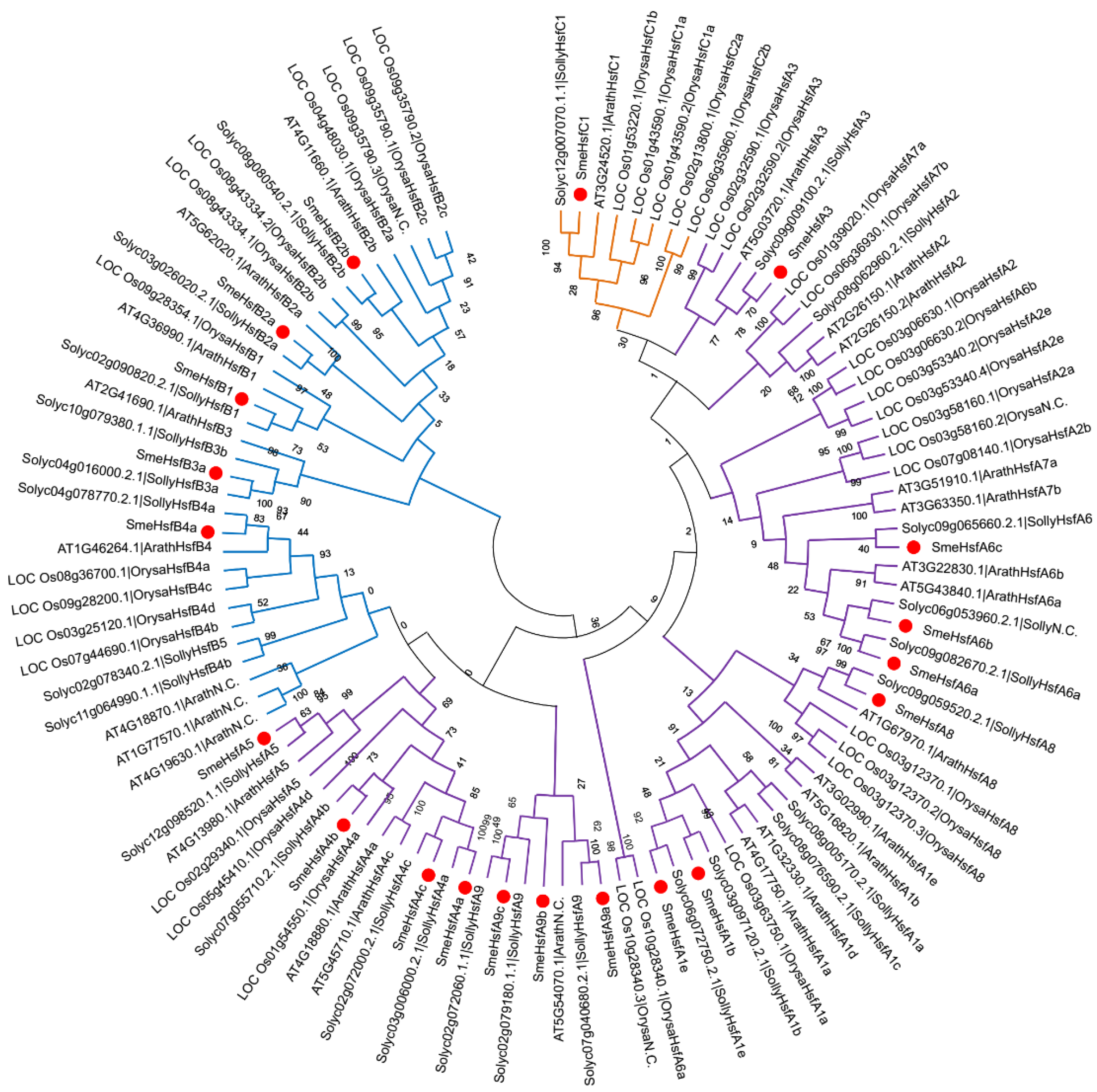
2.3. Phylogenetic Analysis of SmeHsfs
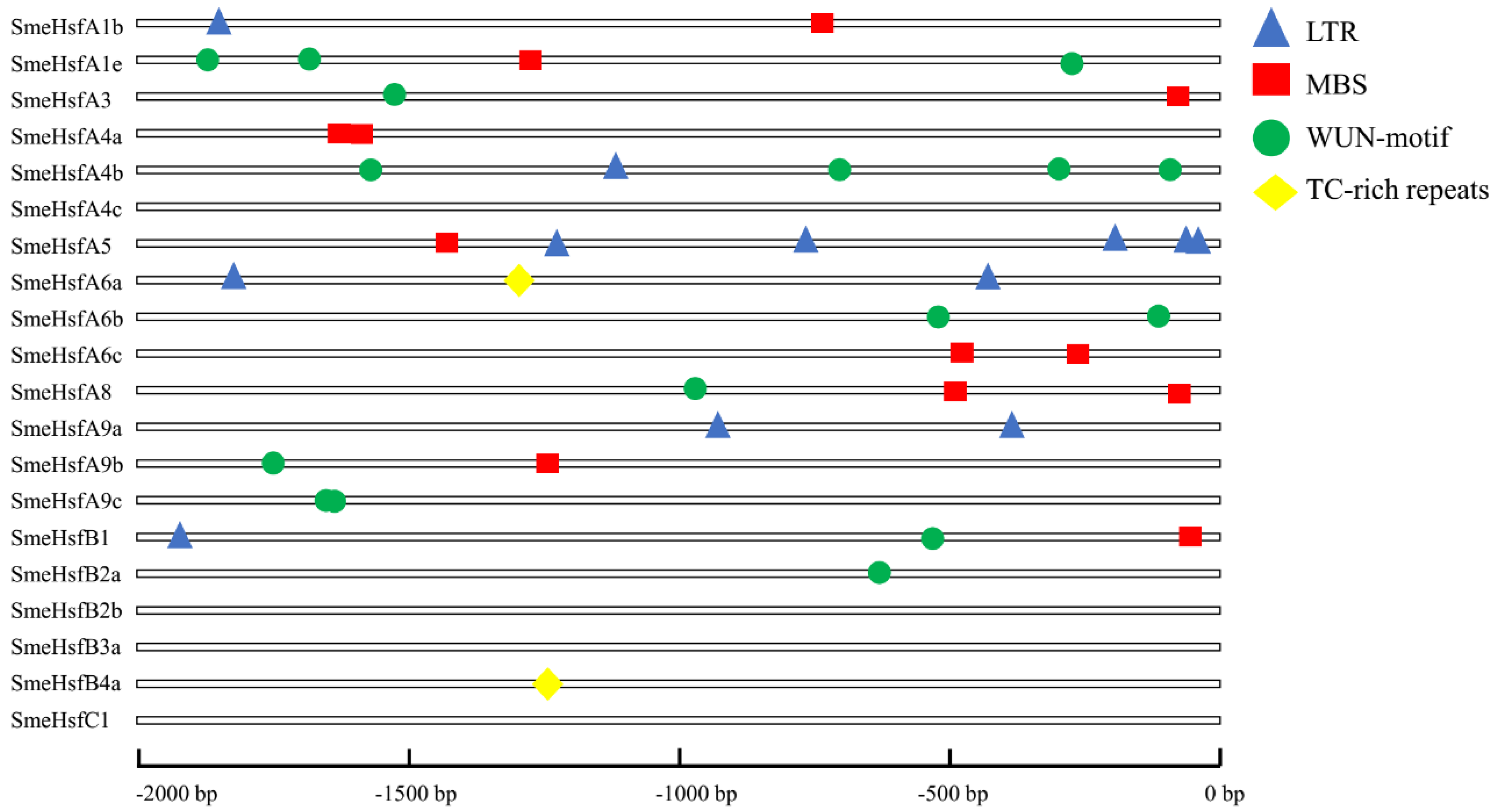
2.4. Putative Regulatory cis-Elements of the SmeHsf Promoters
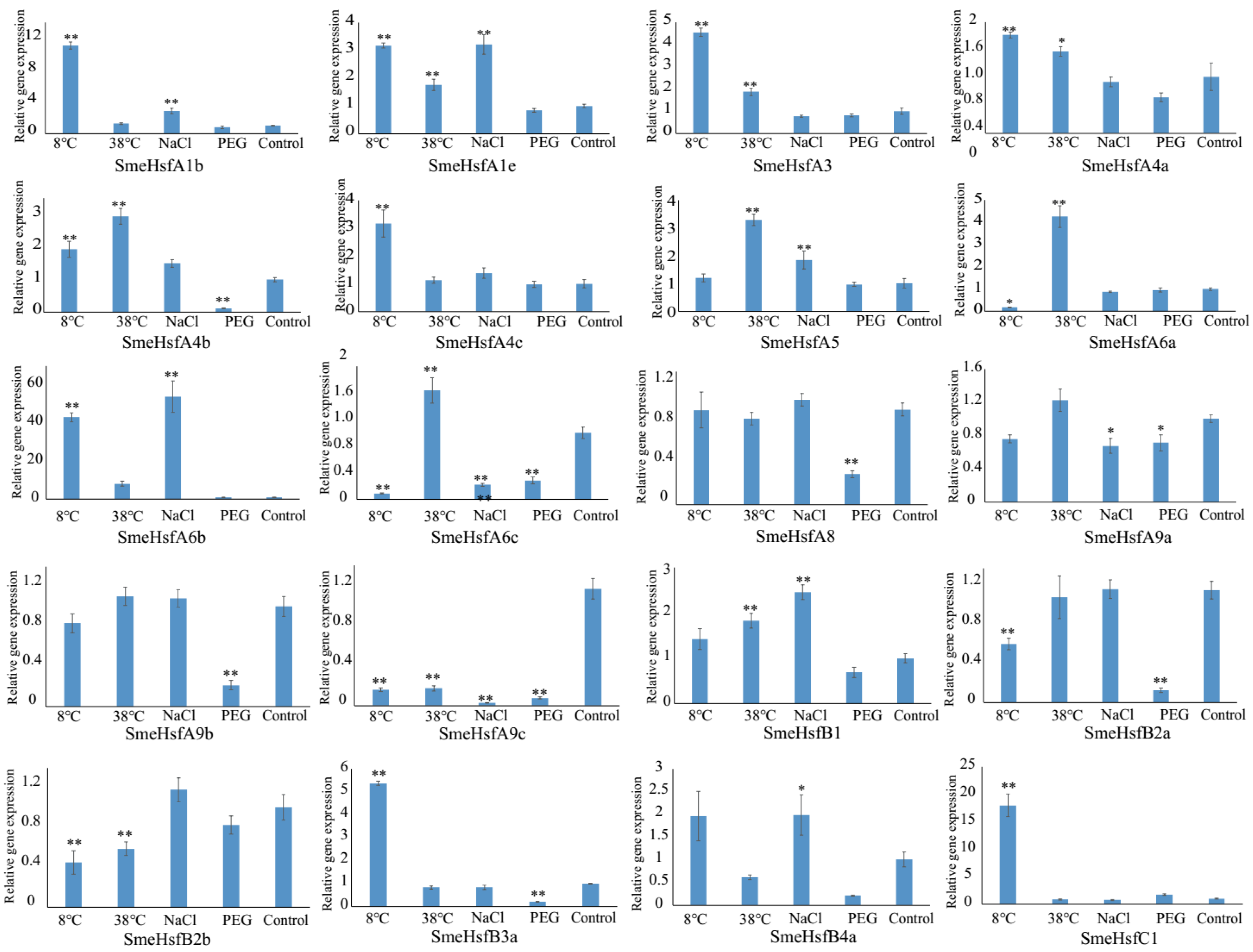
2.5. qRT-PCR Analysis of SmeHsf Responses to Different Abiotic Stresses
3. Discussion
4. Materials and Methods
4.1. Identification and Characterization of Hsfs in Eggplant
4.2. Gene Structure, Domain and Motif Analyses
4.3. Phylogenetic Analysis and Classification of SmeHsf Genes
4.4. cis-Element Analysis of SmeHsf Promoters
4.5. Plant Materials and Stress Treatments
4.6. RNA Extraction and qRT-PCR Analysis
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Wu, C. Heat shock transcription factors: Structure and regulation. Annu. Rev. Cell. Dev. Biol. 1995, 11, 441–469. [Google Scholar] [CrossRef] [PubMed]
- Brash, A.R. Lipoxygenases: Occurrence, functions, catalysis, and acquisition of substrate. J. Biol. Chem. 1999, 274, 23679–23682. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2002, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M.N.V. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Park, M.; Kitazumi, A.; Herath, V.; Mohanty, B.; Yun, S.J.; Reyes, B.G.D.L. Cis-regulatory signatures of orthologous stress-associated bZIP transcription factors from rice, sorghum and Arabidopsis based on phylogenetic footprints. BMC Genom. 2012, 13, 497. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, G.; Garg, V.; Kudapa, H.; Doddamani, D.; Pazhamala, L.T.; Khan, A.W.; Thudi, M.; Lee, S.; Varshney, R.K. Genome-wide dissection of AP2/ERF and HSP90 gene families in five legumes and expression profiles in chickpea and pigeonpea. Plant Biotechnol. J. 2016, 14, 1563–1577. [Google Scholar] [CrossRef]
- Wang, J.; Sun, N.; Deng, T.; Zhang, L.; Zuo, K. Genome-wide cloning, identification, classification and functional analysis of cotton heat shock transcription factors in cotton (Gossypium hirsutum). BMC Genom. 2014, 15, 961. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.; Ma, X.; Luo, D.; Gong, Z.; Lu, M. The plant heat stress transcription factors (HSFs): Structure, regulation, and function in response to abiotic stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef]
- Akerfelt, M.; Morimoto, R.I.; Sistonen, L. Heat shock factors: Integrators of cell stress, development and lifespan. Nat. Rev. Mol. Cell Biol. 2010, 11, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Nover, L.; Bharti, K.; Doring, P.; Mishra, S.K.; Ganguli, A.; Scharf, K. Arabidopsis and the heat stress transcription factor world: How many heat stress transcription factors do we need? Cell Stress Chaperon. 2001, 6, 177–189. [Google Scholar] [CrossRef]
- Bienz, M.; Pelham, H.R.B. Mechanisms of heat-shock gene activation in higher eukaryotes. Adv. Genet. 1987, 24, 31–72. [Google Scholar]
- Damberger, F.F.; Pelton, J.G.; Harrison, C.J.; Nelson, H.C.M.; Wemmer, D.E. Solution structure of the DNA-binding domain of the heat shock transcription factor determined by multidimensional heteronuclear magnetic resonance spectroscopy. Protein Sci. 1994, 3, 1806–1821. [Google Scholar] [CrossRef] [PubMed]
- Vuister, G.W.; Kim, S.; Orosz, A.; Marquardt, J.L.; Wu, C.; Bax, A. Solution structure of the DNA-binding domain of Drosophila heat shock transcription factor. Nat. Struct. Mol. Biol. 1994, 1, 605–614. [Google Scholar] [CrossRef]
- Peteranderl, R.; Rabenstein, M.; Shin, Y.; Liu, C.W.; Wemmer, D.E.; King, D.S.; Nelson, H.C.M. Biochemical and biophysical characterization of the trimerization domain from the heat shock transcription factor. Biochemistry 1999, 38, 3559–3569. [Google Scholar] [CrossRef]
- Scharf, K.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. BBA-Gene Regul. Mech. 2012, 1819, 104–119. [Google Scholar] [CrossRef]
- Baniwal, S.K.; Bharti, K.; Chan, K.Y.; Fauth, M.; Ganguli, A.; Kotak, S.; Mishra, S.K.; Nover, L.; Port, M.; Scharf, K. Heat stress response in plants: A complex game with chaperones and more than twenty heat stress transcription factors. J. Biosci. 2004, 29, 471–487. [Google Scholar] [CrossRef]
- Shim, D.; Hwang, J.; Lee, J.; Lee, S.; Choi, Y.; An, G.; Martinoia, E.; Lee, Y. Orthologs of the class A4 heat shock transcription factor HsfA4a confer cadmium tolerance in wheat and rice. Plant Cell 2009, 21, 4031–4043. [Google Scholar] [CrossRef]
- Ikeda, M.; Mitsuda, N.; Ohmetakagi, M. Arabidopsis HsfB1 and HsfB2b act as repressors of the expression of heat-inducible hsfs but positively regulate the acquired thermotolerance. Plant Physiol. 2011, 157, 1243–1254. [Google Scholar] [CrossRef]
- Ma, J.; Xu, Z.; Wang, F.; Tan, G.; Li, M.; Xiong, A. Genome-wide analysis of HSF family transcription factors and their responses to abiotic stresses in two Chinese cabbage varieties. Acta Physiol. Plant. 2014, 36, 513–523. [Google Scholar] [CrossRef]
- Xue, G.P.; Sadat, S.; Drenth, J.; Mcintyre, C.L. The heat shock factor family from Triticum aestivum in response to heat and other major abiotic stresses and their role in regulation of heat shock protein genes. J. Exp. Bot. 2014, 65, 539–557. [Google Scholar] [CrossRef]
- Ogawa, D.; Yamaguchi, K.; Nishiuchi, T. High-level overexpression of the Arabidopsis HsfA2 gene confers not only increased themotolerance but also salt/osmotic stress tolerance and enhanced callus growth. J. Exp. Bot. 2007, 58, 3373–3383. [Google Scholar] [CrossRef] [PubMed]
- Banti, V.; Mafessoni, F.; Loreti, E.; Alpi, A.; Perata, P. The heat-inducible transcription factor HSFA2 enhances anoxia tolerance in Arabidopsis. Plant Physiol. 2010, 152, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liao, H.; Charng, Y. The role of class A1 heat shock factors (HSFA1s) in response to heat and other stresses in Arabidopsis. Plant Cell Environ. 2011, 34, 738–751. [Google Scholar] [CrossRef]
- Bechtold, U.; Albihlal, W.S.; Lawson, T.; Fryer, M.J.; Sparrow, P.A.C.; Richard, F.; Persad, R.; Bowden, L.; Hickman, R.; Martin, C. Arabidopsis HEAT SHOCK TRANSCRIPTION FACTORA1b overexpression enhances water productivity, resistance to drought, and infection. J. Exp. Bot. 2013, 64, 3467–3481. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Tripp, J.; Winkelhaus, S.; Tschiersch, B.; Theres, K.; Nover, L.; Scharf, K.-D. In the complex family of heat stress transcription factors, HsfA1 has a unique role as master regulator of thermotolerance in tomato. Genes Dev. 2002, 16, 1555–1567. [Google Scholar] [CrossRef]
- Scharf, K.-D.; Heider, H.; Höhfeld, I.; Lyck, R.; Schmidt, E.; Nover, L. The tomato Hsf system: HsfA2 needs interaction with HsfA1 for efficient nuclear import and may be localized in cytoplasmic heat stress granules. Mol. Cell Biol. 1998, 18, 2240–2251. [Google Scholar] [CrossRef]
- Giorno, F.; Woltersarts, M.; Grillo, S.; Scharf, K.; Vriezen, W.H.; Mariani, C. Developmental and heat stress-regulated expression of HsfA2 and small heat shock proteins in tomato anthers. J. Exp. Bot. 2010, 61, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, H.; Zhao, H.; Gao, T.; Song, A.; Jiang, J.; Chen, F.; Chen, S. Chrysanthemum CmHSFA4 gene positively regulates salt stress tolerance in transgenic chrysanthemum. Plant Biotechnol. J. 2018, 16, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Giorno, F.; Guerriero, G.; Baric, S.; Mariani, C. Heat shock transcriptional factors in Malus domestica: Identification, classification and expression analysis. BMC Genom. 2012, 13, 639. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Niu, C.; Yang, C.; Jinn, T. The heat stress factor HSFA6b Connects ABA signaling and ABA-mediated heat responses. Plant Physiol. 2016, 172, 1182–1199. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wu, J.; Ji, Q.; Wang, C.; Luo, L.; Yuan, Y.; Wang, Y.; Wang, J. Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis. J. Genet. Genom. 2008, 35, 105–118. [Google Scholar] [CrossRef]
- Chauhan, H.; Khurana, N.; Agarwal, P.; Khurana, P. Heat shock factors in rice (Oryza sativa L.): Genome-wide expression analysis during reproductive development and abiotic stress. Mol. Genet. Genom. 2011, 286, 171–187. [Google Scholar] [CrossRef]
- Wang, F.; Dong, Q.; Jiang, H.; Zhu, S.; Chen, B.; Xiang, Y. Genome-wide analysis of the heat shock transcription factors in Populus trichocarpa and Medicago truncatula. Mol. Biol. Rep. 2012, 39, 1877–1886. [Google Scholar] [CrossRef]
- Lohani, N.; Golicz, A.A.; Singh, M.; Bhalla, P.L. Genome-wide analysis of the Hsf gene family in Brassica oleracea and a comparative analysis of the Hsf gene family in B. oleracea, B. rapa and B. napus. Funct. Integr. Genom. 2019, 19, 515–531. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Li, Z.; Luo, S.; Sun, B. Effects of heat stress on gene expression in eggplant ( Solanum melongema L.) seedlings. Afr. J. Biotechnol. 2011, 10, 18078–18084. [Google Scholar]
- Berz, J.; Simm, S.; Schuster, S.; Scharf, K.; Schleiff, E.; Ebersberger, I. HEATSTER: A database and web server for identification and classification of heat stress transcription factors in plants. Bioinform. Biol. Insights 2019, 13, 117793221882136. [Google Scholar] [CrossRef]
- Agarwal, P.; Khurana, P. Functional characterization of HSFs from wheat in response to heat and other abiotic stress conditions. Funct. Integr. Genom. 2019, 19, 497–513. [Google Scholar] [CrossRef]
- Duan, S.; Liu, B.; Zhang, Y.; Li, G.; Guo, X. Genome-wide identification and abiotic stress-responsive pattern of heat shock transcription factor family in Triticum aestivum L. BMC Genom. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; De Peer, Y.V.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucl. Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chou, S.; Murshid, A.; Prince, T.L.; Schreiner, S.; Stevenson, M.A.; Calderwood, S.K. The role of heat shock factors in stress-induced transcription. Methods Mol. Biol. 2011, 787, 21–32. [Google Scholar] [PubMed]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant heat stress response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Gho, H.J.; Nguyen, M.X.; Kim, S.-R.; An, G. Genome-wide expression analysis of HSP70 family genes in rice and identification of a cytosolic HSP70 gene highly induced under heat stress. Funct. Integr. Genom. 2013, 13, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Snyman, M.; Cronjé, M. Modulation of heat shock factors accompanies salicylic acid-mediated potentiation of Hsp70 in tomato seedlings. J. Exp. Bot. 2008, 59, 2125–2132. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Pan, S.K.; Cheng, S.F.; Zhang, B.; Bachem, C.W.B.; De Boer, J.M.; Borm, T.J.A.; Kloosterman, B.; Van Eck, H.J.; Datema, E. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar]
- Hirakawa, H.; Shirasawa, K.; Miyatake, K.; Nunome, T.; Negoro, S.; Ohyama, A.; Yamaguchi, H.; Sato, S.; Isobe, S.; Tabata, S. Draft genome sequence of eggplant (Solanum melongena L.): The representative solanum species indigenous to the old world. DNA Res. 2014, 21, 649–660. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.; Kim, Y.; Lee, J.M.; Lee, H.; Seo, E.; Choi, J.Y.; Cheong, K.; Kim, K. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef]
- Sierro, N.; Battey, J.N.D.; Ouadi, S.; Bakaher, N.; Bovet, L.; Willig, A.; Goepfert, S.; Peitsch, M.C.; Ivanov, N.V. The tobacco genome sequence and its comparison with those of tomato and potato. Nat. Commun. 2014, 5, 3833. [Google Scholar] [CrossRef]
- Mittal, D.; Chakrabarti, S.; Sarkar, A.; Singh, A.; Grover, A. Heat shock factor gene family in rice: Genomic organization and transcript expression profiling in response to high temperature, low temperature and oxidative stresses. Plant Physiol. Bioch. 2009, 47, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Dossa, K.; Diouf, D.; Cisse, N. Genome-wide investigation of hsf genes in sesame reveals their segmental duplication expansion and their active role in drought stress response. Front. Plant Sci. 2016, 7, 1522. [Google Scholar] [CrossRef]
- Zhou, M.; Zheng, S.; Liu, R.; Lu, J.; Lu, L.; Zhang, C.; Liu, Z.; Luo, C.; Zhang, L.; Yant, L. Genome-wide identification, phylogenetic and expression analysis of the heat shock transcription factor family in bread wheat (Triticum aestivum L.). BMC Genom. 2019, 20, 505. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jiang, J.; Han, X.; Zhang, Y.; Zhuo, R. Identification, expression analysis of the Hsf family, and characterization of class A4 in Sedum Alfredii hance under cadmium stress. Int. J. Mol. Sci. 2018, 1216. [Google Scholar] [CrossRef]
- Lv, L.L.; Feng, X.F.; Li, W.; Li, K. High temperature reduces peel color in eggplant (Solanum melongena) as revealed by RNA-seq analysis. Genome 2019, 62, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Tian, F.; Yang, D.; Meng, Y.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucl. Acids Res. 2017, 45. [Google Scholar] [CrossRef]
- Elgebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A. The Pfam protein families database in 2019. Nucl. Acids Res. 2019, 47. [Google Scholar]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, 193–196. [Google Scholar] [CrossRef]
- Zimmermann, L.; Stephens, A.; Nam, S.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.; Alva, V. A completely reimplemented MPI bioinformatics toolkit with a new HHpred server at its core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Peter, J.A.C.; Tiago, A.; Jeffrey, T.C.; Brad, A.C.; Cymon, J.C.; Andrew, D.; Iddo, F.; Thomas, H.; Frank, K.; Bartek, W.; et al. Biopython: Freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a Toolkit for Biologists integrating various HTS-data handling tools with a user-friendly interface. bioRxiv 2018, 289660. [Google Scholar]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.C.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucl. Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef]
- Kotak, S.; Port, M.; Ganguli, A.; Bicker, F.; Von Koskulldoring, P. Characterization of C-terminal domains of Arabidopsis heat stress transcription factors (Hsfs) and identification of a new signature combination of plant class A Hsfs with AHA and NES motifs essential for activator function and intracellular localization. Plant J. 2004, 39, 98–112. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Pang, Q.; Li, Z.; Luo, S.; Chen, R.; Jin, Q.; Li, Z.; Li, D.; Sun, B.; Sun, G. Selection and stability analysis of reference gene for qRT-PCR in eggplant under high temperature stress. Acta Hortic. Sin. 2017, 44, 475–486. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Gene Name | Gene Code | Subgroup | Protein Length (aa) | Molecular Weight (kDa) | Aromaticity | Instability Index | Isoelectric Point | GRAVY 1 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | SmeHsfA1b | Sme2.5_02334.1_g00004.1 | A1 | 496 | 54.84 | 0.05 | 52.58 | 5.14 | −0.48 |
| 2 | SmeHsfA1e | Sme2.5_00204.1_g00007.1 | A1 | 478 | 53.81 | 0.06 | 51.96 | 5.88 | −0.61 |
| 3 | SmeHsfA3 | Sme2.5_00292.1_g00007.1 | A3 | 494 | 55.08 | 0.10 | 53.86 | 4.60 | −0.58 |
| 4 | SmeHsfA4a | Sme2.5_01013.1_g00005.1 | A4 | 403 | 46.04 | 0.07 | 42.59 | 5.15 | −0.78 |
| 5 | SmeHsfA4b | Sme2.5_01314.1_g00005.1 | A4 | 421 | 48.36 | 0.09 | 54.15 | 5.35 | −0.74 |
| 6 | SmeHsfA4c | Sme2.5_04312.1_g00009.1 | A4 | 377 | 43.01 | 0.07 | 47.45 | 5.14 | −0.76 |
| 7 | SmeHsfA5 | Sme2.5_09846.1_g00002.1 | A5 | 475 | 53.31 | 0.07 | 57.57 | 5.51 | −0.78 |
| 8 | SmeHsfA6a | Sme2.5_00065.1_g00020.1 | A6 | 357 | 41.70 | 0.10 | 45.43 | 5.15 | −0.93 |
| 9 | SmeHsfA6b | Sme2.5_08000.1_g00008.1 | A6 | 324 | 37.93 | 0.08 | 47.19 | 5.50 | −0.83 |
| 10 | SmeHsfA6c | Sme2.5_04149.1_g00004.1 | A6 | 343 | 39.84 | 0.09 | 59.73 | 5.99 | −0.79 |
| 11 | SmeHsfA8 | Sme2.5_08951.1_g00003.1 | A8 | 374 | 43.27 | 0.10 | 58.33 | 4.72 | −0.62 |
| 12 | SmeHsfA9a | Sme2.5_00023.1_g00025.1 | A9 | 410 | 47.37 | 0.09 | 50.08 | 5.29 | −0.55 |
| 13 | SmeHsfA9b | Sme2.5_03412.1_g00012.1 | A9 | 317 | 36.77 | 0.12 | 41.31 | 9.00 | −0.74 |
| 14 | SmeHsfA9c | Sme2.5_04312.1_g00005.1 | A9 | 340 | 38.48 | 0.07 | 48.87 | 5.93 | −0.86 |
| 15 | SmeHsfB1 | Sme2.5_00010.1_g00004.1 | B1 | 481 | 53.57 | 0.07 | 30.35 | 5.24 | −0.67 |
| 16 | SmeHsfB2a | Sme2.5_13301.1_g00001.1 | B2 | 341 | 38.30 | 0.07 | 60.33 | 6.35 | −0.62 |
| 17 | SmeHsfB2b | Sme2.5_02712.1_g00007.1 | B2 | 331 | 36.21 | 0.06 | 55.45 | 5.18 | −0.44 |
| 18 | SmeHsfB3a | Sme2.5_00159.1_g00006.1 | B3 | 213 | 24.62 | 0.08 | 52.04 | 9.44 | −0.79 |
| 19 | SmeHsfB4a | Sme2.5_01029.1_g00008.1 | B4 | 357 | 40.64 | 0.09 | 60.84 | 7.73 | −0.60 |
| 20 | SmeHsfC1 | Sme2.5_04829.1_g00004.1 | C1 | 352 | 39.42 | 0.09 | 68.53 | 6.14 | −0.65 |
| Gene | DBD | HR-A/B | NLS | NES | AHA |
|---|---|---|---|---|---|
| SmeHsfA1b | 13–106 | 143–196 | (205) NNSKKRRLLVSNY | (150) ILM | Na |
| SmeHsfA1e | 10–103 | 140–193 | (211) ITGMNKKRRFP | Na | Na |
| SmeHsfA3 | 78–171 | 196–249 | Na | Na | Na |
| SmeHsfA4a | 10–103 | 136–189 | Na | Na | (341) DVFWEQFLTE |
| SmeHsfA4b | 16–109 | 139–192 | Na | (236) LEM | (245) INFWERFLYG; (354) DVFWQQFLTE |
| SmeHsfA4c | 10–104 | 136–189 | (203) NDRKRRFPG | Na | (343) DVFWEQFLTE |
| SmeHsfA5 | 13–106 | 133–186 | (202) ISAFSKKRRLP | (193) LAQKLESMDI | (422) DVFWEQFLTE |
| SmeHsfA6a | 34–127 | 159–212 | (220) EIRNKRKRQID | Na | (314) EGFWEDLLNE |
| SmeHsfA6b | 29–122 | 153–206 | (115) LLRTIKRRKTTNF; (226) EINKKRRRPID | (265) VALNM | Na |
| SmeHsfA6c | 33–126 | 160–213 | (119) LLRNIKRRKTP; (222) QQKGKRKEIEEDITKKRRQPI | (191) LRL | (305) MGFWEELFND |
| SmeHsfA8 | 10–103 | 138–191 | Na | Na | Na |
| SmeHsfA9a | 46–139 | 164–217 | (134) INIKRRKQYP; (229) KQGKKRKLCDAQF | Na | Na |
| SmeHsfA9b | 29–122 | 147–200 | Na | Na | Na |
| SmeHsfA9c | 36–129 | 160–213 | (219) DTRKRPCLV | Na | (269) REFWEKLFED |
| SmeHsfB1 | 6–99 | 161–193 | (254) KEKKKKRGPD | Na | Na |
| SmeHsfB2a | 22–115 | 177–209 | Na | Na | Na |
| SmeHsfB2b | 21–114 | 196–228 | Na | Na | Na |
| SmeHsfB3a | 2–84 | 137–169 | (190) EMERKRKRVEL | Na | Na |
| SmeHsfB4a | 21–114 | 201–233 | (203) NERKRRLPG | (342) LEKNDLGL | Na |
| SmeHsfC1 | 1–83 | 108–147 | (169) REKKRRLMIS | (155) LMEKERSKRLSL | Na |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Hu, H.; Wang, W.; Wei, Q.; Hu, T.; Bao, C. Genome-Wide Identification and Functional Characterization of the Heat Shock Factor Family in Eggplant (Solanum melongena L.) under Abiotic Stress Conditions. Plants 2020, 9, 915. https://doi.org/10.3390/plants9070915
Wang J, Hu H, Wang W, Wei Q, Hu T, Bao C. Genome-Wide Identification and Functional Characterization of the Heat Shock Factor Family in Eggplant (Solanum melongena L.) under Abiotic Stress Conditions. Plants. 2020; 9(7):915. https://doi.org/10.3390/plants9070915
Chicago/Turabian StyleWang, Jinglei, Haijiao Hu, Wuhong Wang, Qingzhen Wei, Tianhua Hu, and Chonglai Bao. 2020. "Genome-Wide Identification and Functional Characterization of the Heat Shock Factor Family in Eggplant (Solanum melongena L.) under Abiotic Stress Conditions" Plants 9, no. 7: 915. https://doi.org/10.3390/plants9070915
APA StyleWang, J., Hu, H., Wang, W., Wei, Q., Hu, T., & Bao, C. (2020). Genome-Wide Identification and Functional Characterization of the Heat Shock Factor Family in Eggplant (Solanum melongena L.) under Abiotic Stress Conditions. Plants, 9(7), 915. https://doi.org/10.3390/plants9070915