Canthaxanthin, a Red-Hot Carotenoid: Applications, Synthesis, and Biosynthetic Evolution



Abstract
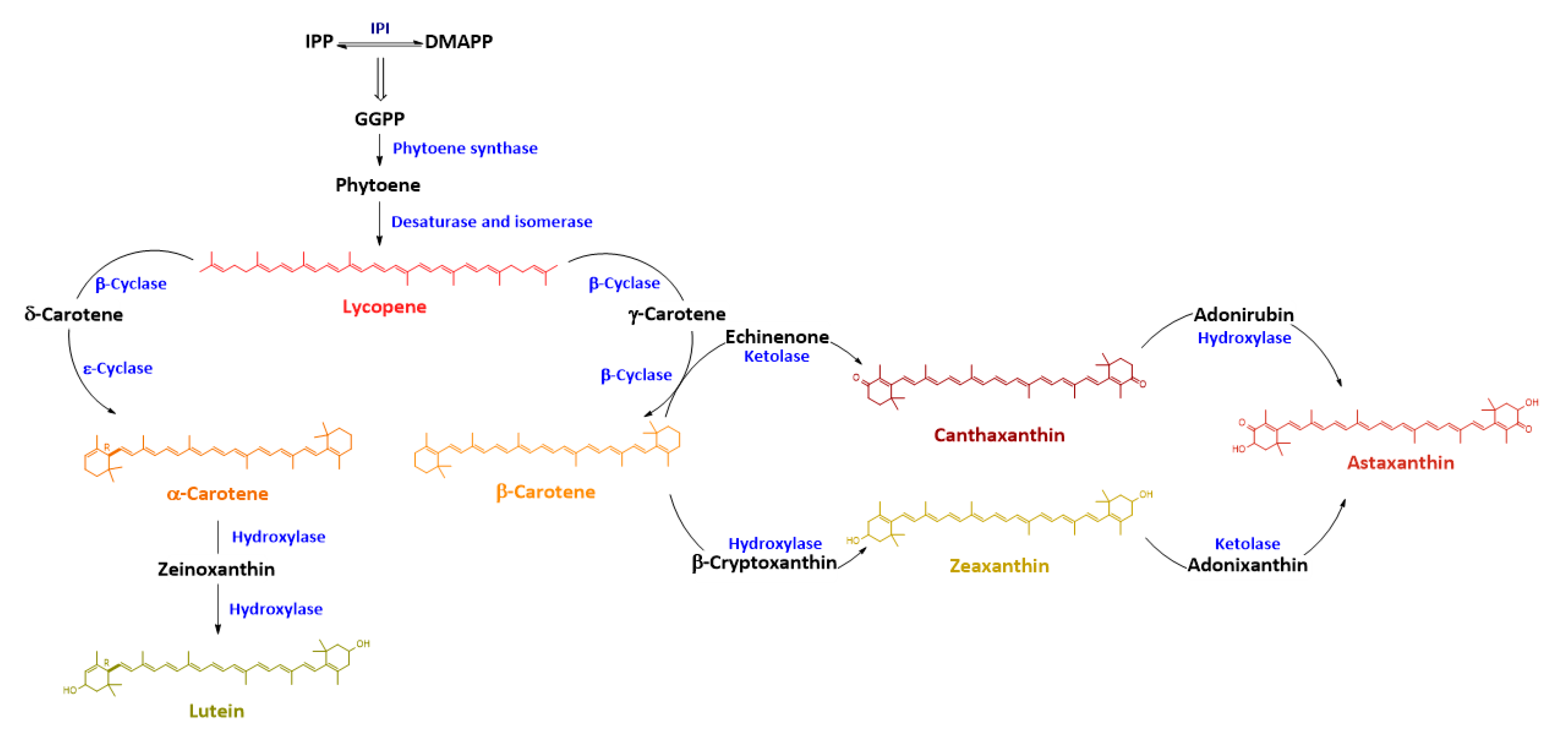
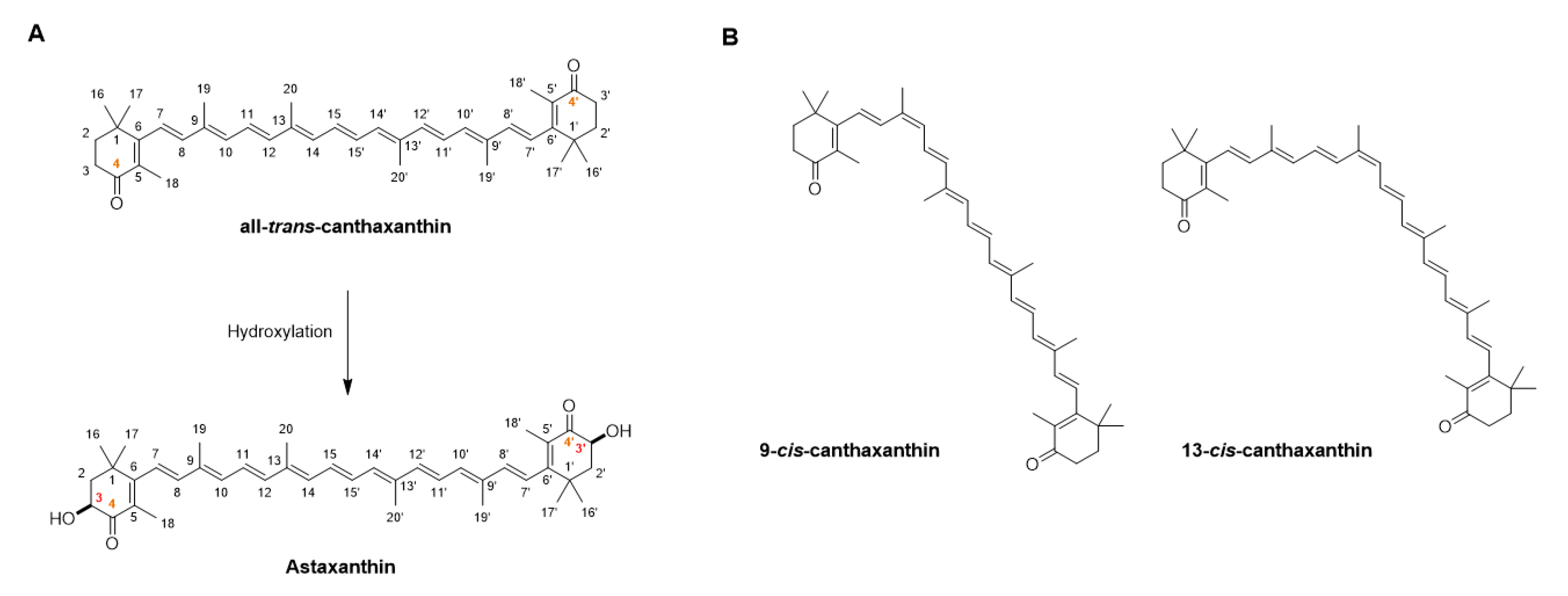
1. Introduction: Characterization and Biosynthesis of Canthaxanthin
2. Economic Value of Ketocarotenoids and Market Potential
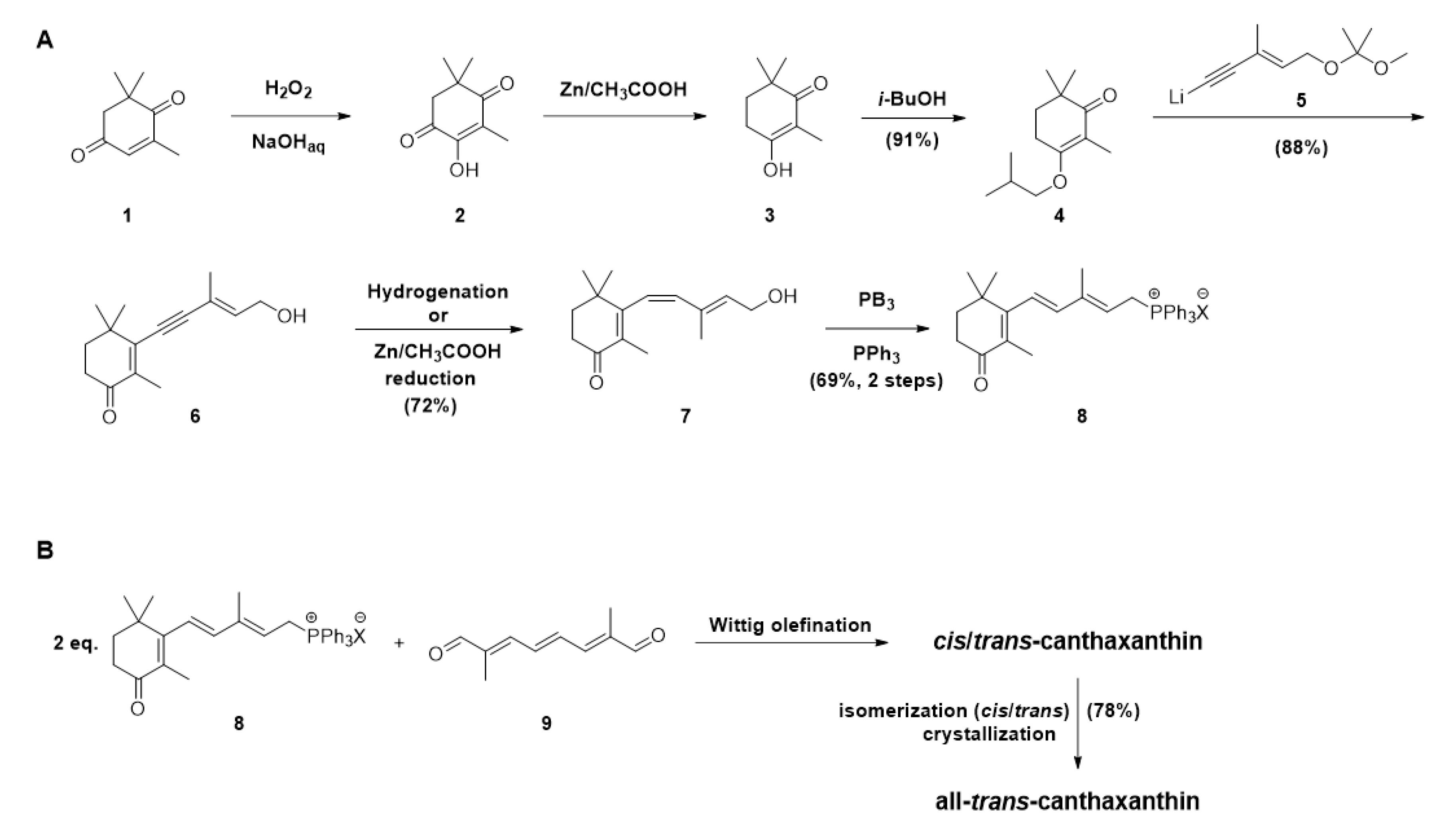
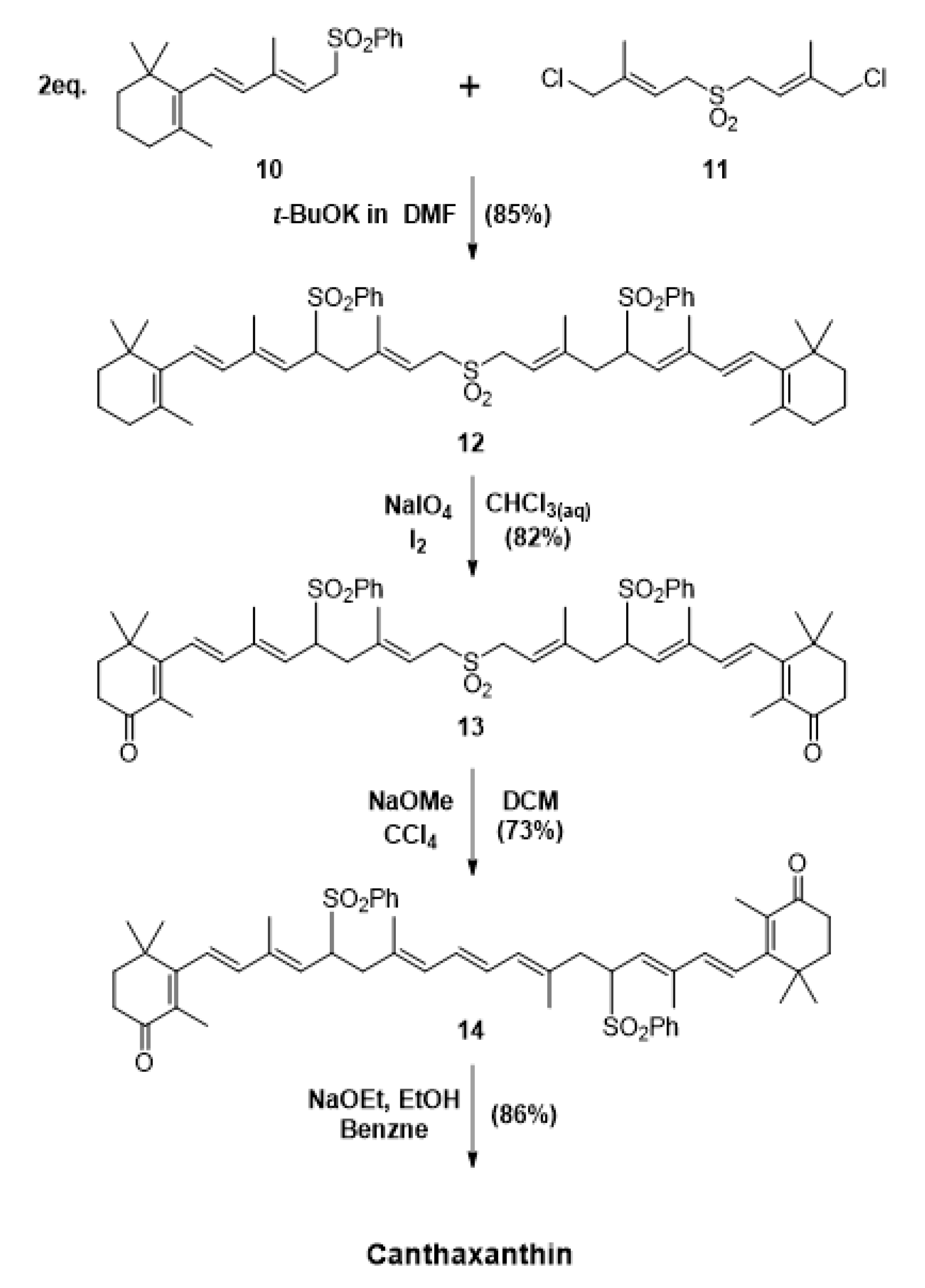
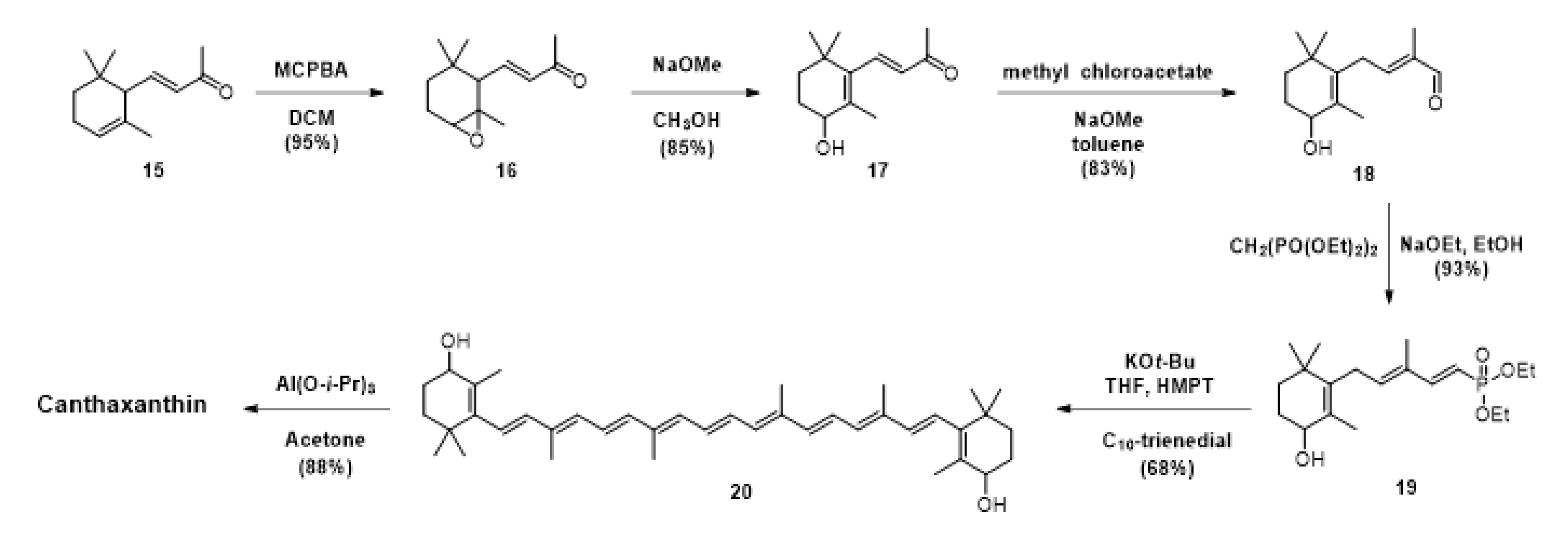
3. Chemical Synthesis of Canthaxanthin, the Main Approach
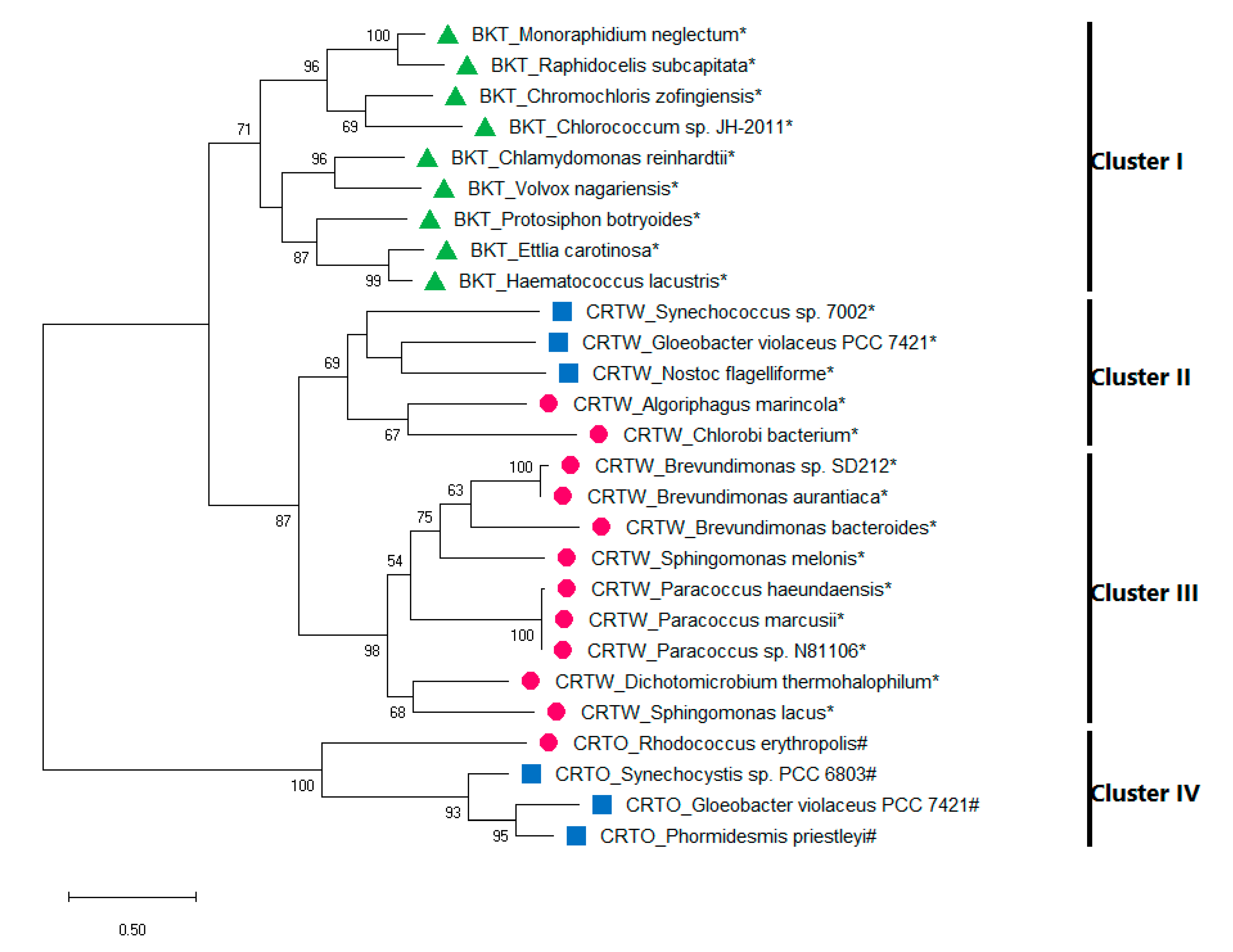
4. Evolution of Canthaxanthin Biosynthesis
5. Canthaxanthin Production in Heterologous Systems
6. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ruiz-Sola, M.Á.; Rodríguez-Concepción, M. Carotenoid biosynthesis in Arabidopsis: A colorful pathway. Arab. Book 2012, 10, e0158. [Google Scholar] [CrossRef]
- Wang, C.; Kim, J.H.; Kim, S.W. Synthetic biology and metabolic engineering for marine carotenoids: New opportunities and future prospects. Mar. Drugs 2014, 12, 4810–4832. [Google Scholar] [CrossRef]
- Zhu, C.; Naqvi, S.; Capell, T.; Christou, P. Metabolic engineering of ketocarotenoid biosynthesis in higher plants. Arch. Biochem. Biophys. 2009, 483, 182–190. [Google Scholar] [CrossRef]
- Mortimer, C.L.; Misawa, N.; Ducreux, L.; Campbell, R.; Bramley, P.M.; Taylor, M.; Fraser, P.D. Product stability and sequestration mechanisms in Solanum tuberosum engineered to biosynthesize high value ketocarotenoids. Plant Biotechnol. J. 2016, 14, 140–152. [Google Scholar] [CrossRef]
- Phillip, D.M.; Young, A.J. Preferential inhibition of the lycopene ε-cyclase by the substituted triethylamine compound MPTA in higher plants. J. Plant Physiol. 2006, 163, 383–391. [Google Scholar] [CrossRef]
- Cardoso, L.A.C.; Karp, S.G.; Vendruscolo, F.; Kanno, K.Y.F.; Zoz, L.I.C.; Carvalho, J.C. Biotechnological production of carotenoids and their applications in food and pharmaceutical products. In Carotenoids; Cvetkovic, D., Nikolic, G., Eds.; IntechOpen: London, UK, 2017; Volume 8, pp. 125–141. [Google Scholar]
- Cunningham, F.X.; Gantt, E. Elucidation of the Pathway to Astaxanthin in the Flowers of Adonis aestivalis. Plant Cell 2011, 23, 3055–3069. [Google Scholar] [CrossRef]
- Zakynthinos, G.; Varzakas, T. Carotenoids: From plants to food industry. Curr. Res. Nutr. Food Sci. 2016, 4, 38–51. [Google Scholar] [CrossRef]
- Venugopalan, V.; Tripathi, S.K.; Nahar, P.; Saradhi, P.P.; Das, R.H.; Gautam, H.K. Characterization of canthaxanthin isomers isolated from a new soil Dietzia sp. and their antioxidant activities. J. Microbiol. Biotechnol. 2013, 23, 237–245. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Britton, G.; Vicario, I.M.; Heredia, F.J. Relationship between the colour and the chemical structure of carotenoid pigments. Food Chem. 2007, 101, 1145–1150. [Google Scholar] [CrossRef]
- Qiu, D.; Zhu, W.L.; Tang, C.K.; Shi, L.F.; Gao, H.Q. Identification of the Composition of Isomeric Canthaxanthin Sample by NMR, HPLC, and Mass Spectrometry. Food Anal. Methods 2014, 7, 597–605. [Google Scholar] [CrossRef]
- Hashimoto, H.; Koyama, Y.; Shimamura, T. Isolation of cis-trans isomers of canthaxanthin by high-performance liquid chromatography using a calcium hydroxide column and identification of their configurations by 1H NMR spectroscopy. J. Chromatogr. A 1988, 448, 182–187. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef]
- Fraser, P.D.; Bramley, P.M. The biosynthesis and nutritional uses of carotenoids. Prog. Lipid Res. 2004, 43, 228–265. [Google Scholar] [CrossRef]
- Martín, J.F.; Gudiña, E.; Barredo, J.L. Conversion of β-carotene into astaxanthin: Two separate enzymes or a bifunctional hydroxylase-ketolase protein? Microb. Cell Fact. 2008, 7, 3. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Rimbach, G. Canthaxanthin: From molecule to function. Mol. Nutr. Food Res. 2017, 61, 1–17. [Google Scholar] [CrossRef]
- Fernández-García, E.; Carvajal-Lérida, I.; Jarén-Galán, M.; Garrido-Fernández, J.; Pérez-Gálvez, A.; Hornero-Méndez, D. Carotenoids bioavailability from foods: From plant pigments to efficient biological activities. Food Res. Int. 2012, 46, 438–450. [Google Scholar] [CrossRef]
- De Miguel, T.; Sieiro, C.; Poza, M.; Villa, T.G. Analysis of canthaxanthin and related pigments from Gordonia jacobaea mutants. J. Agric. Food Chem. 2001, 49, 1200–1202. [Google Scholar] [CrossRef]
- Abe, K.; Hattori, H.; Hirano, M. Accumulation and antioxidant activity of secondary carotenoids in the aerial microalga Coelastrella striolata var. multistriata. Food Chem. 2007, 100, 656–661. [Google Scholar] [CrossRef]
- Yeum, K.-J.; Russell, R.M. Carotenoid bioavailability and bioconversion. Annu. Rev. Nutr. 2002, 22, 483–504. [Google Scholar] [CrossRef]
- Perera, C.O.; Yen, G.M. Functional properties of carotenoids in human health. Int. J. Food Prop. 2007, 10, 201–230. [Google Scholar] [CrossRef]
- Reboul, E. Mechanisms of Carotenoid Intestinal Absorption: Where Do We Stand? Nutrients 2019, 11, 838. [Google Scholar] [CrossRef]
- Cheng, X.; Ku, C.H.; Siow, R.C.M. Regulation of the Nrf2 antioxidant pathway by microRNAs: New players in micromanaging redox homeostasis. Free Radic. Biol. Med. 2013, 64, 4–11. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, e17023. [Google Scholar] [CrossRef]
- Barros, M.P.; Rodrigo, M.J.; Zacarias, L. Dietary Carotenoid Roles in Redox Homeostasis and Human Health. J. Agric. Food Chem. 2018, 66, 5733–5740. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Cantrell, A.; McGarvey, D.J.; Truscott, T.G.; Rancan, F.; Böhm, F. Singlet oxygen quenching by dietary carotenoids in a model membrane environment. Arch. Biochem. Biophys. 2003, 412, 47–54. [Google Scholar] [CrossRef]
- Gradelet, S.; Le Bon, A.M.; Bergès, R.; Suschetet, M.; Astorg, P. Dietary carotenoids inhibit aflatoxin B1-induced liver preneoplastic foci and DNA damage in the rat: Role of the modulation of aflatoxin B1 metabolism. Carcinogenesis 1998, 19, 403–411. [Google Scholar] [CrossRef]
- Elia, A.C.; Prearo, M.; Dörr, A.J.M.; Pacini, N.; Magara, G.; Brizio, P.; Gasco, L.; Abete, M.C. Effects of astaxanthin and canthaxanthin on oxidative stress biomarkers in rainbow trout. J. Toxicol. Environ. Health Part A Curr. Issues 2019, 82, 760–768. [Google Scholar] [CrossRef]
- Palozza, P.; Krinsky, N.I. Astaxanthin and canthaxanthin are potent antioxidants in a membrane model. Arch. Biochem. Biophys. 1992, 297, 291–295. [Google Scholar] [CrossRef]
- Sujak, A. Interactions between canthaxanthin and lipid membranes—Possible mechanisms of canthaxanthin toxicity. Cell. Mol. Biol. Lett. 2009, 14, 395–410. [Google Scholar] [CrossRef]
- Carotenoids Market by Product, Source, and Application: Global Opportunity Analysis and Industry Forecast, 2018–2025. Available online: www.alliedmarketresearch.com/press-release/carotenoids-market.htm (accessed on 10 July 2020).
- Carotenoids Market Size by Source (Natural, Synthetic), by Product (Beta-Carotene, Lutein, Astaxanthin, Canthaxanthin, Lycopene), by Application (Food & Beverages, Pharmaceutical, Cosmetics, Animal Feed, Dietary Supplements), Industry Analysis Report, Reg. Available online: https://www.gminsights.com/industry-analysis/carotenoids-market (accessed on 7 August 2020).
- Canthaxanthin Market Size by Source (Synthetic, Natural), by Application (Dietary Supplement, Personal Care, Pharmaceuticals, Food & Beverages, Animal Feed {Aquaculture, Livestock, Pets}), Industry Analysis Report, Regional Outlook, Growth Potential, Pric. Available online: https://www.gminsights.com/industry-analysis/canthaxanthin-market (accessed on 7 August 2020).
- Global Canthaxanthin Market to Exceed $85 mn by 2024. Available online: https://www.gminsights.com/pressrelease/canthaxanthin-market (accessed on 7 August 2020).
- Haxo, F. Carotenoids of the Mushroom Cantharellus cinnabarinus. Bot. Gaz. 1950, 112, 228–232. [Google Scholar] [CrossRef]
- Krupa, D.; Nakkeeran, E.; Kumaresan, N.; Vijayalakshmi, G.; Subramanian, R. Extraction, purification and concentration of partially saturated canthaxanthin from Aspergillus carbonarius. Bioresour. Technol. 2010, 101, 7598–7604. [Google Scholar] [CrossRef]
- Parajó, J.C.; Santos, V.; Vázquez, M. Optimization of carotenoid production by Phaffia rhodozyma cells grown on xylose. Process Biochem. 1998, 33, 181–187. [Google Scholar] [CrossRef]
- Barredo, J.; García-Estrada, C.; Kosalkova, K.; Barreiro, C. Biosynthesis of Astaxanthin as a Main Carotenoid in the Heterobasidiomycetous Yeast Xanthophyllomyces dendrorhous. J. Fungi 2017, 3, 44. [Google Scholar] [CrossRef]
- Singh, D.P.; Khattar, J.S.; Rajput, A.; Chaudhary, R.; Singh, R. High production of carotenoids by the green microalga Asterarcys quadricellulare PUMCC 5.1.1 under optimized culture conditions. PLoS ONE 2019, 14, e0221930. [Google Scholar] [CrossRef]
- Grung, M.; Metzger, P.; Liaaen-jensen, S. Primary and secondary carotenoids in two races of the green alga Botryococcus braunii. Biochem. Syst. Ecol. 1989, 17, 263–269. [Google Scholar] [CrossRef]
- Malis, S.A.; Cohen, E.; Ben Amotz, A. Accumulation of canthaxanthin in Chlorella emersonii. Physiol. Plant. 1993, 87, 232–236. [Google Scholar] [CrossRef]
- Gouveia, L.; Veloso, V.; Reis, A.; Fernandes, H.; Novais, J.; Empis, J. Evolution of pigment composition in Chlorella vulgaris. Bioresour. Technol. 1996, 57, 157–159. [Google Scholar] [CrossRef]
- Bar, E.; Rise, M.; Vishkautsan, M.; Arad, S. (Malis) Pigment and Structural Changes in Chlorella zofingiensis upon Light and Nitrogen Stress. J. Plant Physiol. 1995, 146, 527–534. [Google Scholar] [CrossRef]
- Li, H.-B.; Fan, K.-W.; Chen, F. Isolation and purification of canthaxanthin from the microalga Chlorella zofingiensis by high-speed counter-current chromatography. J. Sep. Sci. 2006, 29, 699–703. [Google Scholar] [CrossRef]
- Yuan, J.P.; Chen, F.; Liu, X.; Li, X.Z. Carotenoid composition in the green microalga Chlorococcum. Food Chem. 2002, 76, 319–325. [Google Scholar] [CrossRef]
- Grama, B.S.; Chader, S.; Khelifi, D.; Agathos, S.N.; Jeffryes, C. Induction of canthaxanthin production in a Dactylococcus microalga isolated from the Algerian Sahara. Bioresour. Technol. 2014, 151, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Accadia Di, F.; Gribanovski-Sassu, O.; Romagnoli, A.; Tuttobello, L. Isolation and identification of carotenoids produced by a green alga (Dictyococcus cinnabarinus) in submerged culture. Biochem. J. 1966, 101, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Choubert, G.; Heinrich, O. Carotenoid pigments of the green alga Haematococcus pluvialis: Assay on rainbow trout, Oncorhynchus mykiss, pigmentation in comparison with synthetic astaxanthin and canthaxanthin. Aquaculture 1993, 112, 217–226. [Google Scholar] [CrossRef]
- Nobre, B.; Marcelo, F.; Passos, R.; Beirão, L.; Palavra, A.; Gouveia, L.; Mendes, R. Supercritical carbon dioxide extraction of astaxanthin and other carotenoids from the microalga Haematococcus pluvialis. Eur. Food Res. Technol. 2006, 223, 787–790. [Google Scholar] [CrossRef]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Lubián, L.M.; Montero, O.; Moreno-Garrido, I.; Huertas, I.E.; Sobrino, C.; González-Del Valle, M.; Parés, G. Nannochloropsis (Eustigmatophyceae) as source of commercially valuable pigments. J. Appl. Phycol. 2000, 12, 249–255. [Google Scholar] [CrossRef]
- Qin, S.; Liu, G.X.; Hu, Z.Y. The accumulation and metabolism of astaxanthin in Scenedesmus obliquus (Chlorophyceae). Process Biochem. 2008, 43, 795–802. [Google Scholar] [CrossRef]
- Lorquin, J.; Molouba, F.; Dreyfus, B.L. Identification of the carotenoid pigment canthaxanthin from photosynthetic Bradyrhizobium strains. Appl. Environ. Microbiol. 1997, 63, 1151–1154. [Google Scholar] [CrossRef]
- Nelis, H.J.; De Leenheer, P. Reinvestigation of Brevibacterium sp. Strain KY-4313 as a Source of Canthaxanthin. Appl. Environ. Microbiol. 1989, 55, 2505–2510. [Google Scholar] [CrossRef]
- Nasrabadi, M.R.N.; Razavi, S.H. Enhancement of canthaxanthin production from Dietzia natronolimnaea HS-1 in a fed-batch process using trace elements and statistical methods. Braz. J. Chem. Eng. 2010, 27, 517–529. [Google Scholar] [CrossRef]
- Hojjati, M.; Razavi, S.H.; Rezaei, K.; Gilani, K. Stabilization of canthaxanthin produced by Dietzia natronolimnaea HS-1 with spray drying microencapsulation. J. Food Sci. Technol. 2014, 51, 2134–2140. [Google Scholar] [CrossRef] [PubMed]
- Cooney, J.J.; Marks, H.W.; Smith, A.M. Isolation and Identification of Canthaxanthin from Micrococcus roseus. J. Bacteriol. 1966, 92, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Ungers, G.E.; Cooney, J.J. Isolation and characterization of carotenoid pigments of Micrococcus roseus. J. Bacteriol. 1968, 96, 234–241. [Google Scholar] [CrossRef]
- Takaichi, S.; Maoka, T.; Akimoto, N.; Khan, S.T.; Harayama, S. Major carotenoid isolated from Paracoccus schoinia NBRC 100637T is adonixanthin diglucoside. J. Nat. Prod. 2006, 69, 1823–1825. [Google Scholar] [CrossRef]
- Asker, D.; Ohta, Y. Production of canthaxanthin by extremely halophilic bacteria. J. Biosci. Bioeng. 1999, 88, 617–621. [Google Scholar] [CrossRef]
- Asker, D.; Ohta, Y. Haloferax alexandrinus sp. nov., an extremely halophilic canthaxanthin-producing archaeon from a solar saltern in Alexandria (Egypt). Int. J. Syst. Evol. Microbiol. 2002, 52, 729–738. [Google Scholar] [CrossRef]
- Breithaupt, D.E. Modern application of xanthophylls in animal feeding—A review. Trends Food Sci. Technol. 2007, 18, 501–506. [Google Scholar] [CrossRef]
- Widmer, E. Synthetic advances in the carotenoid field. Pure Appl. Chem. 1985, 57, 741–752. [Google Scholar] [CrossRef]
- Rosenberger, M.; McDougal, P.; Bahr, J. Canthaxanthin. A new total synthesis. J. Org. Chem. 1982, 47, 2130–2134. [Google Scholar] [CrossRef]
- Choi, S.; Koo, S.; Pure, J.; Chem, A. Efficient syntheses of the Keto-carotenoids Canthaxanthin, Astaxanthin, and Astacene. J. Org. Chem. 2005, 70, 3328–3331. [Google Scholar] [CrossRef] [PubMed]
- Pi, S.; Xi, M.; Deng, L.; Xu, H.; Feng, C.; Shen, R.; Wu, C. Practical synthesis of canthaxanthin. J. Iran. Chem. Soc. 2020, 17, 493–497. [Google Scholar] [CrossRef]
- Kim, J.E.; Cheng, K.M.; Craft, N.E.; Hamberger, B.; Douglas, C.J. Over-expression of Arabidopsis thaliana carotenoid hydroxylases individually and in combination with a β-carotene ketolase provides insight into in vivo functions. Phytochemistry 2010, 71, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, G. Plant carotenoids: Genomics meets multi-gene engineering. Curr. Opin. Plant Biol. 2014, 19, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Falchi, R.; Vendramin, E.; Zanon, L.; Scalabrin, S.; Cipriani, G.; Verde, I.; Vizzotto, G.; Morgante, M. Three distinct mutational mechanisms acting on a single gene underpin the origin of yellow flesh in peach. Plant J. 2013, 76, 175–187. [Google Scholar] [CrossRef]
- Pierce, E.C.; LaFayette, P.R.; Ortega, M.A.; Joyce, B.L.; Kopsell, D.A.; Parrott, W.A. Ketocarotenoid production in soybean seeds through metabolic engineering. PLoS ONE 2015, 10, e0138196. [Google Scholar] [CrossRef]
- Zhou, P.; Ye, L.; Xie, W.; Lv, X.; Yu, H. Highly efficient biosynthesis of astaxanthin in Saccharomyces cerevisiae by integration and tuning of algal crtZ and bkt. Appl. Microbiol. Biotechnol. 2015, 99, 8419–8428. [Google Scholar] [CrossRef]
- Zhou, P.; Li, M.; Shen, B.; Yao, Z.; Bian, Q.; Ye, L.; Yu, H. Directed coevolution of β-Carotene Ketolase and Hydroxylase and its application in temperature-regulated biosynthesis of Astaxanthin. J. Agric. Food Chem. 2019, 67, 1072–1080. [Google Scholar] [CrossRef]
- Walter, M.H.; Strack, D. Carotenoids and their cleavage products: Biosynthesis and functions. Nat. Prod. Rep. 2011, 28, 663–692. [Google Scholar] [CrossRef]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Marechal-Drouard, L.; et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef]
- Perozeni, F.; Cazzaniga, S.; Baier, T.; Zanoni, F.; Zoccatelli, G.; Lauersen, K.J.; Wobbe, L.; Ballottari, M. Turning a green alga red: Engineering astaxanthin biosynthesis by intragenic pseudogene revival in Chlamydomonas reinhardtii. Plant Biotecnhol. J. 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Scaife, M.A.; Prince, C.A.; Norman, A.; Armenta, R.E. Progress toward an Escherichia coli canthaxanthin bioprocess. Process Biochem. 2012, 47, 2500–2509. [Google Scholar] [CrossRef]
- Choi, S.K.; Nishida, Y.; Matsuda, S.; Adachi, K.; Kasai, H.; Peng, X.; Komemushi, S.; Miki, W.; Misawa, N. Characterization of β-carotene ketolases, CrtW, from marine bacteria by complementation analysis in Escherichia coli. Mar. Biotechnol. 2005, 7, 515–522. [Google Scholar] [CrossRef]
- Gao, X.; Xu, H.; Zhu, Z.; She, Y.; Ye, S. Improved production of echinenone and canthaxanthin in transgenic Nostoc sp. PCC 7120 overexpressing a heterologous crtO gene from Nostoc flagelliforme. Microbiol. Res. 2020, 236, 126455. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, M.; Enfissi, E.M.A.A.; Welsch, R.; Beyer, P.; Zurbriggen, M.D.; Fraser, P.D. Construction of a fusion enzyme for astaxanthin formation and its characterisation in microbial and plant hosts: A new tool for engineering ketocarotenoids. Metab. Eng. 2019, 52, 243–252. [Google Scholar] [CrossRef]
- Hasunuma, T.; Miyazawa, S.-I.; Yoshimura, S.; Shinzaki1, Y.; Tomizawa, K.-I.; Shindo, K.; Choi, S.-K.; Misawa, N.; Miyake, C. Biosynthesis of astaxanthin in tobacco leaves by transplastomic engineering. Plant J. 2008, 55, 857–868. [Google Scholar] [CrossRef]
- Zhu, Q.; Zeng, D.; Yu, S.; Cui, C.; Li, J.; Li, H.; Chen, J.; Zhang, R.; Zhao, X.; Chen, L.; et al. From Golden Rice to aSTARice: Bioengineering Astaxanthin Biosynthesis in Rice Endosperm. Mol. Plant 2018, 11, 1440–1448. [Google Scholar] [CrossRef]
- Fujisawa, M.; Takita, E.; Harada, H.; Sakurai, N.; Suzuki, H.; Ohyama, K.; Shibata, D.; Misawa, N. Pathway engineering of Brassica napus seeds using multiple key enzyme genes involved in ketocarotenoid formation. J. Exp. Bot. 2009, 60, 1319–1332. [Google Scholar] [CrossRef]
- Jayaraj, J.; Devlin, R.; Punja, Z. Metabolic engineering of novel ketocarotenoid production in carrot plants. Transgenic Res. 2008, 17, 489–501. [Google Scholar] [CrossRef]
- Huang, J.; Zhong, Y.; Liu, J.; Sandmann, G.; Chen, F. Metabolic engineering of tomato for high-yield production of astaxanthin. Metab. Eng. 2013, 17, 59–67. [Google Scholar] [CrossRef]
- Nogueira, M.; Enfissi, E.M.A.A.; Martínez Valenzuela, M.E.; Menard, G.N.; Driller, R.L.; Eastmond, P.J.; Schuch, W.; Sandmann, G.; Fraser, P.D.; Martínez, M.E.; et al. Engineering of tomato for the sustainable production of ketocarotenoids and its evaluation in aquaculture feed. Proc. Natl. Acad. Sci. USA 2017, 114, 10876–10881. [Google Scholar] [CrossRef] [PubMed]
- Azadi, P.; Otang, N.V.; Chin, D.P.; Nakamura, I.; Fujisawa, M.; Harada, H.; Misawa, N.; Mii, M. Metabolic engineering of Lilium × formolongi using multiple genes of the carotenoid biosynthesis pathway. Plant Biotechnol. Rep. 2010, 4, 269–280. [Google Scholar] [CrossRef]
- Papp, T.; Csernetics, Á.; Nagy, G.; Bencsik, O.; Iturriaga, E.A.; Eslava, A.P.; Vágvölgyi, C. Canthaxanthin production with modified Mucor circinelloides strains. Appl. Microbiol. Biotechnol. 2013, 97, 4937–4950. [Google Scholar] [CrossRef] [PubMed]
- Pharkya, P.; Nikolaev, E.V.; Maranas, C.D. Review of the BRENDA Database. Metab. Eng. 2003, 5, 71–73. [Google Scholar] [CrossRef]
- Holliday, G.L.; Brown, S.D.; Mischel, D.; Polacco, B.J.; Babbitt, P.C. A strategy for large-scale comparison of evolutionary- and reaction-based classifications of enzyme function. Database 2020, 2020. [Google Scholar] [CrossRef]
- Fernández-González, B.; Sandmann, G.; Vioque, A. A new type of asymmetrically acting β-carotene ketolase is required for the synthesis of echinenone in the cyanobacterium Synechocystis sp. PCC 6803. J. Biol. Chem. 1997, 272, 9728–9733. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, L.; Chi, S.; Wang, G.; Wang, X.; Liu, T.; Tang, X. Phylogenetic analyses of the genes involved in carotenoid biosynthesis in algae. Acta Oceanol. Sin. 2018, 37, 89–101. [Google Scholar] [CrossRef]
- Fang, N.; Wang, C.; Liu, X.; Zhao, X.; Liu, Y.; Liu, X.; Du, Y.; Zhang, Z.; Zhang, H. De novo synthesis of astaxanthin: From organisms to genes. Trends Food Sci. Technol. 2019, 92, 162–171. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, T.; Takaichi, S.; Misawa, N.; Maoka, T.; Miyashita, H.; Mimuro, M. The cyanobacterium Gloeobacter violaceus PCC 7421 uses bacterial-type phytoene desaturase in carotenoid biosynthesis. FEBS Lett. 2005, 579, 2125–2129. [Google Scholar] [CrossRef] [PubMed]
- Steiger, S.; Jackisch, Y.; Sandmann, G. Carotenoid biosynthesis in Gloeobacter violaceus PCC4721 involves a single crtI-type phytoene desaturase instead of typical cyanobacterial enzymes. Arch. Microbiol. 2005, 184, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N. Carotenoid β-ring hydroxylase and ketolase from marine bacteria—Promiscuous enzymes for synthesizing functional xanthophylls. Mar. Drugs 2011, 9, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Steiger, S.; Sandmann, G. Cloning of two carotenoid ketolase genes from Nostoc punctiforme for the heterologous production of canthaxanthin and astaxanthin. Biotechnol. Lett. 2004, 26, 813–817. [Google Scholar] [CrossRef]
- Liang, C.; Zhao, F.; Wei, W.; Wen, Z.; Qin, S. Carotenoid biosynthesis in cyanobacteria: Structural and evolutionary scenarios based on comparative genomics. Int. J. Biol. Sci. 2006, 2, 197–207. [Google Scholar] [CrossRef]
- Enfissi, E.M.A.; Nogueira, M.; D’Ambrosio, C.; Stigliani, A.L.; Giorio, G.; Misawa, N.; Fraser, P.D. The road to astaxanthin production in tomato fruit reveals plastid and metabolic adaptation resulting in an unintended high lycopene genotype with delayed over-ripening properties. Plant Biotechnol. J. 2019, 1–13. [Google Scholar] [CrossRef]
- Harada, H.; Fujisawa, M.; Teramoto, M.; Sakurai, N.; Suzuki, H.; Shibata, D.; Misawa, N. Simple functional analysis of key genes involved in astaxanthin biosynthesis using Arabidopsis cultured cells. Plant Biotechnol. 2009, 26, 81–92. [Google Scholar] [CrossRef]
- Mortimer, C.L.; Misawa, N.; Perez-Fons, L.; Robertson, F.P.; Harada, H.; Bramley, P.M.; Fraser, P.D. The formation and sequestration of nonendogenous ketocarotenoids in transgenic Nicotiana glauca. Plant Physiol. 2017, 173, 1617–1635. [Google Scholar] [CrossRef]
- Wang, R.; Gu, X.; Yao, M.; Pan, C.; Liu, H.; Xiao, W.; Wang, Y.; Yuan, Y. Engineering of β-carotene hydroxylase and ketolase for astaxanthin overproduction in Saccharomyces cerevisiae. Front. Chem. Sci. Eng. 2017, 11, 89–99. [Google Scholar] [CrossRef]
- Moustafa, K.; Makhzoum, A.; Trémouillaux-Guiller, J. Molecular farming on rescue of pharma industry for next generations. Crit. Rev. Biotechnol. 2016, 36, 840–850. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.B.; Chandrasekar, B.; Mandal, M.K.; Kaschani, F.; Kaiser, M.; Both, L.; Hoorn, R.A.L.; Schiermeyer, A.; Abranches, R. Low protease content in Medicago truncatula cell cultures facilitates recombinant protein production. Biotechnol. J. 2018, 13, e1800050. [Google Scholar] [CrossRef] [PubMed]
- Farré, G.; Sanahuja, G.; Naqvi, S.; Bai, C.; Capell, T.; Zhu, C.; Christou, P. Travel advice on the road to carotenoids in plants. Plant Sci. 2010, 179, 28–48. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS). Scientific Opinion on the re-evaluation of canthaxanthin (E 161 g) as a food additive. EFSA J. 2010, 8, 1852. [Google Scholar] [CrossRef]
- Zhang, C. Biosynthesis of Carotenoids and Apocarotenoids by Microorganisms and their Industrial Potential. In Progress in Carotenoid Research; Zepka, L.Q., Jacob-Lopes, E., De Rosso, V.V., Eds.; IntechOpen: London, UK, 2018; Volume 5, pp. 85–105. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Major Ketocarotenoid | Other Carotenoids | Canthaxanthin Amount | Ref. | |
|---|---|---|---|---|---|
| Fungi | Aspergillus carbonarius | Canthaxanthin (partially saturated) | - | 20.6% DW | [37] |
| Cantharellus cinnabarinus | Canthaxanthin | β-carotene | n.a. | [36] | |
| Xanthophyllomyces dendrorhous | Astaxanthin | β-carotene, canthaxanthin, echinenone, zeaxanthin | n.a. | [38,39] | |
| Microalgae | Asterarcys quadricellulare | Astaxanthin | β-carotene, canthaxanthin, lutein | 1.4% DW | [40] |
| Botryococcus braunii | Canthaxanthin | β-carotene, echinenone, lutein, violaxanthin, zeaxanthin | 46% DW | [41] | |
| Chlorella emersonii | Canthaxanthin | β-carotene, neoxanthin, violaxanthin | n.a. | [42] | |
| Chlorella vulgaris | Canthaxanthin | Astaxanthin, violaxanthin | n.a. | [43] | |
| Chlorella zofingiensis | Canthaxanthin | β-carotene, astaxanthin, lutein | 25% TSC 2.1% of crude extract | [44,45] | |
| Chlorococcum | Adonixanthin | β-Carotene, astaxanthin, canthaxanthin, lutein | 3.6–18.9% TC | [46] | |
| Coelastrella striolata | Canthaxanthin | β-carotene, astaxanthin | 4.75% DW | [19] | |
| Dactylococcus dissociatus | Canthaxanthin | Lutein, neoxanthin, violaxanthin | 1.20–3.9 mg/L of culture | [47] | |
| Dictyococcus cinnabarinus | Echinenone | β-carotene, canthaxanthin | n.a. | [48] | |
| Haematococcus pluvialis | Astaxanthin | β-carotene, canthaxanthin, echinenone, lutein, neoxanthin, violaxanthin, zeaxanthin | 2.2–44.3% TC | [49,50,51] | |
| Nannochloropsis | Canthaxanthin | Antheraxanthin, astaxanthin, vaucheriaxanthin, violaxanthin, zeaxanthin | 0.8–16.9% TC | [52] | |
| Scenedesmus obliquus | Astaxanthin | β-carotene, adonirubin, adonixanthin, antheraxanthin, canthaxanthin, echinenone, lutein, neoxanthin, violaxanthin | n.a. | [53] | |
| Bacteria | Bradyrhizobium sp. | Canthaxanthin | Echinenone, spirilloxanthin | 86.6% TC | [54] |
| Brevibacterium sp. | Canthaxanthin | Echinenone | 7.2 mg/L | [55] | |
| Dietzia sp. | Canthaxanthin | β-carotene, astaxanthin | 90–92% TC | [56,57] | |
| Gordonia jacobaea | Canthaxanthin | β-carotene, astaxanthin | 88% TC | [18] | |
| Micrococcus roseus | Canthaxanthin | α/β-carotene derivatives | 85% TC | [58,59] | |
| Paracoccus schoinia | Adonixanthin diglucoside | α-carotene, α-cryptoxanthin, astaxanthin diglucoside, canthaxanthin, echinenone, zeaxanthin, zeaxanthin diglucoside | 1% mol TC | [60] | |
| Archaea | Haloferax alexandrinus TMT | Canthaxanthin | β-carotene | 33.8% TC | [61,62] |
| Species | Genes (Origin) | Major Carotenoid Accumulation | Ref. |
|---|---|---|---|
| Escherichia coli | crtW (A. variabilis) | β-carotene, β-cryptoxanthin, canthaxanthin | [77] |
| crtW (P. N81106) crtW (B. sp. SD212) | Adonixanthin Canthaxanthin | [78] | |
| Nostoc sp. PCC 7120 | crtO (N. flagelliforme) | Echinenone, canthaxanthin | [79] |
| Nicotiana benthamiana | crtZ and crtW (B. sp. SD212) | Canthaxanthin, astaxanthin | [80] |
| Nicotiana tabacum | crtZ and crtW (B. sp. SD212) | Adonixanthin, canthaxanthin, astaxanthin, 4-ketoantheraxanthin | [81] |
| Oryza sativa | psy1 (maize) crtI (P. ananatis) bkt (H. pluvialis or C. reinhardtii) | Astaxanthin, β-carotene, canthaxanthin | [82] |
| Glycine max | crtB (P. ananatis) crtW (B. sp. SD212) bkt (H. pluvialis) | β-carotene, canthaxanthin, astaxanthin | [71] |
| Brassica napus | crtE (P. ananatis) crtW (B. sp. SD212) | α-carotene, echinenone, lutein, canthaxanthin | [83] |
| Daucus carota L. | bkt (H. pluvialis) | Astaxanthin, adonirubin and canthaxanthin | [84] |
| Solanum lycopersicum | bhy (H. pluvialis) bkt (C. reinhardtii) | β-carotene, astaxanthin, adonirubin, canthaxanthin, echinenone | [85] |
| crtZ, crtW (B. sp. SD212) | β-carotene, astaxanthin, phoenicoxanthin, adonixanthin, canthaxanthin, echinenone | [86] | |
| Lilium x formolongi | crtE, crtB, crtI, crtY (P. ananatis) crtZ, crtW (B. sp. SD212) idi (P. sp. N81106) | Echinenone, canthaxanthin, 3-hydroxyechinenone, astaxanthin | [87] |
| Mucor circinelloides | crtW (P. sp. N81106) | β-carotene, echinenone, canthaxanthin | [88] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebelo, B.A.; Farrona, S.; Ventura, M.R.; Abranches, R. Canthaxanthin, a Red-Hot Carotenoid: Applications, Synthesis, and Biosynthetic Evolution. Plants 2020, 9, 1039. https://doi.org/10.3390/plants9081039
Rebelo BA, Farrona S, Ventura MR, Abranches R. Canthaxanthin, a Red-Hot Carotenoid: Applications, Synthesis, and Biosynthetic Evolution. Plants. 2020; 9(8):1039. https://doi.org/10.3390/plants9081039
Chicago/Turabian StyleRebelo, Bárbara A., Sara Farrona, M. Rita Ventura, and Rita Abranches. 2020. "Canthaxanthin, a Red-Hot Carotenoid: Applications, Synthesis, and Biosynthetic Evolution" Plants 9, no. 8: 1039. https://doi.org/10.3390/plants9081039
APA StyleRebelo, B. A., Farrona, S., Ventura, M. R., & Abranches, R. (2020). Canthaxanthin, a Red-Hot Carotenoid: Applications, Synthesis, and Biosynthetic Evolution. Plants, 9(8), 1039. https://doi.org/10.3390/plants9081039