The Role of Microbial Inoculants on Plant Protection, Growth Stimulation, and Crop Productivity of the Olive Tree (Olea europea L.)


 ,
, 

 and
and 
Abstract
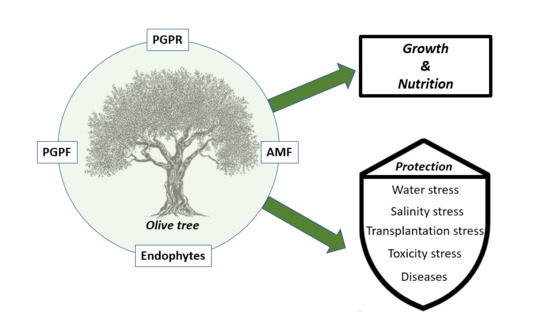
1. Introduction
2. Categorization of Studies
3. Beneficial Microorganisms and Olive Tree’s Growth, Physiology, and Productivity
4. Beneficial Microbes and a Plant’s Response to Abiotic and Transplantation Stress
5. Beneficial Microorganisms Against Pathogens
6. Inoculum-Cultivar Specificity
7. Microbial Inoculation under Nursery and Field Conditions
8. Future Perspectives
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 2011, 344, 1–50. [Google Scholar] [CrossRef]
- Schilirò, E.; Ferrara, M.; Nigro, F.; Mercado-Blanco, J. Genetic Responses Induced in Olive Roots upon Colonization by the Biocontrol Endophytic Bacterium Pseudomonas fluorescens PICF7. PLoS ONE 2012, 7, e48646. [Google Scholar] [CrossRef] [PubMed]
- Cabanás, C.G.L.; Schilirò, E.; Valverde-Corredor, A.; Mercado-Blanco, J. The biocontrol endophytic bacterium Pseudomonas fluorescens PICF7 induces systemic defense responses in aerial tissues upon colonization of olive roots. Front. Microbiol. 2014, 5, 427. [Google Scholar]
- Ouledali, S.; Ennajeh, M.; Zrig, A.; Gianinazzi, S.; Khemira, H. Estimating the contribution of arbuscular mycorrhizal fungi to drought tolerance of potted olive trees (Olea europaea). Acta Physiol. Plant. 2018, 40, 81. [Google Scholar] [CrossRef]
- Ben Amira, M.; Lopez, D.; Triki Mohamed, A.; Khouaja, A.; Chaar, H.; Fumanal, B.; Gousset-Dupont, A.; Bonhomme, L.; Label, P.; Goupil, P.; et al. Beneficial effect of Trichoderma harzianum strain Ths97 in biocontrolling Fusarium solani causal agent of root rot disease in olive trees. Biol. Control. 2017, 110, 70–78. [Google Scholar] [CrossRef]
- Seifi, E.; Teymoor, Y.S.; Alizadeh, M.; Fereydooni, H. Olive mycorrhization: Influences of genotype, mycorrhiza, and growing periods. Sci. Hortic. 2014, 180, 214–219. [Google Scholar] [CrossRef]
- Kara, Z.; Arslan, D.; Güler, M.; Güler, Ş. Inoculation of arbuscular mycorrhizal fungi and application of micronized calcite to olive plant: Effects on some biochemical constituents of olive fruit and oil. Sci. Hortic. 2015, 185, 219–227. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Sánchez-Castro, I.; Cantos, M.; García, J.L.; Azcón, R.; Ruiz-Lozano, J.M.; Beuzón, C.R.; Aroca, R. Effects of different arbuscular mycorrhizal fungal backgrounds and soils on olive plants growth and water relation properties under well-watered and drought conditions. Plant Cell Environ. 2016, 39, 2498–2514. [Google Scholar] [CrossRef]
- Fouad, M.O.; Essahibi, A.; Benhiba, L.; Qaddoury, A. Effectiveness of arbuscular mycorrhizal fungi in the protection of olive plants against oxidative stress induced by drought. Span. J. Agric. Res. 2014, 12, 763–771. [Google Scholar] [CrossRef]
- Merwad, M.A.; Shahin, M.F.M.; Haggag, L.F. Optimizing growth of “Picual” olive seedlings by using organic and biofertilizers as soil application under greenhouse condition. Int. J. Chemtech Res. 2015, 8, 36–42. [Google Scholar]
- Kour, D.; Bakshi, P.; Wali, V.K.; Sharma, N.; Sharma, A.; Iqbal, M. Alternate Bearing in Olive—A Review. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2281–2297. [Google Scholar] [CrossRef]
- Sidhoum, W.; Fortas, Z. Effect of Arbuscular mycorrhizal fungi on growth of semi-woody olive cuttings of the variety “Sigoise” in Algeria. Am. J. Res. Commun. 2013, 1, 244–257. [Google Scholar]
- Bompadre, M.J.; Pérgola, M.; Fernández Bidondo, L.; Colombo, R.P.; Silvani, V.A.; Pardo, A.G.; Ocampo, J.A.; Godeas, A.M. Evaluation of arbuscular mycorrhizal fungi capacity to alleviate abiotic stress of olive (Olea europaea L.) plants at different transplant conditions. Sci. World J. 2014, 6, 378950. [Google Scholar]
- Meddad-Hamza, A.; Beddiar, A.; Gollotte, A.; Lemoine, M.C.; Kuszala, C.; Gianinazzi, S. Arbuscular mycorrhizal fungi improve the growth of olive trees and their resistance to transplantation stress. Afr. J. Biotechnol. 2010, 9, 1159–1167. [Google Scholar]
- Cabanás, C.G.L.; Ruano-Rosa, D.; Legarda, G.; Pizarro-Tobías, P.; Valverde-Corredor, A.; Triviño, J.C.; Roca, A.; Mercado-Blanco, J. Bacillales members from the olive rhizosphere are effective biological control agents against the defoliating pathotype of verticillium dahliae. Agriculture 2018, 8, 90. [Google Scholar] [CrossRef]
- Ghanney, N.; Locantore, P. Potential Biocontrol Effect of the Phylloplane Bacterium Bacillus mojavensis ABC-7 on the Olive Knot Disease. J. Plant Pathol. Microbiol. 2016, 7, 337. [Google Scholar] [CrossRef]
- Nigro, F.; Antelmi, I.; Labarile, R.; Sion, V.; Pentimone, I. Biological control of olive anthracnose. Acta Hortic. 2018, 1199, 439–444. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Valverde-Corredor, A.; Gómez-Lama Cabanás, C.; Sesmero, R.; Mercado-Blanco, J. What Lies Beneath: Root-Associated Bacteria to Improve the Growth and Health of Olive Trees. In Soil Biological Communities and Ecosystem Resilience; Lukac, M., Grenni, P., Gamboni, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 107–122. [Google Scholar]
- Maksoud, M.A.; El-Shamma, M.S.; Saleh, M.A.; Zaied, N.S.; Hafez, O.M. Effect of different compost sorts and biofertilizers on chemlali olive trees grown in calcareous soil. Middle East. J. Sci. Res. 2012, 12, 1046–1049. [Google Scholar]
- Garnault, M.; Duplaix, C.; Leroux, P.; Couleaud, G.; Carpentier, F.; David, O.; Walker, A.S. Spatiotemporal dynamics of fungicide resistance in the wheat pathogen Zymoseptoria tritici in France. Pest. Manag. Sci. 2019, 75, 1794–1807. [Google Scholar] [CrossRef]
- Shaheen, S.A.; El Taweel, A.A.; Omar, M.N.A. Effect of inoculation by some plant growth promoting rhizobacteria (PGPR) on production of “Manzanillo” olive trees. Acta Hortic. 2014, 1018, 245–254. [Google Scholar] [CrossRef]
- Abd-Alhamid, N.; Hassan, H.S.A.; Haggag, L.F.; Hassan, A.M. Effect of mineral and bio-fertilization on vegetative growth, leaf mineral contents and flowering of manzanillo olive trees. Int. J. Chemtech. Res. 2015, 8, 51–61. [Google Scholar]
- Cabanás, C.G.L.; Legarda, G.; Ruano-Rosa, D.; Pizarro-Tobías, P.; Valverde-Corredor, A.; Niqui, J.L.; Triviño, J.C.; Roca, A.; Mercado-Blanco, J. Indigenous Pseudomonas spp. Strains from the Olive (Olea europaea L.) rhizosphere as effective biocontrol agents against Verticillium dahliae: From the host roots to the bacterial genomes. Front. Microbiol. 2018, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Del Mar Alguacil, M.; Torrecillas, E.; Kohler, J.; Roldàn, A. A molecular approach to ascertain the success of “in situ” AM fungi inoculation in the revegetation of a semiarid, degraded land. Sci. Total Environ. 2011, 409, 2874–2880. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.M.D.R.; Melloni, R.; da Silva, L.F.D.O.; Martins, F.B.; Gonçalves, E.D. Arbuscular mycorrhizal fungi in seedling development of olive (Olea europaea L.) in the south of Minas Gerais, Brazil. Rev. Bras. Cienc. Do Solo 2015, 39, 361–366. [Google Scholar] [CrossRef]
- Chenchouni, H.; Mekahlia, M.N.; Beddiar, A. Effect of inoculation with native and commercial arbuscular mycorrhizal fungi on growth and mycorrhizal colonization of olive (Olea europaea L.). Sci. Hortic. 2020, 261, 108969. [Google Scholar] [CrossRef]
- Mechri, B.; Attia, F.; Tekaya, M.; Cheheb, H.; Hammami, M. Colonization of olive trees (Olea europaea L.) with the arbuscular mycorrhizal fungus Glomus sp. modified the glycolipids biosynthesis and resulted in accumulation of unsaturated fatty acids. J. Plant Physiol. 2014, 171, 1217–1220. [Google Scholar] [CrossRef]
- Tekaya, M.; Mechri, B.; Mbarki, N.; Cheheb, H.; Hammami, M.; Attia, F. Arbuscular mycorrhizal fungus Rhizophagus irregularis influences key physiological parameters of olive trees (Olea europaea L.) and mineral nutrient profile. Photosynthetica 2017, 55, 308–316. [Google Scholar] [CrossRef]
- Mechri, B.; Tekaya, M.; Cheheb, H.; Attia, F.; Hammami, M. Accumulation of flavonoids and phenolic compounds in olive tree roots in response to mycorrhizal colonization: A possible mechanism for regulation of defense molecules. J. Plant Physiol. 2015, 185, 40–43. [Google Scholar] [CrossRef]
- Dag, A.; Yermiyahu, U.; Ben-Gal, A.; Zipori, I.; Kapulnik, Y. Nursery and post-transplant field response of olive trees to arbuscular mycorrhizal fungi in an arid region. Crop. Pasture Sci. Crop. Pasture Sci. 2009, 60, 427–433. [Google Scholar] [CrossRef]
- Chatzistathis, T.; Orfanoudakis, M.; Alifragis, D.; Therios, I. Colonization of Greek olive cultivars’ root system by arbuscular mycorrhiza fungus: Root morphology, growth, and mineral nutrition of olive plants. Sci. Agric. 2013, 70, 185–194. [Google Scholar] [CrossRef]
- Boutaj, H.; Meddich, A.; Wahbi, S.; Moukhli, A.; El Alaoui-Talibi, Z.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Improvement of growth and development of olive tree by mycorrhizal autochthonous inoculum. Res. J. Biotechnol. 2020, 15, 76–84. [Google Scholar]
- Semane, F.; Chliyeh, M.; Talbi, Z.; Touati, J.; Selmaoui, K.; Touhami, A.O.; Filali-Maltouf, A.; El Modafar, C.; Moukhli, A.; Benkirane, R.; et al. Effects of a Composite Endomycorrhizal Inoculum on Olive Cuttings under the Greenhouse Conditions. Int. J. Environ. Agric. Biotechnol. 2017, 2, 1070–1083. [Google Scholar] [CrossRef]
- Haggag, L.F.; Merwad, M.A. Effect of NPK and bio-fertilers as soil application on promoting growth of “Toffahi” olive seedlings under greenhouse condition. J. Agric. Tech. 2014, 10, 1607–1617. [Google Scholar]
- Peyvandi, M.; Farahani, F.; Hosseini Mazinani, M.; Noormohamadi, Z.; Ataii, S.; Asgharzade, A. Pseudomonas fluorescent and its ability to promote root formation of olive microshoots. Int. J. Plant Prod. 2010, 4, 63–66. [Google Scholar]
- Montero-Calasanz, M.C.; Santamaría, C.; Albareda, M.; Daza, A.; Duan, J.; Glick, B.R.; Camacho, M. Alternative rooting induction of semi-hardwood olive cuttings by several auxin-producing bacteria for organic agriculture systems. Span. J. Agric. Res. 2013, 11, 146–154. [Google Scholar] [CrossRef]
- Rosa, D.D.; Villa, F.; Da Silva, D.F.; Corbari, F. Rooting of semihardwood cuttings of olive: Indolbutyric acid, calcium and Azospirillum brasilense. Comun. Sci. 2018, 9, 34–40. [Google Scholar] [CrossRef][Green Version]
- Ramasamy, K.; Joe, M.M.; Kim, K.-Y.; Lee, S.-M.; Shagol, C.; Rangasamy, A.; Chung, J.-B.; Islam, M.R.; Sa, T.-M. Synergistic Effects of Arbuscular Mycorrhizal Fungi and Plant Growth Promoting Rhizobacteria for Sustainable Agricultural Production. Korean J. Soil Sci. Fertil. 2011, 44, 637–649. [Google Scholar] [CrossRef]
- Lies, A.; Ferhout, H.; Wahbi, S.; Hafidi, M.; Prin, Y.; Duponnois, R. Selection of efficient microbial biofertilizers from Moroccan soils. Acta Hortic. 2016, 1148, 115–126. [Google Scholar] [CrossRef]
- Costa, S.M.L.; Melloni, R. Relationship of arbuscular mycorrhizal fungi and rhizobacteria on the growth of olive tree seedlings (Olea europaea). Cienc. Florest. 2019, 29, 169–181. [Google Scholar] [CrossRef]
- Hafez, O.M.; Saleh, M.A.; El-Lethy, S.R. Response of some seedlings olive cultivars to foliar spray of yeast and garlic extracts with or without vascular arbuscular mycorrhizal fungi. World Appl. Sci. J. 2013, 24, 1119–1129. [Google Scholar]
- Porras-Soriano, A.; Soriano-Martín, M.L.; Porras-Piedra, A.; Azcón, R. Arbuscular mycorrhizal fungi increased growth, nutrient uptake and tolerance to salinity in olive trees under nursery conditions. J. Plant Physiol. 2009, 166, 1350–1359. [Google Scholar] [CrossRef] [PubMed]
- Chliyeh, M.; Ouazzani Touhami, A.; Filali-Maltouf, A.; El Modafar, C.; Moukhli, A.; Oukabli, A.; Benkirane, R.; Douira, A. Effect of a composite endomycorrhizal inoculum on the growth of olive trees under nurseries conditions in Morocco. Int. J. Pure Appl. Biosci. 2014, 2, 1–14. [Google Scholar]
- Kapulnik, Y.; Tsror, L.; Zipori, I.; Hazanovsky, M.; Wininger, S.; Dag, A. Effect of AMF application on growth, productivity and susceptibility to Verticillium wilt of olives grown under desert conditions. Symbiosis 2010, 52, 103–111. [Google Scholar] [CrossRef]
- Bompadre, M.J.; Rios De Molina, M.C.; Colombo, R.P.; Fernandez Bidondo, L.; Silvani, V.A.; Pardo, A.G.; Ocampo, J.A.; Godeas, A.M. Differential efficiency of two strains of the arbuscular mycorrhizal fungus Rhizophagus irregularis on olive (Olea europaea) plants under two water regimes. Symbiosis 2013, 61, 105–112. [Google Scholar] [CrossRef]
- M’barki, N.; Chehab, H.; Aissaoui, F.; Dabbaghi, O.; Attia, F.; Mahjoub, Z.; Laamari, S.; Chihaoui, B.; del Giudice, T.; Jemai, A.; et al. Effects of mycorrhizal fungi inoculation and soil amendment with hydrogel on leaf anatomy, growth and physiology performance of olive plantlets under two contrasting water regimes. Acta Physiol. Plant. 2018, 40, 116. [Google Scholar] [CrossRef]
- Jiménez-Moreno, M.J.; Moreno-Márquez, M.d.C.; Moreno-Alías, I.; Rapoport, H.; Fernández-Escobar, R. Interaction between mycorrhization with Glomus intraradices and phosphorus in nursery olive plants. Sci. Hortic. 2018, 233, 249–255. [Google Scholar] [CrossRef]
- Khabou, W.; Hajji, B.; Zouari, M.; Rigane, H.; Abdallah, F. Ben Arbuscular mycorrhizal fungi improve growth and mineral uptake of olive tree under gypsum substrate. Ecol. Eng. 2014, 73, 290–296. [Google Scholar] [CrossRef]
- Briccoli Bati, C.; Santilli, E.; Lombardo, L. Effect of arbuscular mycorrhizal fungi on growth and on micronutrient and macronutrient uptake and allocation in olive plantlets growing under high total Mn levels. Mycorrhiza 2014, 25, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Ouledali, S.; Ennajeh, M.; Ferrandino, A.; Khemira, H.; Schubert, A.; Secchi, F. Influence of arbuscular mycorrhizal fungi inoculation on the control of stomata functioning by abscisic acid (ABA) in drought-stressed olive plants. South. Afr. J. Bot. 2019, 121, 152–158. [Google Scholar] [CrossRef]
- Silva, T.F.; Melloni, R.; Melloni, E.G.P.; Gonçalves, E.D. Non-symbiotic diazotrophic bacteria and the rooting of olive semi-hardwood cuttings (Olea europaea L.). Cienc. Florest. 2017, 27, 61–71. [Google Scholar] [CrossRef]
- Maksoud, M.A.; Saleh, M.A.; Fouad, A.A. The Beneficial Effect of Biofertilizers and Antioxidants on Olive Trees under Calcareous Soil Conditions. World J. Agric. Sci. 2009, 5, 350–352. [Google Scholar]
- El-Shazly, M.M.; Ghieth, W.M. Effect of Some Biofertilizers and Humic Acid Application on Olive Seedlings Growth under Irrigation with Saline Water. Alex. Sci. Exch. J. 2019, 40, 263–279. [Google Scholar] [CrossRef]
- Bouaichi, A.; Benkirane, R.; El-Kinany, S.; Habbadi, K.; Lougraimzi, H.; Sadik, S.; Benbouazza, A.; Achbani, E.H. Potential effect of antagonistic bacteria in the management of olive knot disease caused by Pseudomonas savastanoi pv. savastanoi. J. Microbiol. Biotechnol. Food Sci. 2019, 8, 1035–1040. [Google Scholar] [CrossRef]
- Sanei, S. Suppression of Verticillium Wilt of Olive by Pseudomonas fluorescens. Am. J. Exp. Agric. 2011, 1, 294–305. [Google Scholar] [CrossRef]
- Markakis, E.A.; Tjamos, S.E.; Antoniou, P.P.; Paplomatas, E.J.; Tjamos, E.C. Biological control of Verticillium wilt of olive by Paenibacillus alvei, strain K165. BioControl 2016, 61, 293–303. [Google Scholar] [CrossRef]
- Hibar, K.; Gamaoun, W.; Triki, M.A. Isolation, identification and biological control of the major pathogens causing root rot and wilt diseases of young olive trees in. J. New Sci. 2017, 39, 2121–2130. [Google Scholar]
- Abdel-Monaim, M.F.; El-Morsi, M.E.A.; Hassan, M.A.E. Control of root rot and wilt disease complex of some evergreen fruit transplants by using plant growth promoting rhizobacteria in the New Valley Governorate, Egypt. J. Phytopathol. 2014, 1, 23–33. [Google Scholar]
- Bouzoumita, A.; Metoui, M.; Jemni, M.; Kabaeir, N.; Belhouchette, K.; Ferchichi, A. The efficacy of various bacterial organisms for biocontrol of fusarium root rot of olive in tunisia. Pol. J. Environ. Stud. 2020, 29, 11–16. [Google Scholar] [CrossRef]
- Prieto, P.; Schilirò, E.; Maldonado-González, M.M.; Valderrama, R.; Barroso-Albarracín, J.B.; Mercado-Blanco, J. Root Hairs Play a Key Role in the Endophytic Colonization of Olive Roots by Pseudomonas spp. with Biocontrol Activity. Microb. Ecol. 2011, 32, 435–445. [Google Scholar] [CrossRef]
- Maldonado-González, M.M.; Prieto, P.; Ramos, C.; Mercado-Blanco, J. From the root to the stem: Interaction between the biocontrol root endophyte Pseudomonas fluorescens PICF7 and the pathogen Pseudomonas savastanoi NCPPB 3335 in olive knots. Microb. Biotechnol. 2013, 6, 275–287. [Google Scholar] [CrossRef]
- Prieto, P.; Navarro-Raya, C.; Valverde-Corredor, A.; Amyotte, S.G.; Dobinson, K.F.; Mercado-Blanco, J. Colonization process of olive tissues by Verticillium dahliae and its in planta interaction with the biocontrol root endophyte Pseudomonas fluorescens PICF7. Microb. Biotechnol. 2009, 2, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Cabanás, C.G.L.; Sesmero, R.; Valverde-Corredor, A.; Javier López-Escudero, F.; Mercado-Blanco, J. A split-root system to assess biocontrol effectiveness and defense-related genetic responses in above-ground tissues during the tripartite interaction Verticillium dahliae-olive-Pseudomonas fluorescens PICF7 in roots. Plant Soil 2017, 417, 433–452. [Google Scholar] [CrossRef]
- Slama, H.B.; Triki, M.A.; Bouket, A.C.; Mefteh, F.B.; Alenezi, F.N.; Luptakova, L.; Cherif-Silini, H.; Vallat, A.; Oszako, T.; Gharsallah, N.; et al. Screening of the high-rhizosphere competent limoniastrum monopetalum’ culturable endophyte microbiota allows the recovery of multifaceted and versatile biocontrol agents. Microorganisms 2019, 7, 249. [Google Scholar] [CrossRef] [PubMed]
- Cheffi, M.; Bouket, A.C.; Alenezi, F.N.; Luptakova, L.; Belka, M.; Vallat, A.; Rateb, M.E.; Tounsi, S.; Triki, M.A.; Belbahri, L. Olea europaea L. Root endophyte Bacillus Velezensis oee1 counteracts oomycete and fungal harmful pathogens and harbours a large repertoire of secreted and volatile metabolites and beneficial functional genes. Microorganisms 2019, 7, 314. [Google Scholar] [CrossRef]
- Carrero-Carrón, I.; Trapero-Casas, J.L.; Olivares-García, C.; Monte, E.; Hermosa, R.; Jiménez-Díaz, R.M. Trichoderma asperellum is effective for biocontrol of Verticillium wilt in olive caused by the defoliating pathotype of Verticillium dahliae. Crop. Prot. 2016, 88, 45–52. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Prieto, P.; Rincón, A.M.; Gómez-Rodríguez, M.V.; Valderrama, R.; Barroso, J.B.; Mercado-Blanco, J. Fate of Trichoderma harzianum in the olive rhizosphere: Time course of the root colonization process and interaction with the fungal pathogen Verticillium dahliae. BioControl 2016, 61, 269–282. [Google Scholar] [CrossRef]
- Varo, A.; Moral, J.; Lozano-Tóvar, M.D.; Trapero, A. Development and validation of an inoculation method to assess the efficacy of biological treatments against Verticillium wilt in olive trees. BioControl 2016, 61, 283–292. [Google Scholar] [CrossRef]
- Otero, M.L.; Roca, M.; Zapata, R.; Ladux, J.L.; Ortiz, J.; Zanelli, M.; Matías, A.C.; Pérez, B.A. Effect of solarization, organic matter, and Trichoderma on the severity of Verticillium wilt in olive trees (Olea europaea L.) and soil inoculum density. Acta Hortic. 2014, 1057, 121–126. [Google Scholar] [CrossRef]
- Boutaj, H.; Meddich, A.; Wahbi, S.; Moukhli, A.; El Alaoui-Talibi, Z.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Effect of arbuscular mycorrhizal fungi on verticillium wilt development of olive trees caused by Verticillium dahliae. Res. J. Biotechnol. 2019, 14, 79–88. [Google Scholar]
- Varo-Suárez, A.; Raya-Ortega, M.C.; Agustí-Brisach, C.; García-Ortiz-Civantos, C.; Fernández-Hernández, A.; Mulero-Aparicio, A.; Trapero, A. Evaluation of organic amendments from agro-industry waste for the control of verticillium wilt of olive. Plant Pathol. 2018, 67, 860–870. [Google Scholar] [CrossRef]
- Arici, S.E.; Demirtas, A.E. The effectiveness of rhizosphere microorganisms to control Verticillium wilt disease caused by Verticillium dahliae Kleb. in olives. Arab. J. Geosci. 2019, 12, 781. [Google Scholar] [CrossRef]
- Mulero-Aparicio, A.; Varo, A.; Agustí-Brisach, C.; López-Escudero, F.J.; Trapero, A. Biological control of Verticillium wilt of olive in the field. Crop. Prot. 2020, 128, 104993. [Google Scholar] [CrossRef]

{kind=link}
| Microbial Inoculant | Type | Product | Effect | Application | Age | Location | References |
|---|---|---|---|---|---|---|---|
| Rhizophagus irregularis, Glomus mosseae | AMF | - | Growth | Root | Young | Nursery | [6] |
| R. irregularis | AMF | - | Growth and Nutrition * | Root | Young | Nursery | [7] |
| Glomus sp., Acaulospora sp., Glomus macrocarpum, Glomus multicaulis, Scutellospora sp. | AMF | - | Growth | Root | Young | Nursery | [12] |
| Glomus sp., R. irregularis | AMF | - | Growth | Root | Young | Experimental Field | [24] |
| Glomus clarum, Gigaspora rosea, Scutellospora scrobiculata | AMF | - | Growth | Root | Young | Nursery | [25] |
| Glomus sp., Septoglomus constrictum, R. irregularis, Funneliformis mosseae, Gigaspora margarita | AMF | - | Growth | Root | Young | Nursery | [26] |
| R. irregularis | AMF | - | Growth and Nutrition | Root | Young | Nursery | [27] |
| R. irregularis | AMF | - | Growth and Nutrition ** | Root | Young | Nursery | [28] |
| R. irregularis | AMF | - | Growth and Nutrition ** | Root | Young | Nursery | [29] |
| R. irregularis, G. mosseae | AMF | - | Growth and Nutrition | Root | Young | Nursery | [30] |
| Glomus sp., Gigaspora sp. | AMF | - | Growth and Nutrition | Root | Young | Nursery | [31] |
| Glomus sp., Acaulospora sp., Gigaspora sp., Entrophospora sp., Scutellospora sp., Glomus irregulare DAOM 197198 | AMF | - | Growth and Nutrition | Root | Young | Nursery | [32] |
| Acaulospora scrobiculata, G. macrocarpum, R. irregularis, Glomus versiforme, Gigaspora sp., Scutellospora fulgida, Glomus geosporum, Glomus aureum, Glomus microcarpum, Glomus aurantium, G. corymbiforme, G. clarum, Scutellospora heterogama | AMF | - | Growth | Root | Young | Nursery | [33] |
| Azotobacter chrococcum, Bacillus megaterium, Bacillus circulans | PGPR | - | Growth | Root | Adult | Orchard | [22] |
| Azospirillum sp., Bacillus mucilaginosus, B. megatherium | PGPR | - | Growth and Nutrition | Root | Young | Nursery | [34] |
| Pseudomonas fluorescent P19, P21 | PGPR | - | Growth | Root | Young | Nursery | [35] |
| Pantoea sp. AG9, Chryseobacterium sp. AG13, Pseudomonas sp. CT364, Chryseobacterium sp. CT348, Azospirillum brasilense Cd | PGPR | - | Growth | Root | Young | Nursery | [36] |
| Azospirillum brasilense | PGPR | Azototal® | Growth | Root | Young | Nursery | [37] |
| Azotobacter chroococcum, Azospirillum brasilense, B. megaterium var. phosphaticum, G. mosseae NRC31, Pseudomonas sp., Bacillus cereus, Glomus fasciculatum NRC15 | AMF and PGPR | - | Growth and Nutrition | Root | Young | Nursery | [10] |
| Pseudomonas sp., Paenibacillus sp., Acaulospora scrobiculata, Gigaspora rosea, Rhizophagus clarus | AMF and PGPR | - | Growth and Nutrition | Root | Young | Nursery | [40] |
| Bacillus polymyxa, Saccharomyces cereivisea | PGPR and Fungi | Biovine® | Growth and Nutrition | Foliar | Adult | Orchard | [21] |
| AMF consortium, S. cerevisiae | AMF and Fungi | - | Growth and Nutrition | Root/Foliar | Young | Nursery | [41] |
| Microbial Inoculant | Type | Product | Stress Conditions | Application | Age | Location | References |
|---|---|---|---|---|---|---|---|
| G. etunicatum, G. microaggregatum, R. irregularis, G. claroideum, G. mosseae, G. geosporum | AMF | Symbivit® | Water Stress | Root | Young | Experimental Field | [4] |
| Glomus sp., R. irregularis, Dominikia sp., Funneliformis sp., Funneliformis geosporum, Septoglomus constrictum, Septoglomus viscosum | AMF | - | Water Stress | Root | Young | Nursery | [8] |
| Rhizophagus manihotis, Funneliformis mosseae | AMF | - | Water Stress | Root | Young | Nursery | [9] |
| R. irregularis GC2, GA5 | AMF | - | Transplantation | Root | Young | Nursery | [13] |
| G. mosseae, R. irregularis | AMF | - | Transplantation | Root | Young | Nursery | [14] |
| G. mosseae, R. irregularis, G. claroideum | AMF | - | Salinity | Root | Young | Nursery | [42] |
| R. irregularis, G. trimurales, G. manihoti, G. diaphanum, G. pansihalos, G. geosporum, G. glomerulatum, G. fasciculatum, G. etunicatum, Acaulospora sp., Acaulospora mellea, Entrophospora kentinensis, Scutellospora nigra, Scutellospora fulgida, Scutellospora heterogama, | AMF | - | Transplantation | Root | Young | Nursery | [43] |
| R. irregularis, G. mosseae | AMF | - | Salinity | Root | Young | Experimental Field | [44] |
| R. irregularis GC2, GA5 | AMF | - | Water Stress | Root | Young | Nursery | [45] |
| R. irregularis | AMF | - | Water Stress | Root | Young | Nursery | [46] |
| R. irregularis | AMF | - | Nutrient stress | Root | Young | Nursery | [47] |
| R. irregularis | AMF | - | Toxicity | Root | Young | Nursery | [48] |
| Glomus sp., Scutellospora heterogama, Paraglomus laccatum, Diversispora celata | AMF | Endorize IV® | Toxicity | Root | Young | Nursery | [49] |
| R. irregularis, G. etunicatum, G. microaggregatum, G. claroideum, G. mosseae, G. geosporum | AMF | Symbivit® | Water Stress | Root | Young | Nursery | [50] |
| Azotobacter chrococcum | PGPR | - | Toxicity | Root | Young | Nursery | [19] |
| Azospirillum brasilense BR11001t, Azospirillum amazonense BR11040t, Herbaspirillum seropedicae BR11175t, Burkholderia brasiliensis BR11340t | PGPR | - | Temperature and pH | Root | Young | Nursery | [51] |
| B. megatherium | PGPR | Phosphorine® | Toxicity | Root | Young | Nursery | [52] |
| Azotobacter chroococcum, G. macrocarbium | PGPR and AMF | - | Salinity | Root | Young | Experimental Field | [53] |
| Microbial Inoculant | Type | Product | Disease | Application | Age | Location | References |
|---|---|---|---|---|---|---|---|
| Pseudomonas fluorescens PICF7 | Endophyte | - | Verticillium wilt | Root | Young | Nursery | [2] |
| P. fluorescens PICF7 | Endophyte | - | Verticillium wilt | Root | Young | Nursery | [3] |
| Trichoderma harzianum Ths97 | PGPF | - | Root rot | Root | Young | Nursery | [5] |
| Paenibacillus polymyxa, Paenibacillus terrae, Bacillus sp. PIC28, P. fluorescensPICF7, Pseudomonas sp. PICF141 | PGPR | - | Verticillium wilt | Root | Young | Nursery | [15] |
| Bacillus mojavensis A-BC-7 | PGPR | - | Olive knot | Stem | Young | Nursery | [16] |
| B. subtilis QST 713, Aureobasidium pullulans A3, A5, A6 | PGPR Endophyte | Serenade Max® | Anthracnose | Foliar | Adult | Orchard | [17] |
| Pseudomonas spp. PIC25, PIC105, PICF141 | PGPR | - | Verticillium wilt | Root | Young | Nursery | [23] |
| R. irregularis, G. mosseae | AMF | - | Verticillium wilt | Root | Young | Experimental Field | [44] |
| B. subtilis 2515-1 | PGPR | - | Olive knot | Stem | Young | Nursery | [54] |
| P. fluorescens | PGPR | - | Verticillium wilt | Root | Young | Nursery | [55] |
| Paenibacillus alvei K165 | PGPR | - | Verticillium wilt | Root | Young | Experimental Field | [56] |
| B. subtilis Y1336 | PGPR | Biobac® | Root rot and wilt | Root | Young | Nursery | [57] |
| Azotobacter sp. AZM1, B. cereus BCM8, B. megatherium BMM5, B. subtilis BSM1 | PGPR | - | Root rot | Root | Young | Nursery | [58] |
| B. licheniformis, Enterobacter colcae | PGPR | - | Root rot | Root | Young | Nursery | [59] |
| P. fluorescens PICF7, P. putida PICP2 | Endophyte | - | Verticillium wilt | Root | Young | Nursery | [60] |
| P. fluorescens PICF7 | Endophyte | - | Olive knot | Root | Young | Nursery | [61] |
| P. fluorescens PICF7 | Endophyte | - | Verticillium wilt | Root | Young | Nursery | [62] |
| P. fluorescens PICF7 | Endophyte | - | Verticillium wilt | Root | Young | Nursery | [63] |
| B. licheniformis LMRE 36 | Endophyte | - | Root rot | Root | Young | Nursery | [64] |
| B. velezensis OEE1 | Endophyte | - | Olive knot | Root | Young | Nursery | [65] |
| Trichoderma asperellum Bt3, T25 | PGPF | - | Verticillium wilt | Root | Young | Nursery | [66] |
| T. harzianum CECT 2413 | PGPF | - | Verticillium wilt | Root | Young | Nursery | [67] |
| Fusarium oxysporum FO12, Phoma sp. Ph02 | PGPF | - | Verticillium wilt | Root | Young | Nursery | [68] |
| T. harziarum | PGPF | - | Verticillium wilt | Root | Adult | Orchard | [69] |
| Glomus sp., Acaulospora sp., Gigaspora sp., Entrophospora sp., Scutellospora sp., G. irregulare DAOM 197198 | AMF | - | Verticillium wilt | Root | Young | Nursery | [70] |
| Fusarium oxysporum FO12, Lactobacillus plantarum, Lactobacillus casei, Rhodobacter sphaeroides, Rhodopseudomonas palustris Saccharomyces sp., Streptococcus lactis, Streptomyces sp. | PGPR and Fungi | EM-1® | Verticillium wilt | Root | Young | Nursery | [71] |
| G. aggregatum, G. mosseae, G. clarum, G. monosporus, G. deserticola, G. brasilianum, G. etunicatum, G. margarita, R. irregularis, T. harzianum Rifai KRL-AG2 | AMF and PGPF | T-22Planter Box® | Verticillium wilt | Root | Adult | Orchard | [72] |
| Fusarium oxysporum FO12 | PGPF | Verticillium wilt | Root | Young/Adult | Nursery/Orchard | [73] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bizos, G.; Papatheodorou, E.M.; Chatzistathis, T.; Ntalli, N.; Aschonitis, V.G.; Monokrousos, N. The Role of Microbial Inoculants on Plant Protection, Growth Stimulation, and Crop Productivity of the Olive Tree (Olea europea L.). Plants 2020, 9, 743. https://doi.org/10.3390/plants9060743
Bizos G, Papatheodorou EM, Chatzistathis T, Ntalli N, Aschonitis VG, Monokrousos N. The Role of Microbial Inoculants on Plant Protection, Growth Stimulation, and Crop Productivity of the Olive Tree (Olea europea L.). Plants. 2020; 9(6):743. https://doi.org/10.3390/plants9060743
Chicago/Turabian StyleBizos, Georgios, Efimia M. Papatheodorou, Theocharis Chatzistathis, Nikoletta Ntalli, Vassilis G. Aschonitis, and Nikolaos Monokrousos. 2020. "The Role of Microbial Inoculants on Plant Protection, Growth Stimulation, and Crop Productivity of the Olive Tree (Olea europea L.)" Plants 9, no. 6: 743. https://doi.org/10.3390/plants9060743
APA StyleBizos, G., Papatheodorou, E. M., Chatzistathis, T., Ntalli, N., Aschonitis, V. G., & Monokrousos, N. (2020). The Role of Microbial Inoculants on Plant Protection, Growth Stimulation, and Crop Productivity of the Olive Tree (Olea europea L.). Plants, 9(6), 743. https://doi.org/10.3390/plants9060743