Weed Functional Diversity as Affected by Agroecological Service Crops and No-Till in a Mediterranean Organic Vegetable System

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
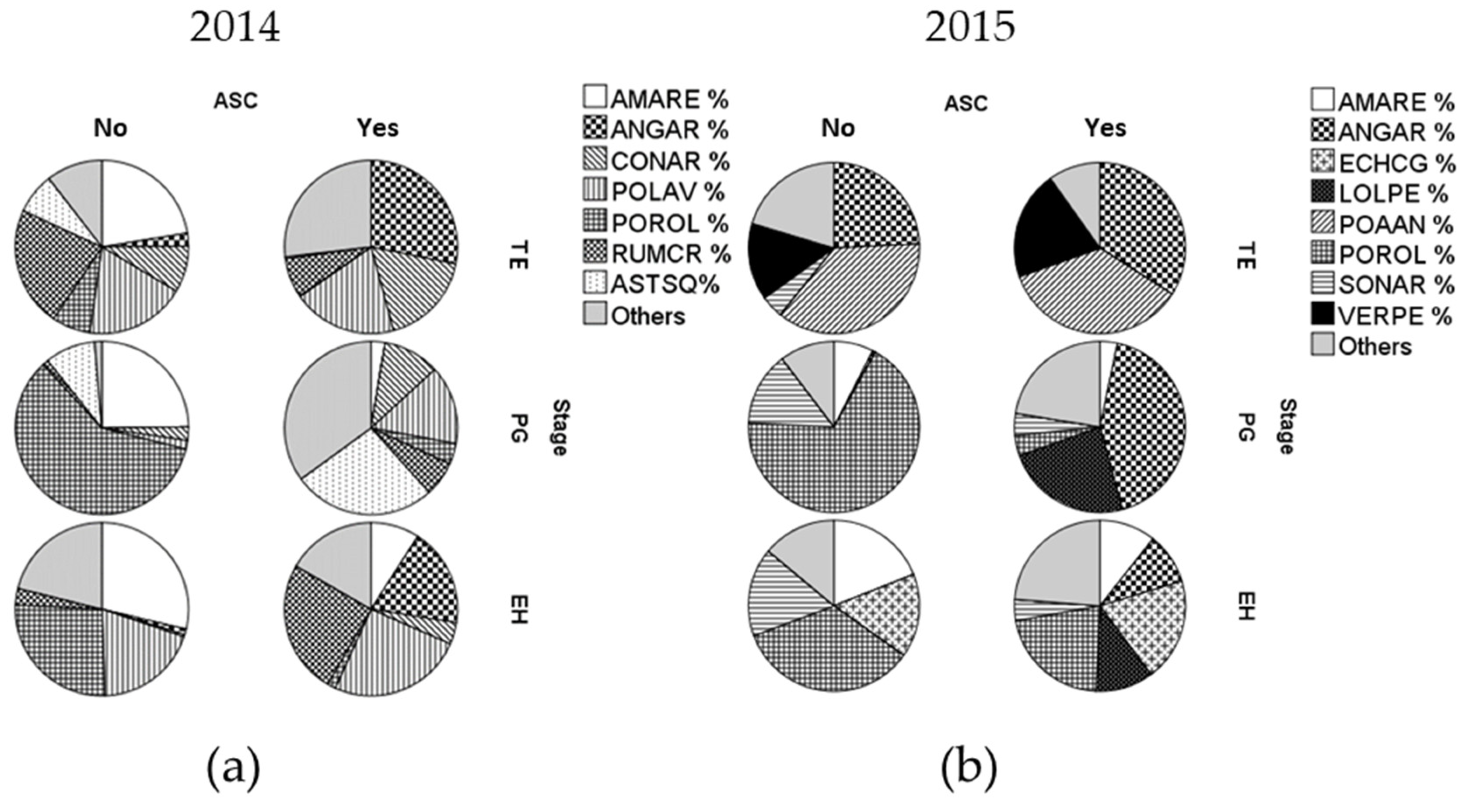
2.1. Effect of ASC Introduction on Weed Density/Cover, Species Richness, Community Composition and Functional Traits
2.2. Effect of ASC Species on Weed Density/Cover, Species Richness and Community Functional Traits
3. Discussion
3.1. Effect of ASC Introduction on Weed Density/Cover, Species Richness and Community Composition and Functional Traits
3.2. Effect of ASC Species on Weed Density/Cover, Species Richness and Community Composition and Functional Traits
4. Materials and Methods
4.1. Site Characterization
4.2. Experiment Setup and Treatments
4.3. Measurements
4.4. Selection of Traits
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Clark, M.; Williams, D.R.; Kimmel, K.; Polasky, S.; Packer, C. Future threats to biodiversity and pathways to their prevention. Nature. 2017, 546, 73. [Google Scholar] [CrossRef] [PubMed]
- Garbach, K.; Milder, J.C.; DeClerck, F.A.; Montenegro de Wit, M.; Driscoll, L.; Gemmill-Herren, B. Examining multi-functionality for crop yield and ecosystem services in five systems of agroecological intensification. Int. J. Agric. Sustain. 2017, 15, 11–28. [Google Scholar] [CrossRef]
- Wezel, A.; Casagrande, M.; Celette, F.; Vian, J.F.; Ferrer, A.; Peigné, J. Agroecological practices for sustainable agriculture. A review. Agron. Sustain. Dev. 2014, 34, 1–20. [Google Scholar] [CrossRef]
- Wood, S.A.; Karp, D.S.; DeClerck, F.; Kremen, C.; Naeem, S.; Palm, C.A. Functional traits in agriculture: Agrobiodiversity and ecosystem services. Trends Ecol. Evol. 2015, 30, 531–539. [Google Scholar] [CrossRef]
- Canali, S.; Diacono, M.; Campanelli, G.; Montemurro, F. Organic No-Till with Roller Crimpers: Agro-ecosystem Services and Applications in Organic Mediterranean Vegetable Productions. Sustain. Agric. Res. 2015, 4, 70–79. [Google Scholar] [CrossRef]
- Ciaccia, C.; Testani, E.; Roccuzzo, G.; Canali, S. The role of agrobiodiversity in sustainable food systems design and management. In Genetic Diversity in Horticultural Plants; Nandwani, D., Ed.; Springer: Cham, Switzerland, 2019; pp. 245–271. [Google Scholar]
- Ciaccia, C.; Canali, S.; Campanelli, G.; Testani, E.; Montemurro, F.; Leteo, F.; Delate, K. Effect of roller-crimper technology on weed management in organic zucchini production in a Mediterranean climate zone. Renew. Agric. Food Syst. 2016, 31, 111–121. [Google Scholar] [CrossRef]
- Gallandt, E. Weed Management in Organic Farming. In Recent Advances in Weed Management; Chauhan, B.S., Mahajan, G., Eds.; Springer: New York, NY, USA, 2014; pp. 63–85. [Google Scholar]
- Armengot, L.; Blanco-Moreno, J.M.; Bàrberi, P.; Bocci, G.; Carlesi, S.; Aendekerk, R.; Berner, A.; Celette, F.; Grosse, M.; Huiting, H.; et al. Tillage as a driver of change in weed communities: A functional perspective. Agric. Ecosyst. Environ. 2016, 222, 276–285. [Google Scholar] [CrossRef]
- Patterson, E.S.; Sanderson, R.A.; Eyre, M.D. Soil tillage reduces arthropod biodiversity and has lag effects within organic and conventional crop rotations. J. Appl. Entomol. 2019, 143, 430–440. [Google Scholar] [CrossRef]
- Kleijn, D.; Bommarco, R.; Fijen, T.P.; Garibaldi, L.A.; Potts, S.G.; van der Putten, W.H. Ecological intensification: Bridging the gap between science and practice. Trends Ecol. Evol. 2018, 34, 13. [Google Scholar] [CrossRef]
- Iocola, I.; Bassu, S.; Farina, R.; Antichi, D.; Basso, B.; Bindi, M.; Dalla Marta, A.; Danuso, F.; Doro, L.; Ferrise, R.; et al. Can conservation tillage mitigate climate change impacts in Mediterranean cereal systems? A soil organic carbon assessment using long term experiments. Eur. J. Agron. 2017, 90, 96–107. [Google Scholar] [CrossRef]
- Nichols, V.; Verhulst, N.; Cox, R.; Govaerts, B. Weed dynamics and conservation agriculture principles: A review. Field Crop Res. 2015, 183, 56–68. [Google Scholar] [CrossRef]
- Armengot, L.; Berner, A.; Blanco-Moreno, J.M.; Mäder, P.; Sans, F.X. Long-term feasibility of reduced tillage in organic farming. Agron. Sustain. Dev. 2015, 35, 339–346. [Google Scholar] [CrossRef]
- Wayman, S.; Cogger, C.; Benedict, C.; Collins, D.; Burke, I.; Bary, A. Cover crop effects on light, nitrogen, and weeds in organic reduced tillage. Agroecol. Sust. Food 2015, 39, 647–665. [Google Scholar] [CrossRef]
- Canali, S.; Campanelli, G.; Ciaccia, C.; Leteo, F.; Testani, E.; Montemurro, F. Conservation tillage strategy based on the roller crimper technology for weed control in Mediterranean vegetable organic cropping systems. Eur. J. Agron. 2013, 50, 11–18. [Google Scholar] [CrossRef]
- Antichi, D.; Sbrana, M.; Martelloni, L.; Abou Chehade, L.; Fontanelli, M.; Raffaelli, M.; Mazzoncini, M.; Peruzzi, A.; Frasconi, C. Agronomic performances of organic field vegetables managed with conservation agriculture techniques: A study from Central Italy. Agronomy 2019, 9, 810. [Google Scholar] [CrossRef]
- Navarro-Miró, D.; Blanco-Moreno, J.M.; Ciaccia, C.; Chamorro, L.; Testani, E.; Kristensen, H.L.; Hefner, M.; Tamm, K.; Bender, I.; Jakop, M.; et al. Agroecological service crops managed with roller crimper reduce weed density and weed species richness in organic vegetable systems across Europe. Agron. Sustain. Dev. 2019, 39, 55. [Google Scholar] [CrossRef]
- Testani, E.; Ciaccia, C.; Campanelli, G.; Leteo, F.; Salvati, L.; Canali, S. Mulch-Based No-Tillage Effects on Weed Community and Management in an Organic Vegetable System. Agronomy 2019, 9, 594. [Google Scholar] [CrossRef]
- Creamer, N.G.; Dabney, S.M. Killing cover crops mechanically: Review of recent literature and assessment of new research results. Am. J. Altern. Agron. 2002, 17, 32–40. [Google Scholar] [CrossRef]
- Halde, C.; Entz, M.H. Plant species and mulch application rate affected decomposition of cover crop mulches used in organic rotational no-till systems. Can. J. Plant Sci. 2016, 96, 59–71. [Google Scholar] [CrossRef]
- Radicetti, E.; Mancinelli, R.; Campiglia, E. Impact of managing cover crop residues on the floristic composition and species diversity of the weed community of pepper crop (Capsicum. annuum. L.). Crop Prot. 2013, 44, 109–119. [Google Scholar] [CrossRef]
- MacLaren, C.; Swanepoel, P.; Bennett, J.; Wright, J.; Dehnen-Schmutz, K. Cover Crop Biomass Production Is More Important than Diversity for Weed Suppression. Crop Sci. 2019, 59, 733–748. [Google Scholar] [CrossRef]
- Wayman, S.; Cogger, C.; Benedict, C.; Burke, I.; Collins, D.; Bary, A. The influence of cover crop variety, termination timing and termination method on mulch, weed cover and soil nitrate in reduced-tillage organic systems. Renew. Agric. Food Sys. 2015b, 30, 450–460. [Google Scholar] [CrossRef]
- Campiglia, E.; Radicetti, E.; Mancinelli, R. Cover crops and mulches influence weed management and weed flora composition in strip-tilled tomato (Solanum. lycopersicum.). Weed Res. 2015, 55, 416–425. [Google Scholar] [CrossRef]
- Bàrberi, P.; Bocci, G.; Carlesi, S.; Armengot, L.; Blanco-Moreno, J.M.; Sans, F.X. Linking species traits to agroecosystem services: A functional analysis of weed communities. Weed Res. 2018, 58, 76–88. [Google Scholar] [CrossRef]
- Trinchera, A.; Ciaccia, C.; Testani, E.; Baratella, V.; Campanelli, G.; Leteo, F.; Canali, S. Mycorrhiza-mediated interference between cover crop and weed in organic winter cereal agroecosystems: The mycorrhizal colonization intensity indicator. Ecol. Evol. 2019, 9, 5593–5604. [Google Scholar] [CrossRef]
- Kruidhof, H.; Bastiaans, L.; Kropff, M.J. Ecological weed management by cover cropping: Effects on weed growth in autumn and weed establishment in spring. Weed Res. 2008, 48, 492–502. [Google Scholar] [CrossRef]
- van Rijn, P.C.; Wäckers, F.L. Nectar accessibility determines fitness, flower choice and abundance of hoverflies that provide natural pest control. J. Appl. Ecol. 2016, 53, 925–933. [Google Scholar] [CrossRef]
- Barberi, P.; Burgio, G.; Dinelli, G.; Moonen, A.C.; Otto, S.; Vazzana, C.; Zanin, G. Functional biodiversity in the agricultural landscape: Relationships between weeds and arthropod fauna. Weed Res. 2010, 50, 388–401. [Google Scholar] [CrossRef]
- Reberg-Horton, S.C.; Grossman, J.M.; Kornecki, T.S.; Meijer, A.D.; Price, A.J.; Place, G.T.; Webster, T.M. Utilizing cover crop mulches to reduce tillage in organic systems in the Southeast. Renew. Agric. Food Syst. 2011, 27, 41–48. [Google Scholar] [CrossRef]
- Moreau, D.; Busset, H.; Matejicek, A.; Munier-Jolain, N. The ecophysiological determinants of nitrophily in annual weed species. Weed Res. 2014, 54, 335–346. [Google Scholar] [CrossRef]
- Gaba, S.; Fried, G.; Kazakou, E.; Chauvel, B.; Navas, M.L. Agroecological weed control using a functional approach: A review of cropping systems diversity. Agron. Sustain. Dev. 2014, 34, 103–119. [Google Scholar] [CrossRef]
- Mohler, C.L. Ecological bases for the cultural control of annual weeds. J. Prod. Agric. 1996, 9, 468–474. [Google Scholar] [CrossRef]
- Both, B.D.; Swanton, C.J. Assembly theory applied to weed communities. Weed Sci. 2002, 50, 2–13. [Google Scholar] [CrossRef]
- Liebert, J.A.; DiTommaso, A.; Ryan, M.R. Rolled Mixtures of Barley and Cereal Rye for Weed Suppression in Cover Crop–Based Organic No-Till Planted Soybean. Weed Sci. 2017, 65, 426–439. [Google Scholar] [CrossRef]
- Ciaccia, C.; Testani, E.; Campanelli, G.; Sestili, S.; Leteo, F.; Tittarelli, F.; Riva, F.; Canali, S.; Trinchera, A. Ecological service providing crops effect on melon-weed competition and allelopathic interactions. Org. Agric. 2015, 5, 199–207. [Google Scholar] [CrossRef]
- Wells, M.S.; Brinton, C.M.; Reberg-Horton, S.C. Weed Suppression and Soybean Yield in a No-till Cover-Crop Mulched System as Influenced by Six Rye Cultivars. Renew. Agric. Food Syst. 2015, 31, 429–440. [Google Scholar] [CrossRef]
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Campanelli, G.; Testani, E.; Canali, S.; Ciaccia, C.; Leteo, F.; Trinchera, A. Effects of cereals as agro-ecological service crops and no-till on organic melon, weeds and N dynamics. Biol. Agric. Hortic. 2019, 35, 275–287. [Google Scholar] [CrossRef]
- UNESCO-FAO. Etude Écologique de la Zone Méditerranéenne. Carte Bioclimatique de la zone Méditerranéenne Ecological Study of the Mediterranean Area: Bioclimatic Map of the Mediterranean Sea; UNESCO: Paris, France; FAO: Rome, Italy, 1963; p. 60. [Google Scholar]
- U.S. Department of Agriculture. Soil survey laboratory methods manual. In Natural Resource Conservation Service, Soil Survey Investigation Report No. 42, version. 3.0; U.S. Department of Agriculture: Washington, DC, USA, 1996. [Google Scholar]
- Pignatti, S. Geobotanica. In Trattato di Botanica; Cappelletti, C., Ed.; UTET: Turin, Italy, 1976; pp. 801–997. [Google Scholar]
- Wikum, D.A.; Shanholtzer, G. Application of the Braun-Blanquet cover-abundance scale for vegetation analysis in land development studies. Environ. Manag. 1978, 2, 323–329. [Google Scholar] [CrossRef]
- Pierce, S.; Negreiros, D.; Cerabolini, B.E.; Kattge, J.; Díaz, S.; Kleyer, M.; Shipley, B.; Wright, J.S.; Soudzilovskaia, N.A.; Onipchenko, V.G.; et al. A global method for calculating plant CSR ecological strategies applied across biomes world-wide. Funct. Ecol. 2017, 31, 444–457. [Google Scholar] [CrossRef]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Biondicazione attraverso le piante vascolari. Valori di indicazione secondo Ellenberg (Zeigerwerte) per le specie della Flora d’Italia. Braun-Blanquetia 2006, 39, 1–97. [Google Scholar]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package Version 1.0–1.2. 2014. Available online: http://7CRAN.R-project.org/package=FD (accessed on 25 June 2014).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 19 October 2018).
- Bates, D.; Maechler, M.; Dai, B. lme4: Linear Mixed-Effects Models Using S4 Classes. Version 0. 999375-27. 2008. Available online: http://lme4.r-forge.r-project.org/ (accessed on 19 February 2009).
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest: Tests for Random and Fixed Effects for Linear Mixed Effect Models (Lmer Objects of lme4 Package). R package version 2.0–6. 2014. Available online: http://CRAN.R-project.org/package=lmerTest (accessed on 19 October 2018).
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 33. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CRP | PRN | SBL | SWT | CH | SLA | BFF | DFF | L | T | N | R | TD/C | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ASC | ||||||||||||||
| TE | Sig. | *** | n.s. | n.s. | n.s. | n.s. | * | * | ** | ** | n.s. | n.s. | ** | * |
| PG | Sig. | n.s. | * | *** | *** | n.s. | *** | *** | *** | * | *** | *** | n.s. | *** |
| EH | Sig. | n.s. | n.s. | n.s. | ** | *** | n.s. | *** | n.s. | n.s. | n.s. | ** | * | n.s. |
| Year | ||||||||||||||
| TE | Sig. | *** | n.s. | n.s. | *** | ** | n.s. | * | *** | ** | *** | * | n.s. | n.s. |
| PG | Sig. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | *** | n.s. | n.s. | *** |
| EH | Sig. | n.s. | n.s. | n.s. | ** | *** | n.s. | *** | n.s. | n.s. | n.s. | ** | * | n.s. |
| Year x ASC | ||||||||||||||
| TE | Sig. | ** | n.s. | n.s. | n.s. | n.s. | ** | n.s. | *** | *** | ** | ** | * | n.s. |
| PG | Sig. | n.s. | n.s. | ** | n.s. | n.s. | n.s. | * | n.s. | n.s. | n.s. | n.s. | * | n.s. |
| EH | Sig. | n.s. | *** | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | * | n.s. | n.s. | n.s. |
| CRP | PRN | SBL | SWT | CH | SLA | BFF | DFF | L | T | N | R | TD/C | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TE 2014 | N < Y | n.s. | n.s. | n.s. | n.s. | N < Y | N > Y | N < Y | N > Y | n.s. | n.s. | n.s. | N > Y |
| TE 2015 | N < Y | n.s. | n.s. | n.s. | n.s. | N < Y | N > Y | N < Y | n.s. | N < Y | n.s. | N > Y | N > Y |
| PG 2014 | n.s. | n.s. | N > Y | N < Y | N < Y | N < Y | N > Y | N < Y | n.s. | N > Y | N > Y | n.s. | N > Y |
| PG 2015 | n.s. | n.s | n.s | N < Y | n.s. | N < Y | n.s. | N < Y | n.s. | N > Y | N > Y | n.s. | N > Y |
| EH 2014 | n.s. | N < Y | N > Y | N < Y | N < Y | N < Y | N > Y | n.s. | N > Y | N > Y | N > Y | N < Y | n.s. |
| EH 2015 | n.s. | n.s. | n.s. | N < Y | n.s. | N < Y | n.s. | N < Y | n.s. | n.s. | n.s. | n.s. | N > Y |
| CRP | PRN | SBL | SWT | CH | SLA | BFF | DFF | L | T | N | R | TD/C | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ASC species | ||||||||||||||
| TE | Sig. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| PG | Sig. | n.s. | n.s. | n.s. | n.s. | *** | n.s. | n.s. | ** | n.s. | ** | n.s. | * | *** |
| EH | Sig. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| Year | ||||||||||||||
| TE | Sig. | *** | n.s. | ** | *** | ** | * | n.s. | n.s. | *** | *** | *** | * | *** |
| PG | Sig. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | *** | n.s. | n.s. | *** |
| EH | Sig. | n.s. | n.s. | n.s. | ** | *** | n.s. | *** | n.s. | n.s. | n.s. | ** | * | * |
| Year x ASC sp. | ||||||||||||||
| TE | Sig. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | * |
| PG | Sig. | * | n.s. | n.s. | n.s. | *** | ** | ** | ** | n.s. | ** | n.s. | n.s. | ** |
| EH | Sig. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| CRP | PRN | SBL | SWT | CH | SLA | BFF | DFF | L | T | N | R | TD/C | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TE 2014 | n.s. | n.s. | n.s. | n.s. | n.s. | * | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | * |
| TE 2015 | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | * | * |
| PG 2014 | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s | n.s. | * | *** |
| PG 2015 | n.s. | n.s. | n.s. | n.s. | *** | * | * | * | n.s. | * | n.s. | * | * |
| EH 2014 | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | * | n.s. |
| EH 2015 | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciaccia, C.; Armengot Martinez, L.; Testani, E.; Leteo, F.; Campanelli, G.; Trinchera, A. Weed Functional Diversity as Affected by Agroecological Service Crops and No-Till in a Mediterranean Organic Vegetable System. Plants 2020, 9, 689. https://doi.org/10.3390/plants9060689
Ciaccia C, Armengot Martinez L, Testani E, Leteo F, Campanelli G, Trinchera A. Weed Functional Diversity as Affected by Agroecological Service Crops and No-Till in a Mediterranean Organic Vegetable System. Plants. 2020; 9(6):689. https://doi.org/10.3390/plants9060689
Chicago/Turabian StyleCiaccia, Corrado, Laura Armengot Martinez, Elena Testani, Fabrizio Leteo, Gabriele Campanelli, and Alessandra Trinchera. 2020. "Weed Functional Diversity as Affected by Agroecological Service Crops and No-Till in a Mediterranean Organic Vegetable System" Plants 9, no. 6: 689. https://doi.org/10.3390/plants9060689
APA StyleCiaccia, C., Armengot Martinez, L., Testani, E., Leteo, F., Campanelli, G., & Trinchera, A. (2020). Weed Functional Diversity as Affected by Agroecological Service Crops and No-Till in a Mediterranean Organic Vegetable System. Plants, 9(6), 689. https://doi.org/10.3390/plants9060689