Comparing Salt Tolerance at Seedling and Germination Stages in Local Populations of Medicago ciliaris L. to Medicago intertexta L. and Medicago scutellata L.

 ,
,  , , ,
, , , 

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Effect of Salt on Seed Germination
2.1.1. Salt Stress Effect on Final Germination Percentage (FGP)
2.1.2. Salt Stress Effect on Germination Rate (T50)
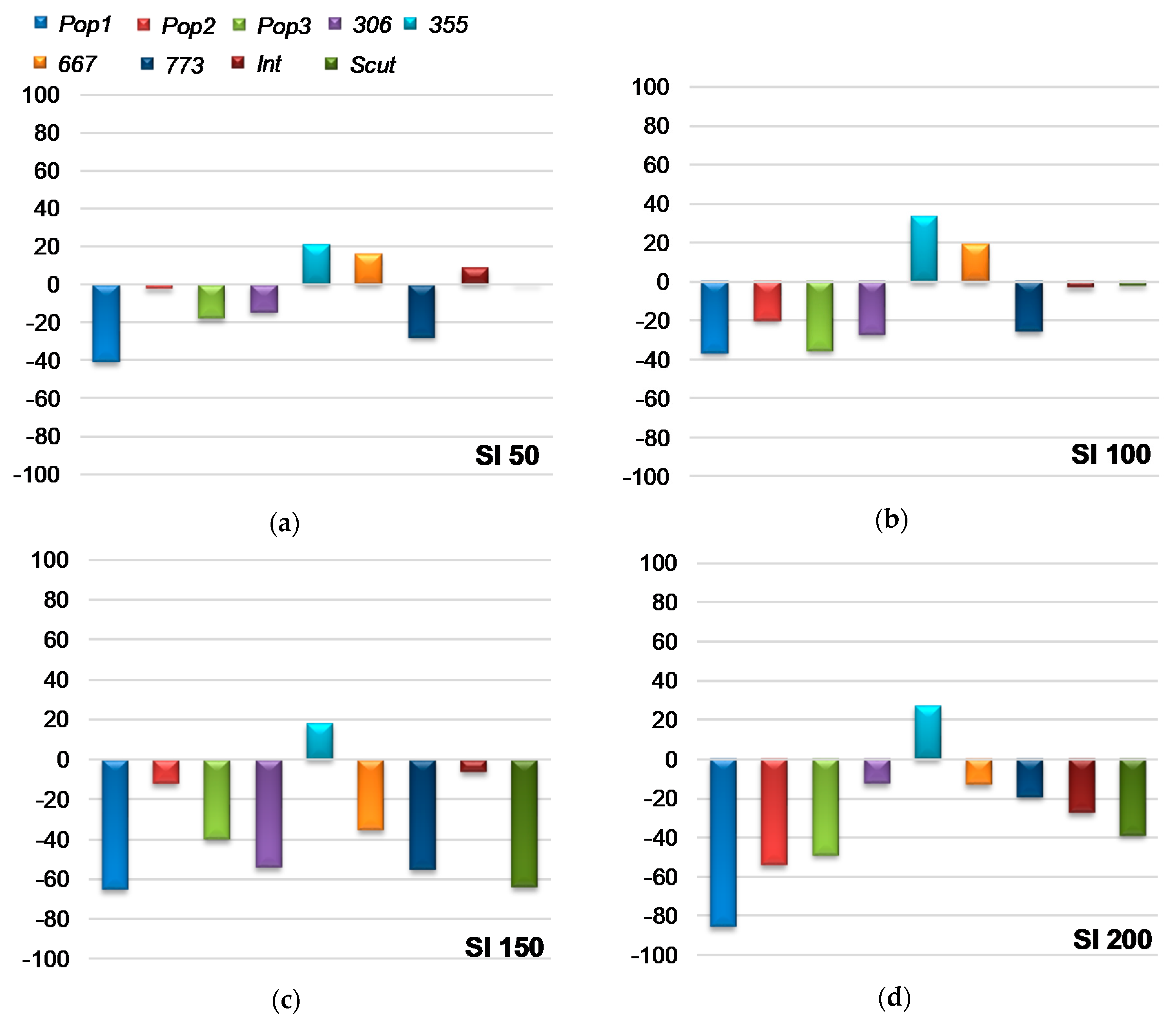
2.1.3. Sensitivity Index to Salt Stress for Seed Germination (SI)
2.2. Hydroponic Culture Assay
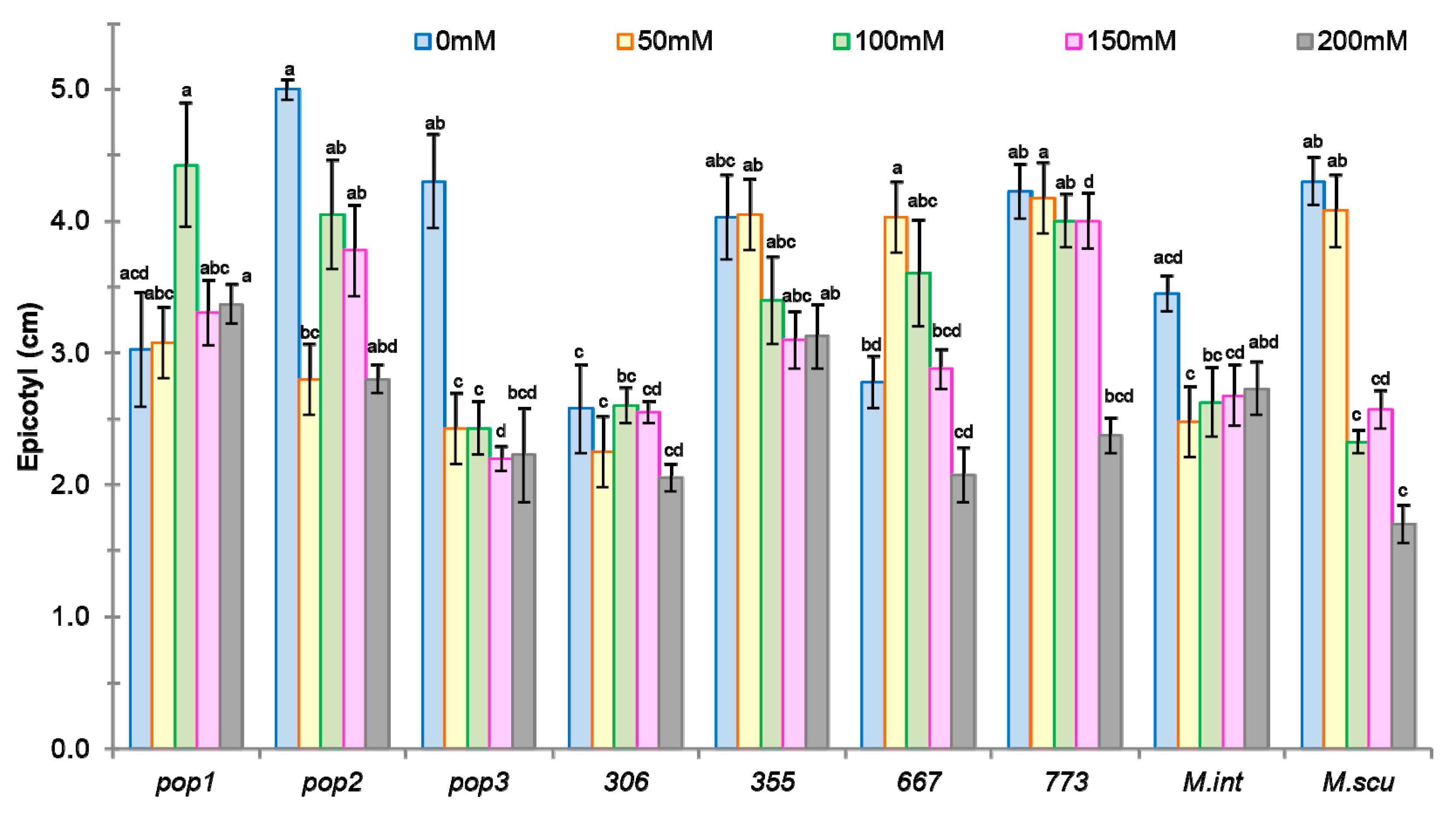
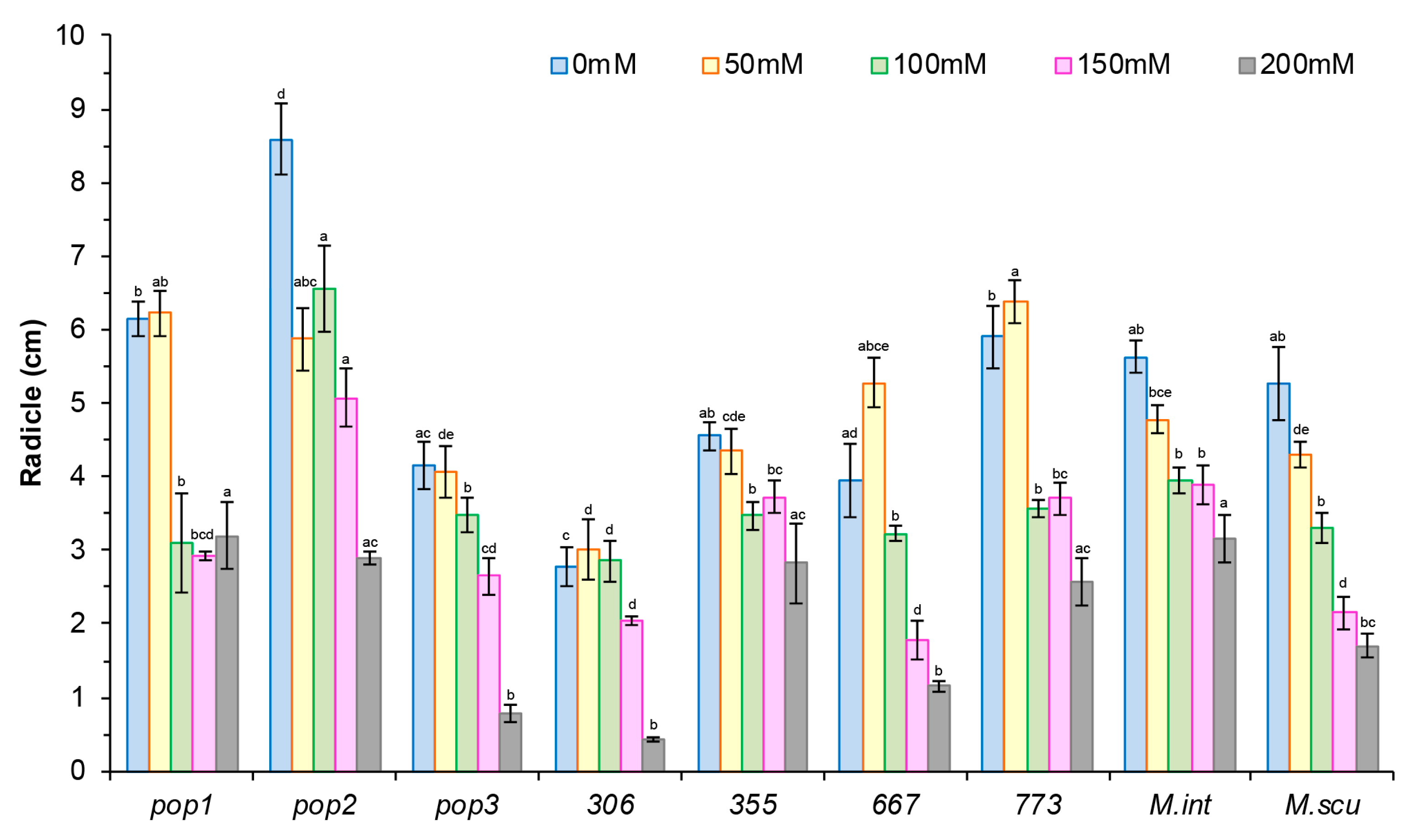
2.2.1. Effect of NaCl on Growth
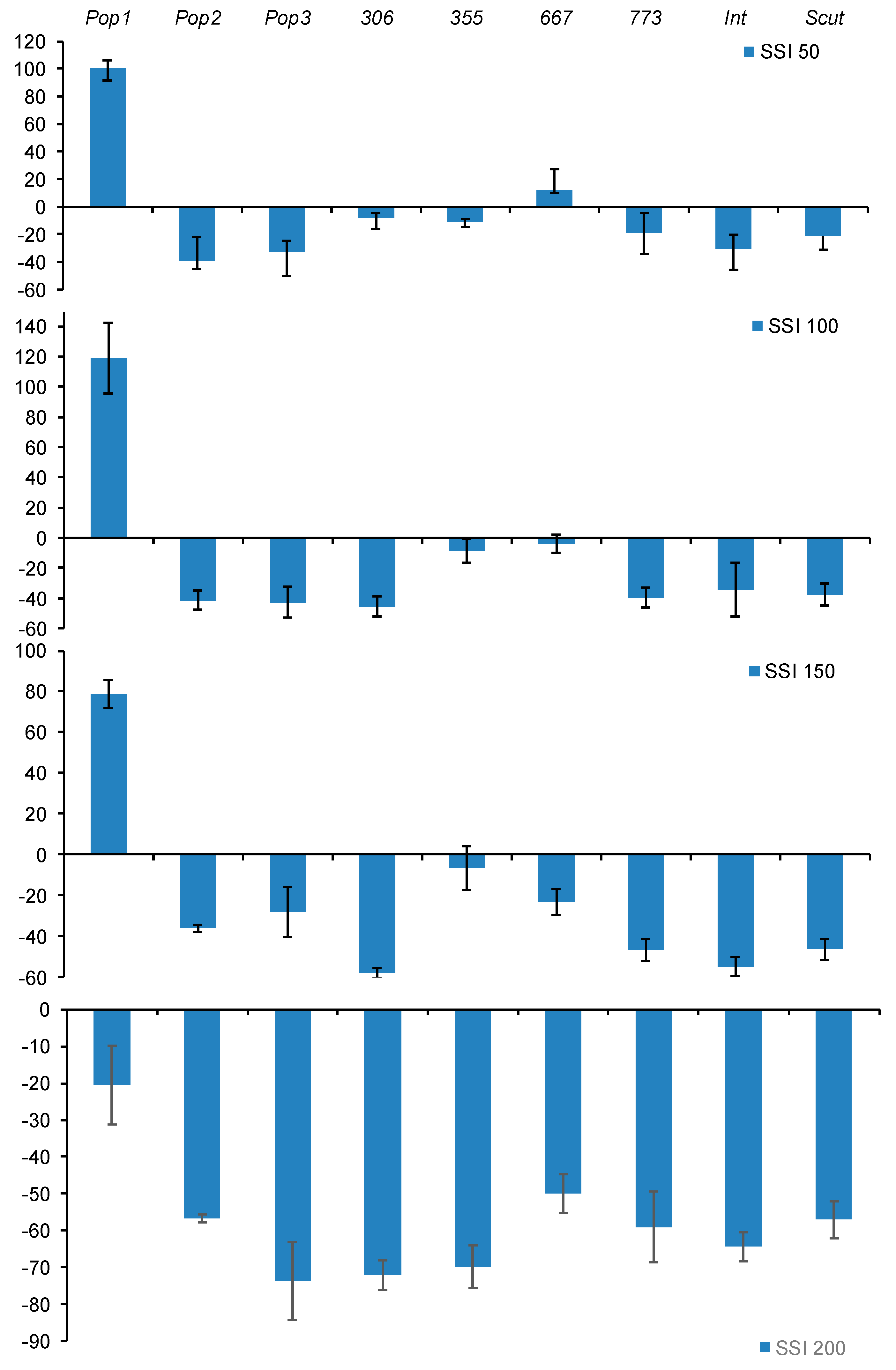
2.2.2. The Sensitivity Index to Salt Stress for Growth (SI), Noted SI(DW)
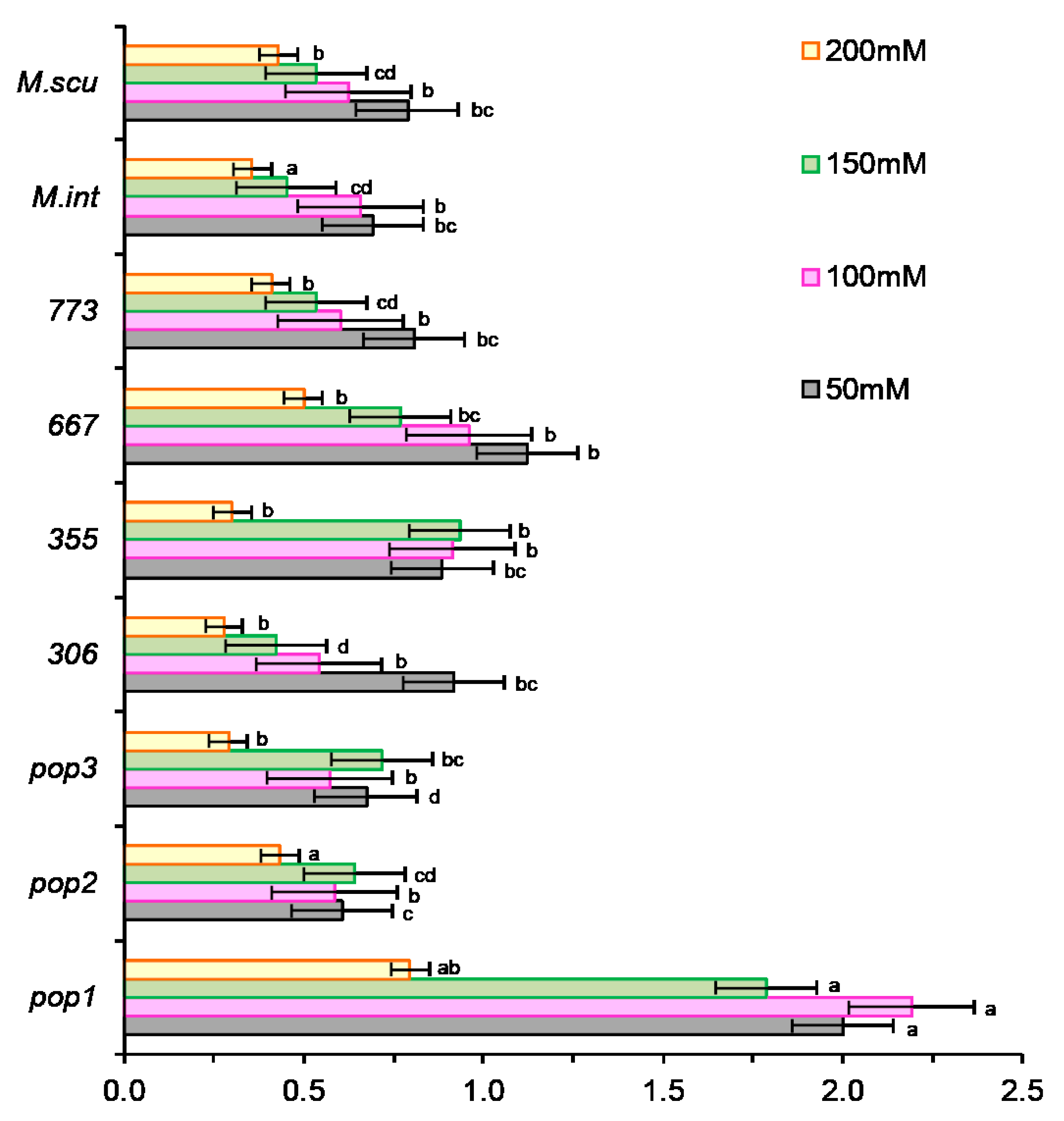
2.2.3. The Salt Tolerance Index at Early Growing Stage (TI)
2.3. Biochemical Parameters
2.3.1. Chlorophyll a Content
2.3.2. Chlorophyll b and Total Chlorophyll Content
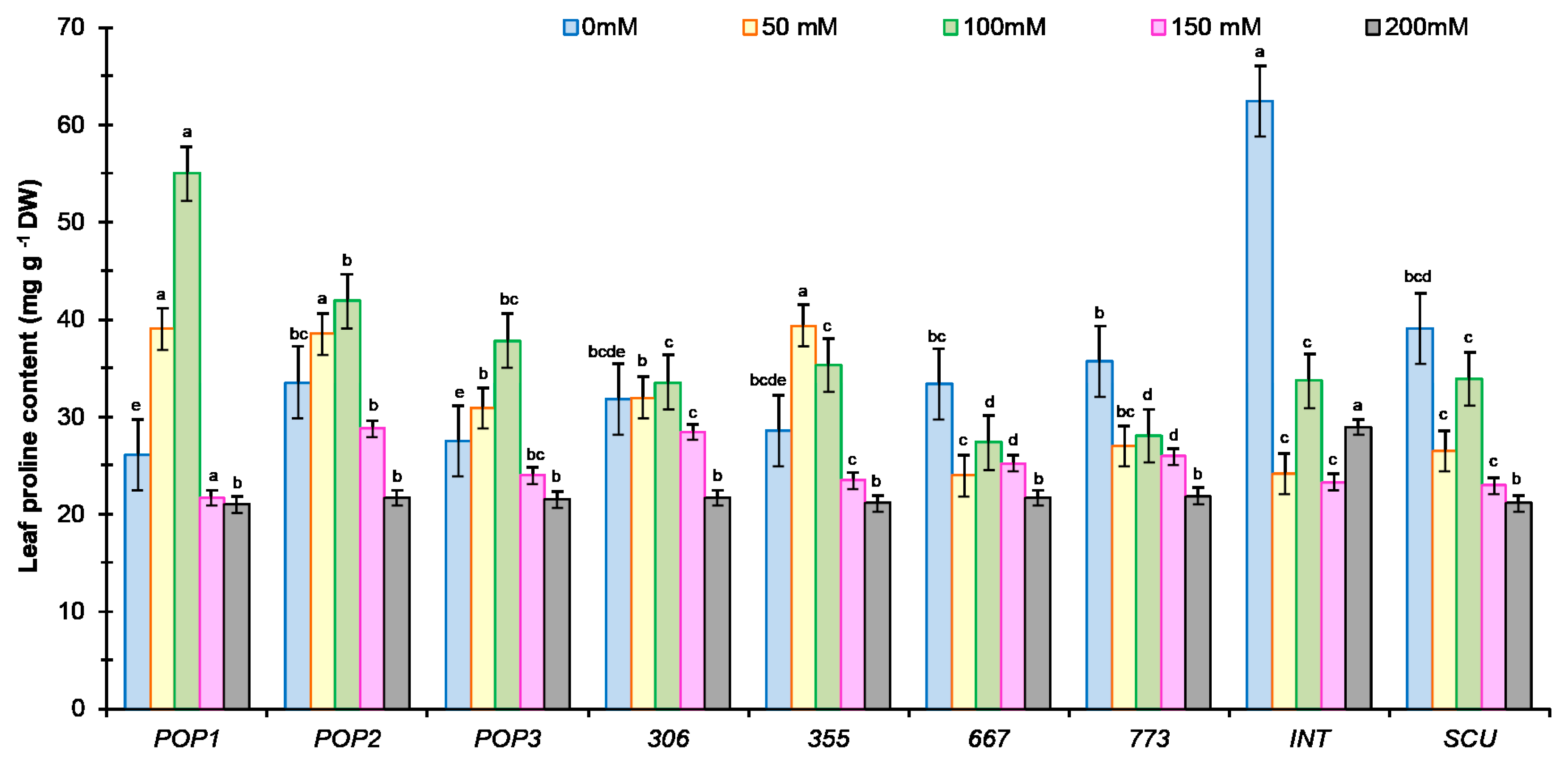
2.3.3. Proline Content
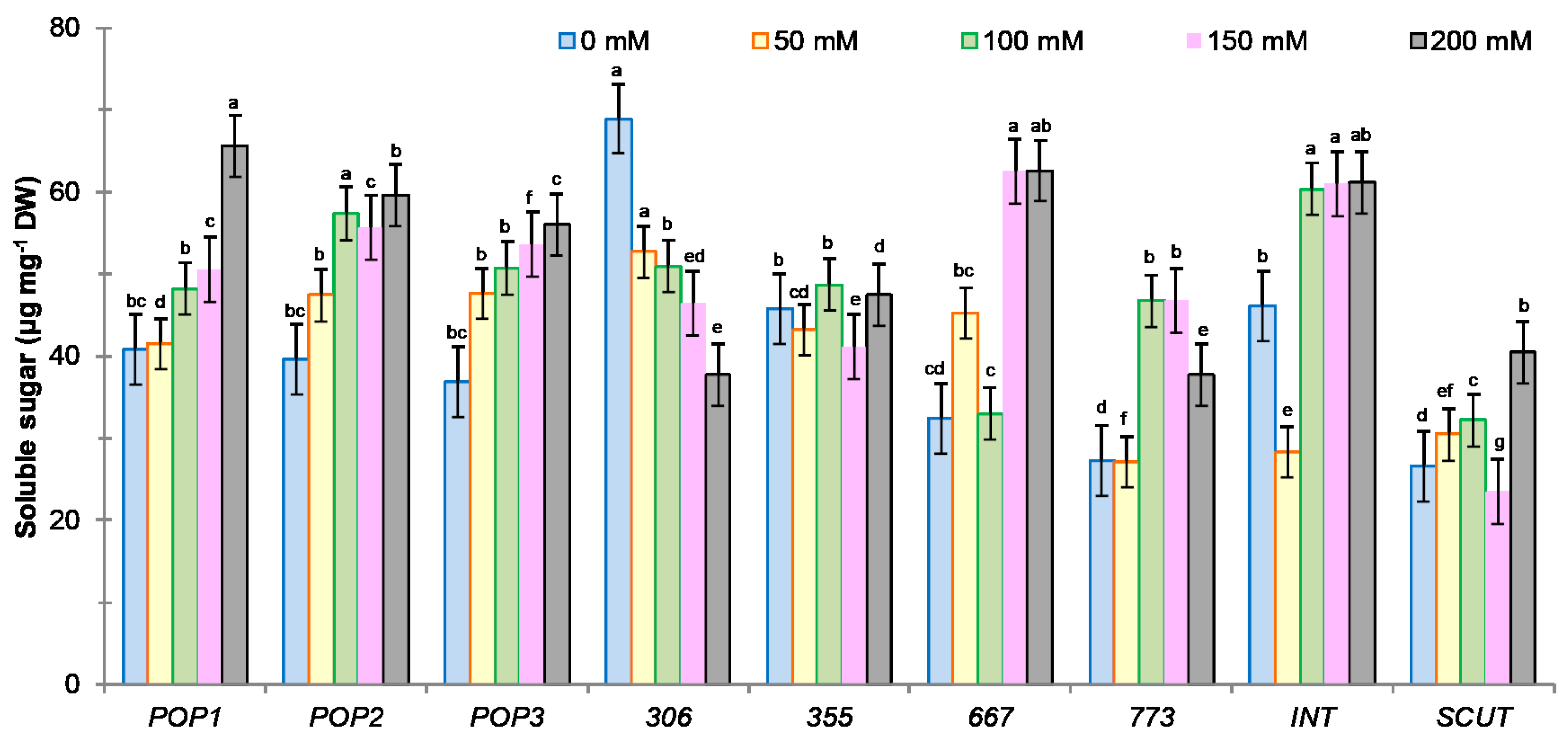
2.3.4. Sugar Content
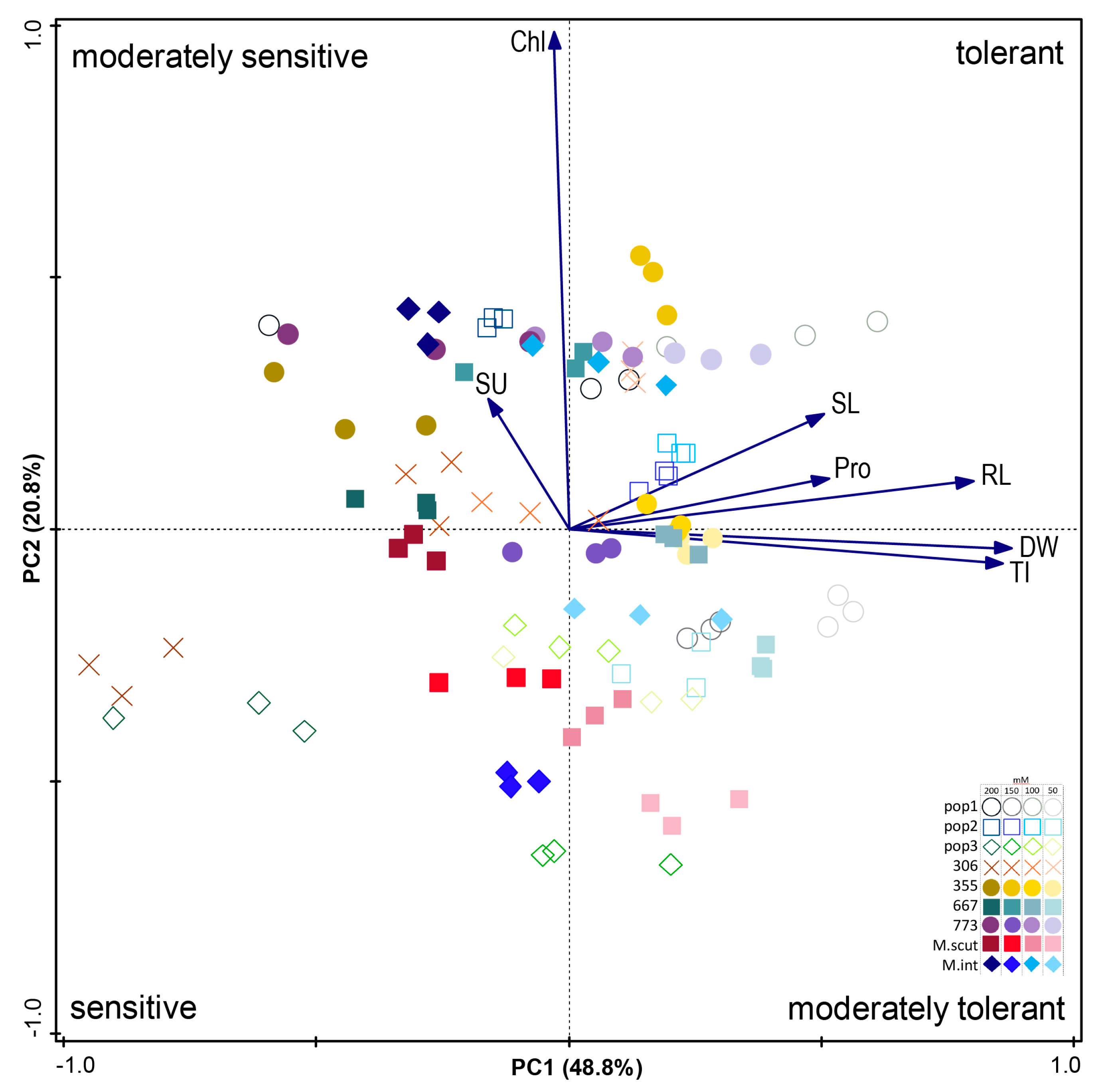
2.3.5. PCA Analysis for Traits
3. Discussion
4. Materials and Methods
4.1. Plant Genotypes and Seed Sources
4.2. Germination Assays
4.3. Hydroponic Culture Assay under Salt Stress
4.4. Parameters Measured
4.5. Chlorophyll Content
- Chl a (μg mL−1) = 12.25.·A663 − 2.79.·A647
- Chl b (μg mL−1) = 21.5.·A647 − 5.10.·A663
- Total Chlorophyll (μg mL−1) = (20.2 × A645) + (8.02 × A663)
4.6. Proline Content
4.7. Total Sugars
4.8. Salt Tolerance Index
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shokat, S.; Großkinsky, D.K. Tackling salinity in sustainable agriculture-What developing countries may learn from approaches of the developed world. Sustainability 2019, 11, 4558. [Google Scholar] [CrossRef]
- Minhas, P.S.; Ramos, T.B.; Ben-Gal, A.; Pereira, L.S. Coping with salinity in irrigated agriculture: Crop evapotranspiration and water management issues. Agric. Water Manag. 2020, 227, 105832. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Jensen, C.R.; Liu, F. Improving crop production in the arid Mediterranean climate. Field Crop. Res. 2012, 128, 34–47. [Google Scholar] [CrossRef]
- Srivastava, N. Reclamation of Saline and Sodic Soil Through Phytoremediation. In Environmental Concerns and Sustainable Development; Springer: Singapore, 2020; pp. 279–306. [Google Scholar]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Louati, D.; Majdoub, R.; Rigane, H.; Abida, H. Effects of Irrigating with Saline Water on Soil Salinization (Eastern Tunisia). Arab. J. Sci. Eng. 2018, 43, 3793–3805. [Google Scholar] [CrossRef]
- Mbarki, S.; Sytar, O.; Cerda, A.; Zivcak, M.; Rastogi, A.; He, X.; Zoghlami, A.; Abdelly, C.; Brestic, M. Strategies to mitigate the salt stress effects on photosynthetic apparatus and productivity of crop plants. In Salinity Responses and Tolerance in Plants, Volume 1: Targeting Sensory, Transport and Signaling Mechanisms; Springer International Publishing: Cham, Switzerland, 2018; pp. 85–136. ISBN 9783319756714. [Google Scholar]
- Safdar, H.; Amin, A.; Shafiq, Y.; Ali, A.; Yasin, R. A review: Impact of salinity on plant growth. Nat. Sci. 2019, 17, 34–40. [Google Scholar] [CrossRef]
- Acosta-motos, J.R.; Penella, C.; Hernández, J.A.; Sánchez-blanco, M.J.; Navarro, J.M.; Gómez-bellot, M.J. Towards a Sustainable Agriculture: Strategies Involving Phytoprotectants against Salt Stress. Agronomy 2020, 10, 194. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; Mann, A.; Devi, G.; Sharma, H.; Singh, R.; Sanwal, S.K. Phytoamelioration of the Salt-Affected Soils Through Halophytes. In Ecophysiology, Abiotic Stress Responses and Utilization of Halophytes; Springer: Singapore, 2019; pp. 313–326. [Google Scholar]
- Ismail, S.; Rao, N.K.; Dagar, J.C. Identification, Evaluation, and Domestication of Alternative Crops for Saline Environments. In Research Developments in Saline Agriculture; Springer: Berlin, Germany, 2019; pp. 505–536. [Google Scholar]
- Abiala, M.A.; Abdelrahman, M.; Burritt, D.J.; Tran, L.S.P. Salt stress tolerance mechanisms and potential applications of legumes for sustainable reclamation of salt-degraded soils. Land Degrad. Dev. 2018, 29, 3812–3822. [Google Scholar] [CrossRef]
- Mbarki, S.; Cerdà, A.; Zivcak, M.; Brestic, M.; Rabhi, M.; Mezni, M.; Jedidi, N.; Abdelly, C.; Pascual, J.A. Alfalfa crops amended with MSW compost can compensate the effect of salty water irrigation depending on the soil texture. Process. Saf. Environ. Prot. 2018, 115, 8–16. [Google Scholar] [CrossRef]
- Morton, M.J.L.; Awlia, M.; Al-Tamimi, N.; Saade, S.; Pailles, Y.; Negrão, S.; Tester, M. Salt stress under the scalpel-dissecting the genetics of salt tolerance. Plant. J. 2019, 97, 148–163. [Google Scholar] [CrossRef]
- Venâncio, C.; Pereira, R.; Lopes, I. The influence of salinization on seed germination and plant growth under mono and polyculture. Environ. Pollut. 2020, 260, 113993. [Google Scholar] [CrossRef]
- Hernández, J.A. Salinity tolerance in plants: Trends and perspectives. Int. J. Mol. Sci. 2019, 20, 2408. [Google Scholar] [CrossRef] [PubMed]
- Kotula, L.; Kwa, H.Y.; Nichols, P.G.H.; Colmer, T.D. Tolerance and recovery of the annual pasture legumes Melilotus siculus, Trifolium michelianum and Medicago polymorpha to soil salinity, soil waterlogging and the combination of these stresses. Plant. Soil 2019, 444, 267–280. [Google Scholar] [CrossRef]
- Smith, A.P.; Moore, A.D. Whole farm implications of lucerne transitions in temperate crop-livestock systems. Agric. Syst. 2020, 177, 102686. [Google Scholar] [CrossRef]
- Nedjimi, B.; Zemmiri, H. Salinity Effects on Germination of Artemisia herba–alba Asso: Important Pastoral Shrub from North African Rangelands. Rangel. Ecol. Manag. 2019, 72, 189–194. [Google Scholar] [CrossRef]
- Natasha, K. Effect of sodium chloride, potassium chloride on germination and growth of Foxtail millet (Setaria italica L.). Pure Appl. Biol. 2019, 8, 1398–1407. [Google Scholar] [CrossRef]
- Bouslama, M.; Denden, M.; Hajlaoui, H. Etude de la variabilité intraspécifique de tolérance au stress salin du pois chiche (Cicer arietinum L.) au stade germination. Tropicultura 2007, 25, 168–173. [Google Scholar]
- Ouerghi, K.; Abdi, N.; Maazaoui, H.; Hmissi, I.; Bouraoui, M.; Sifi, B. Physiological and morphological characteristics of pea (Pisum sativum L.) seeds under salt stress. J. New Sci. 2016, 28, 1559–1565. [Google Scholar]
- Debez, A.; Ben Slimen, I.D.; Bousselmi, S.; Atia, A.; Farhat, N.; El Kahoui, S.; Abdelly, C. Comparative analysis of salt impact on sea barley from semi-arid habitats in Tunisia and cultivated barley with special emphasis on reserve mobilization and stress recovery aptitude. Plant. Biosyst. 2019, 153, 1–9. [Google Scholar] [CrossRef]
- Rahman, M.M.; Mostofa, M.G.; Rahman, M.A.; Miah, M.G.; Saha, S.R.; Karim, M.A.; Keya, S.S.; Akter, M.; Islam, M.; Tran, L.S.P. Insight into salt tolerance mechanisms of the halophyte Achras sapota: An important fruit tree for agriculture in coastal areas. Protoplasma 2019, 256, 181–191. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenomenon. Planta 2020, 251, 3. [Google Scholar] [CrossRef]
- Nawaz, K.; Hussain, K.; Majeed, A.; Khan, F.; Afghan, S.; Ali, K. Fatality of salt stress to plants: Morphological, physiological and biochemical aspects. Afr. J. Biotechnol. 2010, 9, 5475–5480. [Google Scholar]
- Bayuelo-Jiménez, J.S.; Craig, R.; Lynch, J.P. Salinity tolerance of Phaseolus species during germination and early seedling growth. Crop. Sci. 2002, 42, 1584–1594. [Google Scholar] [CrossRef]
- Rashid, M.H.O.; Islam, S.; Bari, M. In vitro screening for salt stress tolerance of native and exotic potato genotypes by morphological and physiological parameters. J. Bio-Sci. 2019, 28, 21–32. [Google Scholar] [CrossRef]
- Taffouo, V.; Meguekam, L.; Kenne, M.; Yayi, E.; Magnitsop, A.; Akoa, A.; Ourry, A. Germination et accumulation des métabolites chez les plantules de légumineuses cultivées sous stress salin. Agron. Afr. 2009, 20, 129–139. [Google Scholar] [CrossRef][Green Version]
- Ibrahim, M.H.; Abas, N.A.; Zahra, S.M. Impact of Salinity Stress on Germination of Water Spinach (Ipomoea aquatica). Annu. Res. Rev. Biol. 2019, 31, 1–12. [Google Scholar] [CrossRef]
- Lovato, M.B.; Martins, P.S. Genetic variability in salt tolerance during germination of Stylosanthes humilis H.B.K. and association between salt tolerance and isozymes. Braz. J. Genet. 1997, 20, 435–441. [Google Scholar] [CrossRef]
- Kayess, M.O.; Rahman, M.L.; Ahmed, K.; Khan, M.R.; Hossan, M.S.; Hossain, M.S.; Khanam, M.M.; Chandra Pal, D. Effect of Salinity Stress on Different Root and Shoot Traits of Selected Tomato Cultivars. Asian J. Adv. Res. Rep. 2020, 8, 1–9. [Google Scholar] [CrossRef]
- Farissi, M.; Ghoulam, C.; Bouizgaren, A. Changes in water deficit saturation and photosynthetic pigments of Alfalfa populations under salinity and assessment of proline role in salt tolerance. Agric. Sci. Res. J. 2013, 3, 29–35. [Google Scholar]
- Moghaddam, M.; Farhadi, N.; Panjtandoust, M.; Ghanati, F. Seed germination, antioxidant enzymes activity and proline content in medicinal plant Tagetes minuta under salinity stress. Plant. Biosyst. 2019, 153, 1–8. [Google Scholar] [CrossRef]
- Pujol, J.A.; Calvo, J.F.; Ramírez-Díaz, L. Recovery of germination from different osmotic conditions by four halophytes from southeastern Spain. Ann. Bot. 2000, 85, 279–286. [Google Scholar] [CrossRef]
- Mirza, H.; Kamrun, N.; Masayuki, F.; Hirosuke, O.; Islam, T.M.; Shahzad, B.; Fahad, S.; Tanveer, M.; Saud, S.; Khan, I.A. Plant Responses and Tolerance to Salt Stress. In Approaches for Enhancing Abiotic Stress Tolerance in Plants; CRC Press: Boca Raton, FL, USA, 2019; pp. 61–78. [Google Scholar]
- Shoukat, E.; Ahmed, M.Z.; Abideen, Z.; Azeem, M.; Ibrahim, M.; Gul, B.; Khan, M.A. Short and long term salinity induced differences in growth and tissue specific ion regulation of Phragmites karka. Flora Morphol. Distrib. Funct. Ecol. Plants 2020, 263, 151550. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Klapaki, G. Response of two greenhouse pepper hybrids to NaCl salinity during different growth stages. Sci. Hortic. Amst. 2000, 86, 247–260. [Google Scholar] [CrossRef]
- Mahdavi, B.; Sanavy, S.A.M.M. Germination and seedling growth in grasspea (Lathyrus sativus) cultivars under salinity conditions. Pak. J. Biol. Sci. 2007, 10, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Shi, F.; Fukuda, K.; Yang, Y. Effects of salt and alkali stresses on germination, growth, photosynthesis and ion accumulation in alfalfa (Medicago sativa L.). Soil Sci. Plant. Nutr. 2010, 56, 725–733. [Google Scholar] [CrossRef]
- Ranjbar, G.H. Anagholi, a Evaluation of physiological indices of salinity tolerance in forage Sorghum (Sorghum bicolor) lines. Int. Res. J. Appl. Basic Sci. 2012, 3, 305–308. [Google Scholar]
- Benabderrahim, M.A.; Guiza, M.; Haddad, M. Genetic diversity of salt tolerance in tetraploid alfalfa (Medicago sativa L.). Acta Physiol. Plant. 2020, 42, 5. [Google Scholar] [CrossRef]
- Bhatt, A.; Bhat, N.R.; Santo, A.; Phartyal, S.S. Influence of Temperature, Light and Salt on the Germination of Deverra Triradiata Seeds. Seed Sci. Technol. 2019, 47, 25–31. [Google Scholar] [CrossRef]
- Bukhari, S.A.H.; Peerzada, A.M.; Javed, M.H.; Dawood, M.; Hussain, N.; Ahmad, S. Growth and Development Dynamics in Agronomic Crops Under Environmental Stress. In Agronomic Crops; Springer: Singapore, 2019; pp. 83–114. [Google Scholar]
- Thouraya, R.; Imen, I.; Imen, H.; Riadh, I.; Ahlem, B.; Hager, J. Effet du stress salin sur le comportement physiologique et métabolique de trois variétés de piment (Capsicum annuum L.). J. Appl. Biosci. 2013, 66, 5060. [Google Scholar] [CrossRef][Green Version]
- Wu, H.; Guo, J.; Wang, C.; Li, K.; Zhang, X.; Yang, Z.; Li, M.; Wang, B. An effective screening method and a reliable screening trait for salt tolerance of Brassica napus at the germination stage. Front. Plant. Sci. 2019, 10, 530. [Google Scholar] [CrossRef]
- Naeem, M.S.; Jin, Z.L.; Wan, G.L.; Liu, D.; Liu, H.B.; Yoneyama, K.; Zhou, W.J. 5-Aminolevulinic acid improves photosynthetic gas exchange capacity and ion uptake under salinity stress in oilseed rape (Brassica napus L.). Plant. Soil 2010, 332, 405–415. [Google Scholar] [CrossRef]
- Poór, P.; Czékus, Z.; Ördög, A. Role and Regulation of Glucose as a Signal Molecule to Salt Stress. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 193–205. [Google Scholar]
- Jawahar, G.; Rajasheker, G.; Maheshwari, P.; Punita, D.L.; Jalaja, N.; Kumari, P.H.; Kumar, S.A.; Afreen, R.; Karumanchi, A.R.; Rathnagiri, P.; et al. Osmolyte Diversity, Distribution, and Their Biosynthetic Pathways. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 449–458. [Google Scholar]
- Lucho, S.R.; do Amaral, M.N.; Auler, P.A.; Bianchi, V.J.; Ferrer, M.Á.; Calderón, A.A.; Braga, E.J.B. Salt Stress-Induced Changes in In Vitro Cultured Stevia rebaudiana Bertoni: Effect on Metabolite Contents, Antioxidant Capacity and Expression of Steviol Glycosides-Related Biosynthetic Genes. J. Plant. Growth Regul. 2019, 38, 1341–1353. [Google Scholar] [CrossRef]
- Hamrouni, L.; Hanana, M.; Abdelly, C.; Ghorbel, A. Exclusion du chlorure et inclusion du sodium: Deux mécanismes concomitants de tolérance à la salinité chez la vigne sauvage Vitis vinifera subsp. sylvestris (var. ’séjnène’). Biotechnol. Agron. Soc. Environ. 2011, 15, 387–400. [Google Scholar]
- Shafi, A.; Zahoor, I.; Mushtaq, U. Proline Accumulation and Oxidative Stress: Diverse Roles and Mechanism of Tolerance and Adaptation Under Salinity Stress. In Salt Stress, Microbes, and Plant Interactions: Mechanisms and Molecular Approaches; Springer: Singapore, 2019; pp. 269–300. [Google Scholar]
- Hasanuzzaman, M.; Fujita, M.; Oku, H.; Islam, M.T.; Khanna-Chopra, R.; Kumar Semwal, V.; Lakra, N.; Pareek, A. Proline—A Key Regulator Conferring Plant Tolerance to Salinity and Drought. In Plant Tolerance to Environmental Stress; CRC Press: Boca Raton, FL, USA, 2019; pp. 59–80. [Google Scholar]
- Forlani, G.; Bertazzini, M.; Cagnano, G. Stress-driven increase in proline levels, and not proline levels themselves, correlates with the ability to withstand excess salt in a group of 17 Italian rice genotypes. Plant. Biol. 2019, 21, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Cheima, J. Genetic diversity of pod traits in local populations of Medicago ciliaris L. Iosr J. Agric. Vet. Sci. 2012, 1, 44–48. [Google Scholar] [CrossRef]
- Roberts, S.J.; Koenraadt, H. Validated Seed Health Testing Methods. In International Rules for Seed Testing; ISTA(The International Seed Testing Association): Basserdorf, Switzerland, 2020; pp. 1–276. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants Without Soil; Circular Agricultural Experiment Station, University of California: Berkeley, CA, USA, 1950; Volume 347, No.2nd edit; p. 32. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant. Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Do, B.C.; Dang, T.T.; Berrin, J.G.; Haltrich, D.; To, K.A.; Sigoillot, J.C.; Yamabhai, M. Cloning, expression in Pichia pastoris, and characterization of a thermostable GH5 mannan endo-1,4-beta-mannosidase from Aspergillus niger BK01. Microb. Cell Fact. 2009, 8, 59. [Google Scholar] [CrossRef]
- El Goumi, Y.; Fakiri, M.; Lamsaouri, O.; Benchekroun, M. Salt stress effect on seed germination and some physiological traits in three Moroccan barley (Hordeum vulgare L.) cultivars. J. Mater. Environ. Sci. 2014, 5, 625–632. [Google Scholar]
- Smilauer, P.; Leps, J. Multivariate Analysis of Ecological Data Using Canoco 5; Cambridge University Press: Cambridge, UK, 2014; ISBN 9781139627061. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Populations | Salt Concentration | ||||
|---|---|---|---|---|---|
| 0 mM | 50 mM | 100 mM | 150 mM | 200 mM | |
| Pop1 | 98a | 70bc | 63bc | 34de | 22bc |
| Pop2 | 100a | 100a | 85ab | 88a | 24bc |
| Pop3 | 100a | 66.7bc | 57c | 54cd | 38.4b |
| 306 | 95ab | 85ab | 65c | 44cde | 16c |
| 355 | 80bc | 86.7ab | 83abc | 78ab | 21bc |
| 667 | 62.5d | 75bc | 80bc | 40cde | 26.4bc |
| 773 | 95ab | 56.7c | 63bc | 42cde | 30.4bc |
| M. scut | 67.5cd | 70bc | 90ab | 60bc | 21bc |
| M. int | 100a | 100a | 95a | 46e | - |
| Factors | Average | Sum Square | Average Square | F | p |
|---|---|---|---|---|---|
| salt concentration(S) | 60.7 | 179,795.5556 | 35,959.1111 | 180.05 | <0.0001 |
| Population (Pop) | 15,020 | 1877.5000 | 9.40 | <0.0001 | |
| Repetition (Rep) | 739.2593 | 184.8148 | 0.93 | 0.4501 | |
| Salt×Pop | 42,984.4444 | 1074.6111 | 5.38 | <0.0001 |
| Population | NaCl Concentration | ||||
|---|---|---|---|---|---|
| 0 mM | 50 mM | 100 mM | 150 mM | 200 mM | |
| Pop1 | 1.8ab | 2.2a | 1.8ab | 1.2a | 1.6a |
| Pop2 | 1.2b | 1.6a | 1b | 1.4a | 3a |
| Pop3 | 1.6ab | 2.8a | 3.6a | 3.6a | 2.6a |
| 306 | 1.6ab | 2.2a | 3.6a | 2.6a | 1.6a |
| 355 | 1.8ab | 1.8a | 1d | 3.4a | 1.2a |
| 667 | 2a | 2.2a | 2.2bc | 3a | 2.6a |
| 773 | 1.2b | 1.2a | 1.6cd | 2.8a | 1.4a |
| M. scut | 1.8ab | 1.6a | 2.8ab | 3.4a | 3.4a |
| M. int | 1.6ab | 1.2a | 1.2cd | 2a | 3.4a |
| Means of treatments (n = 10 seeds/5 repetitions) | 1.7b | 1.9b | 2.3ab | 2.6a | 2.3ab |
| Characteristic | MS Populations | MS Salt Stress | MS Interaction | F Population | F Salt Stress | F Interaction |
|---|---|---|---|---|---|---|
| DW g plant−1 | 0.004556 | 0.049997 | 0.002733 | 8.918**** | 97.872**** | 5.360**** |
| Epicotyl (cm) | 4.477 | 7.454 | 1.508 | 17.219**** | 28.670**** | 5.800**** |
| Radicle (cm) | 21.744 | 60.324 | 2.104 | 54.885**** | 152.262**** | 5.310**** |
| NaCl Concentration (mM) | Populations | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Pop1 | Pop2 | Pop3 | 306 | 355 | 667 | 773 | M. int | M. Scut | |
| Chl a | |||||||||
| 0 | 1.09 ± 0.03c | 0.72 ± 0.03d | 0.52 ± 0.02e | 2.34 ± 0.02a | 0.3 ± 0.02f | 1.21 ± 0.06b | 0.57 ± 0.01e | 0.34 ± 0.02f | 0.53 ± 0.01e |
| 50 | 0.71 ± 0.01c | 0.48 ± 0.02e | 0.24 ± 0.003g | 1.17 ± 0.01a | 0.60 ± 0.01d | 0.58 ± 0.03d | 1.14 ± 0.01a | 0.78 ± 0.01b | 0.29 ± 0.04f |
| 100 | 1.23 ± 0.03c | 0.53 ± 0.003g | 0.24 ± 0.02h | 0.68 ± 0.01f | 0.93 ± 0.04d | 0.76 ± 0.02e | 1.30 ± 0.03b | 1.57 ± 0.01a | 0.49 ± 0.01g |
| 150 | 0.72 ± 0.005c | 0.73 ± 0.01c | 0.36 ± 0.01f | 0.61 ± 0.03de | 0.57 ± 0.01e | 1.37 ± 0.01a | 0.92 ± 0.005b | 0.24 ± 0.03g | 0.65 ± 0.01d |
| 200 | 0.36 ± 0.01f | 1.11 ± 0.02a | 0.15 ± 0.01h | 0.26 ± 0.01g | 0.49 ± 0.02e | 0.87 ± 0.01d | 0.96 ± 0.005c | 1.03 ± 0.005b | 0.48 ± 0.05e |
| Chl b | |||||||||
| 0.47 ± 0.02e | 0.12 ± 0.02f | 0.69 ± 0.01d | 0.70 ± 0.04d | 0.74 ± 0.03d | 1.49 ± 0.03a | 1.20 ± 0.03c | 1.370.05b | 0.410.02e | |
| 50 | 0.19 ± 0.02c | 0.19 ± 0.03c | 0.42 ± 0.03b | 0.95 ± 0.004a | 0.45 ± 0.01b | 0.13 ± 0.01d | 0.92 ± 0.02a | 0.13 ± 0.01d | 0.15 ± 0.01dc |
| 100 | 1.15 ± 0.05a | 0.77 ± 0.02b | 0.34 ± 0.01d | 0.52 ± 0.04c | 0.25 ± 0.03e | 0.42 ± 0.07d | 0.76 ± 0.03b | 0.34 ± 0.01d | 0.15 ± 0.04f |
| 150 | 0.17 ± 0.01d | 0.52 ± 0.02c | 0.03 ± 0.01d | 0.75 ± 0.09b | 2.24 ± 0.18a | 0.77 ± 0.01b | 0.080.01d | 0.21 ± 0.03d | 0.130.02d |
| 200 | 0.149 ± 0.01a | 1.16 ± 0.02bc | 0.50 ± 0.02e | 0.59 ± 0.03e | 1.16 ± 0.08bc | 0.50 ± 0.02e | 1.25 ± 0.01b | 1.11 ± 0.01c | 0.74 ± 0.01d |
| Tot Chl | |||||||||
| 0 | 1.56 ± 0.02e | 0.83 ± 0.01i | 1.21 ± 0.04f | 3.04 ± 0.02a | 1.04 ± 0.01g | 2.71 ± 0.02b | 1.77 ± 0.03c | 1.71 ± 0.02d | 0.94 ± 0.03h |
| 50 | 0.90 ± 0.01d | 0.77 ± 0.003f | 0.66 ± 0.03f | 2.12 ± 0.01a | 1.05 ± 0.003c | 0.71 ± 0.02e | 2.05 ± 0.01b | 0.91 ± 0.01d | 0.44 ± 0.01g |
| 100 | 2.38 ± 0.02a | 1.31 ± 0.02d | 0.76 ± 0.01f | 1.20 ± 0.03c | 1.18 ± 0.01e | 1.18 ± 0.04e | 2.06 ± 0.04b | 1.99 ± 0.04c | 0.64 ± 0.04g |
| 150 | 0.88 ± 0.01de | 1.25 ± 0.005c | 0.39 ± 0.005f | 1.36 ± 0.09c | 2.86 ± 0.17a | 2.14 ± 0.01b | 1 ± 0.005d | 0.45 ± 0.01f | 0.79 ± 0.01c |
| 200 | 1.86 ± 0.01f | 2.27 ± 0.05b | 0.65 ± 0.02ef | 0.85 ± 0.06d | 1.65 ± 0.06de | 1.37 ± 0.02a | 2.21 ± 0.01c | 2.14 ± 0.01b | 1.22 ± 0.01c |
| Species/Populations | Code /NGBT* | Province | Alt (m) | AAR*** (mm) | Tmax (°C) | Tmin (°C) | CE (mM/cm) |
|---|---|---|---|---|---|---|---|
| Medicago ciliaris | |||||||
| Pop1 | 9151 | Raoued El Hessiane | 5.75 | 398.5 | 35.1 | 5.4 | 0.39 |
| Pop2 | 9152 | Kalaat El Andalous | 5.85 | 398.5 | 35.1 | 5.4 | 0.41 |
| Pop3 | 9153 | Ghar El Melh | 6.19 | 497.8 | 32.9 | 6.7 | 0.16 |
| 306 | 9141 | Siliana | 575 | 341.1 | 38.6 | 1.6 | 0.27 |
| 355 | 9142 | Dougga | 320 | 493.5 | 21.7 | 9.9 | 0.24 |
| 667 | 9143 | Zaghouan | 300 | 496 | 32.6 | 6.7 | 2.41 |
| 773 | 9144 | Mateur 1 | 5 | 527.1 | 33.4 | 4.9 | 0.74 |
| Medicago scutellata | Wafra* | Saîda/Manouba | 114 | 450 | 45 | 6 | - |
| Medicago intertexta** | - | SidiAissa/Nabeul | 120 | 358 | 36 | 8.4 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mbarki, S.; Skalicky, M.; Vachova, P.; Hajihashemi, S.; Jouini, L.; Zivcak, M.; Tlustos, P.; Brestic, M.; Hejnak, V.; Zoghlami Khelil, A. Comparing Salt Tolerance at Seedling and Germination Stages in Local Populations of Medicago ciliaris L. to Medicago intertexta L. and Medicago scutellata L. Plants 2020, 9, 526. https://doi.org/10.3390/plants9040526
Mbarki S, Skalicky M, Vachova P, Hajihashemi S, Jouini L, Zivcak M, Tlustos P, Brestic M, Hejnak V, Zoghlami Khelil A. Comparing Salt Tolerance at Seedling and Germination Stages in Local Populations of Medicago ciliaris L. to Medicago intertexta L. and Medicago scutellata L. Plants. 2020; 9(4):526. https://doi.org/10.3390/plants9040526
Chicago/Turabian StyleMbarki, Sonia, Milan Skalicky, Pavla Vachova, Shokoofeh Hajihashemi, Latifa Jouini, Marek Zivcak, Pavel Tlustos, Marian Brestic, Vaclav Hejnak, and Aziza Zoghlami Khelil. 2020. "Comparing Salt Tolerance at Seedling and Germination Stages in Local Populations of Medicago ciliaris L. to Medicago intertexta L. and Medicago scutellata L." Plants 9, no. 4: 526. https://doi.org/10.3390/plants9040526
APA StyleMbarki, S., Skalicky, M., Vachova, P., Hajihashemi, S., Jouini, L., Zivcak, M., Tlustos, P., Brestic, M., Hejnak, V., & Zoghlami Khelil, A. (2020). Comparing Salt Tolerance at Seedling and Germination Stages in Local Populations of Medicago ciliaris L. to Medicago intertexta L. and Medicago scutellata L. Plants, 9(4), 526. https://doi.org/10.3390/plants9040526