Freezing Tolerance and Expression of β-amylase Gene in Two Actinidia arguta Cultivars with Seasonal Changes

Abstract
1. Introduction
2. Results
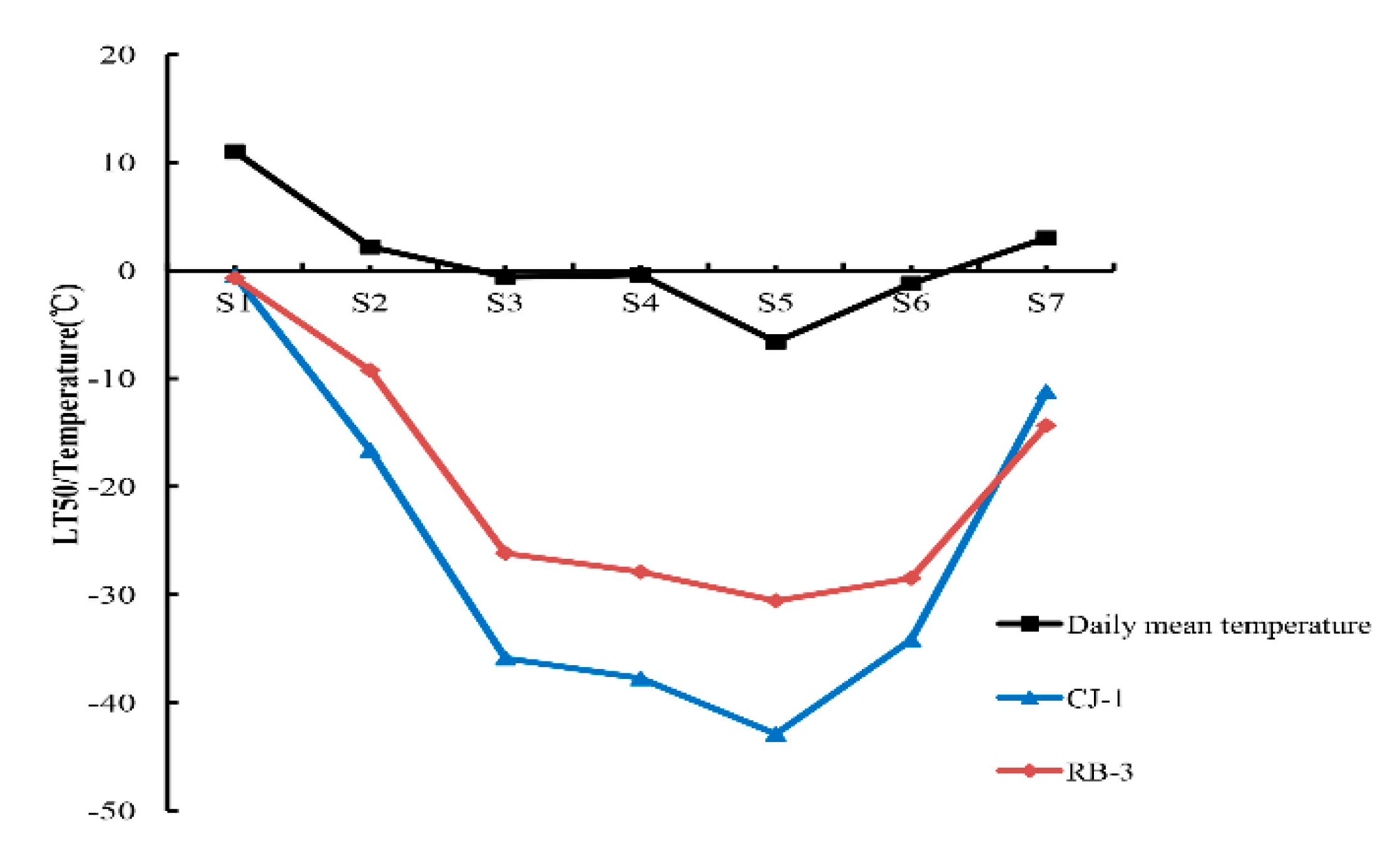
2.1. Seasonal Changes in Air Temperature
2.2. Freezing Tolerance in Two Actinidia Arguta
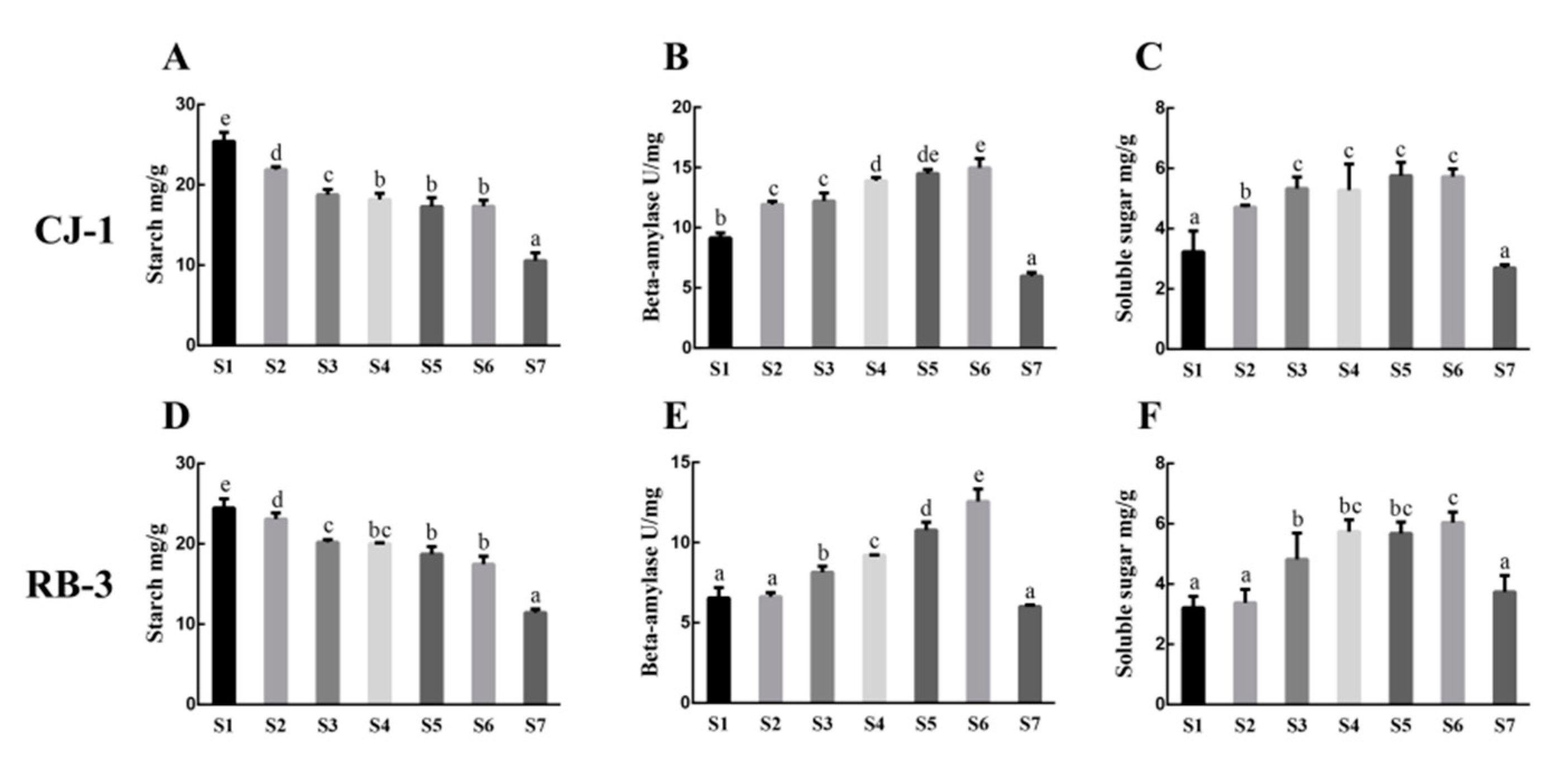
2.3. Metabolite Contents
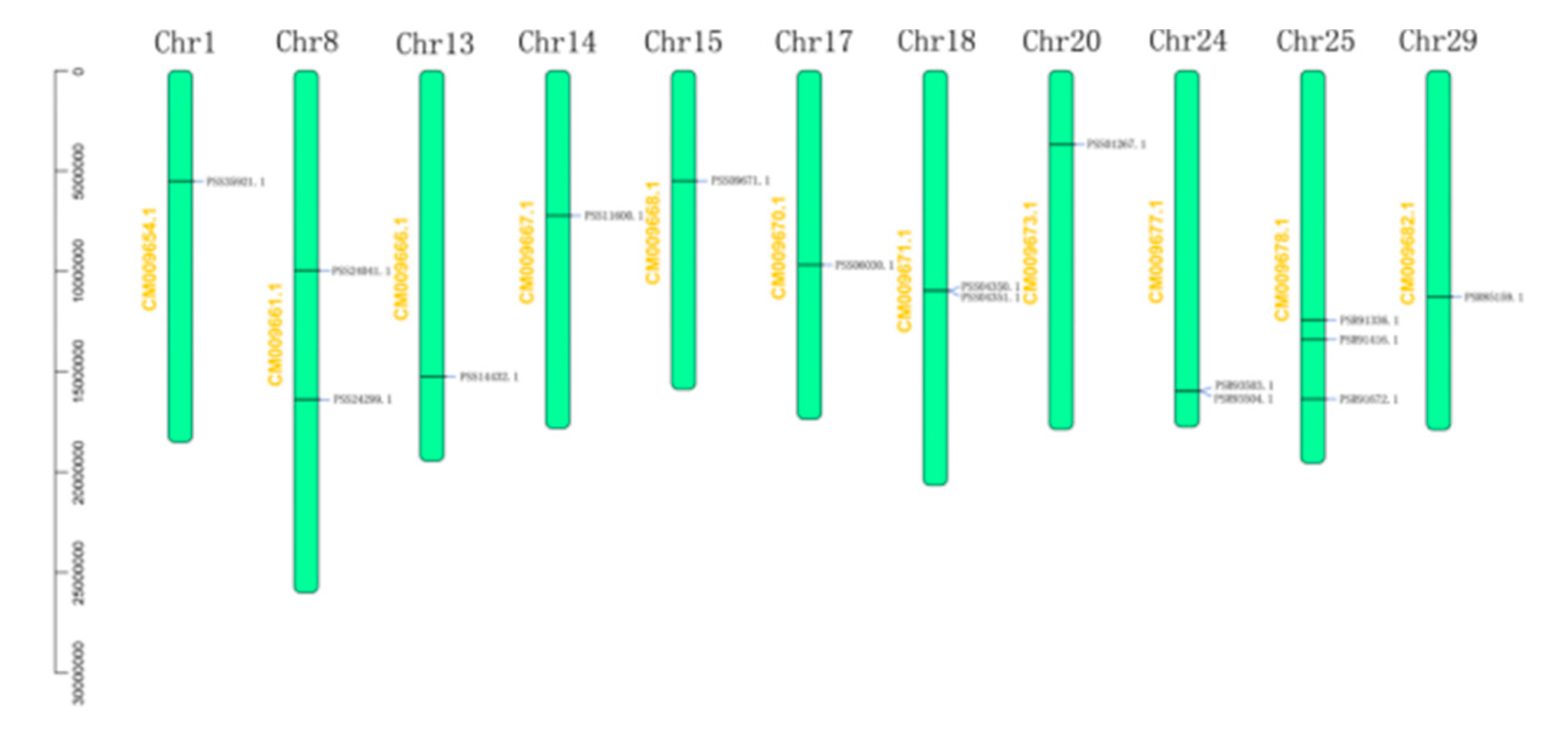
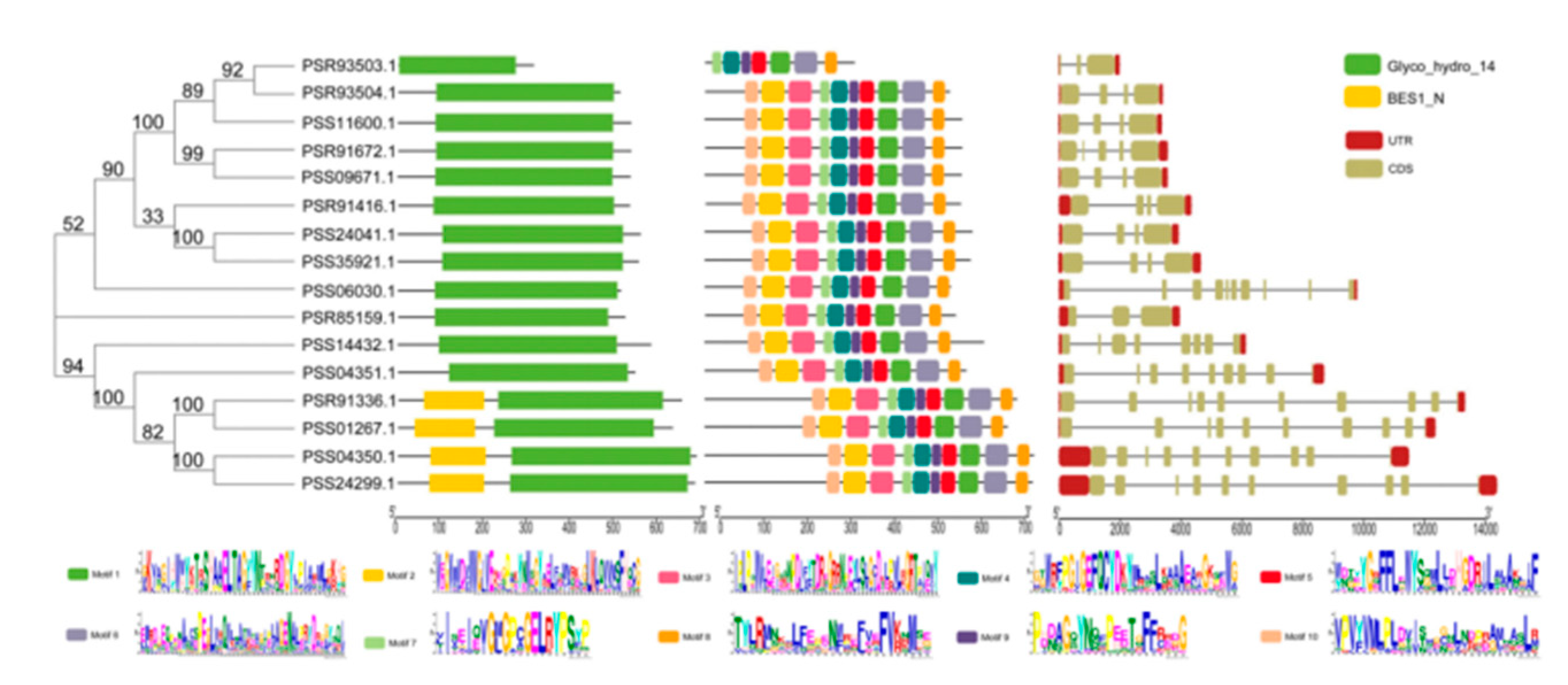
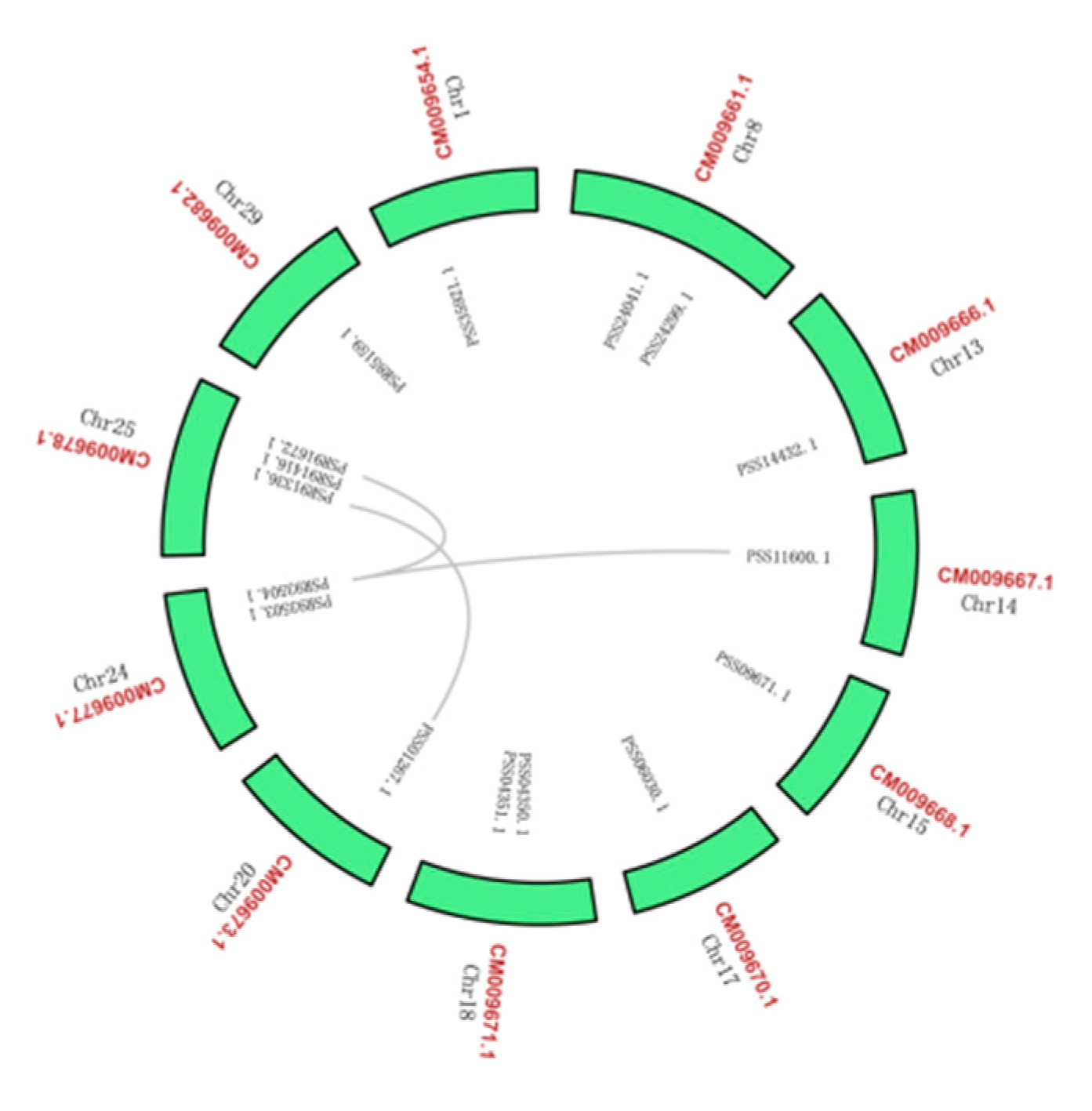
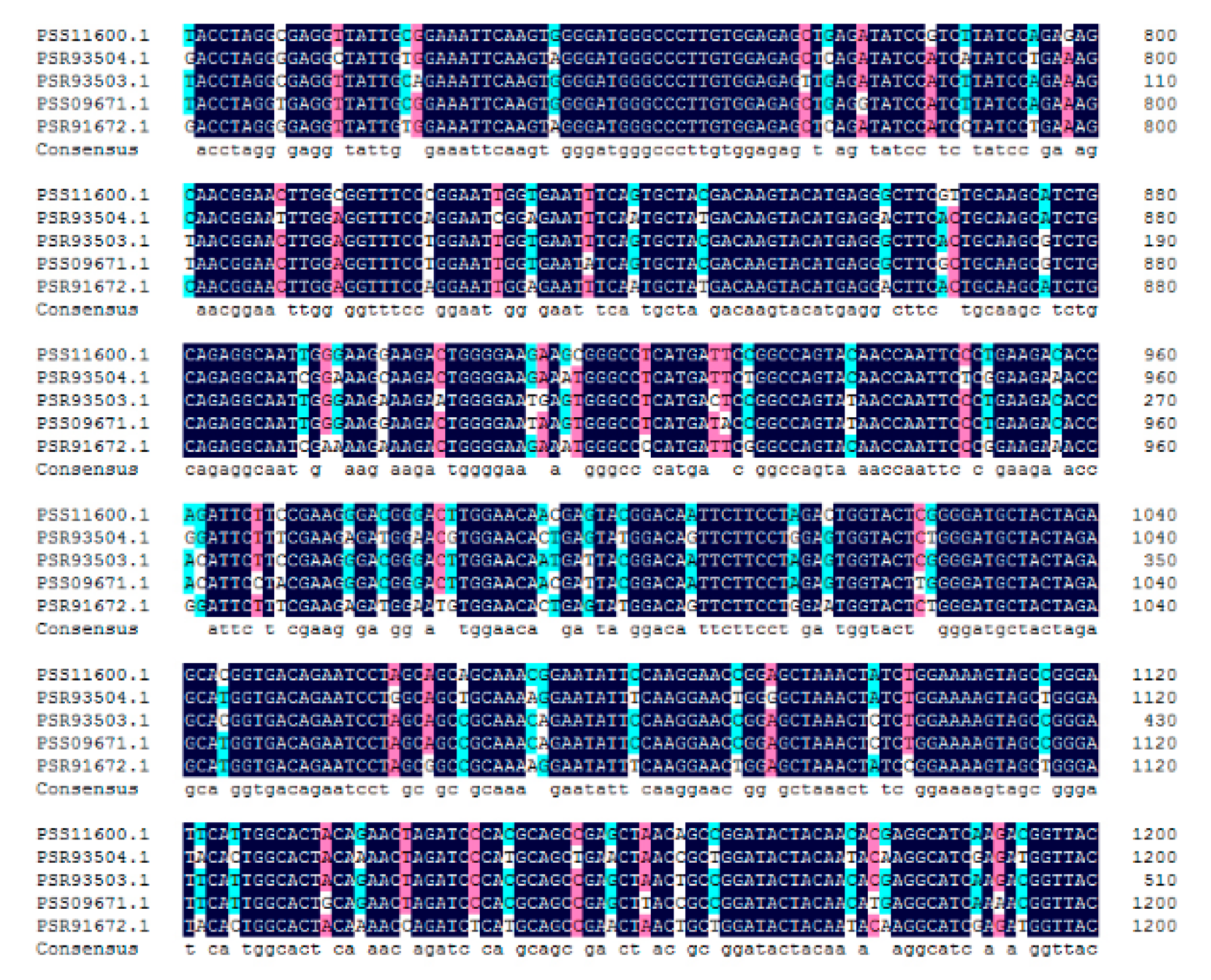
2.4. Bioinformatic Analysis of AcBAM
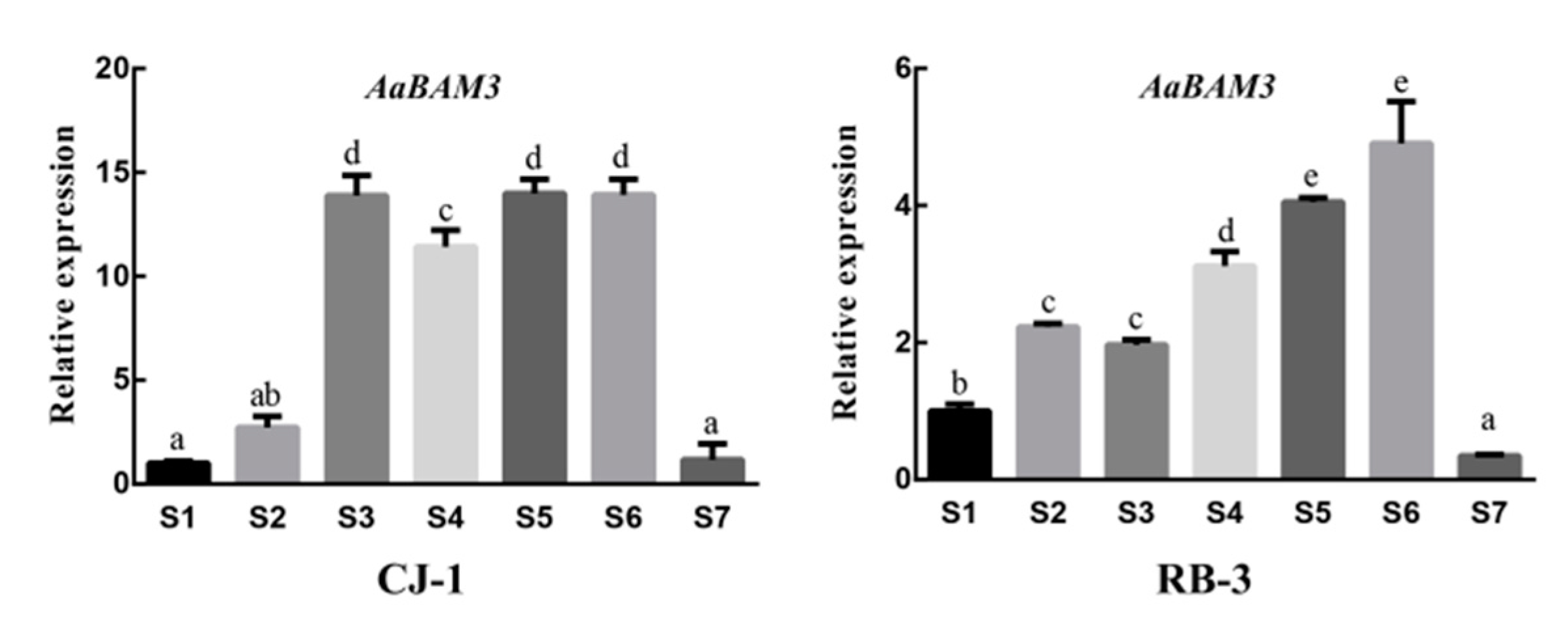
2.5. Expression Analysis of BAM
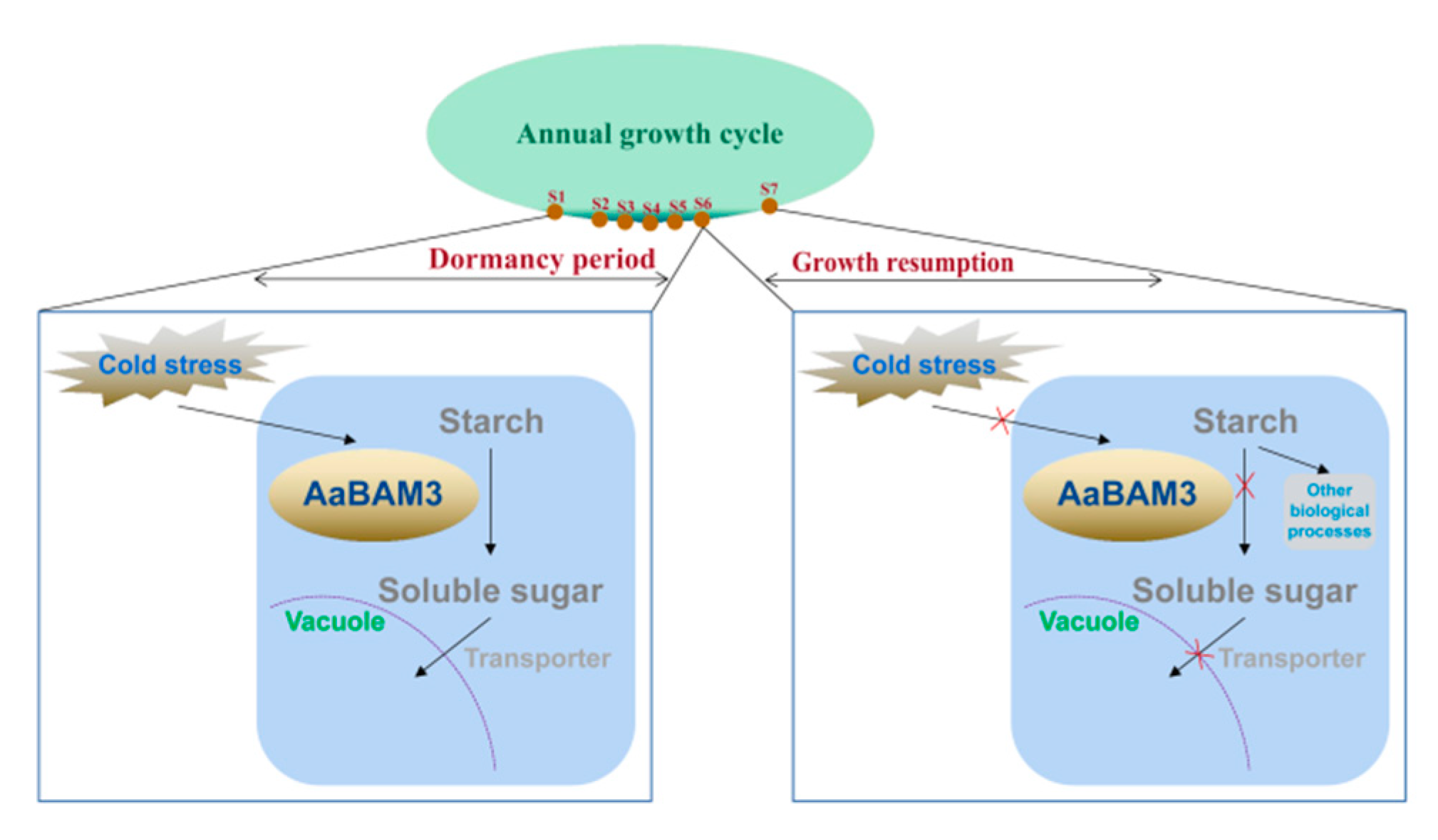
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Conditions
4.2. Determinations of Freezing Tolerance
4.3. The Measurement of Carbohydrate Metabolites Contents
4.4. RNA Extraction and Reverse Transcription
4.5. Analysis of AcBAM Gene Family
4.6. Isolation and Expression of Genes
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sun, X.; Zhenfei, Z.; Langlang, Z.; Linchuan, F.; Jisen, Z.; Qingfeng, W.; Shaohua, L.; Zhenchang, L.; Haiping, X. Overexpression of ethylene response factors VaERF080 and VaERF087 from Vitis amurensis enhances cold tolerance in Arabidopsis. Sci. Hortic. 2019, 243, 320–326. [Google Scholar] [CrossRef]
- An, J.P.; Li, R.; Qu, F.J.; You, C.X.; Wang, X.F.; Hao, Y.J. An apple NAC transcription factor negatively regulates cold tolerance via CBF-dependent pathway. J. Plant Physiol. 2018, 221, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Lin, O.; Leus, L.; Keyser, E.D.; Labeke, M.C.V. Seasonal changes in cold hardiness and carbohydrate metabolism in four garden rose cultivars. J. Plant Physiol. 2018, 232, 188–199. [Google Scholar]
- Pietsch, G.M.; Anderson, N.O.; Li, P.H. Cold tolerance and short day acclimation in perennial Gaura coccinea and G. drummondii. Sci. Hortic. 2009, 120, 418–425. [Google Scholar] [CrossRef]
- Horrer, D.; Flütsch, S.; Pazmino, D.; Matthews, J.A.; Thalmann, M.; Nigro, A.; Leonhardt, N.; Lawson, T.; Santelia, D. Blue Light Induces a Distinct Starch Degradation Pathway in Guard Cells for Stomatal Opening. Curr. Biol. 2016, 26, 362–370. [Google Scholar] [CrossRef]
- Lee, B.H.; Hojoung, L.; Liming, X.; JianKang, Z. A mitochondrial complex I defect impairs cold-regulated nuclear gene expression. Plant Cell 2002, 14, 1235–1251. [Google Scholar] [CrossRef]
- Zanella, M.; Borghi, G.L.; Pirone, C.; Thalmann, M.; Pazmino, D.; Costa, A.; Santelia, D.; Trost, P.; Sparla, F. β-amylase 1 (BAM1) degrades transitory starch to sustain proline biosynthesis during drought stress. J. Exp. Bot. 2016, 67, 1819–1826. [Google Scholar] [CrossRef]
- Zhao, L.; Gong, X.; Gao, J.; Dong, H.; Zhang, S.; Tao, S.; Huang, X. Transcriptomic and evolutionary analyses of white pear (Pyrus bretschneideri) β-amylase genes reveals their importance for cold and drought stress responses. Gene 2019, 689, 102–113. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef]
- Dong, S.; Beckles, D.M. Dynamic changes in the starch-sugar interconversion within plant source and sink tissues promote a better abiotic stress response. J. Plant Physiol. 2019, 234, 80–93. [Google Scholar] [CrossRef]
- Yue, C.; Cao, H.L.; Wang, L.; Zhou, Y.H.; Huang, Y.T.; Hao, X.Y.; Wang, Y.C.; Wang, B.; Yang, Y.J.; Wang, X.C. Effects of cold acclimation on sugar metabolism and sugar-related gene expression in tea plant during the winter season. Plant Mol. Biol. 2015, 88, 591–608. [Google Scholar] [CrossRef] [PubMed]
- Fulton, D.C.; Stettler, M.; Mettler, T.; Vaughan, C.K.; Li, J.; Francisco, P.; Gil, M.; Reinhold, H.; Eicke, S.; Messerli, G.; et al. β-AMYLASE4, a Noncatalytic Protein Required for Starch Breakdown, Acts Upstream of Three Active β-Amylases in Arabidopsis Chloroplasts. Plant Cell 2008, 20, 1040–1058. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Zhu, X.; Duan, N.; Liu, J.H. PtrBAM1, a beta-amylase-coding gene of Poncirus trifoliata, is a CBF regulon member with function in cold tolerance by modulating soluble sugar levels. Plant Cell Environ. 2014, 37, 2754–2767. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Yang, T.; Xing, C.; Dong, H.; Qi, K.; Gao, J.; Tao, S.; Wu, J.; Wu, J.; Zhang, S.; et al. The β-amylase PbrBAM3 from pear (Pyrus betulaefolia) regulates soluble sugar accumulation and ROS homeostasis in response to cold stress. Plant Sci. 2019, 287, 110184. [Google Scholar] [CrossRef]
- Huang, H.W. Huang History of 100 Years of Domestication and Improvement of Kiwifruit and Gene Discovery from Genetic Introgressed Populations in the Wild. Chin. Bull. Bot. 2009, 44, 127–142. [Google Scholar]
- Jo, Y.S.; Cho, H.S.; Park, J.O.; Kim, T.C.; Kim, W.S. Assay of cold tolerance of Actinidia eriantha. Acta Hortic. 2006, 773, 277–282. [Google Scholar]
- Kovi, M.R.; Ergon, A.; Rognli, O.A. Freezing tolerance revisited—Effects of variable temperatures on gene regulation in temperate grasses and legumes. Curr. Opin. Plant Biol. 2016, 33, 140–146. [Google Scholar] [CrossRef]
- Hyung, L.J.; Yu, D.J.; Jin, K.S.; Doil, C.; Jae, L.H. Intraspecies differences in cold hardiness, carbohydrate content and β-amylase gene expression of Vaccinium corymbosum during cold acclimation and deacclimation. Tree Physiol. 2012, 32, 1533–1540. [Google Scholar]
- Marvin, J.W.; Morselli, M. Rapid low temperature hydrolysis of starch to sugars in maple stems and in maple tissue cultures. Cryobiology 1971, 8, 339–344. [Google Scholar] [CrossRef]
- Eris, A.; Gulen, H.; Barut, E.; Cansev, A. Annual patterns of total soluble sugars and proteins related to coldhardiness in olive. J. Hortic. Sci. Biotechnol. 2007, 82, 597–604. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R. Understanding the cellular mechanism of recovery from freeze-thaw injury in spinach, possible role of aquaporins, heat shock proteins, dehydrin and antioxidant system. Physiol. Plant. 2014, 150, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Xu, S.; Wang, Y.; Zhang, E.; Xu, C. Molecular evolution and expression patterns under abiotic stresses of beta-amylase gene family in grasses. Sci. Technol. Rev. 2014, 32, 29–36. [Google Scholar]
- Dong, C.; Jia, L.; Deng, J.; Zhao, Q.; Chen, H.; Hu, H.G. Phylogenetic Analysis ofα-amylase Gene Family in Musa accuminata. South China Fruits 2016, 45, 12–17. [Google Scholar]
- Monroe, J.D.; Storm, A.R. The Arabidopsis beta-amylase (BAM) gene family, Diversity of form and function. Plant Sci. 2018, 276, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Fatma, K.; Guy, C.L. beta-Amylase induction and the protective role of maltose during temperature shock. Plant Physiol. 2004, 135, 1674–1684. [Google Scholar]
- Sun, S. Establishment and Application of Evaluation Method of Freezing Tolerance in Actinidia; Chinese Academy of Agricultural Sciences: Beijing, China, 2018. [Google Scholar]
- Murray, M.B.; Cape, J.N.; Fowler, D. Quantification of frost damage in plants by rate of electrolyte leakage. New Phytol. 2010, 113, 307–311. [Google Scholar] [CrossRef]
- He, W.; Ai, J.; Fan, S.; Yang, Y.; Wang, Z.; Zhao, Y.; Qiao, Y.; Zhang, Y.; Li, X.Y. Study on evaluation method for cold resistance of grape cultivars and rootstock. J. Fruit Sci. 2015, 32, 1135–1142. [Google Scholar]
- Pucher, G.W.; Vickery, H.B. Determination of Starch in Plant Tissues. Ind. Eng. Chem. Anal. Ed. 1936, 8, 92–97. [Google Scholar] [CrossRef]
- Guo, Y.H.; Yang, W.H.; Yu, Y.Z.; Xu, H.X.; Wang, Y.Q.; Zhou, D.Y.; Kuang, M. Determination of Total Sugar Contents in Mature Cotton Fiber Using Anthrone Colorimetry. China Cotton 2011, 10, 23–26. [Google Scholar]
- Pilkington, S.M.; Crowhurst, R.; Schaffer, R.J. A manually annotated Actinidia chinensis var. chinensis (kiwifruit) genome highlights the challenges associated with draft genomes and gene prediction in plants. BMC Genom. 2018, 257, 257. [Google Scholar] [CrossRef]
- Li, Y.; Fang, J.; Qi, X.; Lin, M.; Zhong, Y.; Sun, L. A key structural gene, AaLDOX, is involved in anthocyanin biosynthesis in all red-fleshed kiwifruit (Actinidia arguta) based on transcriptome analysis. Gene 2018, 648, 31–41. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | 23 October | 22 November | 7 December | 22 December | 5 January | 20 January | 5 March |
|---|---|---|---|---|---|---|---|
| Phase | S1 | S2 | S3 | S4 | S5 | S6 | S7 |
| Accession | Location | pI | Molecular Weight (Da) | Size (aa) |
|---|---|---|---|---|
| PSR85159.1 | Chr29: 11230497–11233893 | 6.71 | 58895.73 | 532 |
| PSR91336.1 | Chr25: 12382772–12394190 | 5.76 | 74551.84 | 664 |
| PSR91416.1 | Chr25: 13363180–13366908 | 6.8 | 61047.39 | 542 |
| PSR91672.1 | Chr25: 16324193–16327258 | 7.22 | 61707.94 | 546 |
| PSR93503.1 | Chr24: 15922030–15923754 | 8.52 | 35748.08 | 317 |
| PSR93504.1 | Chr24: 15932333–15935251 | 8.72 | 59006.06 | 520 |
| PSS01267.1 | Chr20: 3639423–3650017 | 6.02 | 72661.93 | 644 |
| PSS04350.1 | Chr18: 10923906–10933753 | 5.24 | 78372.16 | 700 |
| PSS04351.1 | Chr18: 10935330–10942790 | 5.46 | 63106.07 | 556 |
| PSS06030.1 | Chr17: 9640160–9648546 | 8.83 | 59864.53 | 523 |
| PSS09671.1 | Chr15: 5454601–5457662 | 7.56 | 61368.5 | 546 |
| PSS11600.1 | Chr14: 7186173–7189084 | 8.76 | 61812.1 | 547 |
| PSS14432.1 | Chr13: 15225015–15230273 | 5.51 | 66860.73 | 594 |
| PSS24041.1 | Chr8: 9912429–9915793 | 5.78 | 63364.85 | 569 |
| PSS24299.1 | Chr8:16339786–16352100 | 5.6 | 77792.48 | 697 |
| PSS35921.1 | Chr1: 5464816–5468823 | 6.06 | 62737.09 | 565 |
| Primer Name | Sequence | Type |
|---|---|---|
| AaBAM3-F | 5′-ATGAATGCCAGCTTGATGGC-3′ | PCR |
| AaBAM3-R | 5′-TTACACTAGAGCAGCCTCCTTCG-3′ | |
| qBAM-F | 5′-ACCTACCAATAGCGCGGATG-3′ | RT-qPCR |
| qBAM-R | 5′-AACTAACCCTTCTGGCGAGC-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.; Fang, J.; Lin, M.; Qi, X.; Chen, J.; Wang, R.; Li, Z.; Li, Y.; Muhammad, A. Freezing Tolerance and Expression of β-amylase Gene in Two Actinidia arguta Cultivars with Seasonal Changes. Plants 2020, 9, 515. https://doi.org/10.3390/plants9040515
Sun S, Fang J, Lin M, Qi X, Chen J, Wang R, Li Z, Li Y, Muhammad A. Freezing Tolerance and Expression of β-amylase Gene in Two Actinidia arguta Cultivars with Seasonal Changes. Plants. 2020; 9(4):515. https://doi.org/10.3390/plants9040515
Chicago/Turabian StyleSun, Shihang, Jinbao Fang, Miaomiao Lin, Xiujuan Qi, Jinyong Chen, Ran Wang, Zhi Li, Yukuo Li, and Abid Muhammad. 2020. "Freezing Tolerance and Expression of β-amylase Gene in Two Actinidia arguta Cultivars with Seasonal Changes" Plants 9, no. 4: 515. https://doi.org/10.3390/plants9040515
APA StyleSun, S., Fang, J., Lin, M., Qi, X., Chen, J., Wang, R., Li, Z., Li, Y., & Muhammad, A. (2020). Freezing Tolerance and Expression of β-amylase Gene in Two Actinidia arguta Cultivars with Seasonal Changes. Plants, 9(4), 515. https://doi.org/10.3390/plants9040515