Cellular and Subcellular Compartmentation of the 2C-Methyl-D-Erythritol 4-Phosphate Pathway in the Madagascar Periwinkle

 ,
,  , ,
, ,  , ,
, ,  , , and
, , and 
Abstract
1. Introduction
2. Results
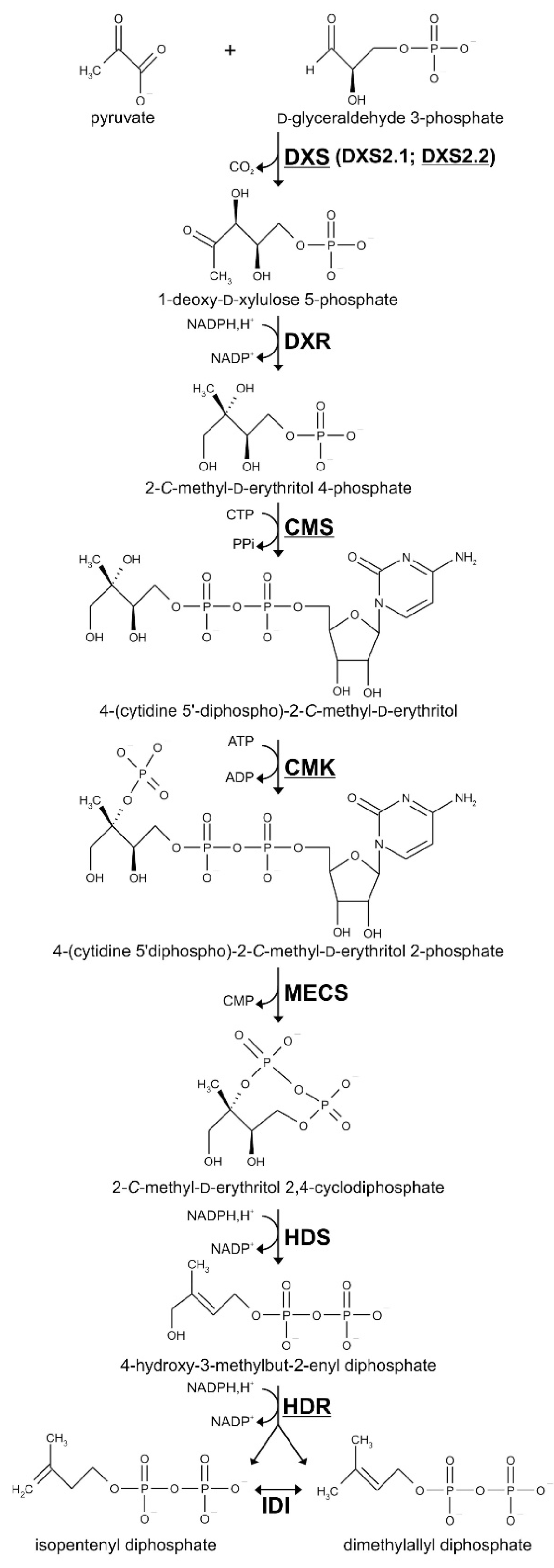
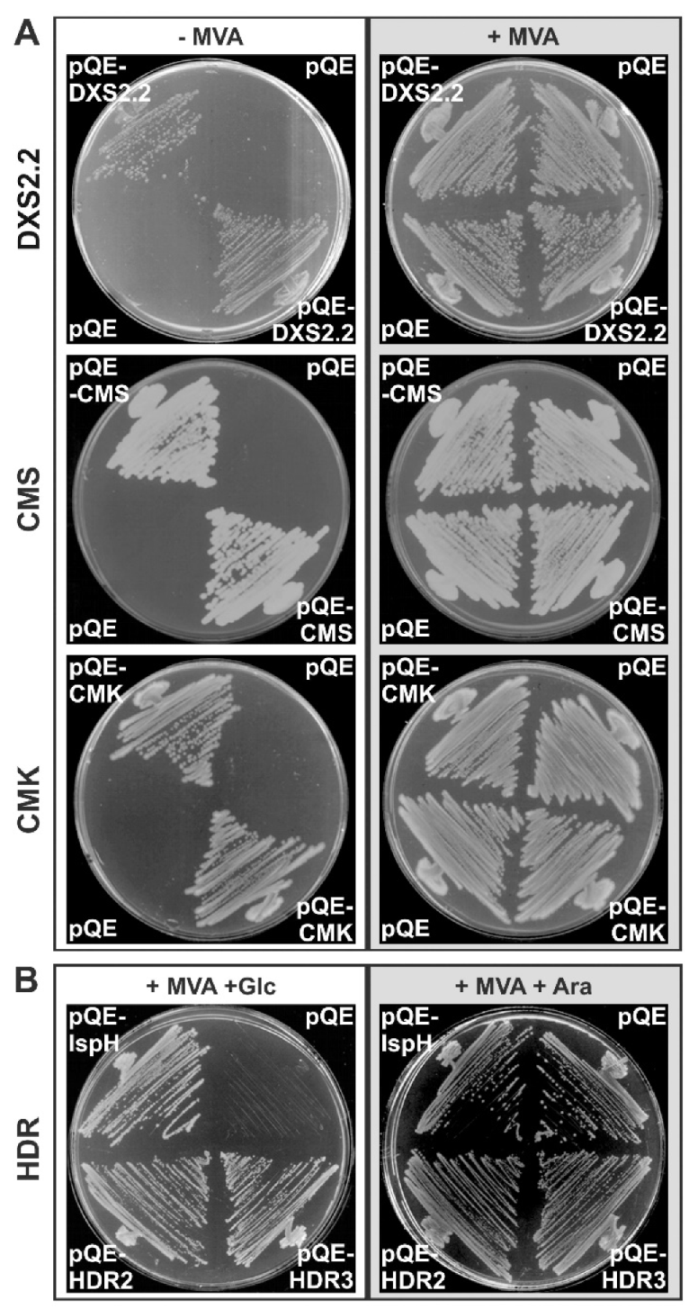
2.1. Cloning and Functional Validation of cDNAs Encoding a Second DXS Isoform, CMS, CMK and HDR
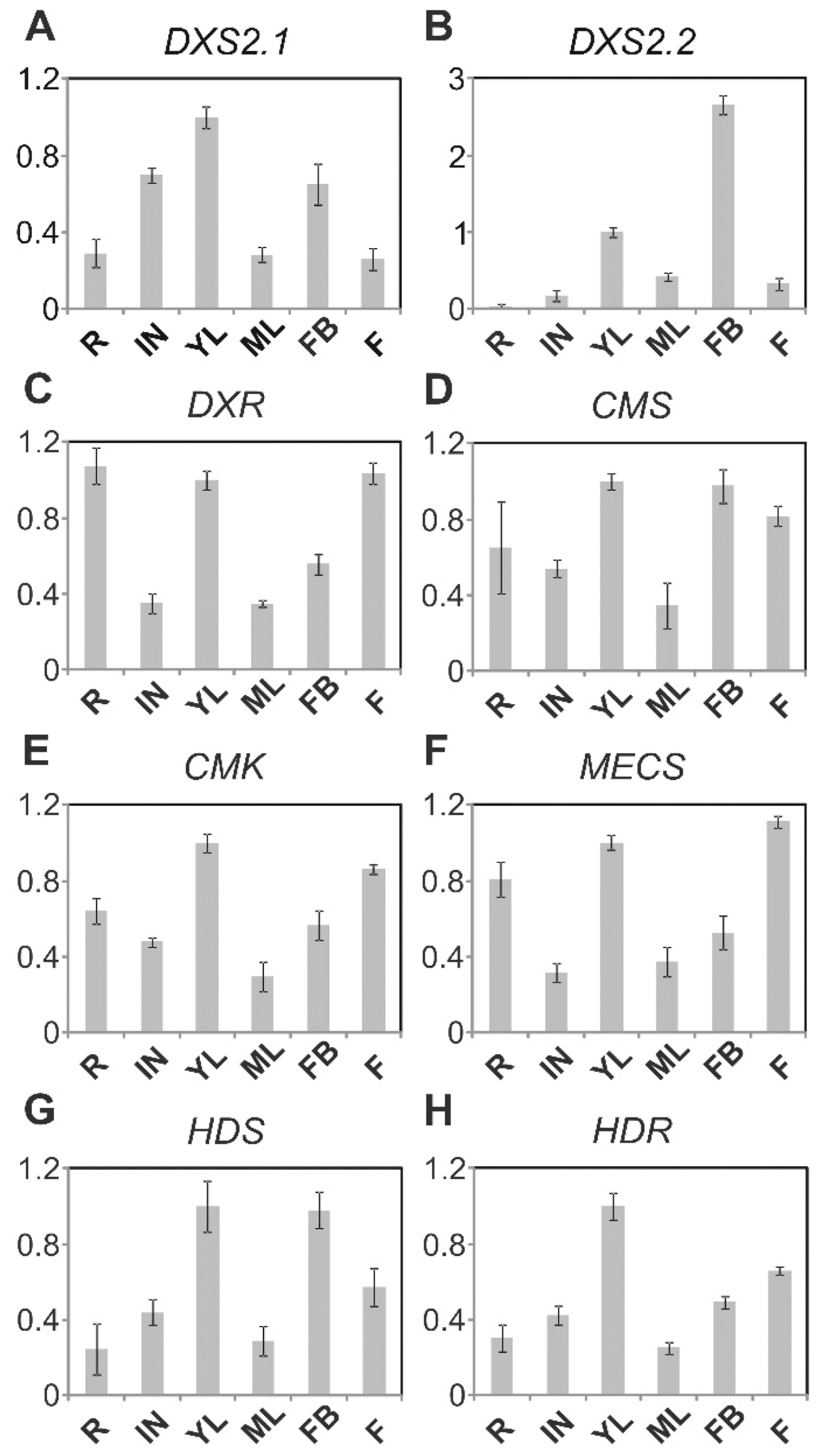
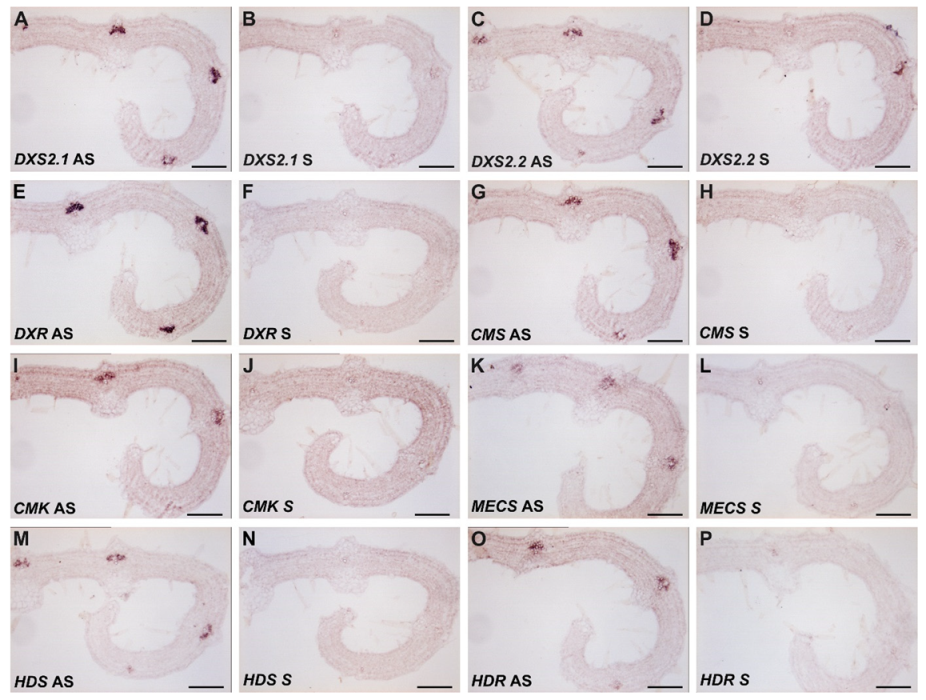
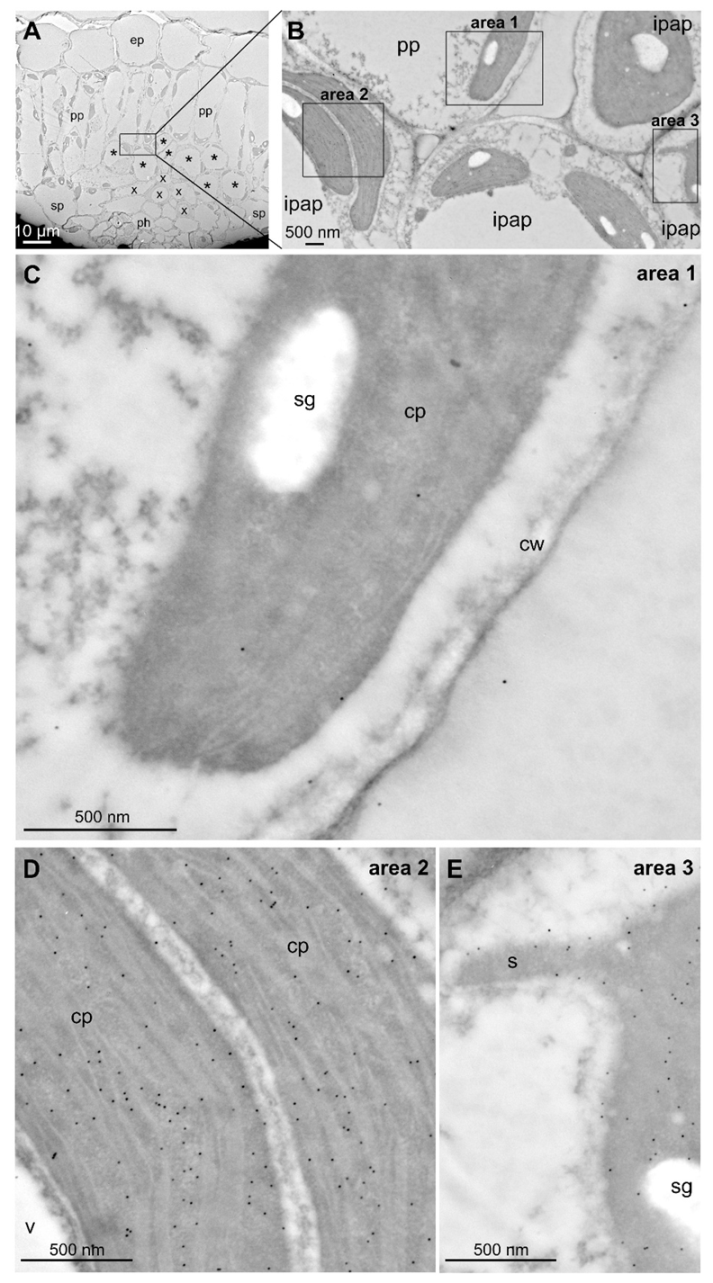
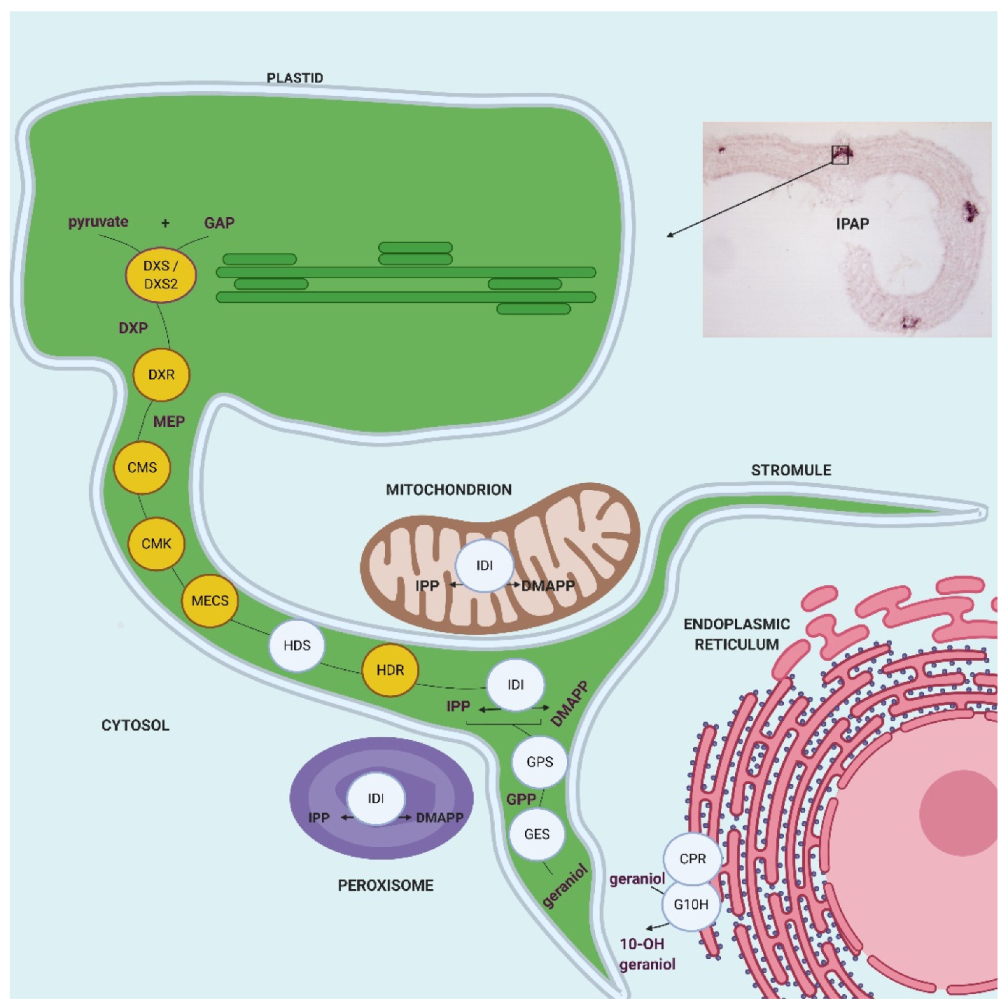
2.2. MEP Pathway Genes Are Highly Coexpressed in IPAP Cells of Young Leaves
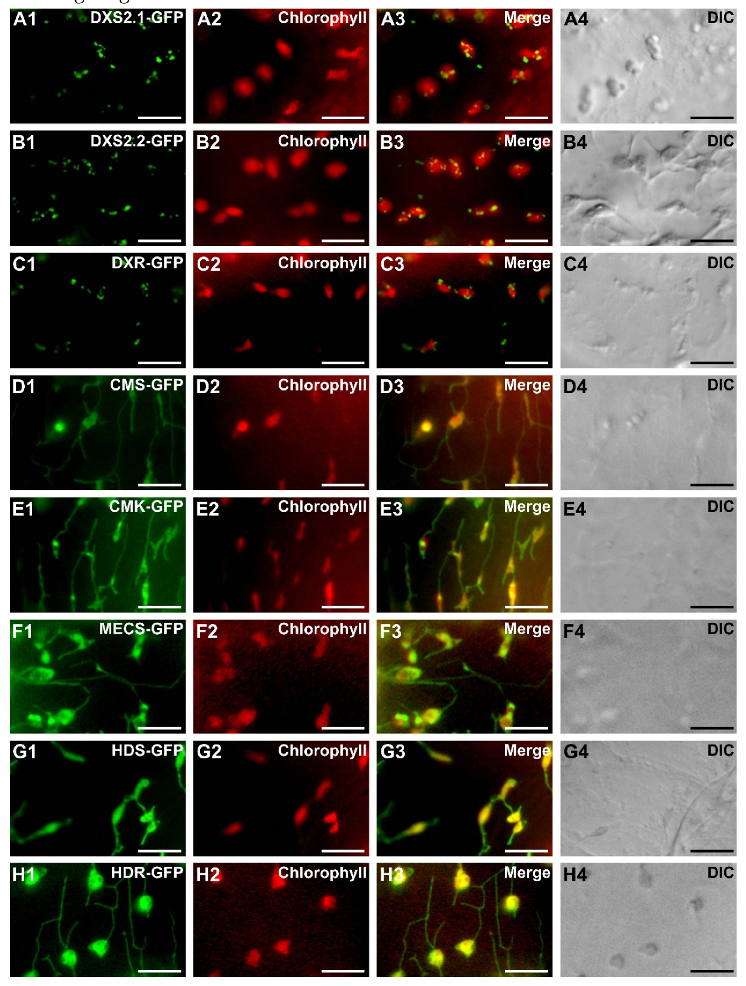
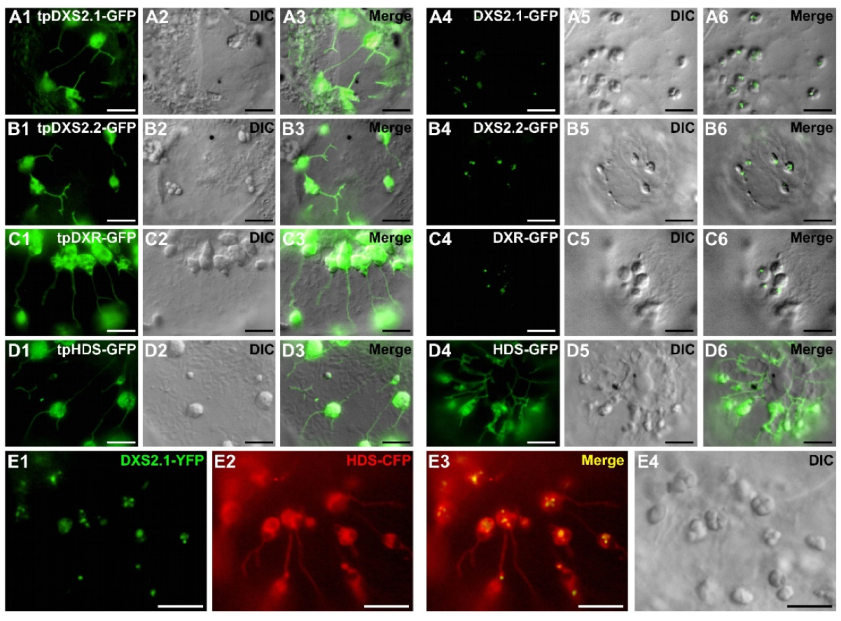
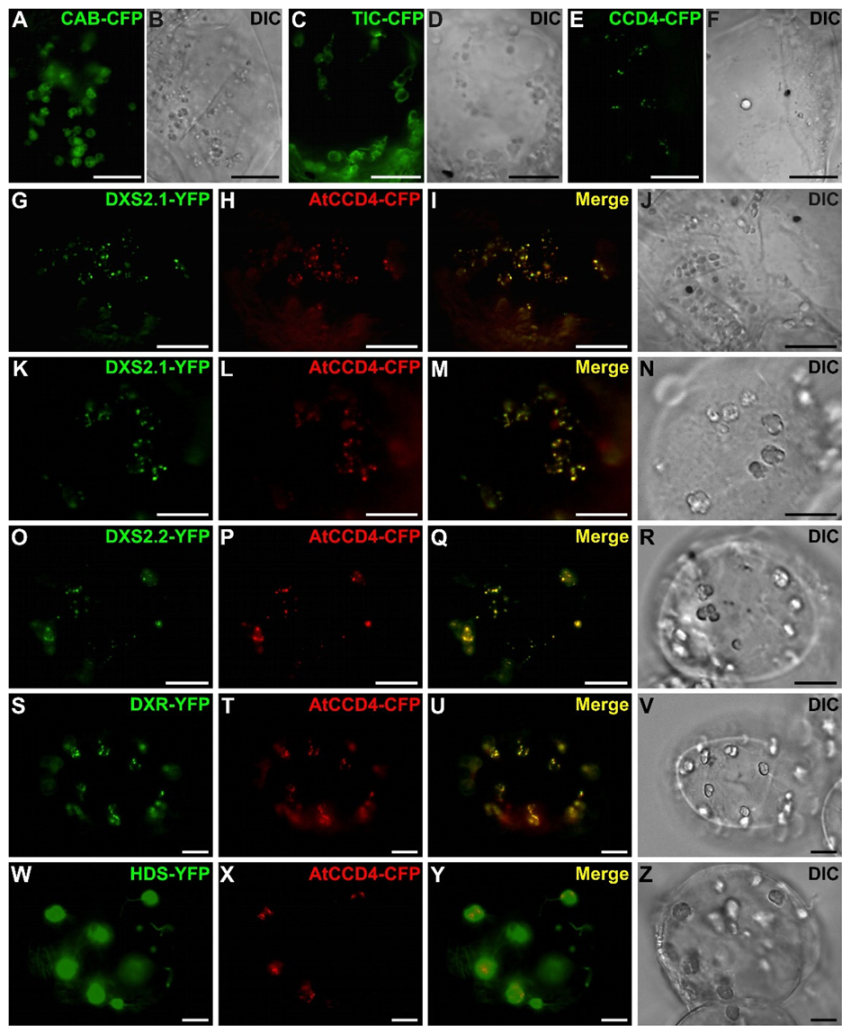
2.3. MEP Pathway Enzymes Are Targeted to Plastids
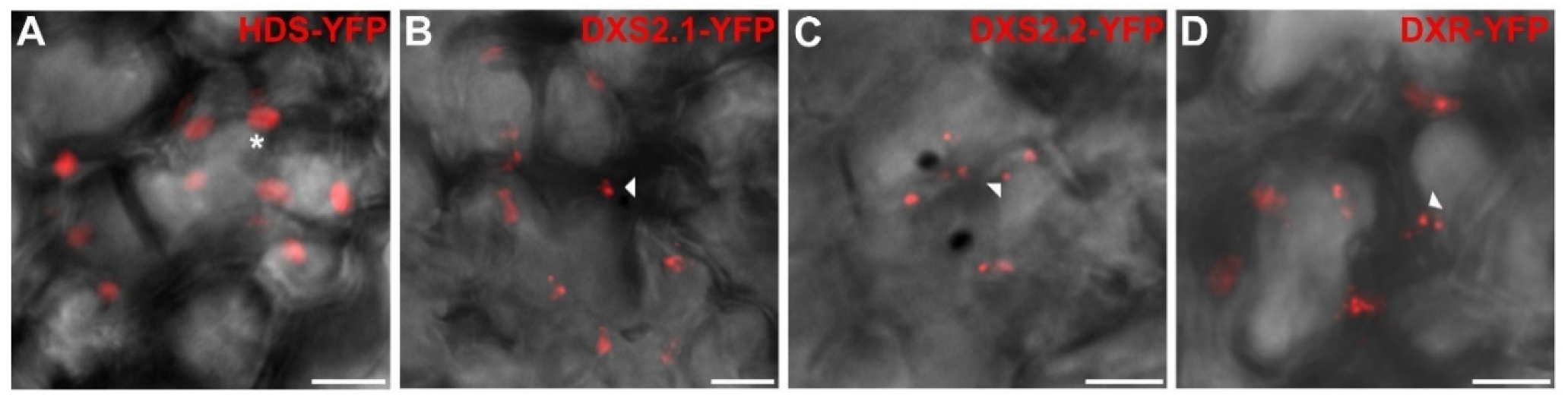
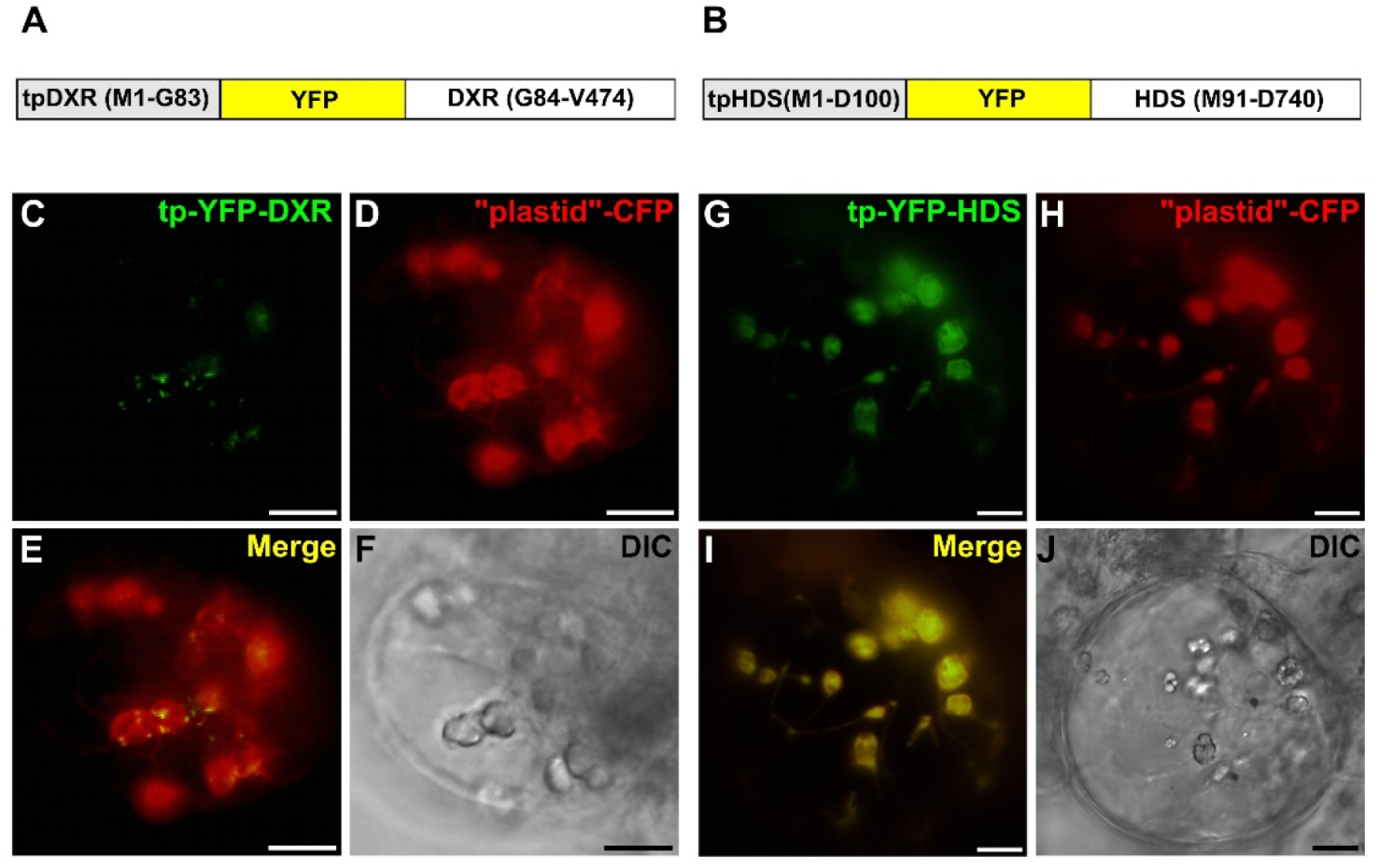
2.4. Punctuated Pattern of Localization of DXS and DXR Requires the Whole Protein Sequence
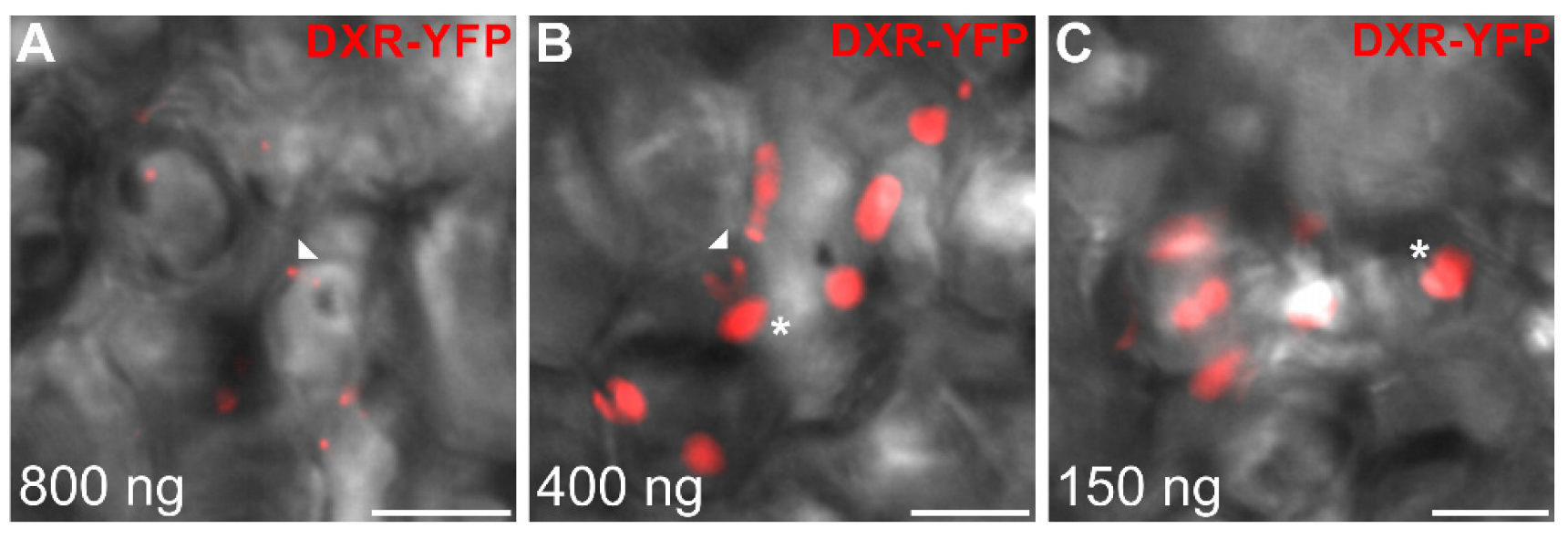
2.5. Punctuated Localization of DXS and DXR is Likely Caused by Protein Overexpression
3. Discussion
4. Material and Methods
4.1. Plant Material and Cell Culture Conditions
4.2. RNA Extraction and cDNA Synthesis
4.3. Cloning of the cDNA Sequences from the C. Roseus MEP Pathway Enzymes
4.4. Protein Production and E. coli Complementation
4.5. Gene Expression Measurements (Real-Time PCR Analysis)
4.6. Tissue Fixation, Embedding in Paraffin and Microtomy
4.7. RNA In Situ Hybridization
4.8. Bioinformatic Sequence Analysis
4.9. Coexpression Network Analysis
4.10. FP-Fusion Constructs for MEP Pathway Enzymes Localization Studies
4.11. Organelle Markers
4.12. Cell Transformations and Epifluorescence Microscopy
4.13. Transmission Electron Microscopy Immunogold Labeling
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bouvier, F.; Rahier, A.; Camara, B. Biogenesis, molecular regulation and function of plant isoprenoids. Prog. Lipid Res. 2005, 44, 357–429. [Google Scholar] [CrossRef] [PubMed]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- Guirimand, G.; Courdavault, V.; St-Pierre, B.; Burlat, V. Biosynthesis and Regulation of Alkaloids. In Plant Developmental Biology-Biotechnological Perspectives: Volume 2; Pua, E.C., Davey, M.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 139–160. ISBN 9783642046704. [Google Scholar]
- O’Connor, S.E.; Maresh, J.J. Chemistry and biology of monoterpene indole alkaloid biosynthesis. Nat. Prod. Rep. 2006, 23, 532–547. [Google Scholar] [CrossRef] [PubMed]
- Dugé De Bernonville, T.; Carqueijeiro, I.; Lanoue, A.; Lafontaine, F.; Sánchez Bel, P.; Liesecke, F.; Musset, K.; Oudin, A.; Glévarec, G.; Pichon, O.; et al. Folivory elicits a strong defense reaction in Catharanthus roseus: Metabolomic and transcriptomic analyses reveal distinct local and systemic responses. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Guirimand, G.; Courdavault, V.; Lanoue, A.; Mahroug, S.; Guihur, A.; Blanc, N.; Giglioli-Guivarc’h, N.; St-Pierre, B.; Burlat, V. Strictosidine activation in Apocynaceae: Towards a “nuclear time bomb”? BMC Plant Biol. 2010, 10, 182. [Google Scholar] [CrossRef]
- Luijendijk, T.J.C.; Van Der Meijden, E.D.; Verpoorte, R. Involvement of strictosidine as a defensive chemical in Catharanthus roseus. J. Chem. Ecol. 1996, 22, 1355–1366. [Google Scholar] [CrossRef]
- Facchini, P.J.; De Luca, V. Opium poppy and Madagascar periwinkle: Model non-model systems to investigate alkaloid biosynthesis in plants. Plant J. 2008, 54, 763–784. [Google Scholar] [CrossRef]
- Kellner, F.; Geu-Flores, F.; Sherden, N.H.; Brown, S.; Foureau, E.; Courdavault, V.; O’Connor, S.E. Discovery of a P450-catalyzed step in vindoline biosynthesis: A link between the aspidosperma and eburnamine alkaloids. Chem. Commun. 2015, 51, 7626–7628. [Google Scholar] [CrossRef]
- Qu, Y.; Easson, M.L.A.E.; Froese, J.; Simionescu, R.; Hudlicky, T.; DeLuca, V. Completion of the seven-step pathway from tabersonine to the anticancer drug precursor vindoline and its assembly in yeast. Proc. Natl. Acad. Sci. USA 2015, 112, 6224–6229. [Google Scholar] [CrossRef]
- Tatsis, E.C.; Carqueijeiro, I.; Dugé de Bernonville, T.; Franke, J.; Dang, T.-T.T.; Oudin, A.; Lanoue, A.; Lafontaine, F.; Stavrinides, A.K.; Clastre, M.; et al. A three enzyme system to generate the Strychnos alkaloid scaffold from a central biosynthetic intermediate. Nat. Commun. 2017, 8, 316. [Google Scholar] [CrossRef]
- Caputi, L.; Franke, J.; Farrow, S.C.; Chung, K.; Payne, R.M.E.; Nguyen, T.-D.; Dang, T.-T.T.; Soares Teto Carqueijeiro, I.; Koudounas, K.; Dugé de Bernonville, T.; et al. Missing enzymes in the biosynthesis of the anticancer drug vinblastine in Madagascar periwinkle. Science 2018, 360, 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Easson, M.E.A.M.; Simionescu, R.; Hajicek, J.; Thamm, A.M.K.; Salim, V.; De Luca, V. Solution of the multistep pathway for assembly of corynanthean, strychnos, iboga, and aspidosperma monoterpenoid indole alkaloids from 19E-geissoschizine. Proc. Natl. Acad. Sci. USA 2018, 115, 3180–3185. [Google Scholar] [CrossRef] [PubMed]
- Carqueijeiro, I.; Langley, C.; Grzech, D.; Koudounas, K.; Papon, N.; O’Connor, S.E.; Courdavault, V. Beyond the semi-synthetic artemisinin: Metabolic engineering of plant-derived anti-cancer drugs. Curr. Opin. Biotechnol. 2020, 65, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Dugé de Bernonville, T.; Papon, N.; Clastre, M.; O’Connor, S.E.; Courdavault, V. Identifying Missing Biosynthesis Enzymes of Plant Natural Products. Trends Pharmacol. Sci. 2020, 41, 142–146. [Google Scholar] [CrossRef]
- Courdavault, V.; O’Connor, S.E.; Oudin, A.; Besseau, S.P.N. Towards the Microbial Production of Plant-Derived Anticancer Drugs. Trends Cancer 2020, in press. [Google Scholar] [CrossRef]
- St-Pierre, B.; Besseau, S.; Clastre, M.; Courdavault, V.; Courtois, M.; Crèche, J.; Ducos, E.; de Bernonville, T.D.; Dutilleul, C.; Glévarec, G.; et al. Deciphering the Evolution, Cell Biology and Regulation of Monoterpene Indole Alkaloids. Adv. Bot. Res. 2013, 68, 73–109. [Google Scholar]
- De Luca, V.; Marineau, C.; Brisson, N. Molecular cloning and analysis of cDNA encoding a plant tryptophan decarboxylase: Comparison with animal dopa decarboxylases. Proc. Natl. Acad. Sci. USA 1989, 86, 2582–2586. [Google Scholar] [CrossRef]
- Miettinen, K.; Dong, L.; Navrot, N.; Schneider, T.; Burlat, V.; Pollier, J.; Woittiez, L.; van der Krol, S.; Lugan, R.; Ilc, T.; et al. The seco-iridoid pathway from Catharanthus roseus. Nat. Commun. 2014, 5, 3606. [Google Scholar] [CrossRef]
- Dugé de Bernonville, T.; Foureau, E.; Parage, C.; Lanoue, A.; Clastre, M.; Londono, M.A.; Oudin, A.; Houillé, B.; Papon, N.; Besseau, S.; et al. Characterization of a second secologanin synthase isoform producing both secologanin and secoxyloganin allows enhanced de novo assembly of a Catharanthus roseus transcriptome. BMC Genom. 2015, 16, 619. [Google Scholar] [CrossRef]
- Contin, A.; van der Heijden, R.; Lefeber, A.W.; Verpoorte, R. The iridoid glucoside secologanin is derived from the novel triose phosphate/pyruvate pathway in a Catharanthus roseus cell culture. FEBS Lett. 1998, 434, 413–416. [Google Scholar] [CrossRef]
- Walter, M.H.; Hans, J.; Strack, D. Two distantly related genes encoding 1-deoxy-d-xylulose 5-phosphate synthases: Differential regulation in shoots and apocarotenoid-accumulating mycorrhizal roots. Plant J. 2002, 31, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.A.; Walter, M.H.; Ralph, S.G.; Dabrowska, P.; Luck, K.; Urós, E.M.; Boland, W.; Strack, D.; Rodríguez-Concepción, M.; Bohlmann, J.; et al. Functional identification and differential expression of 1-deoxy-D-xylulose 5-phosphate synthase in induced terpenoid resin formation of Norway spruce (Picea abies). Plant Mol. Biol. 2007, 65, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Cordoba, E.; Porta, H.; Arroyo, A.; San Román, C.; Medina, L.; Rodríguez-Concepción, M.; León, P. Functional characterization of the three genes encoding 1-deoxy-D-xylulose 5-phosphate synthase in maize. J. Exp. Bot. 2011, 62, 2023–2038. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-R.; Kim, S.-U.; Chang, Y.-J. Differential expression of three 1-deoxy-D: -xylulose-5-phosphate synthase genes in rice. Biotechnol. Lett. 2005, 27, 997–1001. [Google Scholar] [CrossRef] [PubMed]
- Okada, A.; Shimizu, T.; Okada, K.; Kuzuyama, T.; Koga, J.; Shibuya, N.; Nojiri, H.; Yamane, H. Elicitor induced activation of the methylerythritol phosphate pathway toward phytoalexins biosynthesis in rice. Plant Mol. Biol. 2007, 65, 177–187. [Google Scholar] [CrossRef]
- Rohdich, F.; Zepeck, F.; Adam, P.; Hecht, S.; Kaiser, J.; Laupitz, R.; Gräwert, T.; Amslinger, S.; Eisenreich, W.; Bacher, A.; et al. The deoxyxylulose phosphate pathway of isoprenoid biosynthesis: Studies on the mechanisms of the reactions catalyzed by IspG and IspH protein. Proc. Natl. Acad. Sci. USA 2003, 100, 1586–1591. [Google Scholar] [CrossRef]
- Ramos-Valdivia, A.C.; van der Heijden, R.; Verpoorte, R. Isopentenyl diphosphate isomerase: A core enzyme in isoprenoid biosynthesis. A review of its biochemistry and function. Nat. Prod. Rep. 1997, 14, 591–603. [Google Scholar] [CrossRef]
- Simkin, A.J.; Miettinen, K.; Claudel, P.; Burlat, V.; Guirimand, G.; Courdavault, V.; Papon, N.; Meyer, S.; Godet, S.; St-Pierre, B.; et al. Characterization of the plastidial geraniol synthase from Madagascar periwinkle which initiates the monoterpenoid branch of the alkaloid pathway in internal phloem associated parenchyma. Phytochemistry 2013, 85, 36–43. [Google Scholar] [CrossRef]
- Courdavault, V.; Papon, N.; Clastre, M.; Giglioli-Guivarc’h, N.; St-Pierre, B.; Burlat, V. A look inside an alkaloid multisite plant: The Catharanthus logistics. Curr. Opin. Plant Biol. 2014, 19, 43–50. [Google Scholar] [CrossRef]
- Dugé de Bernonville, T.; Clastre, M.; Besseau, S.; Oudin, A.; Burlat, V.; Glévarec, G.; Lanoue, A.; Papon, N.; Giglioli-Guivarc’h, N.; St-Pierre, B.; et al. Phytochemical genomics of the Madagascar periwinkle: Unravelling the last twists of the alkaloid engine. Phytochemistry 2015, 113, 9–23. [Google Scholar] [CrossRef]
- Burlat, V.; Oudin, A.; Courtois, M.; Rideau, M.; St-Pierre, B. Co-expression of three MEP pathway genes and geraniol 10-hydroxylase in internal phloem parenchyma of Catharanthus roseus implicates multicellular translocation of intermediates during the biosynthesis of monoterpene indole alkaloids and isoprenoid-derive. Plant J. 2004, 38, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Geu-Flores, F.; Sherden, N.H.; Courdavault, V.; Burlat, V.; Glenn, W.S.; Wu, C.; Nims, E.; Cui, Y.; O’Connor, S.E. An alternative route to cyclic terpenes by reductive cyclization in iridoid biosynthesis. Nature 2012, 492, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Guirimand, G.; Guihur, A.; Phillips, M.A.; Oudin, A.; Glévarec, G.; Melin, C.; Papon, N.; Clastre, M.; St-Pierre, B.; Rodríguez-Concepción, M.; et al. A single gene encodes isopentenyl diphosphate isomerase isoforms targeted to plastids, mitochondria and peroxisomes in Catharanthus roseus. Plant Mol. Biol. 2012, 79, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Oudin, A.; Courtois, M.; Rideau, M.; Clastre, M. The iridoid pathway in Catharanthus roseus alkaloid biosynthesis. Phytochem. Rev. 2007, 6, 259–276. [Google Scholar] [CrossRef]
- Guirimand, G.; Burlat, V.; Oudin, A.; Lanoue, A.; St-Pierre, B.; Courdavault, V. Optimization of the transient transformation of Catharanthus roseus cells by particle bombardment and its application to the subcellular localization of hydroxymethylbutenyl 4-diphosphate synthase and geraniol 10-hydroxylase. Plant Cell Rep. 2009, 28, 1215–1234. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.R.; Hines, K.M. Stromules: Probing Formation and Function. Plant Physiol. 2018, 176, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Mueller-Schuessele, S.J.; Michaud, M. Plastid Transient and Stable Interactions with Other Cell Compartments. Methods Mol. Biol. 2018, 1829, 87–109. [Google Scholar]
- Chahed, K.; Oudin, A.; Giglioli-Guivarc’h, N.; Hamdi, S.; Chénieux, J.C.; Rideau, M.; Clastre, M. 1-Deoxy-d-xylulose 5-phosphate synthase from periwinkle: cDNA identification and induced gene expression in terpenoid indole alkaloid-producing cells. Plant Physiol. Biochem. 2000, 38, 559–566. [Google Scholar] [CrossRef]
- Veau, B.; Courtois, M.; Oudin, A.; Chenieux, J.C.; Rideau, M.; Clastre, M. Cloning and expression of cDNAs encoding two enzymes of the MEP pathway in Catharanthus roseus. Biochim. Biophys. Acta 2000, 1517, 159–163. [Google Scholar] [CrossRef]
- Oudin, A.; Mahroug, S.; Courdavault, V.; Hervouet, N.; Zelwer, C.; Rodríguez-Concepción, M.; St-Pierre, B.; Burlat, V. Spatial distribution and hormonal regulation of gene products from methyl erythritol phosphate and monoterpene-secoiridoid pathways in Catharanthus roseus. Plant Mol. Biol. 2007, 65, 13–30. [Google Scholar] [CrossRef]
- Campos, N.; Rodríguez-Concepción, M.; Sauret-Güeto, S.; Gallego, F.; Lois, L.M.; Boronat, A. Escherichia coli engineered to synthesize isopentenyl diphosphate and dimethylallyl diphosphate from mevalonate: A novel system for the genetic analysis of the 2-C-methyl-d-erythritol 4-phosphate pathway for isoprenoid biosynthesis. Biochem. J. 2001, 353, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Sauret-Güeto, S.; Ramos-Valdivia, A.; Ibáñez, E.; Boronat, A.; Rodríguez-Concepción, M. Identification of lethal mutations in Escherichia coli genes encoding enzymes of the methylerythritol phosphate pathway. Biochem. Biophys. Res. Commun. 2003, 307, 408–415. [Google Scholar] [CrossRef]
- Medicinal Plant Genomics Ressource MPGR RNA seq Data. Available online: http://medicinalplantgenomics.msu.edu/index.shtml (accessed on 10 March 2020).
- Courdavault, V.; Thiersault, M.; Courtois, M.; Gantet, P.; Oudin, A.; Doireau, P.; St-Pierre, B.; Giglioli-Guivarc’h, N. CaaX-prenyltransferases are essential for expression of genes involved in the early stages of monoterpenoid biosynthetic pathway in Catharanthus roseus cells. Plant Mol. Biol. 2005, 57, 855–870. [Google Scholar] [CrossRef] [PubMed]
- Leménager, D.; Ouelhazi, L.; Mahroug, S.; Veau, B.; St-Pierre, B.; Rideau, M.; Aguirreolea, J.; Burlat, V.; Clastre, M. Purification, molecular cloning, and cell-specific gene expression of the alkaloid-accumulation associated protein CrPS in Catharanthus roseus. J. Exp. Bot. 2005, 56, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Carqueijeiro, I.; Sepúlveda, L.J.; Mosquera, A.; Payne, R.; Corbin, C.; Papon, N.; de Bernonville, T.D.; Besseau, S.; Lanoue, A.; Glévarec, G.; et al. Vacuole-Targeted Proteins: Ins and Outs of Subcellular Localization Studies. Methods Mol. Biol. 2018, 1789, 33–54. [Google Scholar]
- Foureau, E.; Carqueijeiro, I.; Dugé de Bernonville, T.; Melin, C.; Lafontaine, F.; Besseau, S.; Lanoue, A.; Papon, N.; Oudin, A.; Glévarec, G.; et al. Prequels to Synthetic Biology: From Candidate Gene Identification and Validation to Enzyme Subcellular Localization in Plant and Yeast Cells. Methods Enzymol. 2016, 576, 167–206. [Google Scholar]
- Besseau, S.; Kellner, F.; Lanoue, A.; Thamm, A.M.K.; Salim, V.; Schneider, B.; Geu-Flores, F.; Höfer, R.; Guirimand, G.; Guihur, A.; et al. A pair of tabersonine 16-hydroxylases initiates the synthesis of vindoline in an organ-dependent manner in Catharanthus roseus. Plant Physiol. 2013, 163, 1792–1803. [Google Scholar] [CrossRef]
- Parage, C.; Foureau, E.; Kellner, F.; Burlat, V.; Mahroug, S.; Lanoue, A.; Dugé de Bernonville, T.; Londono, M.A.; Carqueijeiro, I.; Oudin, A.; et al. Class II Cytochrome P450 Reductase Governs the Biosynthesis of Alkaloids. Plant Physiol. 2016, 172, 1563–1577. [Google Scholar] [CrossRef]
- Larsen, B.; Fuller, V.L.; Pollier, J.; Van Moerkercke, A.; Schweizer, F.; Payne, R.; Colinas, M.; O’Connor, S.E.; Goossens, A.; Halkier, B.A. Identification of Iridoid Glucoside Transporters in Catharanthus roseus. Plant Cell Physiol. 2017, 58, 1507–1518. [Google Scholar] [CrossRef]
- Ytterberg, A.J.; Peltier, J.-B.; van Wijk, K.J. Protein profiling of plastoglobules in chloroplasts and chromoplasts. A surprising site for differential accumulation of metabolic enzymes. Plant Physiol. 2006, 140, 984–997. [Google Scholar] [CrossRef]
- Perello, C.; Llamas, E.; Burlat, V.; Ortiz-Alcaide, M.; Phillips, M.A.; Pulido, P.; Rodriguez-Concepcion, M. Differential Subplastidial Localization and Turnover of Enzymes Involved in Isoprenoid Biosynthesis in Chloroplasts. PLoS ONE 2016, 11, e0150539. [Google Scholar] [CrossRef] [PubMed]
- Pulido, P.; Llamas, E.; Llorente, B.; Ventura, S.; Wright, L.P.; Rodríguez-Concepción, M. Specific Hsp100 Chaperones Determine the Fate of the First Enzyme of the Plastidial Isoprenoid Pathway for Either Refolding or Degradation by the Stromal Clp Protease in Arabidopsis. PLoS Genet. 2016, 12, e1005824. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Smita, S.S.; Singh, A.K.; Shanker, K.; Nagegowda, D.A. Heteromeric and homomeric geranyl diphosphate synthases from Catharanthus roseus and their role in monoterpene indole alkaloid biosynthesis. Mol. Plant 2013, 6, 1531–1549. [Google Scholar] [CrossRef] [PubMed]
- Schattat, M.; Barton, K.; Baudisch, B.; Klösgen, R.B.; Mathur, J. Plastid stromule branching coincides with contiguous endoplasmic reticulum dynamics. Plant Physiol. 2011, 155, 1667–1677. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.R.; Sattarzadeh, A. Trafficking of proteins through plastid stromules. Plant Cell 2013, 25, 2774–2782. [Google Scholar] [CrossRef] [PubMed]
- Mehrshahi, P.; Stefano, G.; Andaloro, J.M.; Brandizzi, F.; Froehlich, J.E.; DellaPenna, D. Transorganellar complementation redefines the biochemical continuity of endoplasmic reticulum and chloroplasts. Proc. Natl. Acad. Sci. USA 2013, 110, 12126–12131. [Google Scholar] [CrossRef] [PubMed]
- Guirimand, G.; Guihur, A.; Ginis, O.; Poutrain, P.; Héricourt, F.; Oudin, A.; Lanoue, A.; St-Pierre, B.; Burlat, V.; Courdavault, V. The subcellular organization of strictosidine biosynthesis in Catharanthus roseus epidermis highlights several trans-tonoplast translocations of intermediate metabolites. FEBS J. 2011, 278, 749–763. [Google Scholar] [CrossRef]
- Mahroug, S.; Courdavault, V.; Thiersault, M.; St-Pierre, B.; Burlat, V. Epidermis is a pivotal site of at least four secondary metabolic pathways in Catharanthus roseus aerial organs. Planta 2006, 223, 1191–1200. [Google Scholar] [CrossRef]
- PSORT, Kenta Nakai; Human Genome Center, Institute for Medical Science, University ot Tokyo: Tokyo, Japan, 2007.
- Predotar, version 1.3; Unité de Recherches en Génomique Info: Versailles, France, 2010.
- Emanuelsson, O.; Nielsen, H.; Von Heijne, G. ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci. 1999, 8, 978–984. [Google Scholar] [CrossRef]
- Ison, J.; Rapacki, K.; Ménager, H.; Kalaš, M.; Rydza, E.; Chmura, P.; Booth, T. Tools and data services registry: A community effort to document bioinformatics resources. Nucleic Acids Res. 2016, 44, D38–D47. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; Von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.K.; Cai, X.; Nebenführ, A. A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 2007, 51, 1126–1136. [Google Scholar] [CrossRef] [PubMed]
- Vidi, P.A.; Kanwischer, M.; Baginsky, S.; Austin, J.R.; Csucs, G.; Dörmann, P.; Kessler, F.; Bréhélin, C. Tocopherol cyclase (VTE1) localization and vitamin E accumulation in chloroplast plastoglobule lipoprotein particles. J. Biol. Chem. 2006, 281, 11225–11234. [Google Scholar] [CrossRef] [PubMed]
- Pulido, P.; Toledo-Ortiz, G.; Phillips, M.A.; Wright, L.P.; Rodríguez-Concepción, M. Arabidopsis J-protein J20 delivers the first enzyme of the plastidial isoprenoid pathway to protein quality control. Plant Cell 2013, 25, 4183–4194. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Identity | PSORT | Predotar | ChloroP | MitoProt | TargetP |
|---|---|---|---|---|---|---|
| DXS/DXS2.1 | 74% 64% 41% | ER (membrane) 0.550 Peroxisome 0.102 ER (lumen) 0.100 | P 0.45 M 0.01 E 0.51 | Y (0.509) 35 residues | 0.636 37 residues | cTP (0.375) mTP (0.217) |
| DXS2.2 | 65% 56% 34% | P (stroma) 0.570 M (matrix) 0.475 Cyto 0.450 | P 0.91 M 0.06 E 0.08 | Y (0.565) 62 residues | 0.327 16 residues | cTP (0.849) mTP (0.301) |
| DXR | 89% | P (stroma) 0.520 Cyto 0.450 M (matrix) 0.360 | P 0.30 M 0.01 E 0.69 | Y (0.509) 83 residues | 0.596 66 residues | cTP (0.303) mTP (0.049) |
| CMS | 76% | P (stroma) 0.865 P (thy mb) 0.515 P (thy space) 0.461 | P 0.07 M 0.20 E 0.71 | Y (0.576) 66 residues | 0.5259 Not predictable | cTP (0.932) mTP (0.083) |
| CMK | 75% | P (stroma) 0.899 P (thy mb) 0.595 P (thy space) 0.595 | P 0.84 M 0.01 E 0.16 | Y (0.582) 57 residues | 0.949 59 residues | cTP (0.895) mTP (0.084) |
| MECS | 85% | Plasma mb 0.700 P (thy mb) 0.547 M (inner mb) 0.415 | P 0.97 M 0.03 E 0.03 | Y (0.582) 55 residues | 0.5829 58 residues | cTP (0.955) mTP (0.024) |
| HDS | 87% | ER (lumen) 0.850 Plasma mb 0.790 Peroxisome 0.300 | P 0.66 M 0.02 E 0.33 | Y (0.526) 40 residues | 0.009 18 residues | cTP (0.496) mTP (0.098) |
| HDR | 79% | M (matrix) 0.850 Peroxisome 0.322 Nucleus 0.300 | P 0.80 M 0.02 E 0.20 | Y (0.530) 34 residues | 0.104 Not predictable | cTP (0.827) mTP (0.129) |
| RANK | 1 | 2 | 8 | 14 | 16 | 20 | 64 | NF |
|---|---|---|---|---|---|---|---|---|
| PCC | 1 | 0,98859 | 0,96838 | 0,96244 | 0,96168 | 0,955751 | 0,921181 | NF |
| CRA LOCUS | 618 | 4910 | 1424 | 7966 | 891 | 4962 | 381 | 2720 |
| NAME | DXS2.1 | CMK | HDS | MECS | CMS | DXR | DXS2.2 | HDR |
| CRA_AA | 23 | 66 | 149 | 40 | 26 | 69 | 9 | NF |
| CRA_AN | 49 | 65 | 144 | 82 | 33 | 65 | 15 | |
| CRA_AM | 36 | 81 | 161 | 89 | 30 | 92 | 9 | |
| CRA_AO | 34 | 37 | 48 | 27 | 10 | 52 | 16 | |
| CRA_AP | 221 | 113 | 239 | 87 | 39 | 138 | 45 | |
| CRA_AE | 85 | 70 | 109 | 57 | 23 | 71 | 16 | |
| CRA_AG | 416 | 227 | 287 | 126 | 46 | 216 | 106 | |
| CRA_AF | 191 | 129 | 242 | 96 | 45 | 132 | 100 | |
| CRA_AH | 52 | 32 | 105 | 58 | 10 | 40 | 19 | |
| CRA_AB | 65 | 44 | 164 | 48 | 13 | 42 | 9 | |
| CRA_AC | 26 | 26 | 106 | 41 | 10 | 22 | 5 | |
| CRA_AD | 22 | 25 | 98 | 36 | 12 | 26 | 4 | |
| CRA_AL | 26 | 21 | 78 | 32 | 7 | 26 | 5 | |
| CRA_AI | 38 | 29 | 85 | 42 | 14 | 23 | 15 | |
| CRA_AJ | 41 | 33 | 74 | 41 | 15 | 28 | 19 | |
| CRA_AK | 35 | 34 | 72 | 32 | 5 | 26 | 5 | |
| CRA_AS | 102 | 103 | 206 | 78 | 31 | 173 | 36 | |
| CRA_AQ | 181 | 146 | 310 | 135 | 51 | 254 | 104 | |
| CRA_AR | 67 | 60 | 151 | 75 | 15 | 123 | 30 | |
| CRA_AT | 141 | 110 | 234 | 66 | 29 | 243 | 42 | |
| CRA_AU | 803 | 534 | 1084 | 382 | 146 | 1374 | 256 | |
| CRA_AV | 186 | 148 | 275 | 86 | 41 | 281 | 79 | |
| CRA_AW | 832 | 573 | 1154 | 335 | 136 | 1288 | 529 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guirimand, G.; Guihur, A.; Perello, C.; Phillips, M.; Mahroug, S.; Oudin, A.; Dugé de Bernonville, T.; Besseau, S.; Lanoue, A.; Giglioli-Guivarc’h, N.; et al. Cellular and Subcellular Compartmentation of the 2C-Methyl-D-Erythritol 4-Phosphate Pathway in the Madagascar Periwinkle. Plants 2020, 9, 462. https://doi.org/10.3390/plants9040462
Guirimand G, Guihur A, Perello C, Phillips M, Mahroug S, Oudin A, Dugé de Bernonville T, Besseau S, Lanoue A, Giglioli-Guivarc’h N, et al. Cellular and Subcellular Compartmentation of the 2C-Methyl-D-Erythritol 4-Phosphate Pathway in the Madagascar Periwinkle. Plants. 2020; 9(4):462. https://doi.org/10.3390/plants9040462
Chicago/Turabian StyleGuirimand, Grégory, Anthony Guihur, Catalina Perello, Michael Phillips, Samira Mahroug, Audrey Oudin, Thomas Dugé de Bernonville, Sébastien Besseau, Arnaud Lanoue, Nathalie Giglioli-Guivarc’h, and et al. 2020. "Cellular and Subcellular Compartmentation of the 2C-Methyl-D-Erythritol 4-Phosphate Pathway in the Madagascar Periwinkle" Plants 9, no. 4: 462. https://doi.org/10.3390/plants9040462
APA StyleGuirimand, G., Guihur, A., Perello, C., Phillips, M., Mahroug, S., Oudin, A., Dugé de Bernonville, T., Besseau, S., Lanoue, A., Giglioli-Guivarc’h, N., Papon, N., St-Pierre, B., Rodríguez-Concepcíon, M., Burlat, V., & Courdavault, V. (2020). Cellular and Subcellular Compartmentation of the 2C-Methyl-D-Erythritol 4-Phosphate Pathway in the Madagascar Periwinkle. Plants, 9(4), 462. https://doi.org/10.3390/plants9040462