1. Introduction
Doubled haploids (DH) are an efficient alternative to conventional inbred lines developed through recurrent selfing in maize breeding. Compared to conventional inbred lines, DH lines facilitate faster variety registration and intellectual protection due to their distinctness, uniformity, and stability, thereby supporting quicker deployment of maize hybrids in the market [
1,
2]. Higher genetic variance and higher heritabilities when using DH lines compared to conventional selfed lines in per se and testcross evaluation facilitate accurate selection decisions. The major steps in maize DH line production pipelines are in vivo induction of haploids, separation of haploids from diploids, doubling the genome in haploid (D
0) plants, and production of D
1 seed from D
0 plants [
3,
4]. Development of protocols for large-scale DH line production and continuous process improvement in the last three decades resulted in higher efficiencies at different steps of DH line development [
3].
DH production in maize relies on in vivo haploid induction using maternal haploid inducers. Maternal haploid inducers with high haploid induction rates were developed in both temperate and tropical genetic backgrounds making large-scale induction of haploids possible from diverse maize germplasm in a cost-efficient manner [
2,
5,
6]. Haploids are separated from diploids based on phenotypic markers integrated in the inducers, like
R1-nj [
7,
8], red root [
6,
9], and high oil [
10]. Recently, efforts were put into automation of haploid identification process using high oil [
11,
12] and
R1-nj markers [
13,
14,
15,
16] that can lead to significant resource savings.
Chromosomal doubling is essential for restoring fertility and production of seed from haploids. However, improving efficiency in the chromosomal doubling process is relatively less researched/published compared to other steps in the DH production pipeline. Spontaneous chromosomal doubling occurs in too low frequencies in most maize genotypes [
17,
18]. Therefore, chromosomal doubling by treating the haploids with mitosis inhibiting chemicals is commonly practiced [
19,
20]. Chromosome doubling protocols involves several labor-intensive steps and use of expensive chemicals. Hence, increasing chromosomal doubling efficiency is critical to reduce the cost per DH line [
3].
Commonly reported chemicals for achieving artificial chromosome doubling in maize haploids include colchicine [
21,
22,
23], antimitotic herbicides [
20], and nitrous oxide (N
2O) gas [
24,
25]. Colchicine is much more toxic compared to N
2O gas and anti-mitotic herbicides [
20,
25]. However, colchicine is more commonly used in DH production pipelines because the colchicine is more readily available for purchase in large quantities and protocols are well-established. Several methods of colchicine application to haploids were described in literature that include (a) treating the seed; (b) immersing germinating haploid seedlings in colchicine solution; (c) immersing the roots of haploid seedlings; or (d) injecting colchicine into shoot apical meristem [
3]. Among these, the seedling immersion method is widely adapted for large-scale DH line production as it showed a better success rate compared to seed treatment [
23], root treatment [
22], or injection into shoot apical meristem [
26]. However, there is no publication showing a robust comparison of maize haploid chromosome doubling efficiencies of these methods as most studies used small sample sizes of haploids and different criteria to assess doubling efficiencies. Therefore, the objectives of the present study were (a) to systematically evaluate chromosomal doubling success rates in five different methods of colchicine application to maize haploids; and (b) to further optimize the colchicine application method to achieve higher success rates.
3. Discussion
Chromosome doubling is the most intricate step in maize DH line production as it involves several critical activities like germination of haploids, preparation and treatment of haploid seedlings with mitosis inhibiting chemicals, post-treatment recovery, and transplanting in the field [
19]. Hence, achieving higher success rates in chromosomal doubling could lead to higher efficiencies in DH production pipelines. Protocols used for doubling chromosomes in maize haploids were developed in the 1990s and involve application of colchicine to seed, whole germinating seedlings or roots, or the shoot apical meristematic region of 8–12-day old seedlings [
21,
22,
27]. A systematic comparison of doubling efficiencies among different colchicine-based chromosome doubling protocols based on large enough samples and replicated experiments was lacking in literature. Additionally, there was no uniform criteria used to assess chromosomal doubling efficiency of different protocols. Some studies considered chromosomal counts through microscopic studies [
21], some considered pollen production [
23], while others considered seed production [
26] as indicators of success of a chromosome doubling treatment of maize haploids. Most studies did not consider mortality as a result of toxicity of chromosome doubling chemicals in determining the success. Melchinger et al. [
20] proposed three parameters namely SR, RR, and OSR to assess the success in chromosomal doubling, as these parameters sufficiently consider all the aspects of the chromosomal doubling step. These parameters were used to assess the efficiency of chromosomal doubling using antimitotic herbicides and N
2O gas in comparison to the standard method of immersing germinating seedlings in colchicine solution [
20,
25]. Among the three parameters, OSR was found to be particularly important for maize DH production pipelines as it considers all the factors important for seed production from haploids.
In this study, the criteria described by Melchinger et al. [
20] were used to assess chromosomal doubling success through five different methods of colchicine application using replicated trials and large sample sizes. The study revealed that immersion of haploid seedlings from root tips up to 2–3 cm above seed level at V2 growth stage (Method II) is the most effective method in doubling the chromosomes. This method resulted in more than double the OSR compared to standard seedling immersion method at VE stage (Method I). At V2 stage if seedlings were immersed in colchicine from root tips up to half the length of roots (Method V), it resulted in significantly lower OSR than immersing the seedlings until 2–3 cm above the seed level. Previous studies that compared immersion of roots only with standard seedling immersion method using colchicine [
22] and antimitotic herbicides [
20] showed very low success for root immersion compared to seedling immersion. Together, immersion of the crown region encompassing the shoot apical meristem along with roots appears to be essential to achieve higher levels of OSR. The seedling immersion method (Method I) used in this study was slightly different from seedling immersion method used earlier [
20,
25] in that the seedlings were treated in a lower concentration of colchicine (0.04% vs. 0.06%) and for much longer time (12 h vs. 8 h). Earlier studies [
20,
25] reported an OSR of ≈10% for the seedling immersion method in the majority of the experiments. Lower OSR for this method in our experiments could be the result of using a lower concentration of colchicine.
Our experiments also revealed that application of colchicine directly to seedling trays in greenhouse (Method III) and to imbibed seed (Method IV) were very inefficient in causing chromosomal doubling. A previous report on seed treatment by Chalyk [
23] also indicated that seed treatment did not result in chromosomal doubling. However, another report by Gayen et al. [
21] indicated that when seeds were treated with 0.06% colchicine with a portion of plumule cut or not cut, it showed higher doubling rates than treating germinating seedlings at the same colchicine concentration. However, the sample sizes of 21 and 50 seeds in these two earlier studies [
23] and [
21], respectively were inadequate to arrive at meaningful conclusions compared to the present study with large sample sizes.
Since Method II involving immersion of whole roots and crown region was found to be more efficient in doubling haploid chromosomes, we tested if increasing concentrations of colchicine and DMSO could further increase OSR. Increasing DMSO concentration from 0.1% to 0.5% led to a slight but nonsignificant increase in OSR. However, increasing colchicine concentration had a very strong positive effect on OSR. It may be possible to achieve even higher doubling rates in Method II than what is reported in this study by further increasing colchicine concentrations. When evaluated in multiple populations, Method II consistently showed significantly higher SR, RR, and OSR than Method I in all the 12 populations studied. In both Methods I and II, OSR varied among the populations. Earlier studies also pointed to genotypic influence on artificial doubling of haploid chromosomes [
18] or spontaneous chromosomal doubling [
17,
28,
29].
The improved method of immersing seedling roots and crown region in 0.07%–0.1% colchicine and 0.5% DMSO is the most efficient method of chromosomal doubling reported so far. This method has several advantages compared to the currently used seedling immersion method (Method I). As OSR is several-fold higher in Method II compared to Method I, use of Method II in DH pipelines, as reported in this study, could result in significant resource savings. For example, field space in induction nursery, greenhouse space, and field space in D
0 nursery can be reduced significantly as the number of haploids to be generated and handled will be much lower through Method II. Currently, when using Method I, significantly high labor costs are incurred as it involves labor-intensive steps like seed germination in paper towels, checking for germination for 3–4 days, cutting the shoot tips, doubling treatment, potting in greenhouse, caring in greenhouse for 8–12 days, and transplanting in the field. As Method II involves relatively fewer steps like germination in greenhouse trays, caring for seedlings for 8–12 days, doubling treatment followed by direct transplanting in the field, relatively less labor is needed. In addition, it is much easier for people to handle relatively bigger seedlings at V2 stage in Method II than handling germinating seedlings in Method I. Higher success rate in Method II also facilitates application of genomic technologies like genomic selection at haploid stage that leads to delivery of DH lines with desirable haplotypes. In addition, increased seed quantity on DH ears from Method II also facilitates phenotypic and genotypic selection on the majority of DH lines produced without needing an additional cycle of seed increase. However, the increased concentrations of colchicine and DMSO in Method II compared to Method I, will increase the cost of chromosomal doubling treatments but overall DH line production cost might not be significantly affected as the cost of chemicals is a much smaller contributor to the overall DH production costs [
20].
Despite the benefits outlined above, it must be noted that colchicine is very toxic to humans [
20,
30]. Hence it is desirable to use chemicals that are less toxic. Recently, efforts to develop much safer protocols indicated that it is possible to achieve similar levels of chromosomal doubling efficiencies as colchicine in Method I when using anti-mitotic herbicides and N
2O gas [
20,
25]. However, there are still limitations on using safer alternatives like availability and cost of these chemicals. For example, mitotic herbicides like amiprofos-methyl (APM) and pronamide are reported to result in OSR similar to colchicine application as in Method I [
20]. However, both chemicals are not registered for sale as herbicides in most countries (e.g., in Africa and Asia) and hence are not widely available. Pure laboratory grade chemicals are more costly than colchicine and are available only in limited quantities. Use of N
2O gas requires a specialized gas chamber which may be of significant initial establishment cost [
24,
25]. Considering these limitations with non-colchicine-based alternatives, Method II described here could provide a practical solution until the limitations with colchicine alternatives are effectively addressed. Irrespective of the chemical used for doubling haploid chromosomes, it is important to follow adequate occupational health and safety measures.
4. Materials and Methods
4.1. Genetic Materials and Experimental Locations
In Experiments 1–4 (described in subsequent section), haploids derived from 20 CIMMYT maize populations adapted to mid-altitude/subtropical regions of eastern Africa were used. Haploids were induced in the summer cycle of 2015 as described earlier [
31] using a tropical inducer TAIL9 × TAIL8 [
9] at CIMMYT’s Maize Doubled Haploid Facility at Kiboko, Kenya (2.25° S, 37.73° E). Putative haploid kernels were identified based on
R1-nj (Navajo) anthocyanin marker expression on kernels. A set of 1500 putative haploid kernels were taken from each of the induced populations and a balanced bulk was constituted. Required samples sizes were drawn from this balanced bulk and used in experiments carried out at the Maize Doubled Haploid Facility at Kiboko, Kenya. Experiment 1 was carried out in the winter season of 2016; Experiments 2 and 3 were undertaken in summer season of 2017, and Experiment 4 was implemented in winter season of 2018.
For Experiments 5 and 6, haploids derived from 13 CIMMYT maize populations adapted to the subtropics and four populations adapted to the lowland tropics of Mexico were used. Haploids were induced in the winter season of 2017 as described above at CIMMYT’s Agua Fria experimental station (20.26° N, 97.38° W) in the state of Puebla, Mexico, using tropical hybrid inducer CIM2GTAIL006 × CIM2GTAIL009 [
5]. Putative haploid kernels were identified based on
R1-nj marker expression. A set of 1000 haploids were sampled from each population and mixed to make a balanced bulk. Experiments 5 and 6 were carried out in the winter cycle of 2018 at Metztitlan (20.6° N, 98.76° W) in the state of Hidalgo, Mexico.
For Experiment 7, haploids derived from nine subtropical populations and three tropical lowland populations were used. Haploids were induced in summer season of 2017 at Agua Fria. Haploid induction and identification were carried out as described in Experiments 4 and 5. This experiment was carried out in the winter cycle of 2019 at Agua fria experimental station.
All experiments were conducted in randomized complete block design where replications from each treatment were blocked.
4.2. Details of Experiments
4.2.1. Experiment 1
Three different methods of colchicine application were compared for their doubling efficiency. A set of 3000 putative haploid seeds were used to test the doubling efficiency of each method. The experiment was carried out in three replications for each method with 1000 seeds per replication. Method I involving germination of haploid seed in paper towels, immersing germinating seedlings in 0.04% colchicine + 0.1% DMSO after cutting the coleoptile tip was carried out as described by Chaikam and Mahuku [
19]. Treated seedlings were cared for in greenhouse and field as described by Mahuku [
32]. In Method II, seedlings were germinated in 96-well plastic greenhouse trays with volcanic cinder (gravel) of particle size ≈ 2 mm in a greenhouse. Seedlings were grown for 10–12 days until they reached V2 stage. Seedlings were sprayed with foliar nutrients 6 days after planting. Just before the treatment, seedlings were pulled out carefully from volcanic cinder to avoid root damage and washed in a plastic tub with tap water to remove the medium. Washed seedlings were aligned at seed level, bundled and placed in 2-L plastic round containers. Each container holds 120–130 seedlings. Then, 0.04% colchicine and 0.1% DMSO solution was poured into the beaker from one side until all the seedling roots and coleoptile region 2–3 cm above the seed level were immersed in the solution. Seedlings were maintained in colchicine solution in the lab at ambient temperature for 5 h. Spent colchicine solution was collected in a plastic tank for safe disposal. The beakers with seedlings were filled with tap water, gently rinsed, and the water was emptied into a plastic container for disposal. Later, seedlings were washed three times in a sink with tap water. Seedlings were replanted in trays in volcanic cinder and maintained for another 8 days for recovery. Seedlings were sprayed with foliar nutrients 2 days after potting in greenhouse. After recovery, seedlings were transplanted in a well irrigated field and provided good agronomic management as described by Mahuku [
32].
For colchicine application by Method III, seedlings were grown in a greenhouse for 10–12 days until they reached V2 stage as described above. In the greenhouse, one of the benches was lined up with a thick polythene sheet on the bottom and all sides to create a tub that can hold seedling trays. This tub had a depth of approximately twice the height of seedling tray. Seedling trays were placed in the tub and the tub was filled with 0.04% colchicine + 0.1% DMSO solution until the collar was immersed in colchicine for 5 h. After the treatment, colchicine was emptied by sucking the solution with a pump into a plastic container. Tap water was applied to the tub and then the tub and trays with seedlings were washed three times. Seedlings were maintained in the trays for another 8 days and the seedlings were transplanted in the field and managed in the field as described for Method II. In this experiment, control treatment was seedling immersion method treated with tap water instead of colchicine and DMSO solution.
4.2.2. Experiment 2
In this experiment, Method II (as described above) was compared to the seed immersion method (Method IV). Method IV was adapted from Gayen et al. [
21]. A set of 600 putative haploid seeds were used for each treatment, with each treatment having three replications of 200 seeds each. The seeds were placed in mesh bags and the bags were kept in beakers with tap water at room temperature and imbibed for 20 h. After imbibition, mesh bags with seeds were placed in colchicine solution for 5 h. Five different concentrations of colchicine were tested in Method IV ranging from 0.05% to 0.3% colchicine. DMSO was used at a concentration of 0.1% in all the treatments. After the treatment, seeds were washed three times under tap water, and then directly planted in the field. Method II was carried out as described in Experiment 1. The control experiment was same as Method II except that only tap water was used for treatment instead of colchicine + DMSO solution.
4.2.3. Experiment 3
In this experiment, Method II was compared against Method V involving immersion of only root tissues in colchicine solution. A set of 600 putative haploid seeds were used for each treatment, with each treatment having three replications with 200 seeds per replication. In Method V, seedlings were germinated and grown as described for Method II in Experiment 1. Seedlings were immersed at half the length of the roots from root tips in 0.04% colchicine and 0.1% DMSO solution for 5 h. Method II was carried out as described in Experiment 1. The control experiment was same as Method II except that only tap water was used for treatment instead of colchicine + DMSO solution.
4.2.4. Experiment 4
To determine if seedling recovery in a greenhouse is required after chromosomal doubling treatment in Method II, seedlings were grown in a greenhouse and were subjected to doubling treatment as described in Method II in Experiment 1. For each treatment, a set of 2100 putative haploid seeds was used. The experiment was conducted in three replications with each replication having 700 seeds. Treated seedlings were either directly transplanted or transplanted after 8 days of recovery in the greenhouse. The control experiment was same as Method II except that only tap water was used for treatment instead of colchicine + DMSO solution and the seedlings were transplanted after 8 days of recovery in greenhouse.
4.2.5. Experiment 5
To test if increasing DMSO concentration has any effect on success rates in colchicine-based chromosomal doubling, seedlings were subjected to Method II as described in Experiment 1. Colchicine concentration of 0.04% was used in all the treatments. Three DMSO concentrations (0.1%, 0.25%, 0.5%) were tested. A control without adding DMSO was included. All the seedlings were directly transplanted to the field after treatment. For each treatment a set of 600 putative haploid seeds was used and experiment was conducted in three replications with each replication consisting of 200 seeds.
4.2.6. Experiment 6
To test if altering colchicine concentration has any effect on chromosomal doubling, seedlings were subjected to Method II as described in Experiment 1. Three concentrations of colchicine (0.01%, 0.04%, 0.07%, 0.1%) were tested and in all experiments; the treatment included 0.1% DMSO. A control with no colchicine was included. All the seedlings were directly transplanted to the field after treatment. For each treatment a set of 600 putative haploid seeds was used. The experiment was conducted in three replications with each replication consisting of 200 seeds.
4.2.7. Experiment 7
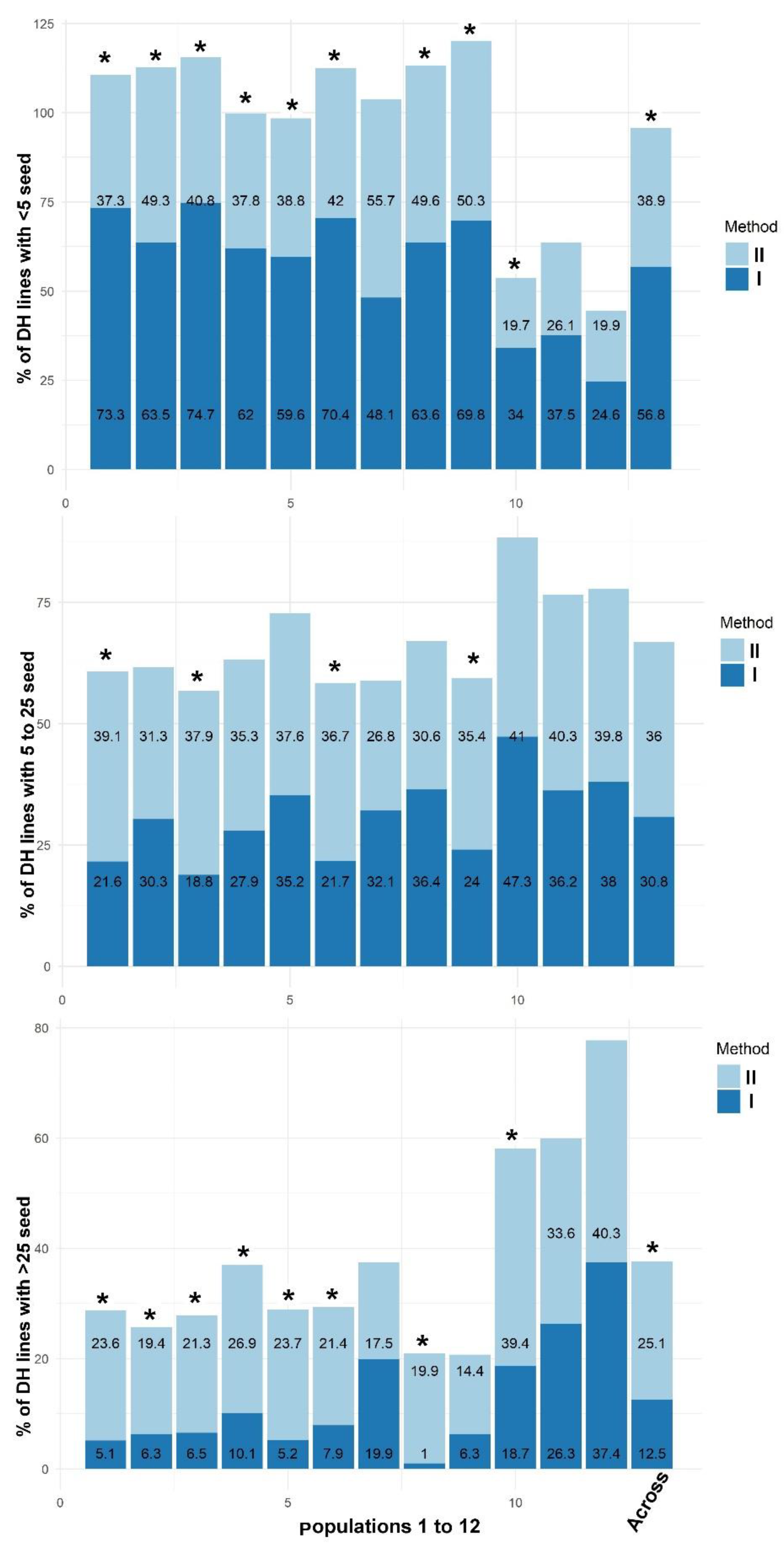
To ascertain the effectiveness of Method II compared to Method I across multiple populations in a large DH production pipeline, haploids derived from 12 populations were subjected to both methods. In Method II, 0.1% colchicine and 0.5% DMSO was used. Method I was as described in Experiment 1. From each population, a set of 1500 putative haploid seeds was used for Method I and another set of 1500 seeds was used for testing Method II. Each method was tested in two replications and 750 seeds per population was used in each replication. Together, a total of 18,000 putative haploids were subjected to Method I across populations and an equal number subjected to Method II. Seed quantity in resulting DH ears from both methods was also compared in this experiment.
4.3. Data Collection and Analysis
In all the experiments except in Experiment 4, the following data were recorded: (1) number of haploid seeds used for experiment; (2) number of seeds germinated; (3) number of seeds/seedlings subjected to treatment; (4) number of seedlings potted in the greenhouse; (5) number of seedlings transplanted in the field; (6) number of D
0 plants surviving at pollination; (7) number of D
0 plants pollinated; (8) number of D
0 plants that produced D
1 seed. In Experiment 4, data on D
0 plants that were pollinated and plants that produced seed was not recorded. The SR, RR, and OSR were calculated as described in [
20] using the following formulae, and were expressed in %:
SR = number of plants surviving at pollination/number of seeds or seedlings subjected to treatment;
RR = number of D0 plants that produced seed/number of surviving D0 plants at pollination;
OSR = number of D0 plants that produced seed/ number of putative haploid seeds or seedlings subjected to chromosome doubling treatment.
In all experiments, except Experiment 2, seed germination and seedling survival rates before chromosomal doubling treatments were not considered in calculating the rates as these losses are not the result of doubling treatments per se. In Experiment 2, for Method IV, seedling germination was considered as the method involved treating the haploid seeds. False positives were also not considered in analyzing the data as they occur at uniform rates in all the treatments.
In Experiment 7, for analysis of seed quantity resulting from fertile D0 plants, the D1 ears obtained were categorized into (1) ears having less than five seeds; (2) ears having 5–25 seeds; and (3) ears having more than 25 seeds. Proportion of D1 ears in each category was obtained by dividing the number of ears in that category by total number of ears. Proportions in each category obtained in Method I were compared to proportions recorded in Method II.
In all experiments, statistical analyses of variance were performed using PROC GLIMMIX procedure in SAS version 9.2 (SAS Institute, Cary, NC, USA).

 ,
, 


{kind=link}