Incidence and Molecular Identification of Apple Necrotic Mosaic Virus (ApNMV) in Southwest China

 and
and 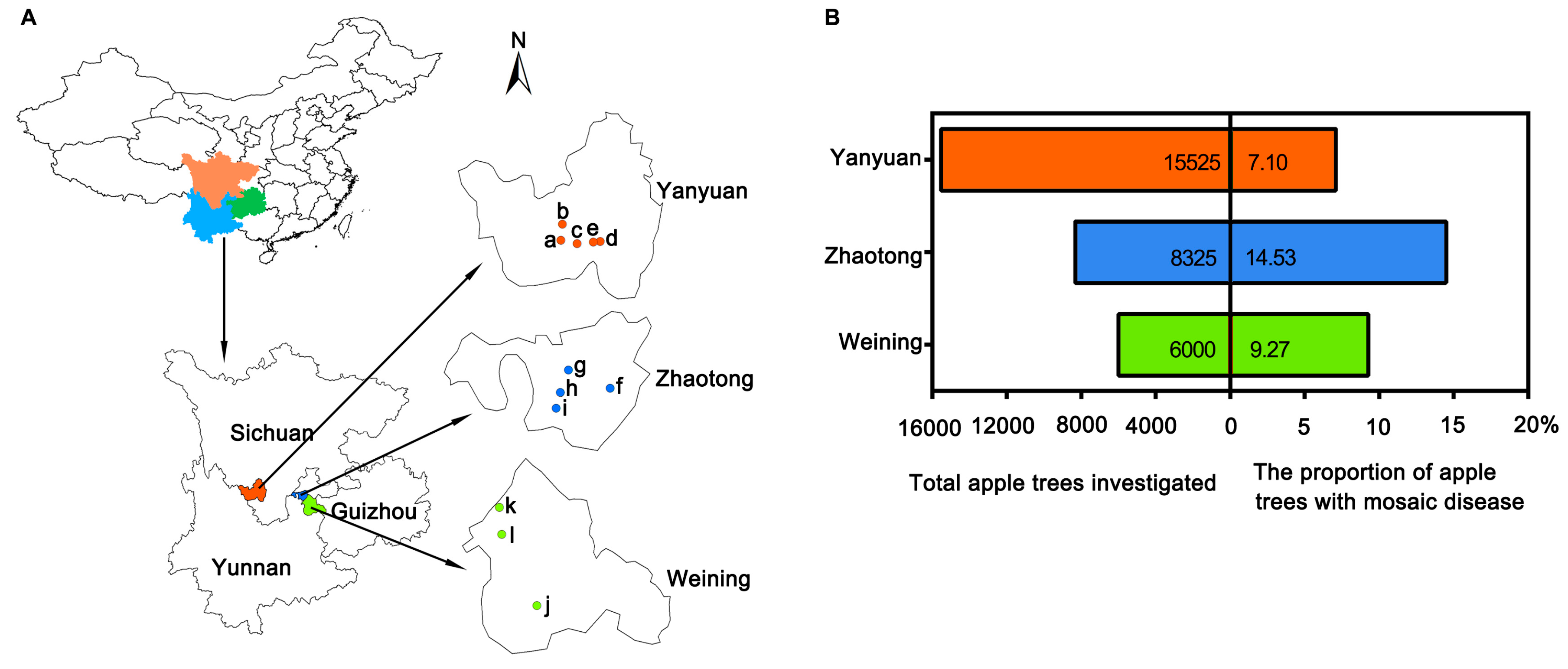{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Apple Mosaic Disease Is Common in Southwest China.
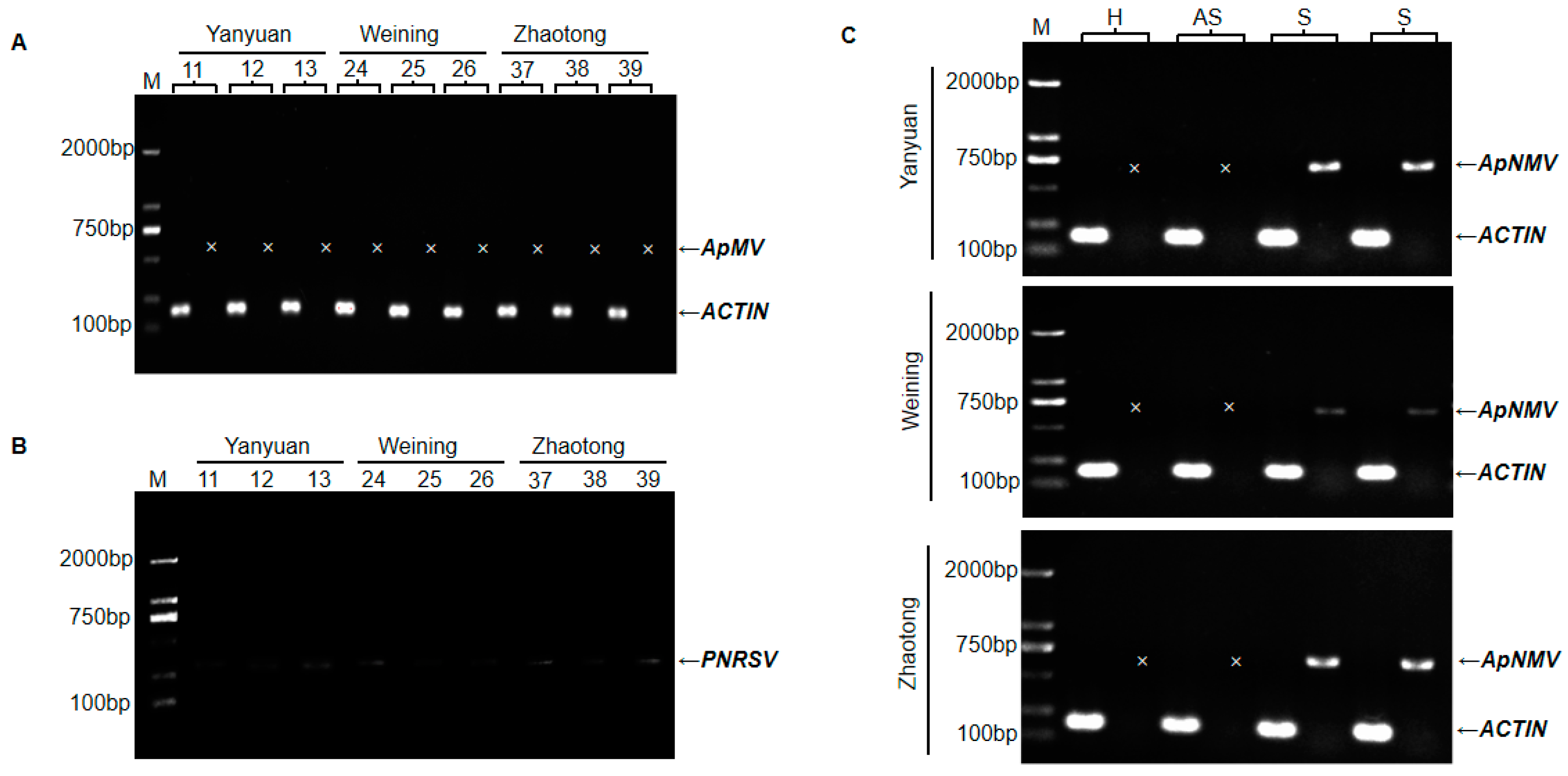
2.2. Pathogen Detection of Apple Mosaic Disease by RT-PCR
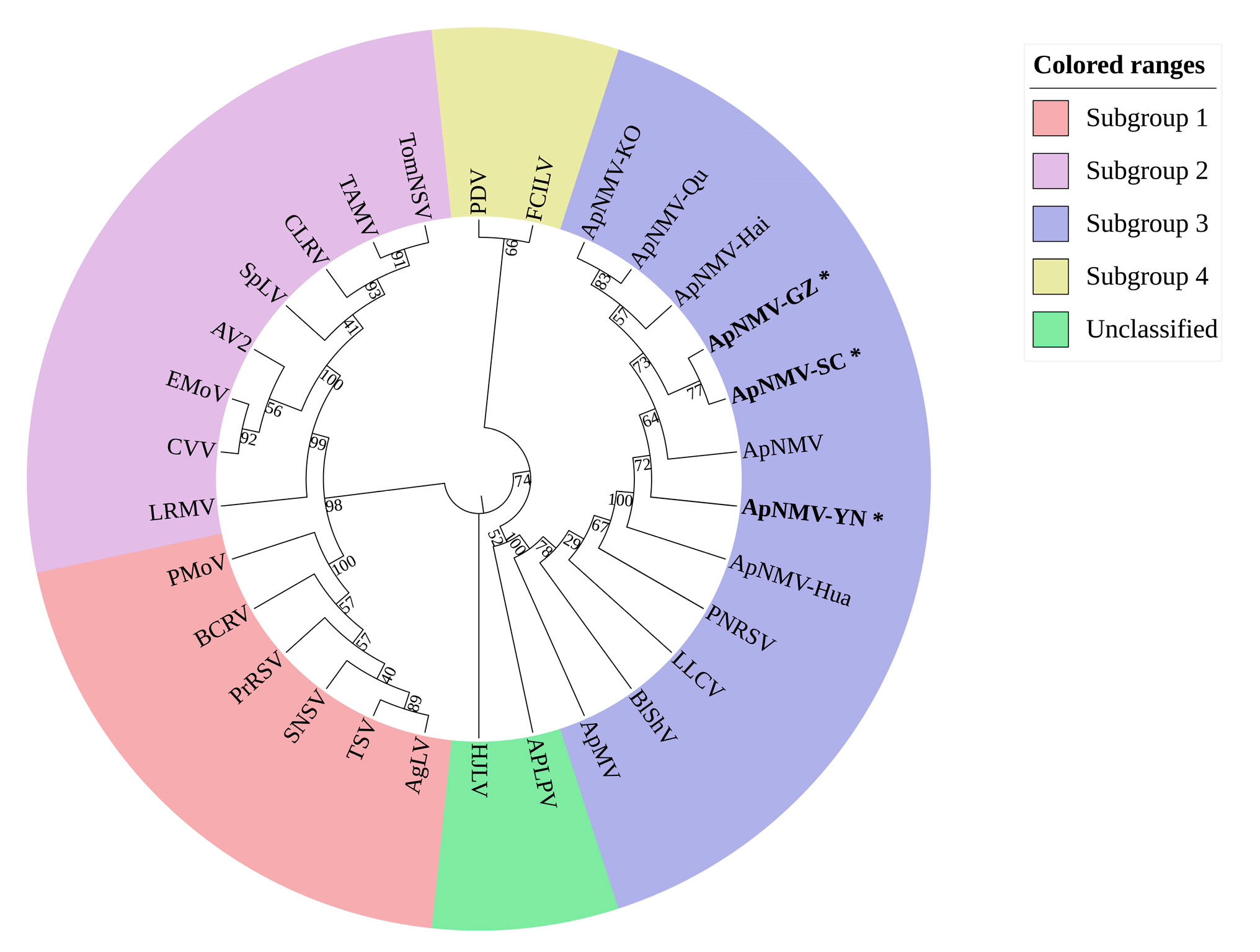
2.3. The Taxonomic Status of ApNMV Isolates in Southwest China.
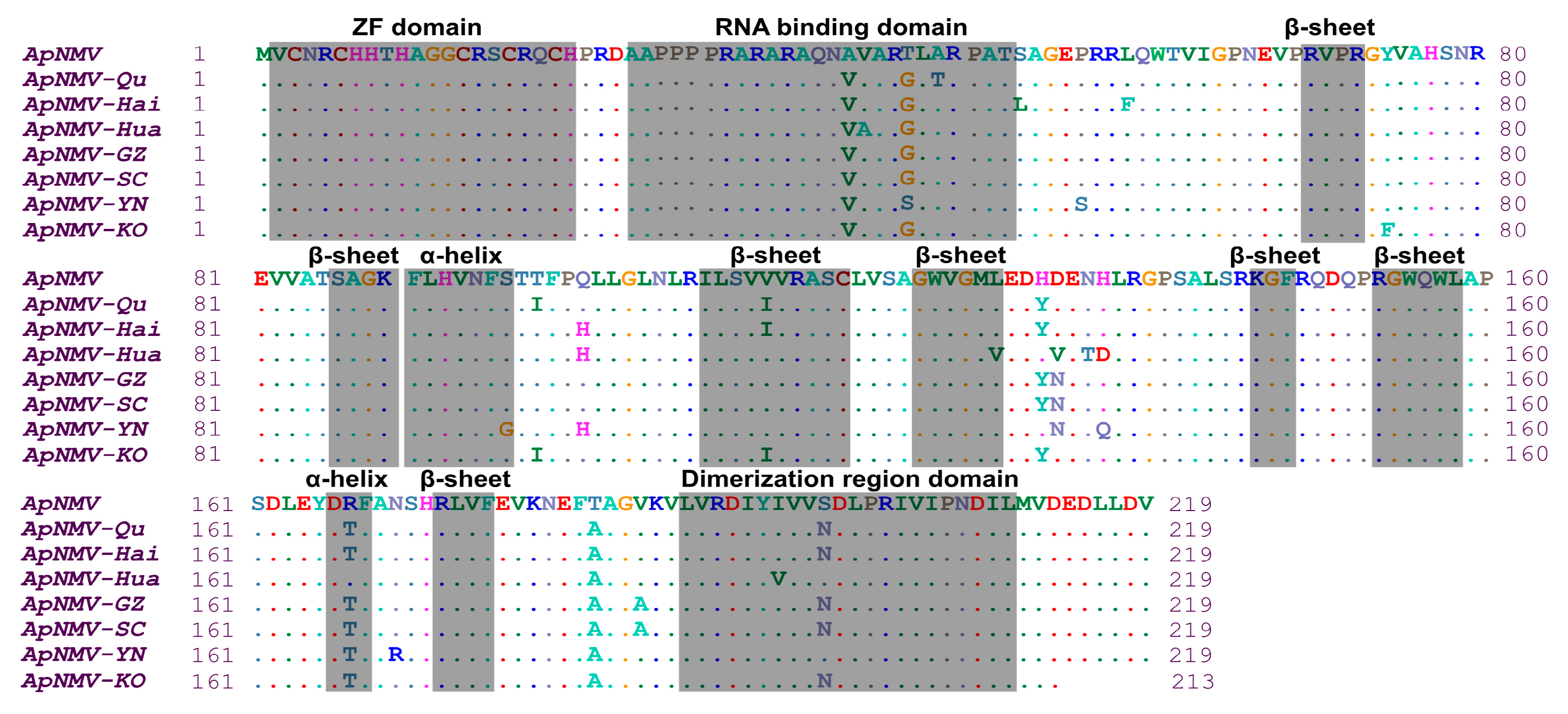
2.4. Diversity Analysis of ApNMV Isolates.
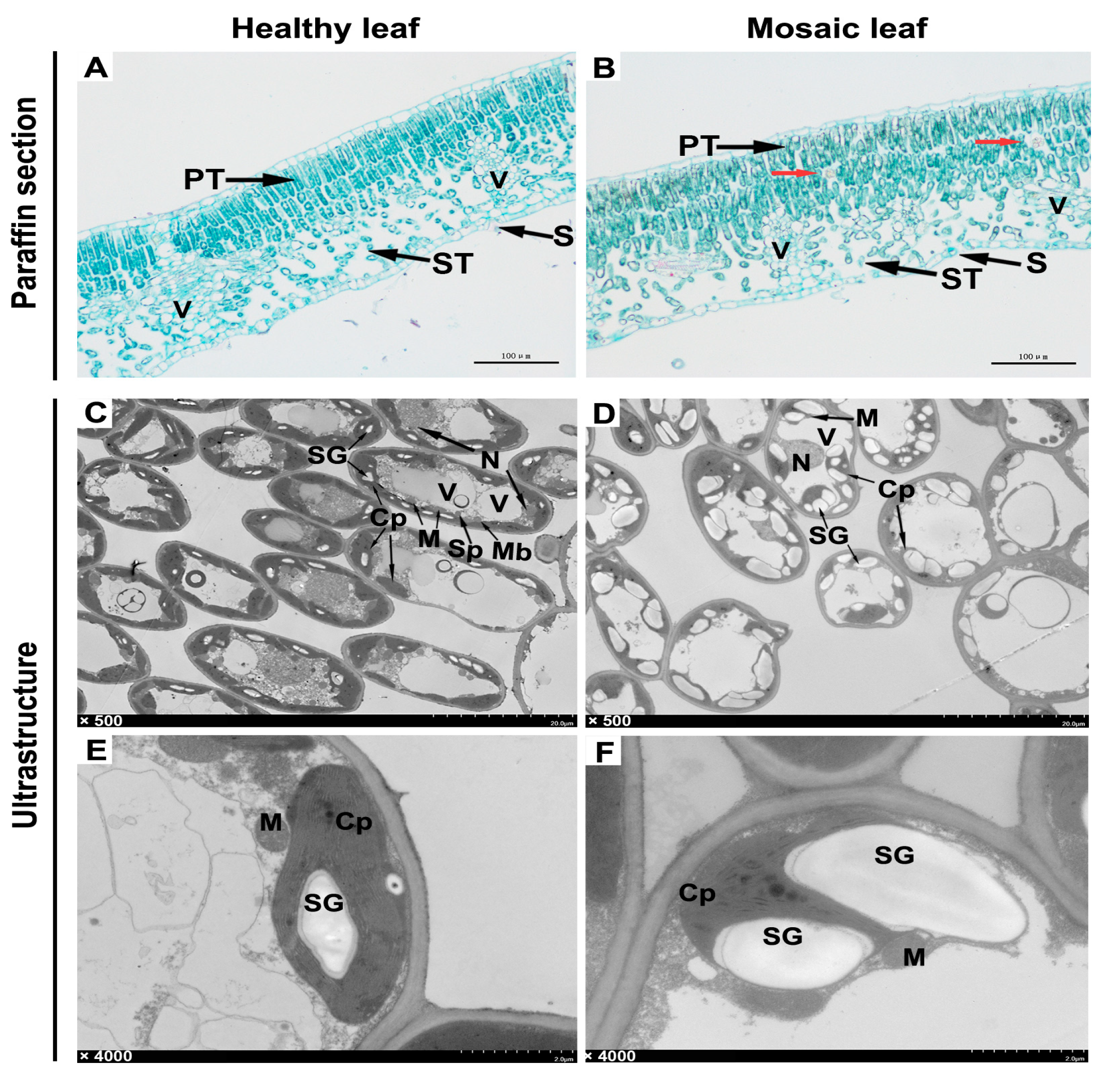
2.5. Histological and Ultrastructural Observation
3. Discussion
4. Materials and Methods
4.1. Field Investigation and Sample Collection of Apple Orchards in Southwest China.
4.2. The Symptoms of Apple Mosaic Disease.
4.3. Total RNA Extraction.
4.4. RT-PCR Detection of ApMV, PNRSV, and ApNMV.
4.5. ApNMV CP Gene Sequencing
4.6. Phylogeny and Diversity Analysis
4.7. Preparation of Paraffin Section and Transmission Electron Microscope Section
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, C.Y.; Chang, Q.; Huo, X.X. Analysis on the Layout of China’s Apple Production Transition. Econ. Geogr. 2018, 38, 141–151. [Google Scholar]
- Wang, Y.N.; Hu, T.L.; Liu, S.X.; Zhao, X.S.; Yang, J.Y.; Cao, K.Q. Research on apple virus in China. Plant Dis. Pests 2010, 1, 35–39. [Google Scholar] [CrossRef]
- Hu, G.J.; Zhang, Z.P.; Dong, Y.F.; Fan, X.D.; Ren, F.; Zhu, H.J. Efficiency of virus elimination from potted apple plants by thermotherapy coupled with shoot-tip grafting. Australas. Plant Pathol. 2015, 44, 167–173. [Google Scholar] [CrossRef]
- Hao, L.; Xie, J.P.; Chen, S.Y.; Wang, S.J.; Gong, Z.Q.; Ling, K.S.; Guo, L.Y.; Fan, Z.F.; Zhou, T.A. multiple RT-PCR assay for simultaneous detection and differentiation of latent viruses and apscarviroids in apple trees. J. Virol. Methods 2016, 234, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.D.; Shi, W.S.; Sunwu, R.Z.; Huang, K.; Wang, J.M. The molecular identification of latent viruses in the main apple production areas in Southwest China. J. Sichuan Univ. (Nat. Sci. Ed.) 2019, 56, 357–362. [Google Scholar] [CrossRef]
- Lim, S.; Moon, J.S.; Cho, I.S.; Kim, H.R.; Lee, S.H. First Report of Apple Hammerhead Viroid Infecting Apple Trees in South Korea. Plant Dis. 2019, 103, 2700–2700. [Google Scholar] [CrossRef]
- Walia, Y.; Dhir, S.; Ram, R.; Zaidi, A.A.; Hallan, V. Identification of the herbaceous host range of Apple scar skin viroid and analysis of its progeny variants. Plant Pathol. 2014, 63, 684–690. [Google Scholar] [CrossRef]
- Aramburua, J.; Rovirab, M. Incidence and natural spread of apple mosaic ilarvirus in hazel in north-east Spain. Plant Pathol. 2000, 49, 423–427. [Google Scholar] [CrossRef]
- Bradford, F.C.; Joley, L. Infectious variegation in the apple. J. Agric. Res. 1933, 46, 901–908. [Google Scholar]
- Grimova, L.; Winkowska, L.; Konrady, M.; Rysanek, P. Apple mosaic virus. Phytopathol. Mediterr. 2016, 55, 1–19. [Google Scholar]
- Noda, H.; Yamagishi, N.; Yaegashi, H.; Xing, F.; Xie, J.P.; Li, S.F.; Zhou, T.; Ito, T.; Yoshikawa, N. Apple necrotic mosaic virus, a novel ilarvirus from mosaic diseased apple trees in Japan and China. J. Gen. Plant Pathol. 2017, 83, 83–90. [Google Scholar] [CrossRef]
- Cho, I.S.; Kwon, S.J.; Yoon, J.Y.; Chung, B.N.; Hammond, J.; Lim, H.S. First reported of apple necrotic mosaic virus infecting apple trees in Korea. J. Plant Pathol. 2017, 99, 815. [Google Scholar]
- Xing, F.; Zhang, Z.X.; Robe, B.L.; Li, S.F. Genomic analysis, sequence diversity, and occurrence of Apple necrotic mosaic virus, a novel ilarvirus associated with Mosaic disease of apple trees in China. Plant Dis. 2018, 102, 1841–1847. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.J.; Dong, Y.F.; Zhang, Z.P.; Fan, X.D.; Ren, F. Molecular characterization of Apple necrotic mosaic virus identified in crabapple (Malus spp.) tree of China. J. Integr. Agric. 2019, 18, 698–701. [Google Scholar] [CrossRef]
- Liang, P.B.; Zhang, Z.X.; Liu, F.; Lu, M.G.; Li, S.F.; Wang, H.Q. Problems of identification of pathogens associated with apple mosaic symptom and the exploration of its potential pathogens. J. Fruit Sci. 2016, 33, 332–339. [Google Scholar] [CrossRef]
- Hu, Y.; Shi, H.W.; Jing, C.C.; Li, K.; Sun, X.C.; Wu, G.T.; Zhou, C.Y.; Qing, L. First report of cucumber mosaic virus infecting apple in China. J. Plant Pathol. 2016, 98, 171–185. [Google Scholar]
- Phyre2. Available online: http://www.sbg.bio.ic.ac.uk/phyre2/html (accessed on 25 January 2020).
- Chamberlain, E.E.; Atkinson, J.D.; Hunter, J.A.; Wood, G.A. Effect of apple mosaic virus on growth and cropping of ‘freyberg’ apple trees. New Zealand J. Agric. Res. 1971, 14, 936–943. [Google Scholar] [CrossRef]
- Wood, G.A.; Chamberlain, E.E.; Atkinson, J.D.; Hunter, J.A. Field studies with apple mosaic virus. N. Z. J. Agric. Res. 1975, 18, 399–404. [Google Scholar] [CrossRef]
- Li, D.H.; Zhao, H.Y.; Hu, Z.Q.; Hu, X.S.; Zhang, Y.H. Studies on the damage, loss of production and control to Apple mosaic disease. J. Northwest Sci.-Tech. Univ. Agric. For. (Nat. Sci. Ed.) 2002, 30, 77–80. [Google Scholar] [CrossRef]
- Ji, P.; Wang, L.C.; Kong, B.H.; Wang, K.; Cao, K.Q.; Ma, J.; Yang, K.; Wang, A.X.; Ma, X.L. Identification of Apple mosaic virus (ApMV) in Zhaotong Yunnan. J. Yunnan Agric. Univ. (Nat. Sci.) 2013, 28, 180–185. [Google Scholar]
- Ji, Z.; Zhao, X.; Duan, H.; Hu, T.; Wang, S.; Wang, Y.; Cao, K. Multiplex RT-PCR detection and distribution of four apple viruses in China. Acta Virol. 2013, 57, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.R.; Zhao, X.S.; Wang, S.T.; Hu, T.L.; Wang, Y.N.; Cao, K.Q. The RT-PCR detection and distribution of ApMV in apple producing area of China. Acta Phytophylacica Sin. 2012, 39, 443–448. [Google Scholar]
- Mandadi, K.K.; Scholthof, K.B.G. Plant Immune Responses Against Viruses: How Does a Virus Cause Disease? Plant Cell 2013, 25, 1489–1505. [Google Scholar] [CrossRef]
- Shiel, P.J.; Berger, P.H. The complete nucleotide sequence of apple mosaic virus (ApMV) RNA 1 and RNA 2: ApMV is more closely related to alfalfa mosaic virus than to other ilarviruses. J. Gen. Virol. 2000, 81, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, T.; Ogata, Y.; Hirata, Y.; Ohki, S.T. Quantitative transcriptional changes associated with chlorosis severity in mosaic leaves of tobacco plants infected with Cucumber mosaic virus. Mol. Plant Pathol. 2014, 15, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Lehto, K.; Tikkanen, M.; Hiriart, J.B.; Paakkarinen, V.; Aro, E.M. Depletion of the photosystem II core complex in mature tobacco leaves infected by the flavum strain of Tobacco mosaic virus. Mol. Plant-Microbe Interact. 2003, 16, 1135–1144. [Google Scholar] [CrossRef]
- Feigelstock, D.A.; Tozzini, A.C.; Hopp, H.E. Coat protein sequence of a resistance-breaking strain of potato virus X isolated in Argentina. Virus Genes 1995, 10, 289–292. [Google Scholar] [CrossRef]
- Cordingley, H.; Grainger, J.; Pearsall, W.H.; Wright, A. The effect of mosaic disease upon certain metabolic products in the tobacco plant. Ann. Appl. Biol. 1934, 21, 78–89. [Google Scholar] [CrossRef]
- Hong, J.; Xu, Y.; Li, J.Y.; Zang, R.C.; Jiang, D.A. Effect of TuMV-infection on photosynthesis of host plants. J. Chin. Electron Microsc. Soc. 2002, 21, 110–113. [Google Scholar] [CrossRef]
- Mochizuki, T.; Ohki, S.T. Single amino acid substitutions at residue 129 in the coat protein of cucumber mosaic virus affect symptom expression and thylakoid structure. Arch. Virol. 2011, 156, 881–886. [Google Scholar] [CrossRef]
- Ohnishi, J.; Hirai, K.; Kanda, A.; Usugi, T.; Meshi, T.; Tsuda, S. The coat protein of Tomato mosaic virus L11Y is associated with virus-induced chlorosis on infected tobacco plants. J. Gen. Plant Pathol. 2009, 75, 297–306. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information Search database (NCBI). Available online: http://www.ncbi.nlm.nih.gov (accessed on 25 July 2019).






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W.; Yao, R.; Sunwu, R.; Huang, K.; Liu, Z.; Li, X.; Yang, Y.; Wang, J. Incidence and Molecular Identification of Apple Necrotic Mosaic Virus (ApNMV) in Southwest China. Plants 2020, 9, 415. https://doi.org/10.3390/plants9040415
Shi W, Yao R, Sunwu R, Huang K, Liu Z, Li X, Yang Y, Wang J. Incidence and Molecular Identification of Apple Necrotic Mosaic Virus (ApNMV) in Southwest China. Plants. 2020; 9(4):415. https://doi.org/10.3390/plants9040415
Chicago/Turabian StyleShi, Wensen, Rundong Yao, Runze Sunwu, Kui Huang, Zhibin Liu, Xufeng Li, Yi Yang, and Jianmei Wang. 2020. "Incidence and Molecular Identification of Apple Necrotic Mosaic Virus (ApNMV) in Southwest China" Plants 9, no. 4: 415. https://doi.org/10.3390/plants9040415
APA StyleShi, W., Yao, R., Sunwu, R., Huang, K., Liu, Z., Li, X., Yang, Y., & Wang, J. (2020). Incidence and Molecular Identification of Apple Necrotic Mosaic Virus (ApNMV) in Southwest China. Plants, 9(4), 415. https://doi.org/10.3390/plants9040415