Genotypic Variation in Resistance Gene-Mediated Calcium Signaling and Hormonal Signaling Involved in Effector-Triggered Immunity or Disease Susceptibility in the Xanthomonas campestris pv. Campestris–Brassica napus Pathosystem





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
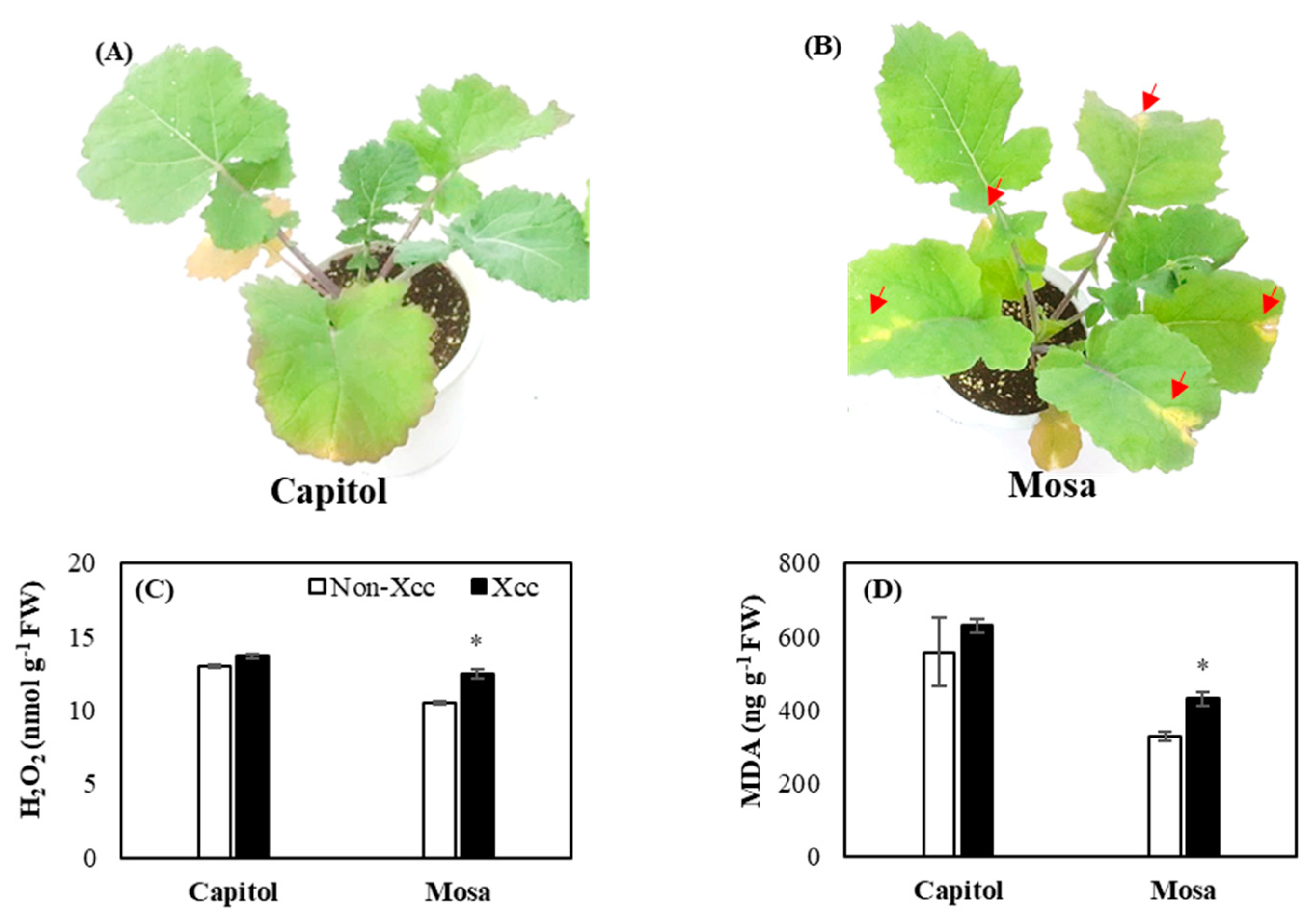
2.1. Development of Symptoms and Oxidative Stress
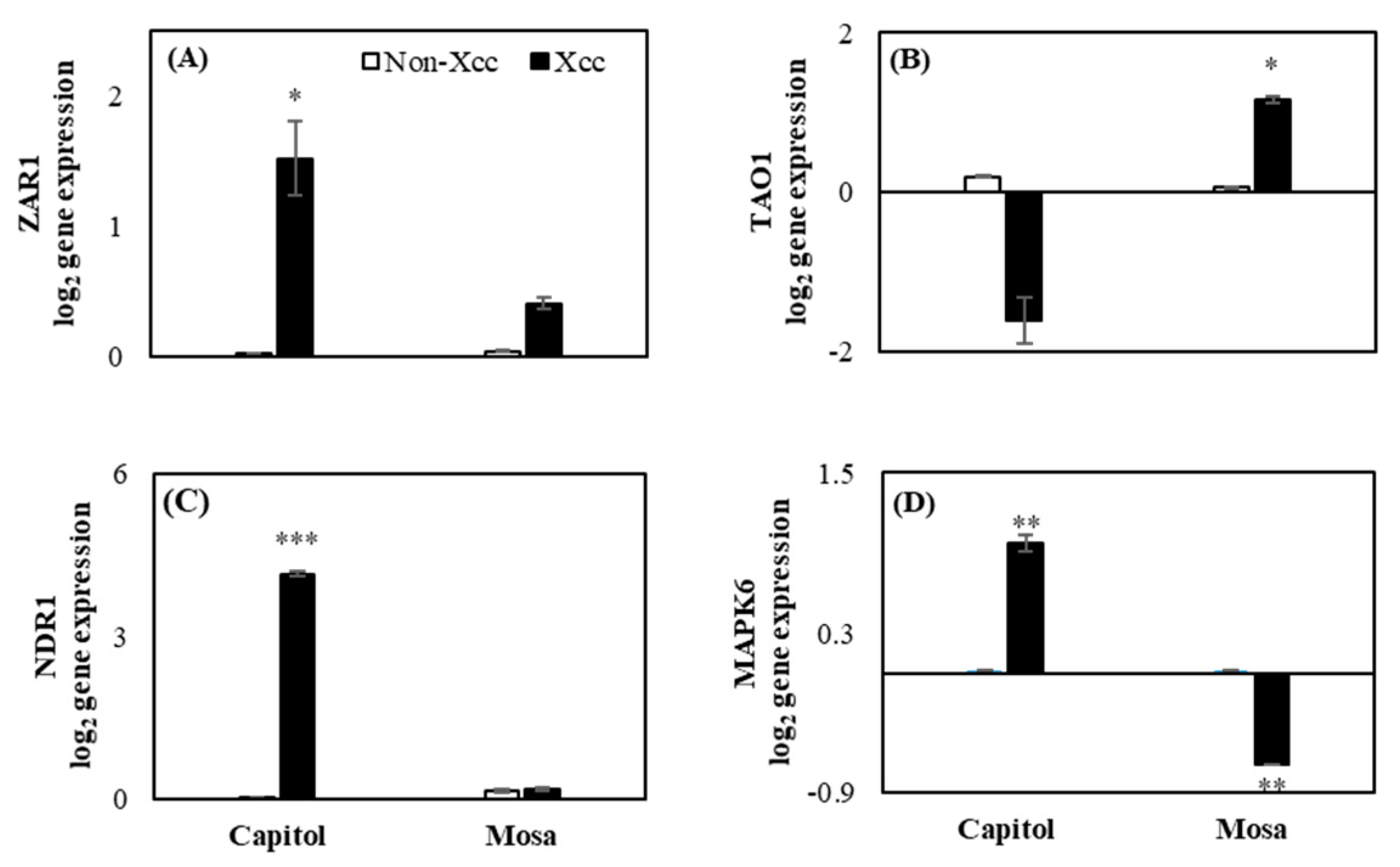
2.2. Expression of the Resistance Genes and Related Genes
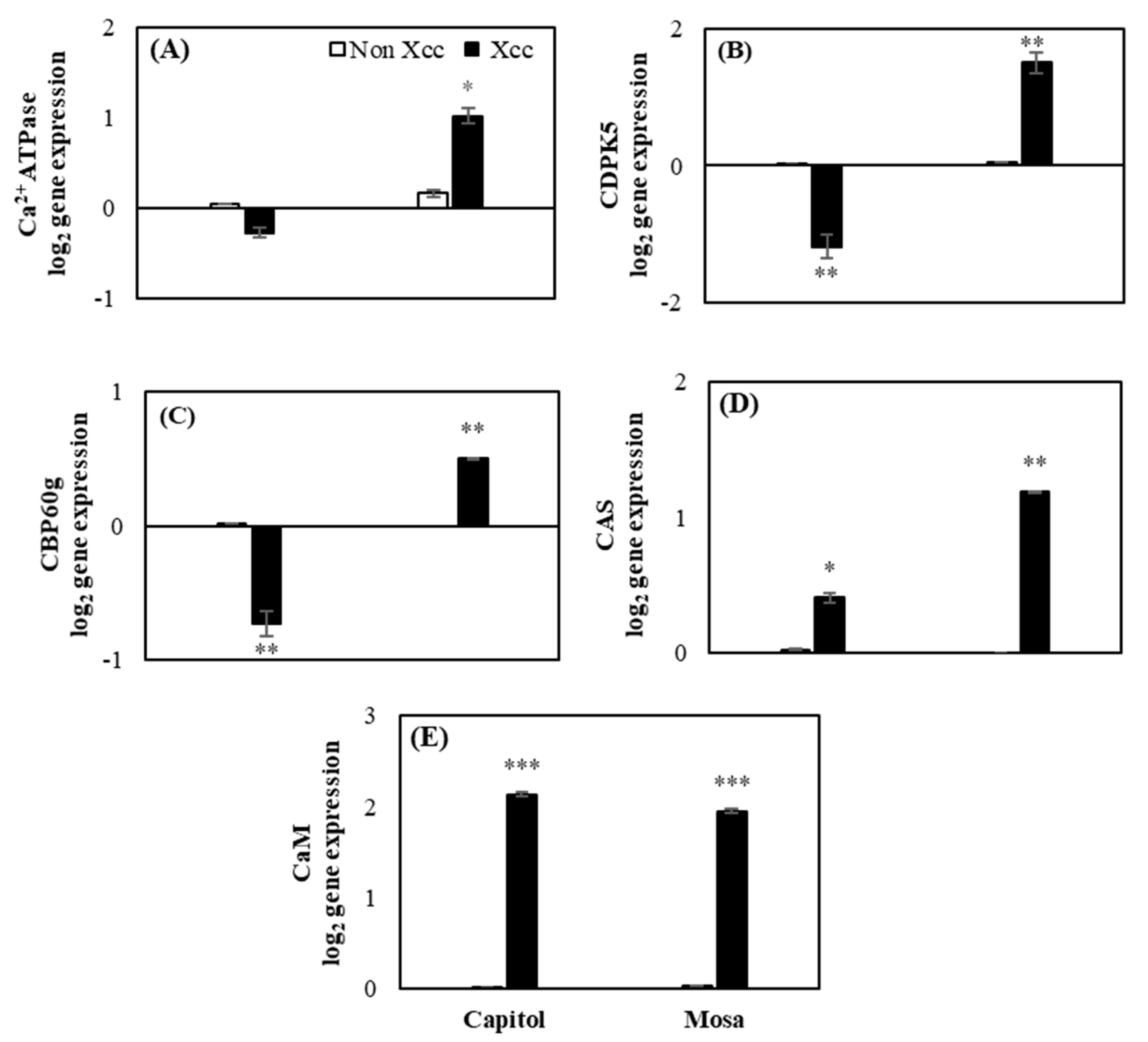
2.3. Expression of the Calcium Signaling-Related Genes
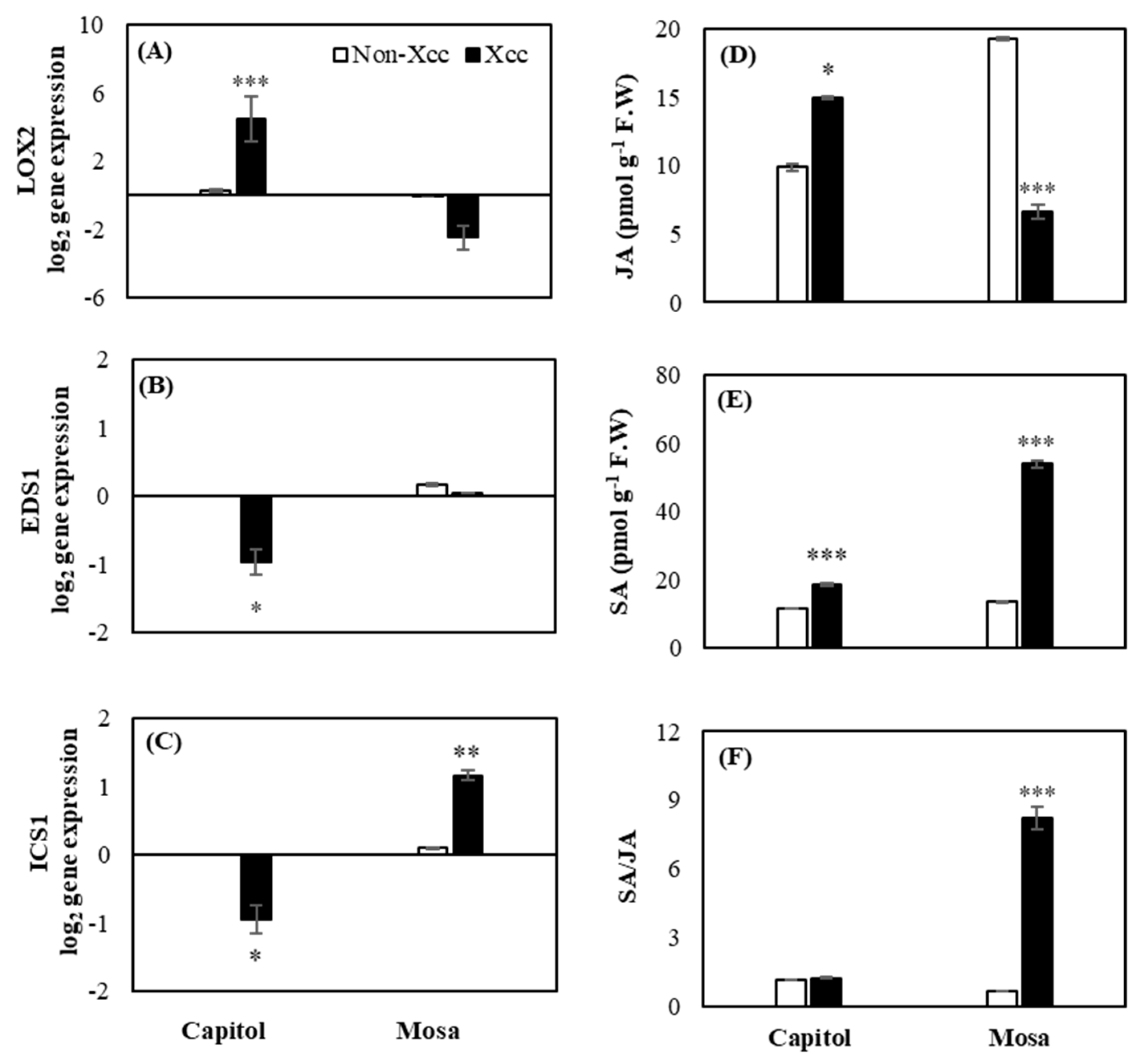
2.4. Expression of Hormonal Synthesis-Related Genes and Endogenous Content of SA and JA
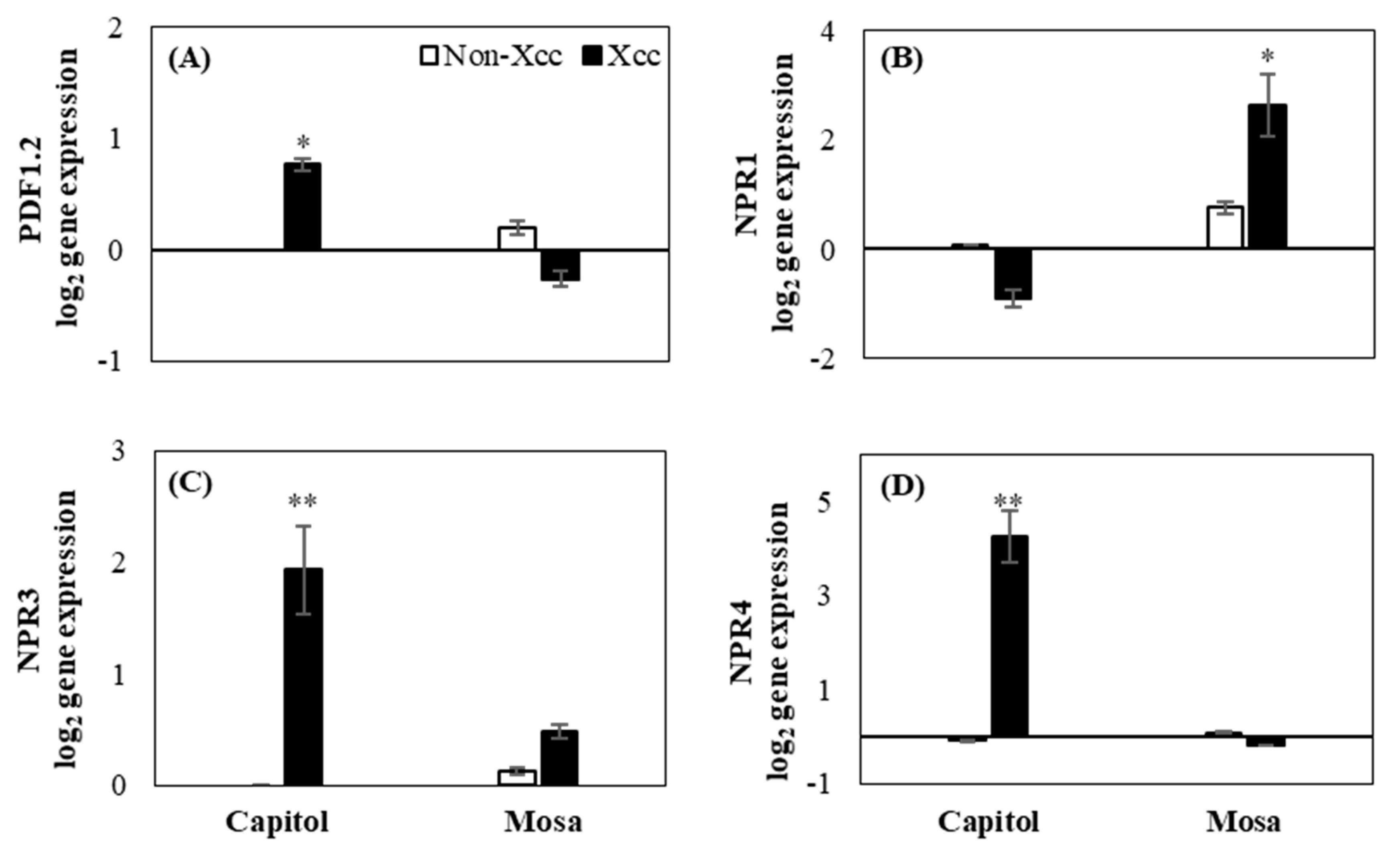
2.5. Expression of Hormonal Signaling-Related Genes
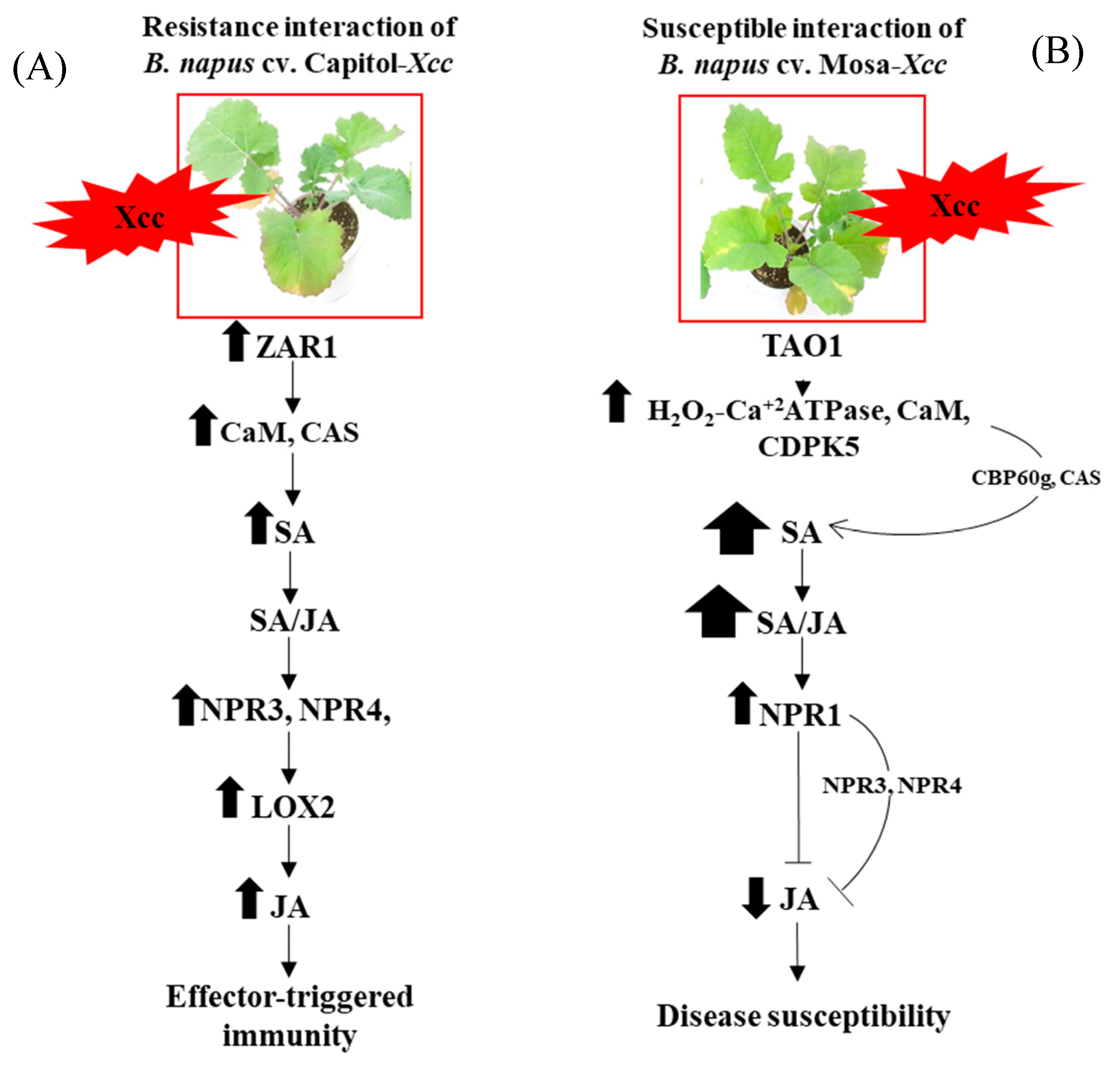
3. Discussion
4. Materials and Methods
4.1. Plant Material and Bacterial Inoculation
4.2. Determination of Hydrogen Peroxide (H2O2) and Lipid Peroxidation
4.3. Phytohormone Analysis
4.4. Isolation of Total RNA and Quantitative Real-Time PCR
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jensen, B.D.; Massomo, S.M.S.; Swai, I.S.; Hockenhull, J.; Andersen, S.B. Field evaluation for resistance to the black rot pathogen Xanthomonas campestris pv. campestris in cabbage (Brassica oleracea). Eur. J. Plant Pathol. 2005, 113, 297–308. [Google Scholar] [CrossRef]
- Ignatov, A.; Kuginuki, Y.; Hida, K. Distribution and inheritance of race-specific resistance to Xanthomonas campestris pv. campestris in Brassica rapa and B. napus. J. Russ. Phytopathol. Soc. 2000, 1, 83–87. [Google Scholar]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.K.; Nayak, S. Functional characterization and signal transduction ability of nucleotide-binding site-leucine-rich repeat resistance genes in plants. Genet. Mol. Res. 2011, 10, 2637–2652. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.X.; Yu, M.; Zhang, N.; Zhou, Z.Q.; Xu, Q.T.; Mei, F.Z.; Qu, L.H. Reactive oxygen species regulate programmed cell death progress of endosperm in winter wheat (Triticum aestivum L.) under waterlogging. Protoplasma 2015, 253, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Lee, B.R.; Park, S.H.; La, V.H.; Bae, D.W.; Kim, T.H. Cultivar variation in hormonal balance is a significant determinant of disease susceptibility to Xanthomonas campestris pv. campestris in Brassica napus. Front. Plant Sci. 2017, 8, 2121. [Google Scholar] [CrossRef]
- Wang, L.; Tsuda, K.; Sato, M.; Cohen, J.D.; Katagiri, F.; Glazebrook, J. Arabidopsis CaM Binding Protein CBP60g Contributes to MAMP-Induced SA Accumulation and Is Involved in Disease Resistance against Pseudomonas syringae. PLoS Pathog. 2009, 5, e1000301. [Google Scholar] [CrossRef]
- Nomura, H.; Komori, T.; Uemura, S.; Kanda, Y.; Shimotani, K.; Nakai, K.; Furuichi, T.; Takebayashi, K.; Sugimoto, T.; Sano, S.; et al. Chloroplast-mediated activation of plant immune signalling in Arabidopsis. Nat. Commun. 2012, 3, 1–11. [Google Scholar] [CrossRef]
- Du, L.; Ali, G.S.; Simons, K.A.; Hou, J.; Yang, T.; Reddy, A.S.N.Y.; Poovaiah, B.W. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1157. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Does, D.V.D.; Zamioudis, C.; Reyes, A.L.; Wees, S.C.M.V. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sonbol, F.M.; Huot, B.; Gu, Y.; Withers, J.; Mwimba, M.; Yao, J.; He, S.Y.; Dong, X. Salicylic acid receptors activate jasmonic acid signaling through a non-canonical pathway to promote effector-triggered immunity. Nat. Commun. 2016, 7, 13099. [Google Scholar] [CrossRef] [PubMed]
- Rahman, T.A.E.; Oirdi, M.E.; Lamothe, R.G.; Bouarab, K. Necrotrophic pathogens use the salicylic acid signaling pathway to promote disease development in tomato. Mol. Plant Microbe Interact. 2012, 25, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Eitas, T.K.; Nimchuk, Z.L.; Dangl, J.L. Arabidopsis TAO1 is a TIR-NB-LRR protein that contributes to disease resistance induced by the Pseudomonas syringae effector AvrB. Proc. Natl. Acad. Sci. USA 2008, 105, 6475–6480. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Lee, B.T.; La, V.H.; Lee, H.; Jung, W.J.; Bae, D.W.; Kim, T.H. p-Coumaric acid induces jasmonic acid-mediated phenolic accumulation and resistance to black rot disease in Brassica napus. Physiol. Mol. Plant. Pathol. 2019, 106, 270–275. [Google Scholar] [CrossRef]
- Islam, M.T. Hormonal regulations in soluble and cell-wall bound phenolic accumulation in two cultivars of Brassica napus contrasting susceptibility to Xanthomonas campestris pv. campestris. Plant Sci. J. 2019, 285, 132–140. [Google Scholar] [CrossRef]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Bohinc, T.; Ban, S.G.; Ban, D.; Trdan, S. Glucosinolates in plant protection strategies: A review. Arch. Biol. Sci. 2012, 64, 821–828. [Google Scholar] [CrossRef]
- Tortosa, M.; Cartea, M.E.; Velasco, P.; Soengas, P.; Rodriguez, V.M. Calcium-signaling proteins mediate the plant transcriptomic response during a well-established Xanthomonas campestris pv. campestris infection. Hortic. Res. 2019, 6, 103. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Vernaldi, S.; Maekawa, T. Evolution and conservation of plant NLR functions. Front. Immunol. 2013, 4, 297. [Google Scholar] [CrossRef]
- Wang, G.; Roux, B.; Feng, F.; He, C.; Noel, L.D.; Zhou, J.M. The Decoy substrate of a pathogen effector and a pseudokinase Specify pathogen-induced modified-self recognition and immunity in plants. Cell Host Microbe 2015, 18, 285–295. [Google Scholar] [CrossRef]
- Poovaiah, B.W.; Du, L.; Wang, H.; Yang, T. Recent Advances in Calcium/Calmodulin-Mediated Signaling with an Emphasis on Plant-Microbe Interactions. Plant Physiol. 2013, 163, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Huda, K.M.D.; Banu, M.S.A.; Tuteja, R.; Tuteja, N. Global calcium transducer P-type Ca2+ATPases open new avenues for agriculture by regulating stress signaling. J. Exp. Bot. 2013, 64, 3099–3109. [Google Scholar] [CrossRef]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.P. Calcium Signalling in Plant Biotic Interactions. Int. J. Mol. Sci. 2018, 19, 665. [Google Scholar] [CrossRef] [PubMed]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, M.; Marczak, M.; Ludwikow, A. Mitogen-Activated Protein Kinase Cascades in Plant Hormone Signaling. Front. Plant Sci. 2018, 9, 1387. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.R.; Lin, Y.; Joe, A.; Guo, M.; Korneli, C.; Yang, H.; Wang, P.; Yu, M.; Cerny, R.L.; Staiger, D.; et al. Structure function analysis of an ADP ribosyltransferase type III effector and its RNA-binding target in plant immunity. J. Biol. Chem. 2011, 286, 43272–43281. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X. Salicylic acid: Biosynthesis, perception and contribution to plant immunity. Curr. Opin. Plant. Biol. 2019, 50, 29–36. [Google Scholar] [CrossRef]
- Adlung, N.; Prochaska, H.; Thieme, S.; Banik, A.; Blüher, D.; John, P.; Nagel, O.; Schulze, S.; Gantner, J.; Delker, C.; et al. Non-host resistance induced by the Xanthomonas Effector XopQ is wide spread within the genus nicotiana and functionally depends on EDS1. Front. Plant Sci. 2016, 7, 1796. [Google Scholar] [CrossRef]
- Venisse, J.S.; Gullner, G.; Brisset, M.N. Evidence for the involvement of an oxidative stress in the initiation of infection of pear by Erwinia amylovora. Plant Physiol. 2019, 125, 2164–2172. [Google Scholar] [CrossRef]
- Lee, B.L.; Kim, K.Y.; Jung, W.J.; Avice, J.C.; Ourry, A.; Kim, T.H. Peroxidases and lignification in relation to the intensity of water-deficit stress in white clover (Trifolium repens L.). J. Exp. Bot. 2007, 58, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high performance liquid chromatography–mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamun, M.A.; Islam, M.T.; Lee, B.-R.; La, V.H.; Bae, D.-W.; Kim, T.-H. Genotypic Variation in Resistance Gene-Mediated Calcium Signaling and Hormonal Signaling Involved in Effector-Triggered Immunity or Disease Susceptibility in the Xanthomonas campestris pv. Campestris–Brassica napus Pathosystem. Plants 2020, 9, 303. https://doi.org/10.3390/plants9030303
Mamun MA, Islam MT, Lee B-R, La VH, Bae D-W, Kim T-H. Genotypic Variation in Resistance Gene-Mediated Calcium Signaling and Hormonal Signaling Involved in Effector-Triggered Immunity or Disease Susceptibility in the Xanthomonas campestris pv. Campestris–Brassica napus Pathosystem. Plants. 2020; 9(3):303. https://doi.org/10.3390/plants9030303
Chicago/Turabian StyleMamun, Md. Al, Md. Tabibul Islam, Bok-Rye Lee, Van Hien La, Dong-Won Bae, and Tae-Hwan Kim. 2020. "Genotypic Variation in Resistance Gene-Mediated Calcium Signaling and Hormonal Signaling Involved in Effector-Triggered Immunity or Disease Susceptibility in the Xanthomonas campestris pv. Campestris–Brassica napus Pathosystem" Plants 9, no. 3: 303. https://doi.org/10.3390/plants9030303
APA StyleMamun, M. A., Islam, M. T., Lee, B.-R., La, V. H., Bae, D.-W., & Kim, T.-H. (2020). Genotypic Variation in Resistance Gene-Mediated Calcium Signaling and Hormonal Signaling Involved in Effector-Triggered Immunity or Disease Susceptibility in the Xanthomonas campestris pv. Campestris–Brassica napus Pathosystem. Plants, 9(3), 303. https://doi.org/10.3390/plants9030303