Community Structure, Diversity and Potential of Endophytic Bacteria in the Primitive New Zealand Medicinal Plant Pseudowintera colorata


Abstract
1. Introduction
2. Results
2.1. Culture Independent Analysis
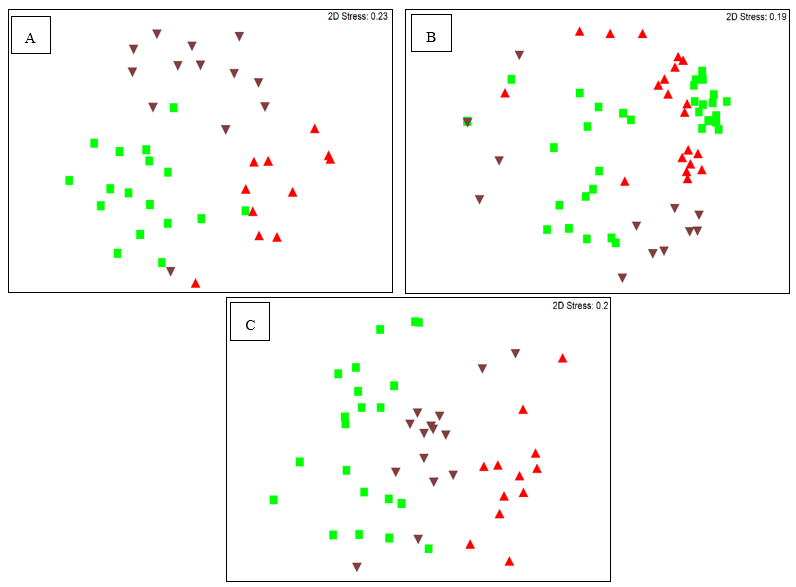
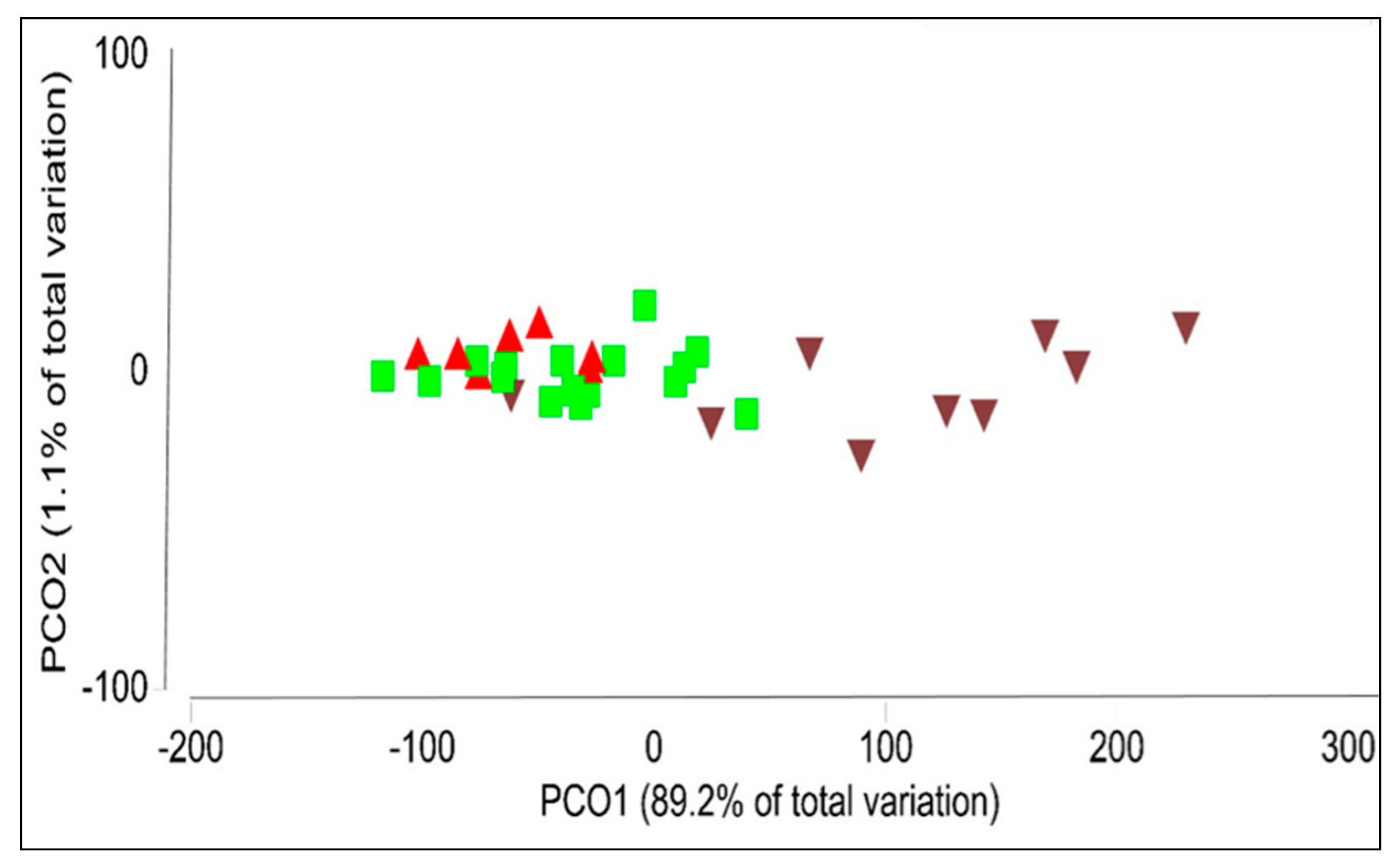
2.1.1. Analysis of the Bacterial Endomicrobiome using DGGE
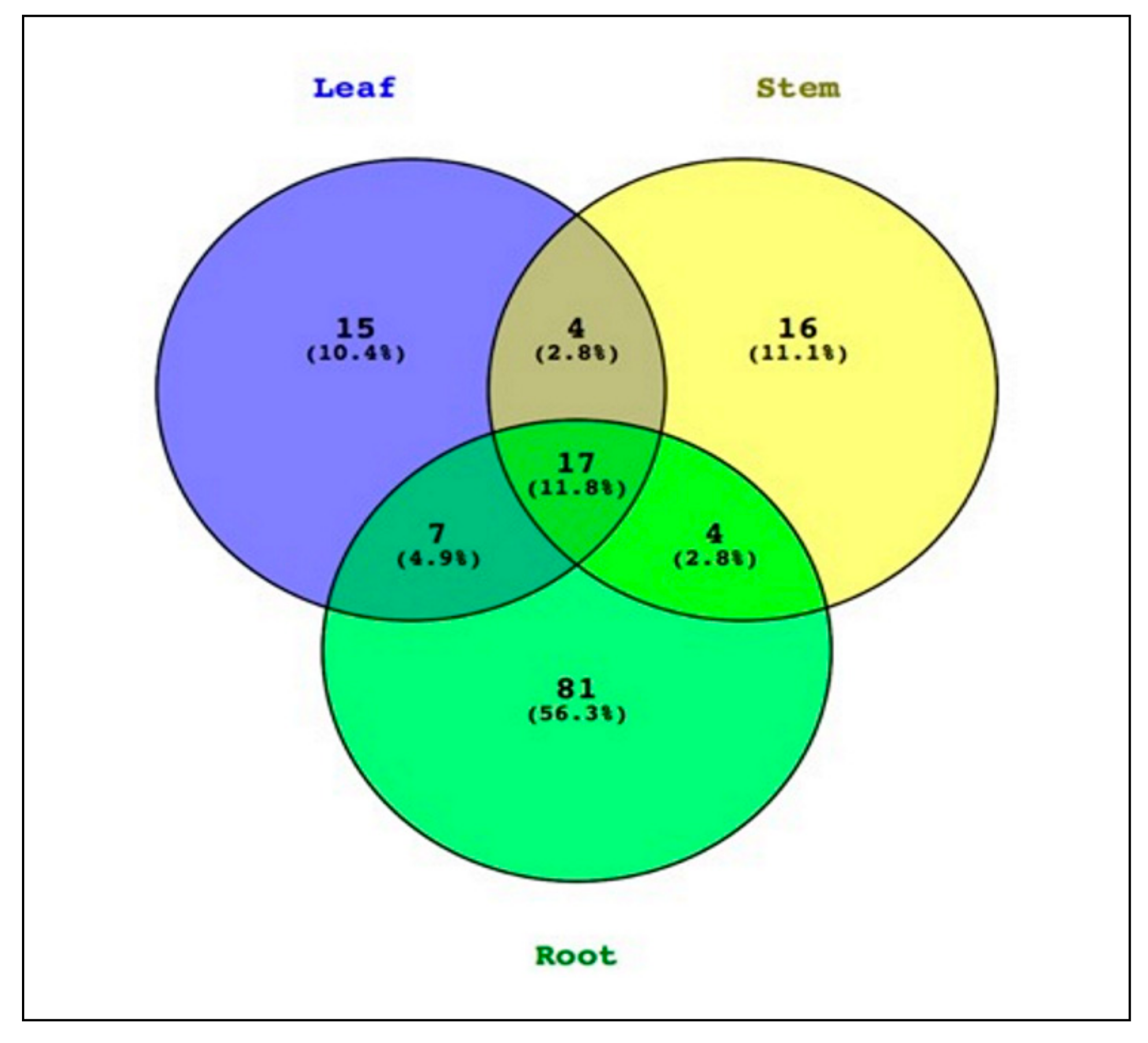
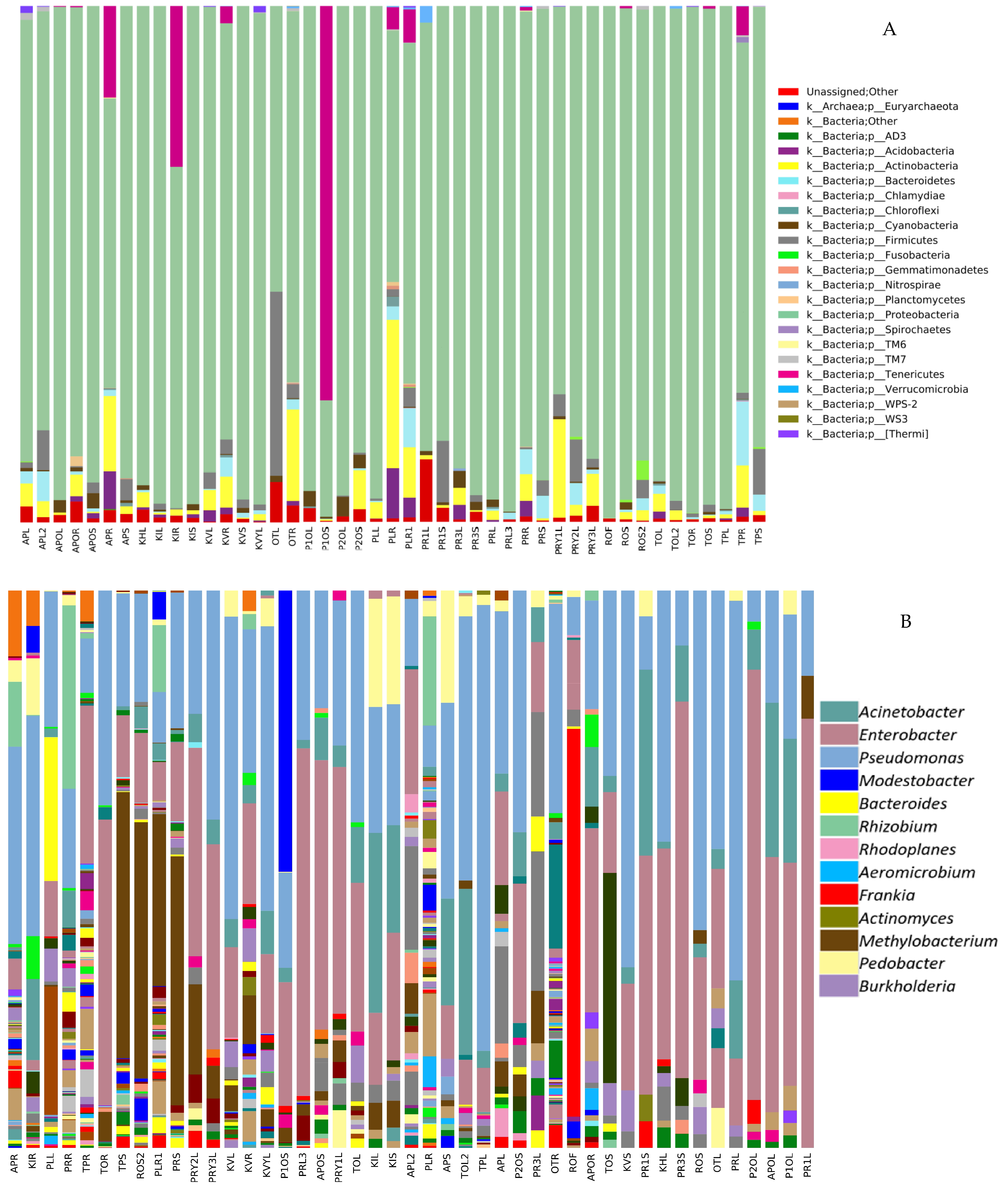
2.1.2. Analyzing the Structure of P. colorata Bacterial Endomicrobiome using Illumina MiSeq Metabarcoding
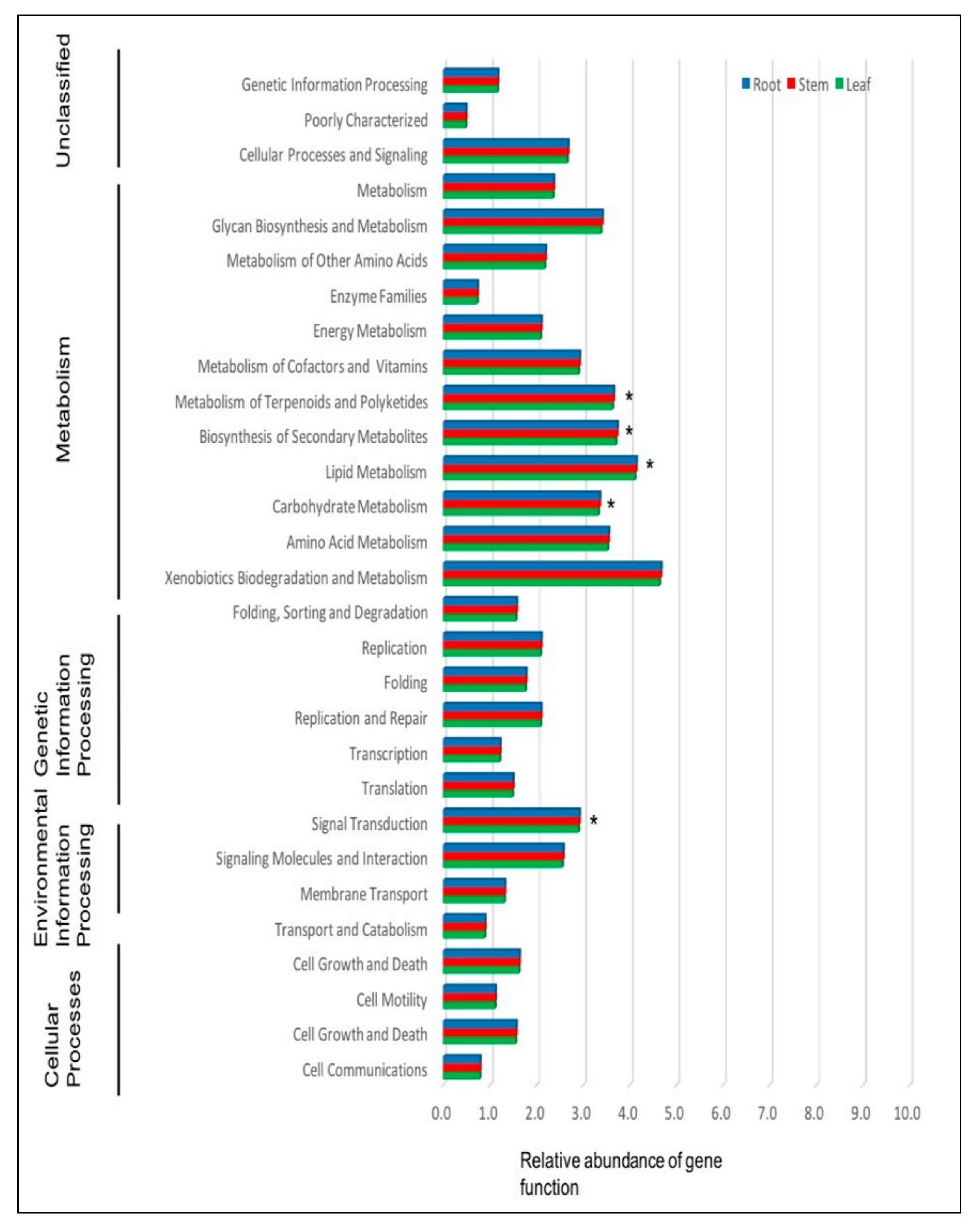
2.1.3. Prediction of the Function of Endophytic Bacteria in P. colorata Using PICRUSt
2.2. Culture Dependent Analysis
2.2.1. Recovery of Endophytic Bacteria from P. colorata
2.2.2. Activity of Endophytic Bacteria against Phytopathogenic Fungi
2.2.3. Identification of Bioactive Bacteria
2.2.4. Influence of Endophytic Bacterial Inoculants on P. colorata Seedlings
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Processing
4.2. Diversity Analysis of the Endophytic Bacteria in P. colorata Using DGGE
4.3. Illumina MiSeq Metabarcoding of Bacterial Endophytes of P. colorata
4.4. Functional Prediction of P. colorata Bacterial Endomicrobiome using PICRUSt
4.5. Biocontrol Activity against Phytopathogenic Fungi
4.6. Identification of Bioactive Bacteria by Sequencing the 16S rRNA Gene
4.7. Effect of Endophytic Bacterial Inoculants on P. colorata Seedlings
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Porras-Alfaro, A.; Bayman, P. Hidden Fungi, Emergent Properties: Endophytes and Microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Boil. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, S.; Garafola, C.; Monchy, S.; Newman, L.; Hoffman, A.; Weyens, N.; Barac, T.; Vangronsveld, J.; van der Lelie, D. Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees. Appl. Environ. Microbiol. 2009, 75, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Boil. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef]
- Castillo, U.F.; Strobel, G.A.; Ford, E.J.; Hess, W.M.; Porter, H.; Jensen, J.B.; Albert, H.; Robison, R.A.; Condron, M.A.M.; Teplow, D.; et al. Munumbicins, wide-spectrum antibiotics produced by Streptomyces NRRL 30562, endophytic on Kennedia nigriscans. FEMS Microbiol. Lett. 2003, 148, 2675–2685. [Google Scholar]
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural Products from Endophytic Microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-H.; Guo, J.-W.; Salam, N.; Li, L.; Zhang, Y.-G.; Han, J.; Mohamad, O.A.; Li, W.-J. Culturable endophytic bacteria associated with medicinal plant Ferula songorica: molecular phylogeny, distribution and screening for industrially important traits. 3 Biotech 2016, 6, 209. [Google Scholar] [CrossRef]
- Garcias-Bonet, N.; Arrieta, J.M.; De Santana, C.N.; Duarte, C.M.; Marbà, N. Endophytic bacterial community of a Mediterranean marine angiosperm (Posidonia oceanica). Front. Microbiol. 2012, 3, 342. [Google Scholar] [CrossRef]
- Da Silva, T.F.; Vollú, R.E.; Jurelevicius, D.; Alviano, D.S.; Alviano, C.S.; Blank, A.F.; Seldin, L. Does the essential oil of Lippia sidoides Cham. (pepper-rosmarin) affect its endophytic microbial community? BMC Microbiol. 2013, 13, 29. [Google Scholar] [CrossRef]
- Poole, I.; Francis, J.E. The First Record of Fossil Wood of Winteraceae from the Upper Cretaceous of Antarctica. Ann. Bot. 2000, 85, 307–315. [Google Scholar] [CrossRef]
- Brooker, S.G.; Cambie, R.C.; Cooper, R.C. New Zealand Medicinal Plants; Heinemann: Auckland, New Zealand, 1987; p. 240. [Google Scholar]
- McCallion, R.F.; Cole, A.L.J.; Walker, J.R.L.; Blunt, J.W.; Munro, M.H.G. Antibiotic substances from New Zealand plants. Planta Med. 1982, 44, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Fujita, K.; Lee, S.H. Antifungal mechanism of polygodial. J. Agric. Food Chem. 2001, 49, 1607–1611. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Fujita, K.; Lee, S.H.; Ha, T.J. Antibacterial activity of Polygodial. Phytother. Res. 2005, 19, 1013–1017. [Google Scholar] [CrossRef]
- Gerard, P.J.; Perry, N.B.; Ruf, L.D.; Foster, L.M. Antifeedant and insecticidal activity of compounds from Pseudowintera colorata (Winteraceae) on the webbing clothes moth, Tineola bisselliella (Lepidoptera: Tineidae) and the Australian carpet beetle, Anthrenocerus australis (Coleoptera: Dermestidae). Bull. Entomol. Res. 1993, 83, 547–552. [Google Scholar] [CrossRef]
- Dinsdale, E.A.; Edwards, R.A.; Hall, D.; Angly, F.; Breitbart, M.; Brulc, J.M.; Furlan, M.; Desnues, C.; Haynes, M.; Li, L.; et al. Functional metagenomics profiling of nine biomes. Nature 2008, 452, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, W.; Liu, L.; Yang, J. Determining microeukaryotic plankton community around Xiamen Island, southeast China, using Illumina MiSeq and PCR-DGGE techniques. PLoS ONE 2015, 10, e0127721. [Google Scholar] [CrossRef]
- Qin, Y.; Fu, Y.; Dong, C.; Jia, N.; Liu, H. Shifts of microbial communities of wheat (Triticum aestivum L.) cultivation in a closed artificial ecosystem. Appl. Microbiol. Biotechnol. 2016, 100, 4085–4095. [Google Scholar] [CrossRef]
- Akinsanya, M.A.; Goh, J.K.; Lim, S.P.; Tinga, A.S.Y. Metagenomics study of endophytic bacteria in Aloe vera using next-generation technology. Genomic Data 2015, 6, 159–163. [Google Scholar] [CrossRef]
- Jin, H.; Yang, X.Y.; Yan, Z.Q.; Liu, Q.; Li, X.Z.; Chen, J.X.; Qin, B. Characterization of rhizosphere and endophytic bacterial communities from leaves, stems and roots of medicinal Stellera chamaejasme L. Syst. Appl. Microbiol. 2014, 37, 376–385. [Google Scholar] [CrossRef]
- Wicaksono, W.A.; Jones, E.E.; Monk, J.; Ridgway, H.J. The Bacterial signature of Leptospermum scoparium (Mānuka) reveals core and accessory communities with bioactive properties. PLoS ONE 2016, 11, e0163717. [Google Scholar] [CrossRef]
- Long, H.H.; Schmidt, D.D.; Baldwin, I.T. Native bacterial endophytes promote host growth in a species-specific manner; phytohormone manipulations do not result in common growth responses. PLoS ONE 2008, 3, e2702. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, J.; Quadt-Hallmann, A.; Miller, W.G.; Sikora, R.A.; Lindow, S.E. Endophytic colonization of plants by the biocontrol agent Rhizobium etli G12 in relation to Meloidogyne incognita infection. Phytopathology 2001, 91, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; van Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellin, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, A.S.; Thijs, S.; Beckers, B.; Gonzalez-Chavez, C.M.; Weyens, N.; Carrillo-Gonzalez, R.; Vangronsveld, J. Community structure and diversity of endophytic bacteria in seeds of three consecutive generations of Crotalaria pumila growing on metal mine residues. Plant Soil 2017, 422, 1–16. [Google Scholar]
- Bragina, A.; Berg, C.; Cardinale, M.; Shcherbakov, A.; Chebotar, V.; Berg, G. Sphagnum mosses harbour highly specific bacterial diversity during their whole lifecycle. The ISME J. 2012, 6, 802–813. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Glavina del Rio, T.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef]
- Müller, H.; Berg, C.; Landa, B.B.; Auerbach, A.; Moissl-Eichinger, C.; Berg, G. Plant genotype-specific archaeal and bacterial endophytes but similar Bacillus antagonists colonize Mediterranean olive trees. Front. Microbiol. 2015, 6, 1–36. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; Bakker, P.A.H.M. Interactions between plants and beneficial Pseudomonas spp.: Exploiting bacterial traits for crop protection. Anton. Leeuw. 2007, 92, 367–389. [Google Scholar] [CrossRef]
- Mühling, M.; Woolven-Allen, J.; Murrell, J.C.; Joint, I. Improved group-specific PCR primers for denaturing gradient gel electrophoresis analysis of the genetic diversity of complex microbial communities. ISME J. 2008, 2, 379–392. [Google Scholar] [CrossRef]
- Dowd, S.E.; Sun, Y.; Secor, P.R.; Rhoads, D.D.; Wolcott, B.M.; James, G.A.; Wolcott, R.D. Survey of bacterial diversity in chronic wounds using pyrosequencing, DGGE, and full ribosome shotgun sequencing. BMC Microbiol. 2008, 8, 43. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Ver Loren van Themaat, E.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Ann. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed]
- Krause, A.; Ramakumar, A.; Bartels, D.; Battistoni, F.; Bekel, T.; Boch, J.; Böhm, M.; Friedrich, F.; Hurek, T.; Krause, L.; et al. Complete genome of the mutualistic, N2-fixing grass endophyte Azoarcus sp. strain BH72. Nature Biotechnol. 2006, 24, 1385–1391. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, A.; Donn, S.; Ryan, P.R.; Mathesius, U.; Devilla, R.; Jones, A.; Watt, M. Microbiome and exudates of the root and rhizosphere of Brachypodium distachyon, a model for wheat. PLoS ONE 2016, 11, e0164533. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.; Daisy, B. Bioprospecting for Microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Cooney, L.J.; Van Klink, J.W.; Hughes, N.M.; Perry, N.B.; Schaefer, H.M.; Menzies, I.J.; Gould, K.S. Red leaf margins indicate increased polygodial content and function as visual signals to reduce herbivory in Pseudowintera colorata. New Phytol. 2012, 194, 488–497. [Google Scholar] [CrossRef]
- Walterson, A.M.; Stavrinides, J. Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiol. Rev. 2015, 39, 968–984. [Google Scholar] [CrossRef]
- Kumar, A.; Guleria, S.; Mehta, P.; Walia, A.; Chauhan, A.; Shirkot, C.K. Plant growth-promoting traits of phosphate solubilizing bacteria isolated from Hippophae rhamnoides L. (Sea-buckthorn) growing in cold desert Trans-Himalayan Lahul and Spiti regions of India. Acta Physiol. Plant. 2015, 37, 1–12. [Google Scholar] [CrossRef]
- Lee, L.H.; Zainal, N.; Azman, A.-S.; Eng, S.-K.; Goh, B.-H.; Yin, W.-F.; Ab Mutalib, N.-S.; Chan, K.-G. Diversity and antimicrobial activities of Actinobacteria isolated from tropical mangrove sediments in Malaysia. Sci. World J. 2014, 698178, 1–14. [Google Scholar]
- Carini, P.; Marsden, P.J.; Leff, J.W.; Morgan, E.E.; Strickland, M.S.; Fierer, N. Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. Nat. Microbiol. 2016, 2, 16242. [Google Scholar] [CrossRef]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef] [PubMed]
- Gomes, N.C.M.; Heuer, H.; Schönfeld, J.; Costa, R.; Mendonca-Hagler, L.; Smalla, K. Bacterial diversity of the rhizosphere of maize (Zea mays) grown in tropical soil studied by temperature gradient gel electrophoresis. Plant Soil 2001, 232, 167–180. [Google Scholar] [CrossRef]
- Purushotham, N.; Jones, E.; Monk, J.; Ridgway, H. Community structure of endophytic Actinobacteria in a native New Zealand medicinal plant Pseudowintera colorata and their influence on plant growth. Microb. Ecol. 2018, 1153–1159. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Neilan, B.A.; Stuart, J.L.; Goodman, A.E.; Cox, P.T.; Hawkins, P.R. Specific amplification and restriction polymorphisms of the cyanobacterial rRNA operon spacer region. Syst. Appl. Microbiol. 1997, 20, 612–621. [Google Scholar] [CrossRef]
- Qin, S.; Miao, Q.; Feng, W.-W.; Wang, Y.; Zhu, X.; Xing, K.; Jiang, J.-H. Biodiversity and plant growth promoting traits of culturable endophytic actinobacteria associated with Jatropha curcas L. growing in Panxi dry-hot valley soil. Appl. Soil Ecol. 2015, 93, 47–55. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Alphaproteobacteria | Betaproteobacteria | Gammaproteobacteria | |||
|---|---|---|---|---|---|---|
| A | B | A | B | A | B | |
| Location | 0.323 | 0.036 * | 0.149 | 0.756 | 0.312 | 0.204 |
| Plant tissue | 0.001 ** | <0.001 ** | 0.001 ** | <0.001 ** | 0.001 ** | <0.001 ** |
| Location vs plant tissue | 0.021 * | 0.253 | 0.001 ** | 0.057 | 0.100 | 0.164 |
| Phytopathogenic Fungi | ||||
|---|---|---|---|---|
| Isolate | Neofusicoccum luteum | Neofusicoccum parvum | Ilyonectria liriodendri | Neonectria ditissima |
| TP1LA1B | +++ | ++ | ++ | ++ |
| TP1LC1B | +++ | ++ | ++ | ++ |
| TOYPRB1R | +++ | ++ | ++ | ++ |
| KIP1SB1B | +++ | ++ | ++ | ++ |
| KRP1BA1 | +++ | +++ | ++ | ++ |
| AP1SA1 | +++ | +++ | - | - |
| KRP1BC1 | +++ | +++ | - | - |
| KRP1BB1 | +++ | +++ | ++ | ++ |
| KRP1BA2 | +++ | +++ | ++ | ++ |
| KVP1BC1 | +++ | ++ | - | - |
| Isolate | Tissue | Reference Strain (GenBank) | Query Cover (%) | Similarity (%) | Accession no. |
|---|---|---|---|---|---|
| TP1LA1B | Leaf | Bacillus amyloliquefaciens strain ML471 | 99 | 99 | KC692205 |
| TP1LC1B | Leaf | Bacillus subtilis strain Y5 | 100 | 99 | GQ148816 |
| TOYPRB1R | Root | Bacillus subtilis strain AU04 | 99 | 98 | MF590152 |
| KIP1SB1B | Stem | Bacillus sp. strain A3 | 99 | 99 | KU904495 |
| KRP1BA1 | Stem | Pseudomonas fluorescens strain 4G628 | 100 | 99 | KY939748 |
| KRP1BA2 | Stem | Pseudomonas fluorescens strain 4G628 | 100 | 99 | KY939748 |
| AP1SA1 | Stem | Pantoea sp. ATY73 | 100 | 98 | HQ219992 |
| KRP1BC1 | Stem | Pseudomonas sp. strain PCH123 | 99 | 98 | MF774109 |
| KRP1BB1 | Stem | Pseudomonas sp. ps10-15 | 98 | 98 | AY303256 |
| KVP1BC1 | Stem | Erwinia sp. strain ES1 | 99 | 98 | KY446019 |
| Endophytic Bacteria | Shoot Height (cm) | Dry Weight (g) | Number of Internodes | |
|---|---|---|---|---|
| Shoot | Root | |||
| Pantoea sp. AP1SA1 | 5.79 a 1 | 0.79 bc | 0.47 b | 6.7 ab |
| Bacillus sp. TP1LA1B | 5.70 a | 1.38 a | 0.69 a | 6.8 a |
| Control | 3.12 b | 0.76 b | 0.46 b | 3.7 c |
| P Value | <0.005 | <0.005 | <0.05 | <0.001 |
| LSD (5%) | 1.63 | 0.24 | 0.17 | 0.49 |
| Sampling Site | Latitude (°South) | Longitude (°East) | Region | North/South Island | Plant Maturity |
|---|---|---|---|---|---|
| Taihape Scenic Reserve | −39.67635 | 175.80560 | Manawatu-Wanganui | North Island | Mature |
| Tongariro National Park | −39.02237 | 175.71810 | Manawatu-Wanganui | North Island | Mature |
| Kaimanawa Forest Park | −38.94721 | 175.94370 | Manawatu-Wanganui | North Island | Mature |
| Lake Rotopounamu Scenic Reserve | −39.02656 | 175.73502 | Manawatu-Wanganui | North Island | Mature |
| Kahurangi National Park | −41.07224 | 172.59166 | Nelson/Tasman | South Island | Mature |
| Paringa Forest | −43.69379 | 169.40724 | West Coast | South Island | Mature and Immature |
| Arthur’s Pass National Park | −42.94215 | 171.56414 | Canterbury | South Island | Mature |
| Kaituna Valley Scenic Reserve | −43.71655 | 172.7554 | Canterbury | South Island | Mature and Immature |
| Peel Forest | −43.91835 | 171.25934 | Canterbury | South Island | Mature and Immature |
| Otago Peninsula Scenic Reserve | −45.88184 | 170.58049 | Otago | South Island | Mature |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purushotham, N.; Jones, E.; Monk, J.; Ridgway, H. Community Structure, Diversity and Potential of Endophytic Bacteria in the Primitive New Zealand Medicinal Plant Pseudowintera colorata. Plants 2020, 9, 156. https://doi.org/10.3390/plants9020156
Purushotham N, Jones E, Monk J, Ridgway H. Community Structure, Diversity and Potential of Endophytic Bacteria in the Primitive New Zealand Medicinal Plant Pseudowintera colorata. Plants. 2020; 9(2):156. https://doi.org/10.3390/plants9020156
Chicago/Turabian StylePurushotham, Neeraj, Eirian Jones, Jana Monk, and Hayley Ridgway. 2020. "Community Structure, Diversity and Potential of Endophytic Bacteria in the Primitive New Zealand Medicinal Plant Pseudowintera colorata" Plants 9, no. 2: 156. https://doi.org/10.3390/plants9020156
APA StylePurushotham, N., Jones, E., Monk, J., & Ridgway, H. (2020). Community Structure, Diversity and Potential of Endophytic Bacteria in the Primitive New Zealand Medicinal Plant Pseudowintera colorata. Plants, 9(2), 156. https://doi.org/10.3390/plants9020156