Genetic Diversity of Potyviruses Associated with Tulip Breaking Syndrome


Abstract
1. Introduction
2. Results
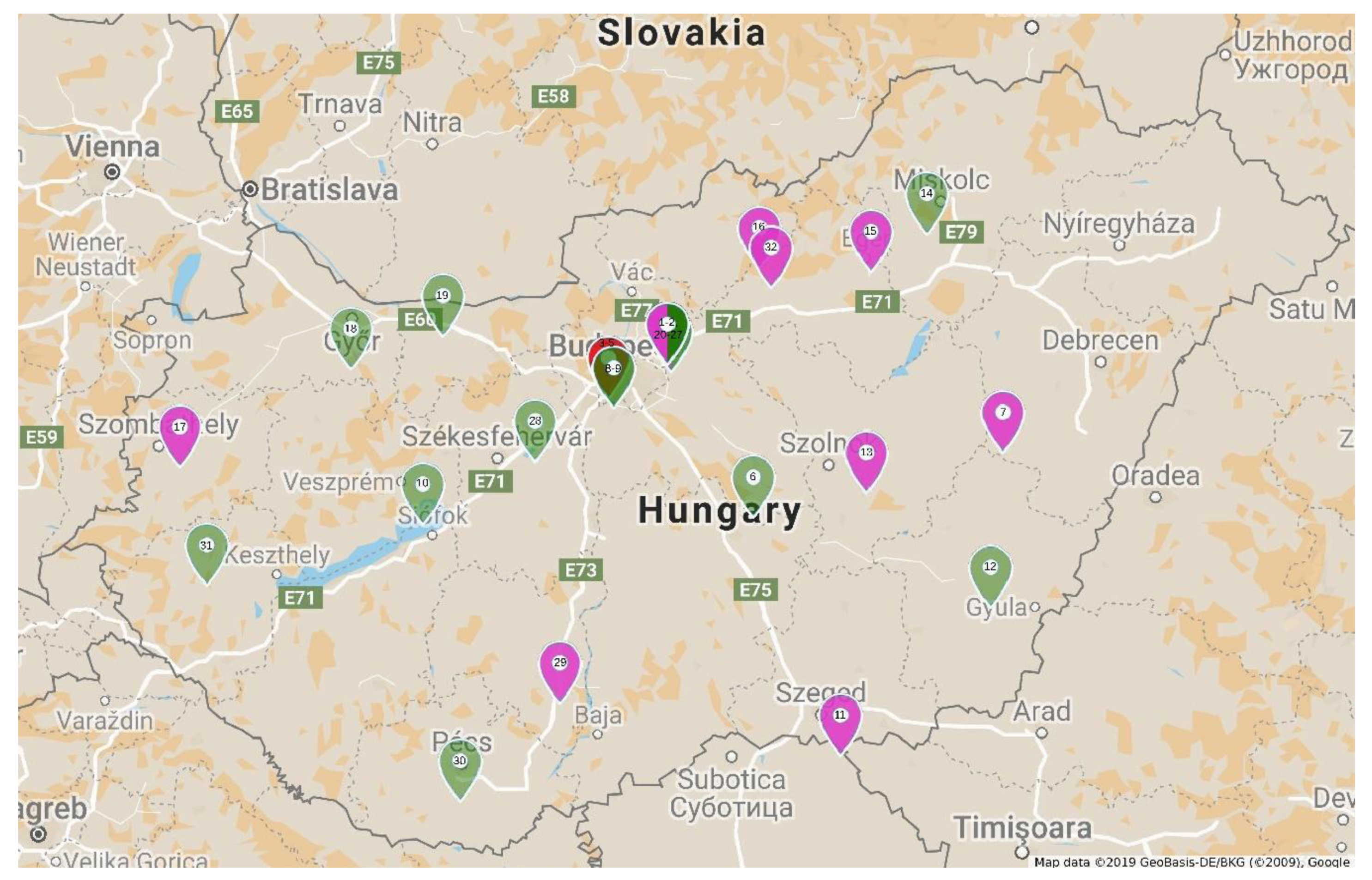
2.1. Detection and Identification of Potyviruses in Tulips
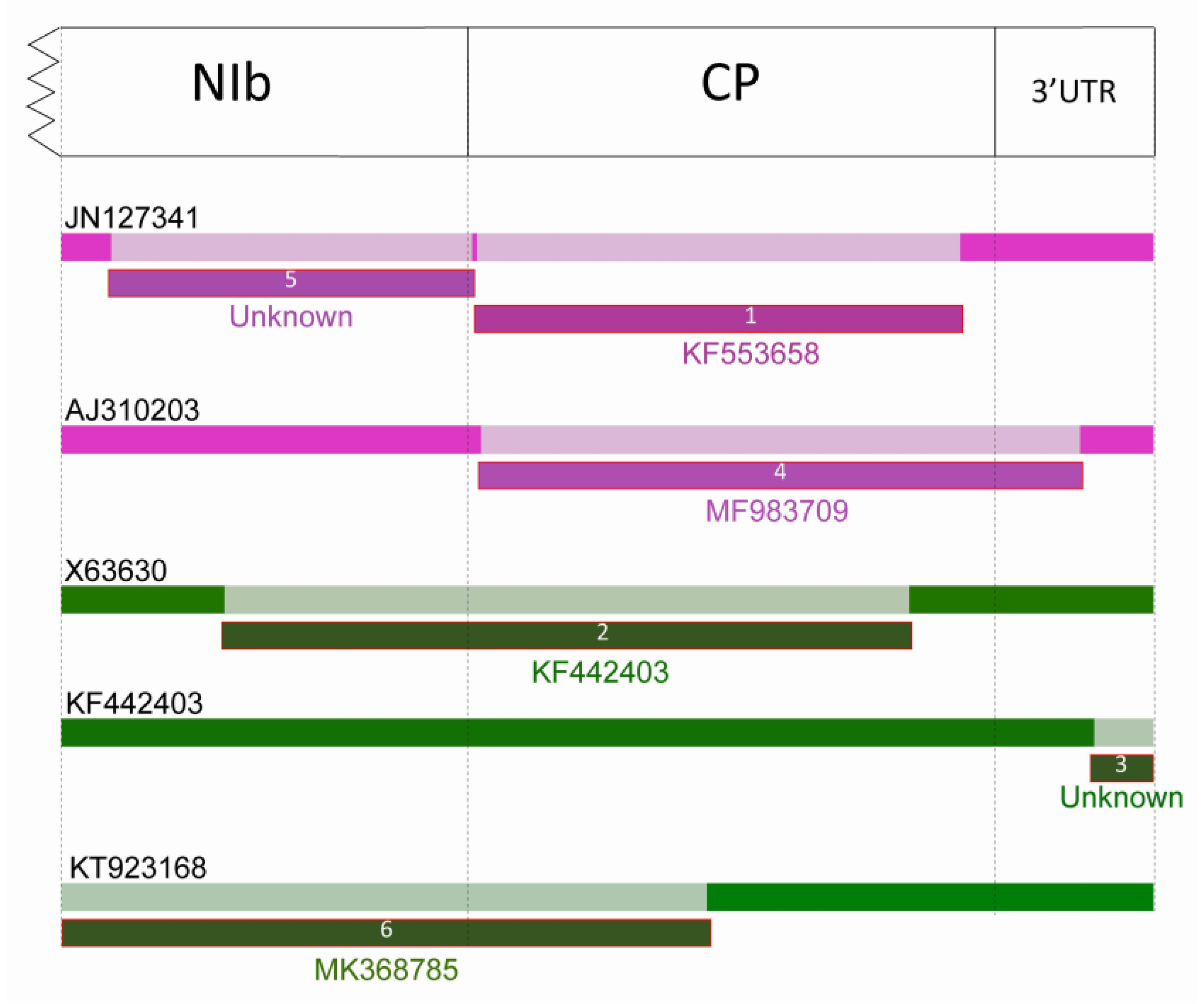
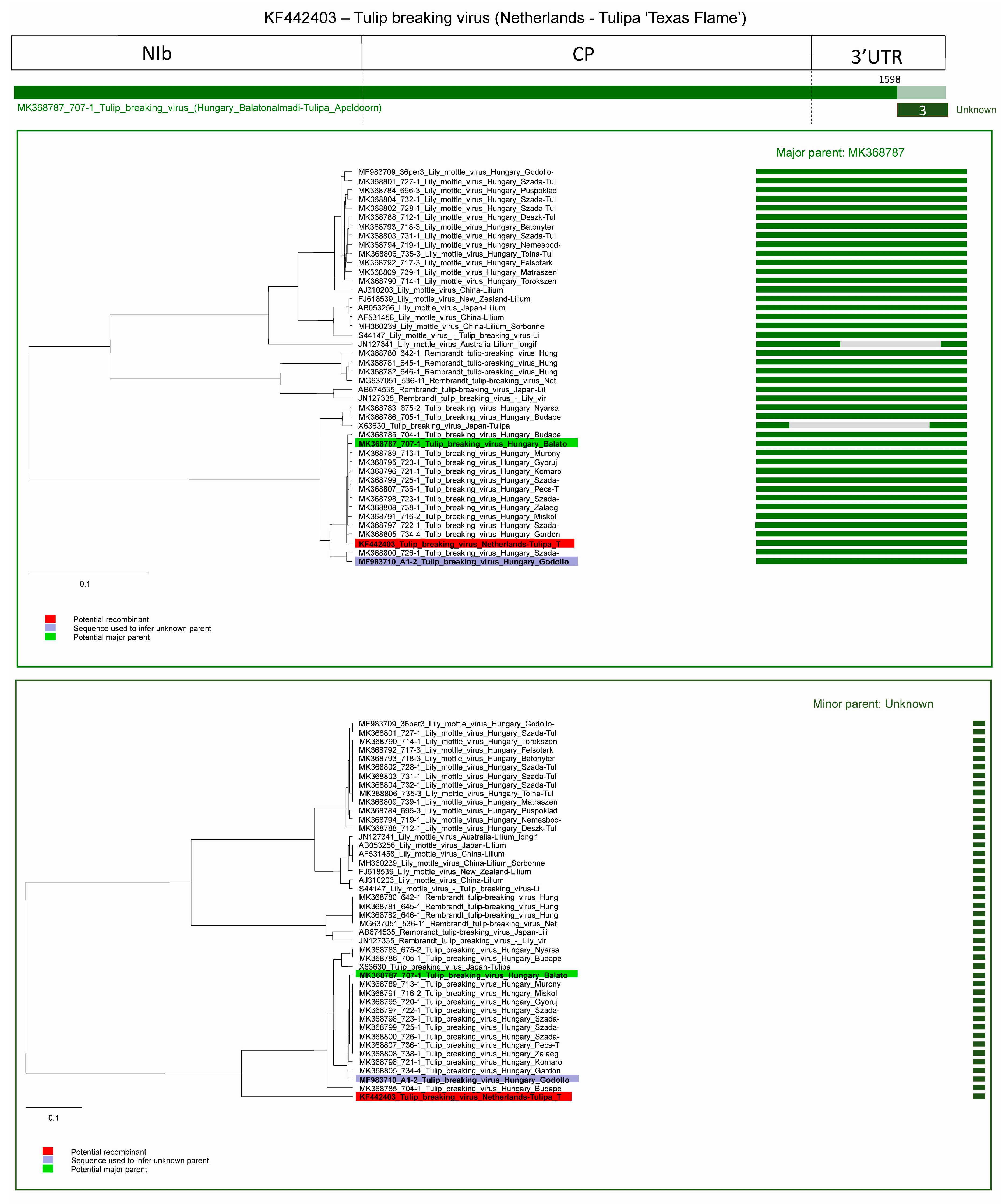
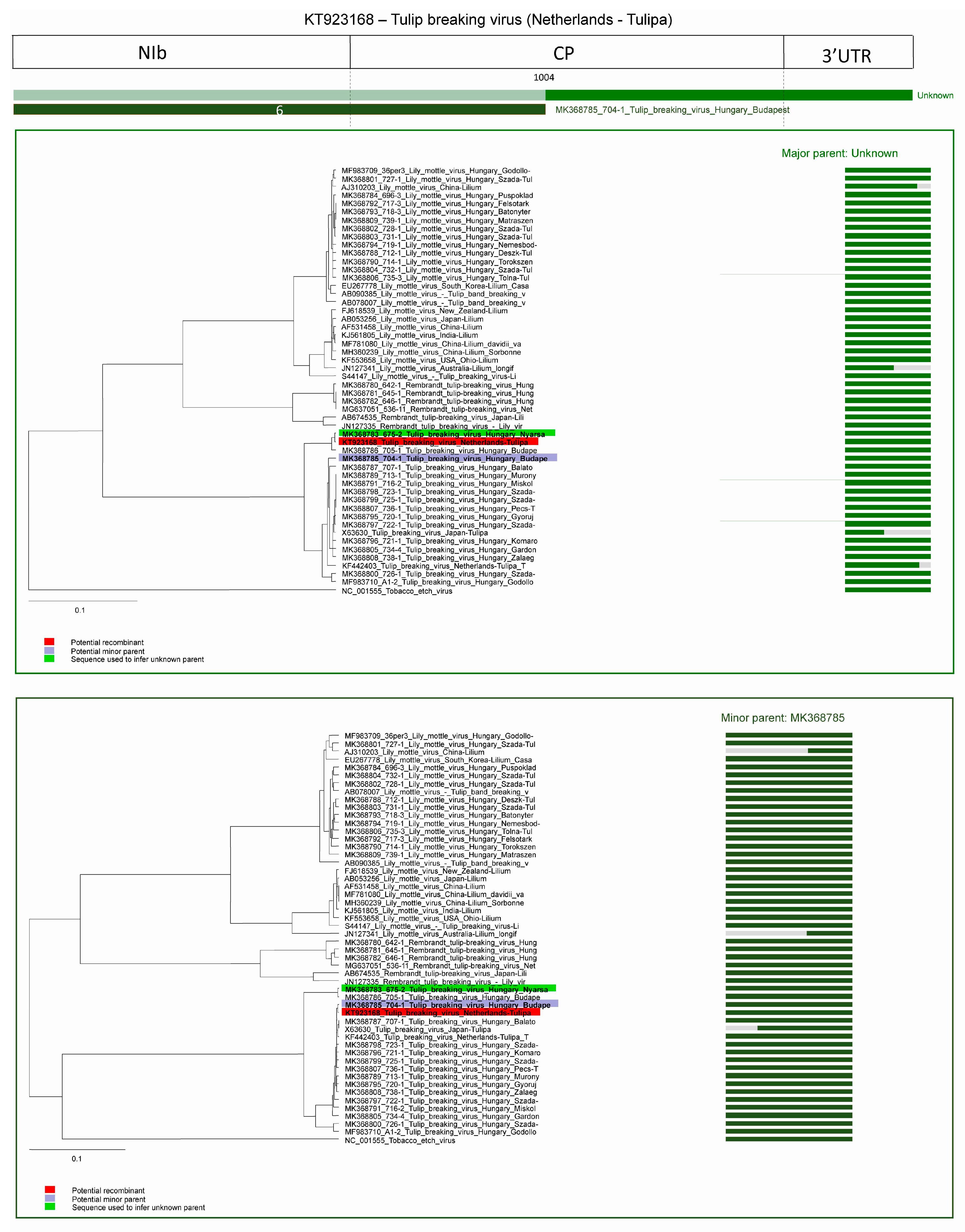
2.2. Recombination
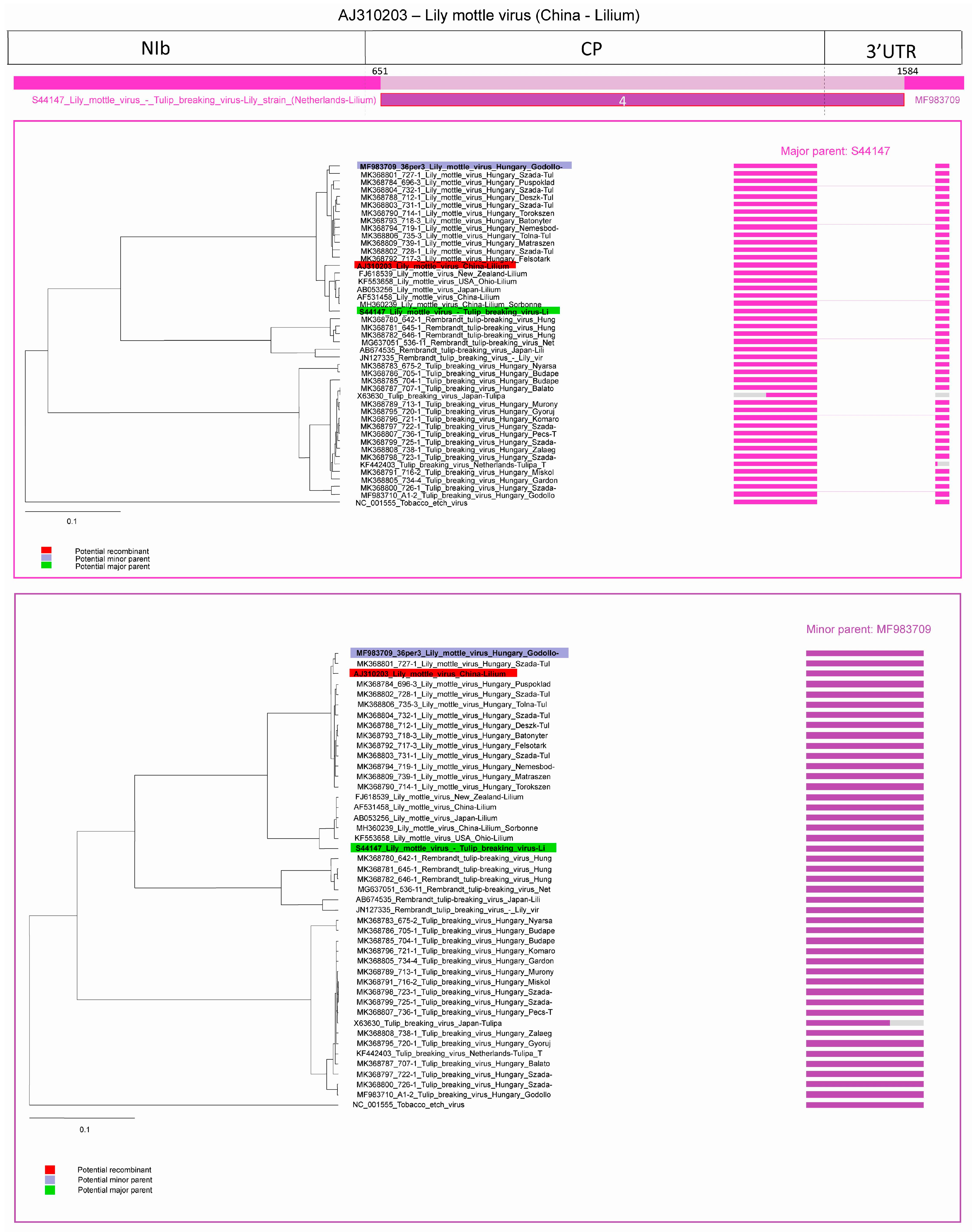
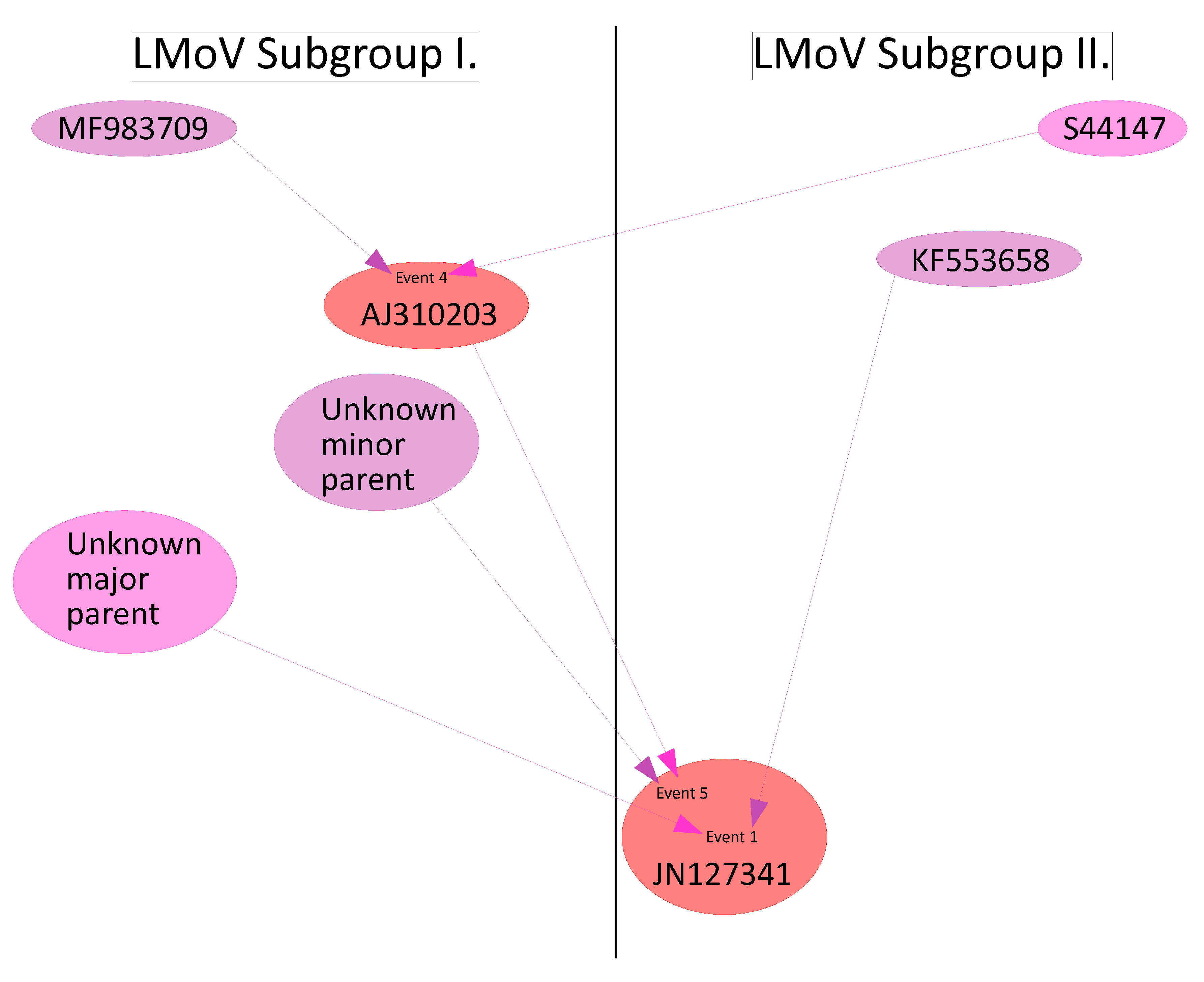
2.2.1. Recombination in LMoV Strains
2.2.2. Recombination in TBV Strains
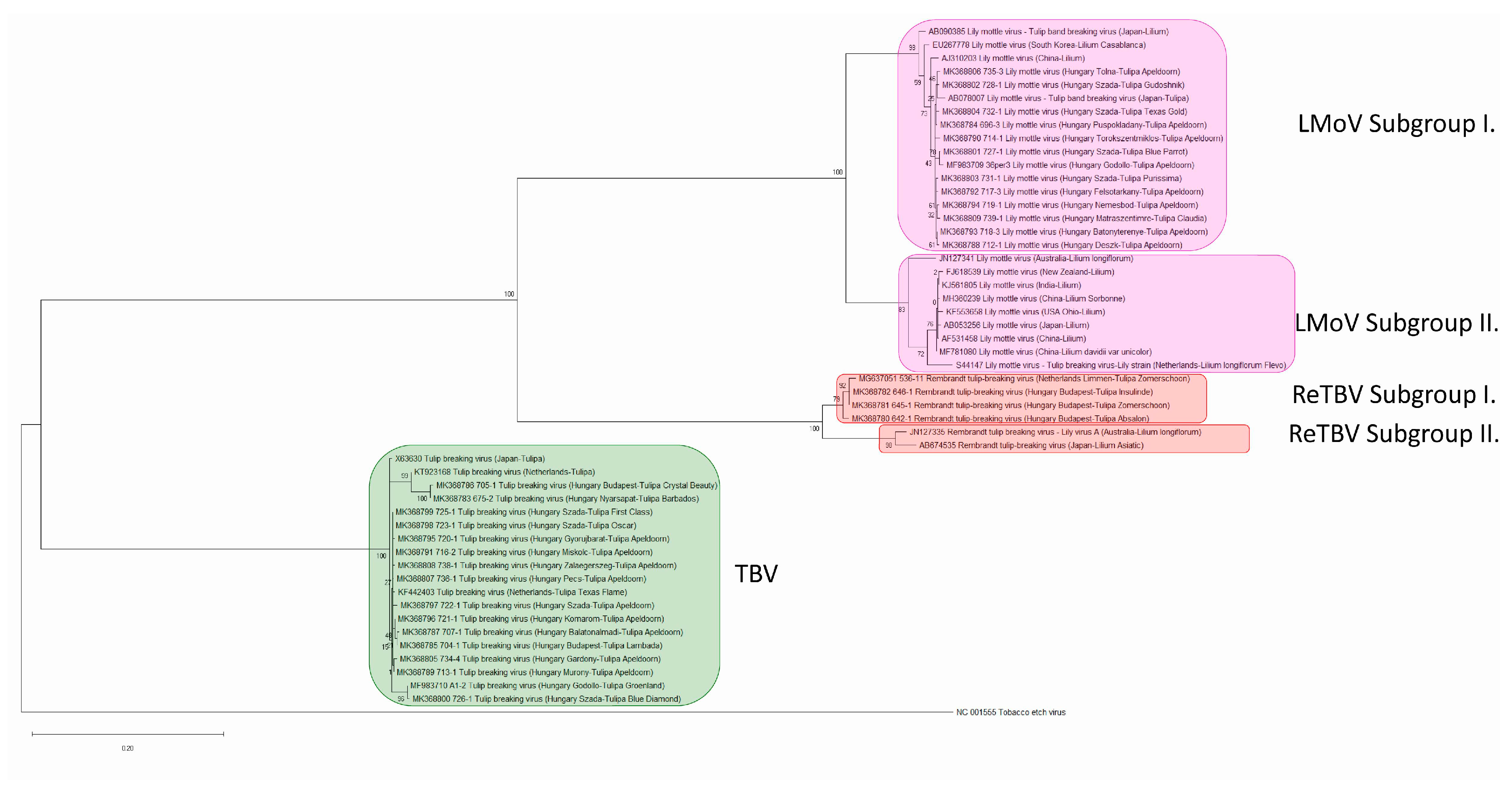
2.3. Phylogenetic Analysis of Tulip Infecting Potyviruses Based on the Coat Protein Gene
2.4. Pairwise Relationships, NIb/CP Polyprotein Cleavage Site, and Species According to the ICTV Taxonomy Criteria
3. Discussion
4. Materials and Methods
4.1. Plant Samples
4.2. Enzyme-Linked Immunosorbent Assay (ELISA)
4.3. RNA Extraction, Reverse Transcription, PCR Cloning and Nucleotide Sequence Determination
4.4. Sequence Analysis and Comparisons
4.5. Phylogenetic Analysis
4.6. Recombination Detection
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability Statement
Ethical Statement
References
- Clusius, C. Caroli Clusii Atrebat Rariorum Alioquot Stirpium per Hispanias Observatarum Historia, Libris Duobus Expressas ad Maximilianum II. Imperatorem; Ex officina Christophori Plantini, Architypographi Regij: Antverpiae, UK, 1576. [Google Scholar]
- Bryan, J.E. Bulbs, Rev. ed.; Timber Press: Portland, OR, USA, 2002; ISBN 978-0-88192-529-6. [Google Scholar]
- Jelitto, L.; Schacht, W.; Epp, M.E.; Baumgardt, J.P.; Fessler, A. Hardy Herbaceous Perennials; Timber Press: Portland, OR, USA, 1995; ISBN 978-0-88192-159-5. [Google Scholar]
- WCSP. World Checklist of Selected Plant Families; Royal Botanic Gardens: Kew, UK, 2020. [Google Scholar]
- Koninklijke Algemeene Vereeniging Voor Bloembollencultuur (KAVB). Ornamental Plant Database; Koninklijke Algemeene Vereeniging Voor Bloembollencultuur (KAVB): Hillegom, The Netherlands, 2020. [Google Scholar]
- Bryan, J.E. Timber Press Pocket Guide TO Bulbs; Timber Press: Portland, OR, USA, 2005; ISBN 978-0-88192-725-2. [Google Scholar]
- Krabbendam, P.; Baardse, A.A. Bloembollenteelt/2, De tulp. In Leidraad Voor Het Land- en Tuinbouwonderwijs, 15th ed.; 36. Serie B; Tjeenk Willink: Zwolle, The Netherlands, 1966. [Google Scholar]
- Hop, M. De Veredeling Van Bloembollen; Stichting Bollenacademie: Hillegom, The Netherlands, 2018; ISBN 8717247013442. [Google Scholar]
- De Hertogh, A.A.; le Nard, M. Tulipa. In The Physiology of Flower Bulbs: A Comprehensive Treatise on the Physiology and Utilization of Ornamental Flowering Bulbous and Tuberous Plants; de Hertogh, A.A., le Nard, M., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 617–682. ISBN 978-0-444-87498-6. [Google Scholar]
- Baardse, A.A.; Krabbendam, P. Eenvoudig Leerboek Voor de Bloembollenteelt: Algemeen Gedeelte, Tulp, Hyacint, Narcis, Gladiool en Boliris. In Leidraad Voor Het Land- en Tuinbouwonderwijs, 2nd ed.; Tjeenk Willink: Zwolle, The Netherlands, 1967. [Google Scholar]
- Bos, L. Viruses of Ornamentals: Historical Perspectives. In Virus and Virus like Diseases of Bulb and Flower Crops; Loebenstein, G., Lawson, R.H., Brunt, A.A., Eds.; Wiley: Chichester, UK, 1995; pp. 15–22. ISBN 978-0-471-95293-0. [Google Scholar]
- Valverde, R.A.; Sabanadzovic, S.; Hammond, J. Viruses that enhance the aesthetics of some ornamental plants: Beauty or beast? Plant Dis. 2012, 96, 600–611. [Google Scholar] [CrossRef] [PubMed]
- De Best, A.L.I.C.; Zwart, M.J.; van Aartrijk, J.; van den Ende, J.E.; Peeters, J.M.M. Ziekten en afwijkingen bij bolgewassen: Liliaceae; Laboratorium voor Bloembollenonderzoek: Lisse, The Netherlands, 2000; Volume 1. [Google Scholar]
- Dekker, E.L.; Derks, A.F.L.M.; Asjes, C.J.; Lemmers, M.E.C.; Bol, J.F.; Langeveld, S.A. Characterization of potyviruses from tulip and lily which cause flower-breaking. J. Gen. Virol. 1993, 74, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Bloembollenkeuringsdienst (BKD). Informatie TBV. Available online: https://www.bkd.eu/wp-content/uploads/2019/04/tbv-beelden-tulp.pdf (accessed on 5 January 2020).
- Némethy, Z. Survey on virus diseases of bulbous flowers in Hungary. Acta Hortic. 1994, 377, 267–274. [Google Scholar] [CrossRef]
- Gleason, M.L.; Daughtrey, M.L.; Chase, A.R.; Moorman, G.W.; Mueller, D.S. Diseases of Herbaceous Perennials; APS Press: St. Paul, MN, USA, 2009; ISBN 978-0-89054-374-0. [Google Scholar]
- Brunt, A.A. (Ed.) Viruses of Plants: Descriptions and Lists from the VIDE Database; CAB International: Wallingford, UK, 1996; ISBN 978-0-85198-794-1. [Google Scholar]
- Van Regenmortel, M.H.V.; Fauquet, C.M.; Bishop, D.H.L.; Carstens, E.B.; Estes, M.K.; Lemon, S.M.; Maniloff, J.; Mayo, M.A.; McGeoch, D.J.; Pringle, C.R.; et al. (Eds.) Virus Taxonomy: Seventh Report of the International Committee on Taxonomy of Viruses; Acad. Press: San Diego, CA, USA, 2000; ISBN 978-0-12-370200-5. [Google Scholar]
- ICTV Report Consortium Fast Track Proposals from the Potyviridae SG; International Committee on Taxonomy of Viruses: London, UK, 2003; pp. 16–29.
- King, A.M.Q.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; Knowles, N.J.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2018). Arch. Virol. 2018, 163, 2601–2631. [Google Scholar] [CrossRef]
- Horst, R.K. Westcott’s Plant Disease Handbook; Springer: Dordrecht, The Netherlands, 2013; ISBN 978-94-007-2140-1. [Google Scholar]
- Mowat, W.P. Tulip. In Virus and Virus like Diseases of Bulb and Flower Crops; Loebenstein, G., Lawson, R.H., Brunt, A.A., Eds.; Wiley: Chichester, UK, 1995; pp. 352–383. ISBN 978-0-471-95293-0. [Google Scholar]
- Fenner, F. (Ed.) Classification and Nomenclature of Viruses: Second Report of the International Committee on Taxonomy of Viruses; S. Karger AG: Basel, Switzerland, 1976; ISBN 978-3-8055-2418-6. [Google Scholar]
- Ohira, K.; Namba, S.; Miyagawa, M.; Kusumi, T.; Tsuchiaki, T. Nucleotide-sequence of the coat protein-coding region of Tulip breaking virus. Virus Genes 1994, 8, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Fauquet, C.M.; Mayo, M.A.; Maniloff, J.; Desselberger, U.; Ball, L.A. (Eds.) Virus taxonomy. Eighth report of the International Committee on the Taxonomy of Virus; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; ISBN 978-0-12-249951-7. [Google Scholar]
- Bloembollenkeuringsdienst (BKD). Knollenmozaïekvirus in Tulp, Een Zeldzaam Verschijnsel. Available online: http://www.bkd.eu/Pages/04_Gewasinformatie/04_Knollenmoza%c3%afe (accessed on 28 April 2010).
- Murphy, F.A.; Fauquet, C.M.; Bishop, D.H.L.; Ghabrial, S.A.; Jarvis, A.W.; Martelli, G.P.; Mayo, M.A.; Summers, M.D. (Eds.) Virus Taxonomy: Sixth report of the International Committee on Taxonomy of Viruses; Springer: Vienna, Austria, 1995; ISBN 978-3-211-82594-5. [Google Scholar]
- Wylie, S.J.; Adams, M.; Chalam, C.; Kreuze, J.; López-Moya, J.J.; Ohshima, K.; Praveen, S.; Rabenstein, F.; Stenger, D.; Wang, A.; et al. ICTV Virus Taxonomy Profile: Potyviridae. J. Gen. Virol. 2017, 98, 352–354. [Google Scholar] [CrossRef]
- Langeveld, S.A.; Dore, J.M.; Memelink, J.; Derks, A.F.L.M.; van der Vlugt, C.I.M.; Asjes, C.J.; Bol, J.F. Identification of potyviruses using the polymerase chain-reaction with degenerate primers. J. Gen. Virol. 1991, 72, 1531–1541. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Chen, J.; Zhao, M.F.; Lin, L.; Chen, J.P.; Antoniw, J.F.; Adams, M.J. Occurrence and sequences of Lily mottle virus and Lily symptomless virus in plants grown from imported bulbs in Zhejiang province, China. Arch. Virol. 2003, 148, 2419–2428. [Google Scholar] [CrossRef]
- Rivas, E.B.; de Almeida Bodi, E.C.; Harakava, R.; Gregori, F.; Goncalves, M.C. Occurrence and molecular analysis of quarantine virus in lily cultivation areas in Brazil. Pesqui. Agropecu. Bras. 2016, 51, 615–622. [Google Scholar] [CrossRef][Green Version]
- Dietrich, C.; Maiss, E. Fluorescent labelling reveals spatial separation of potyvirus populations in mixed infected Nicotiana benthamiana plants. J. Gen. Virol. 2003, 84, 2871–2876. [Google Scholar] [CrossRef]
- Elena, S.F.; Bedhomme, S.; Carrasco, P.; Cuevas, J.M.; de la Iglesia, F.; Lafforgue, G.; Lalić, J.; Pròsper, À.; Tromas, N.; Zwart, M.P. The Evolutionary Genetics of Emerging Plant RNA Viruses. MPMI 2011, 24, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Gagarinova, A.G.; Babu, M.; Strömvik, M.V.; Wang, A. Recombination analysis of Soybean mosaic virus sequences reveals evidence of RNA recombination between distinct pathotypes. Virol. J. 2008, 5, 143. [Google Scholar] [CrossRef] [PubMed]
- Galli, A.; Bukh, J. Comparative analysis of the molecular mechanisms of recombination in hepatitis C virus. Trends Microbiol. 2014, 22, 354–364. [Google Scholar] [CrossRef] [PubMed]
- García, J.A.; Glasa, M.; Cambra, M.; Candresse, T. Plum pox virus and sharka: A model potyvirus and a major disease: Plum pox virus. Mol. Plant Pathol. 2014, 15, 226–241. [Google Scholar] [CrossRef] [PubMed]
- García-Arenal, F.; Fraile, A.; Malpica, J.M. Variation and evolution of plant virus populations. Int. Microbiol. 2003, 6, 225–232. [Google Scholar] [CrossRef]
- Nagy, P.D. Recombination in Plant RNA Viruses. In Plant Virus Evolution; Roossinck, M.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 133–156. ISBN 978-3-540-75762-7. [Google Scholar]
- Gibbs, A.; Ohshima, K. Potyviruses and the digital revolution. Annu. Rev. Phytopathol. 2010, 48, 205–223. [Google Scholar] [CrossRef]
- Kehoe, M.A.; Coutts, B.A.; Buirchell, B.J.; Jones, R.A.C. Hardenbergia mosaic virus: Crossing the barrier between native and introduced plant species. Virus Res. 2014, 184, 87–92. [Google Scholar] [CrossRef]
- Revers, F.; Le Gall, O.; Candresse, T.; Le Romancer, M.; Dunez, J. Frequent occurrence of recombinant potyvirus isolates. J. Gen. Virol. 1996, 77, 1953–1965. [Google Scholar] [CrossRef]
- Valli, A.; López-Moya, J.J.; García, J.A. Recombination and gene duplication in the evolutionary diversification of P1 proteins in the family Potyviridae. J. Gen. Virol. 2007, 88, 1016–1028. [Google Scholar] [CrossRef]
- Desbiez, C.; Lecoq, H. The nucleotide sequence of Watermelon mosaic virus (WMV, Potyvirus) reveals interspecific recombination between two related potyviruses in the 5′ part of the genome. Arch. Virol. 2004, 149, 1619–1632. [Google Scholar] [CrossRef]
- Ohshima, K.; Mitoma, S.; Gibbs, A.J. The genetic diversity of narcissus viruses related to turnip mosaic virus blur arbitrary boundaries used to discriminate potyvirus species. PLoS ONE 2018, 13, e0190511. [Google Scholar] [CrossRef] [PubMed]
- Wylie, S.J.; Jones, R.A.C. Role of recombination in the evolution of host specialization within Bean yellow mosaic virus. Phytopathology 2009, 99, 512–518. [Google Scholar] [CrossRef]
- Zakubanskiy, A.; Mitrofanova, I.; Sheveleva, A.; Chirkov, S. Analysis of canna yellow streak virus complete genomes provides evidence of multiple intraspecies recombination events. J. Plant Pathol. 2018, 100, 575–580. [Google Scholar] [CrossRef]
- Krause-Sakate, R.; Fakhfakh, H.; Peypelut, M.; Pavan, M.A.; Zerbini, F.M.; Marrakchi, M.; Candresse, T.; Le Gall, O. A naturally occurring recombinant isolate of Lettuce mosaic virus. Arch. Virol. 2003, 149, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Navrátil, M.; Šafářová, D. Naturally occurring recombinant isolate of Pea seed-borne mosaic virus—Short Communication. Plant Protect. Sci. 2019, 55, 167–171. [Google Scholar] [CrossRef]
- Chirkov, S.; Ivanov, P.; Sheveleva, A.; Zakubanskiy, A.; Osipov, G. New highly divergent Plum pox virus isolates infecting sour cherry in Russia. Virology 2017, 502, 56–62. [Google Scholar] [CrossRef]
- Chirkov, S.; Ivanov, P.; Sheveleva, A.; Kudryavtseva, A.; Mitrofanova, I. Molecular characterization of Plum pox virus Rec isolates from Russia suggests a new insight into evolution of the strain. Virus Genes 2018, 54, 328–332. [Google Scholar] [CrossRef]
- Glasa, M.; Palkovics, L.; Komínek, P.; Labonne, G.; Pittnerová, S.; Kúdela, O.; Candresse, T.; Šubr, Z. Geographically and temporally distant natural recombinant isolates of Plum pox virus (PPV) are genetically very similar and form a unique PPV subgroup. J. Gen. Virol. 2004, 85, 2671–2681. [Google Scholar] [CrossRef]
- Gürcan, K.; Teber, S.; Candresse, T. Genetic analysis suggests a long and largely isolated evolutionary history of plum pox virus strain D in Turkey. Plant Pathol. 2019; ppa.13115. [Google Scholar] [CrossRef]
- Jelkmann, W.; Sanderson, D.; Berwarth, C.; James, D. First detection and complete genome characterization of a Cherry (C) strain isolate of plum pox virus from sour cherry (Prunus cerasus) in Germany. J. Plant Dis. Prot. 2018, 125, 267–272. [Google Scholar] [CrossRef]
- Green, K.J.; Brown, C.J.; Gray, S.M.; Karasev, A.V. Phylogenetic study of recombinant strains of Potato virus Y. Virology 2017, 507, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Ximba, S.P.F.; Ibaba, J.D.; Gubba, A. Potato virus Y strains infecting potatoes in the Msinga district in the province of KwaZulu-Natal, South Africa. Crop Prot. 2017, 96, 188–194. [Google Scholar] [CrossRef]
- Yasaka, R.; Fukagawa, H.; Ikematsu, M.; Soda, H.; Korkmaz, S.; Golnaraghi, A.; Katis, N.; Ho, S.Y.W.; Gibbs, A.J.; Ohshima, K. The timescale of emergence and spread of turnip mosaic potyvirus. Sci. Rep. 2017, 7, 4240. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, O.A.; Ali, A. Molecular characterization reveals that squash chlorosis mottling virus and zucchini tigré mosaic virus are the same newly emerging potyvirus. Arch. Virol. 2018, 163, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Maina, S.; Coutts, B.A.; Edwards, O.R.; de Almeida, L.; Kehoe, M.A.; Ximenes, A.; Jones, R.A.C. Zucchini yellow mosaic virus populations from East Timorese and Northern Australian cucurbit crops: Molecular properties, genetic connectivity, and biosecurity implications. Plant Dis. 2017, 101, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Maina, S.; Barbetti, M.J.; Edwards, O.R.; Minemba, D.; Areke, M.W.; Jones, R.A.C. Zucchini yellow mosaic virus genomic sequences from Papua New Guinea: Lack of genetic connectivity with Northern Australian or East Timorese genomes, and new recombination findings. Plant Dis. 2019, 103, 1326–1336. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Yamaguchi, Y.; Hirota, R.; Hamamoto, T.; Tomimura, K.; Tan, Z.; Sano, T.; Azuhata, F.; Walsh, J.A.; Fletcher, J.; et al. Molecular evolution of Turnip mosaic virus: Evidence of host adaptation, genetic recombination and geographical spread. J. Gen. Virol. 2002, 83, 1511–1521. [Google Scholar] [CrossRef]
- Tomimura, K.; Špak, J.; Katis, N.; Jenner, C.E.; Walsh, J.A.; Gibbs, A.J.; Ohshima, K. Comparisons of the genetic structure of populations of Turnip mosaic virus in West and East Eurasia. Virology 2004, 330, 408–423. [Google Scholar] [CrossRef]
- Gibbs, A.J.; Ohshima, K.; Phillips, M.J.; Gibbs, M.J. The prehistory of potyviruses: Their initial radiation was during the dawn of agriculture. PLoS ONE 2008, 3, e2523. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Tomitaka, Y.; Ho, S.Y.W.; Duchêne, S.; Vetten, H.-J.; Lesemann, D.; Walsh, J.A.; Gibbs, A.J.; Ohshima, K. Turnip mosaic potyvirus probably first spread to eurasian brassica crops from wild orchids about 1000 years ago. PLoS ONE 2013, 8, e55336. [Google Scholar] [CrossRef]
- Van Scheepen, J. Classified List and International Register of Tulip Names; Koninklijke Algemeene Vereeniging voor Bloembollencultuur (KAVB): Hillegom, The Netherlands, 1996; ISBN 978-90-73350-02-1. [Google Scholar]
- Jordan, R.; Hammond, J. Comparison and differentiation of potyvirus isolates and identification of strain-, virus-, subgroup-specific and potyvirus group-common epitopes using monoclonal antibodies. J. Gen. Virol. 1991, 72, 25–36. [Google Scholar] [CrossRef]
- White, J.L.; Kaper, J.M. A simple method for detection of viral satellite RNAs in small plant tissue samples. J. Virol. Methods 1989, 23, 83–93. [Google Scholar] [CrossRef]
- Salamon, P.; Palkovics, L. Occurrence of Colombian datura virus in Brugmansia hybrids, Physalis peruviana L. and Solanum muricatum Ait. in Hungary. Acta Virol. 2005, 49, 117–122. [Google Scholar] [PubMed]
- Thole, V.; Dalmay, T.; Burgyán, J.; Balázs, E. Cloning and sequencing of potato virus Y (Hungarian isolate) genomic RNA. Gene 1993, 123, 149–156. [Google Scholar] [CrossRef]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef]
- Adams, M.J.; Antoniw, J.F.; Fauquet, C.M. Molecular criteria for genus and species discrimination within the family Potyviridae. Arch. Virol. 2005, 150, 459–479. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK; New York, NY, USA, 2000; ISBN 978-0-19-513584-8. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Antoniw, J.F.; Beaudoin, F. Overview and analysis of the polyprotein cleavage sites in the family Potyviridae. Mol. Plant Pathol. 2005, 6, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Murrell, B.; Khoosal, A.; Muhire, B. Detecting and Analyzing Genetic Recombination Using RDP4. In Bioinformatics; Keith, J.M., Ed.; Springer: New York, NY, USA, 2017; Volume 1525, pp. 433–460. ISBN 978-1-4939-6620-2. [Google Scholar]
- Martin, D.; Rybicki, E. RDP: Detection of recombination amongst aligned sequences. Bioinformatics 2000, 16, 562–563. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Evaluation of methods for detecting recombination from DNA sequences: Computer simulations. Proc. Natl. Acad. Sci. USA 2001, 98, 13757–13762. [Google Scholar] [CrossRef]
- Martin, D.P.; Posada, D.; Crandall, K.A.; Williamson, C. A modified Bootscan algorithm for automated identification of recombinant sequences and recombination breakpoints. Aids Res. Hum. Retrovir. 2005, 21, 98–102. [Google Scholar] [CrossRef]
- Lam, H.M.; Ratmann, O.; Boni, M.F. Improved Algorithmic Complexity for the 3SEQ Recombination Detection Algorithm. Mol. Biol. Evol. 2018, 35, 247–251. [Google Scholar] [CrossRef]
- Padidam, M.; Sawyer, S.; Fauquet, C.M. Possible emergence of new geminiviruses by frequent recombination. Virology 1999, 265, 218–225. [Google Scholar] [CrossRef]
- Smith, J.M. Analyzing the mosaic structure of genes. J. Mol. Evol. 1992, 34. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, M.J.; Armstrong, J.S.; Gibbs, A.J. Sister-Scanning: A Monte Carlo procedure for assessing signals in recombinant sequences. Bioinformatics 2000, 16, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Nei, M. Limitations of the evolutionary parsimony method of phylogenetic analysis. Mol. Biol. Evol. 1990, 7, 82–102. [Google Scholar] [CrossRef]
- Weiller, G.F. Phylogenetic profiles: A graphical method for detecting genetic recombinations in homologous sequences. Mol. Biol. Evol. 1998, 15, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.C.; Worobey, M.; Rambaut, A. Phylogenetic evidence for recombination in dengue virus. Mol. Biol. Evol. 1999, 16, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Tavaré, S. Some Probabilistic and Statistical Problems in the Analysis of DNA Sequences. In Some Mathematical Questions in Biology: DNA Sequence Analysis; American Mathematical Society: Providence, RI, USA, 1986; Volume 17, pp. 57–86. [Google Scholar]
- McGuire, G.; Wright, F. TOPAL 2.0: Improved detection of mosaic sequences within multiple alignments. Bioinformatics 2000, 16, 130–134. [Google Scholar] [CrossRef]
- Felsenstein, J. Phylip Phylogeny Inference Package (Version 3.2). Cladistics 1989, 5, 164–166. [Google Scholar] [CrossRef]
- Voigt, C.A.; Martinez, C.; Wang, Z.-G.; Mayo, S.L.; Arnold, F.H. Protein building blocks preserved by recombination. Nat. Struct. Biol. 2002. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recombination Event | Acronym of Recombinant Virus | Recombinant Sequence | Major Parent | Minor Parent |
|---|---|---|---|---|
| 1 | LMoV | JN127341 | unknown | KF553658 |
| 2 | TBV | X63630 | MK368783 1 | KF442403 2 |
| 3 | TBV | KF442403 | MK368787 1 | unknown |
| 4 | LMoV | AJ310203 | S44147 | MF983709 1 |
| 5 | LMoV | JN127341 | AJ310203 2 | unknown |
| 6 | TBV | KT923168 | unknown | MK368785 1 |
| Average p-Values 1 by Recombination Event Number | ||||||
|---|---|---|---|---|---|---|
| Recombination Event | ||||||
| Method | 1 | 2 | 3 | 4 | 5 | 6 |
| RDP | 1.713 × 10−5 | 5.965 × 10−9 | 1.655 × 10−6 | 1.863 × 10−1 | 5.776 × 10−1 | 2.448 × 10−2 |
| GENECONV | 7.062 × 10−9 | 4.312 × 10−7 | 3.589 × 10−5 | 3.589 × 10−2 | - | 4.265 × 10−1 |
| Bootscan | 3.197 × 10−5 | >1.0 | >1.0 | 2.2 × 10−3 | - | >1.0 |
| MaxChi | >1.0 | >1.0 | >1.0 | >1.0 | >1.0 | 6.678 × 10−1 |
| Chimaera | 9.692 × 10−10 | 5.254 × 10−9 | >1.0 | >1.0 | 1.087 × 10−3 | 6.317 × 10−1 |
| SiScan | 6.921 × 10−16 | 2.561 × 10−26 | 7.68 × 10−17 | 4.867 × 10−27 | 5.221 × 10−20 | 3.991 × 10−7 |
| PhylPro | 5.212 × 10−20 | 1.123 × 10−19 | 1.489 × 10−13 | 9.194 × 10−8 | 2.279 × 10−7 | 3.827 × 10−5 |
| LARD | - | - | - | - | - | - |
| 3Seq | - | - | - | - | - | - |
| Identities of Virus nt Sequences (%) | ||||
|---|---|---|---|---|
| Acronym | LMoV | TBV | ReTBV | TEV |
| LMoV | 85.89–99.76% | |||
| TBV | 63.05–65.29% | 94.78–100% | ||
| ReTBV | 70.16–71.04% | 65.95–67.13% | 89.38–99.88% | |
| TEV | 59.95–60.84% | 63.91–65.82% | 60.71–60.91% | 100% |
| Identities of Virus aa Sequences (%) | ||||
|---|---|---|---|---|
| Acronym | LMoV | TBV | ReTBV | TEV |
| LMoV | 87.23–100% | |||
| TBV | 62.17–67.04% | 97.01–100% | ||
| ReTBV | 66.54–72.66% | 66.79–68.44% | 95.49–100% | |
| TEV | 58.56–59.32% | 64.26–65.01% | 61.24–62.16% | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ágoston, J.; Almási, A.; Salánki, K.; Palkovics, L. Genetic Diversity of Potyviruses Associated with Tulip Breaking Syndrome. Plants 2020, 9, 1807. https://doi.org/10.3390/plants9121807
Ágoston J, Almási A, Salánki K, Palkovics L. Genetic Diversity of Potyviruses Associated with Tulip Breaking Syndrome. Plants. 2020; 9(12):1807. https://doi.org/10.3390/plants9121807
Chicago/Turabian StyleÁgoston, János, Asztéria Almási, Katalin Salánki, and László Palkovics. 2020. "Genetic Diversity of Potyviruses Associated with Tulip Breaking Syndrome" Plants 9, no. 12: 1807. https://doi.org/10.3390/plants9121807
APA StyleÁgoston, J., Almási, A., Salánki, K., & Palkovics, L. (2020). Genetic Diversity of Potyviruses Associated with Tulip Breaking Syndrome. Plants, 9(12), 1807. https://doi.org/10.3390/plants9121807