Proving the Mode of Action of Phytotoxic Phytochemicals



Abstract
1. Introduction
2. Molecular Target Sites and Resulting Secondary and Tertiary Effects
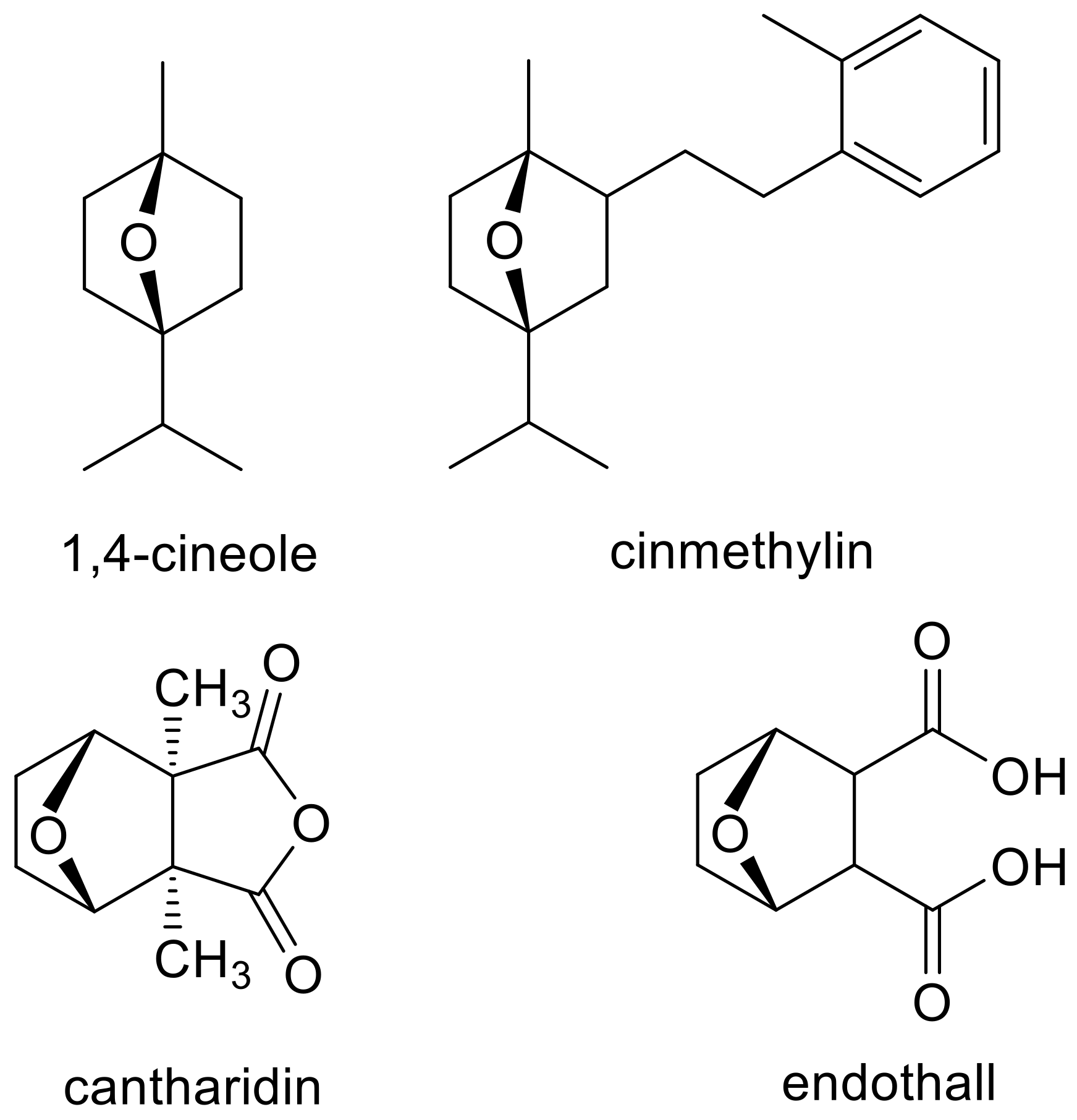
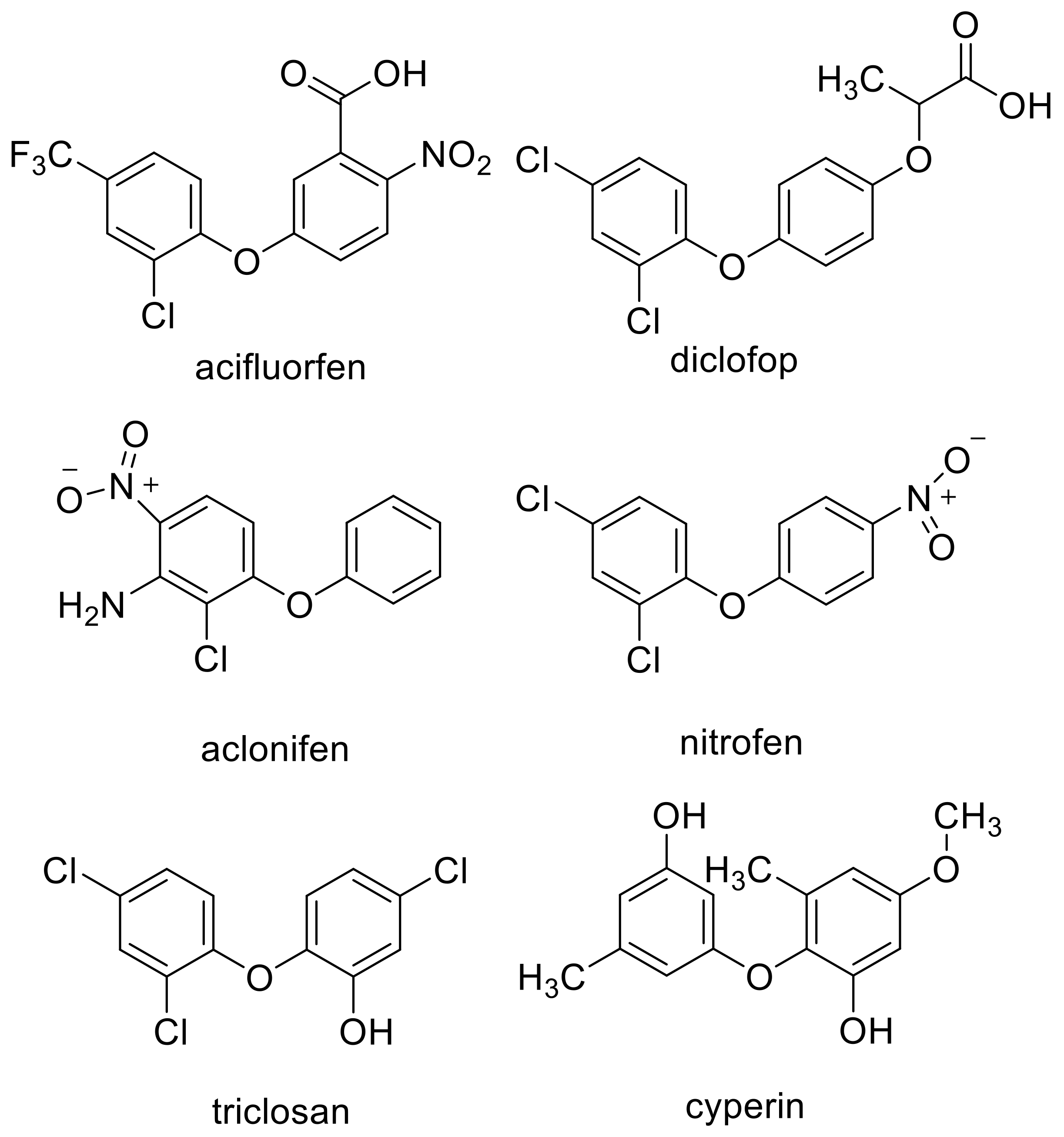
3. Clues from Molecular Structure and In Silico Binding Studies
4. Finding the Primary Molecular Target with Physiology and Biochemistry
4.1. Choosing Test Organisms
4.2. Achieving the Optimal Dose and Timing
4.3. Visible Phenotypic Response Clues
4.4. Reversion and Inhibitor Studies
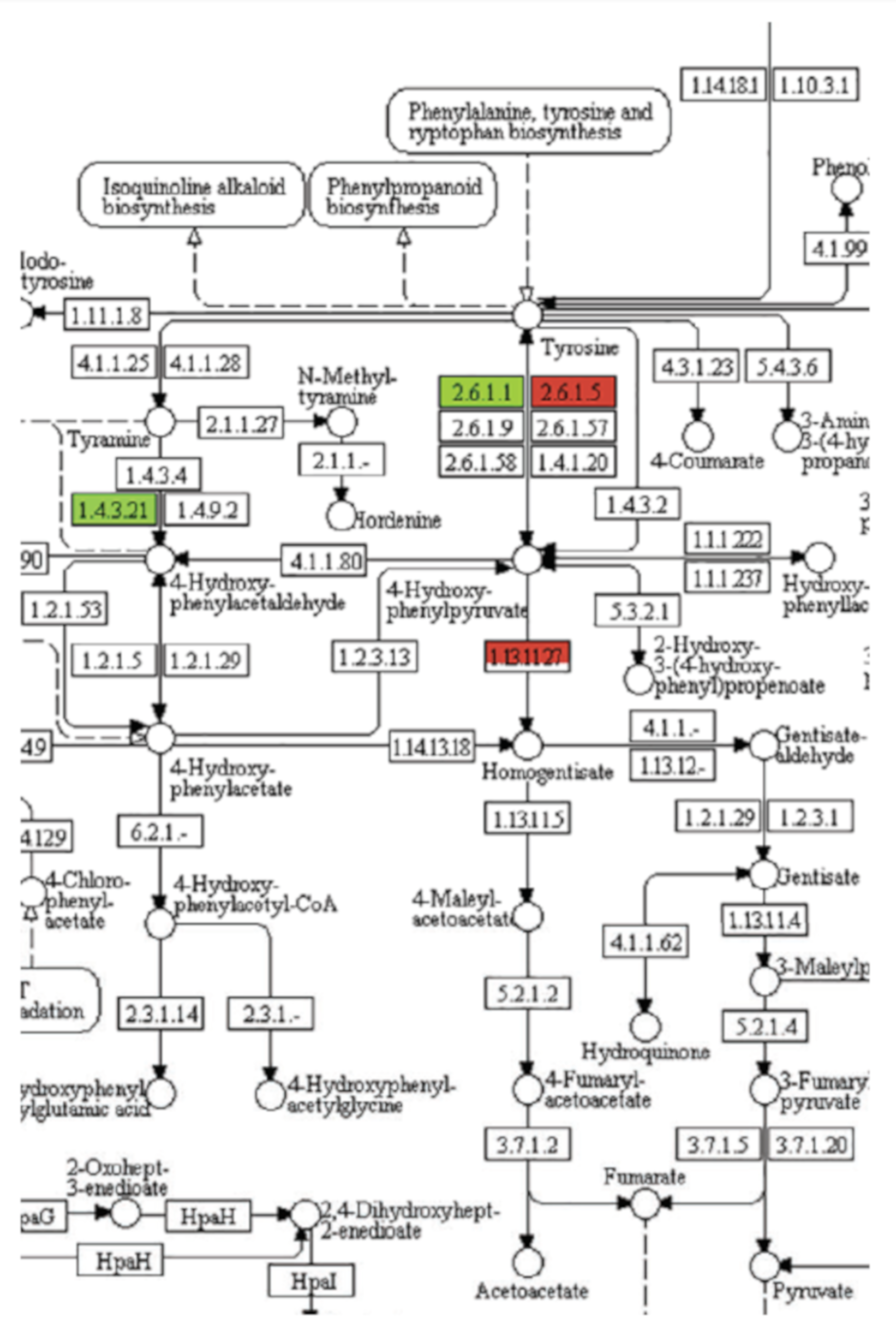
4.5. Omics Methods
4.6. Direct Measurement of Effects of the PP on Molecular Targets
4.7. Complications by Multiple Targets
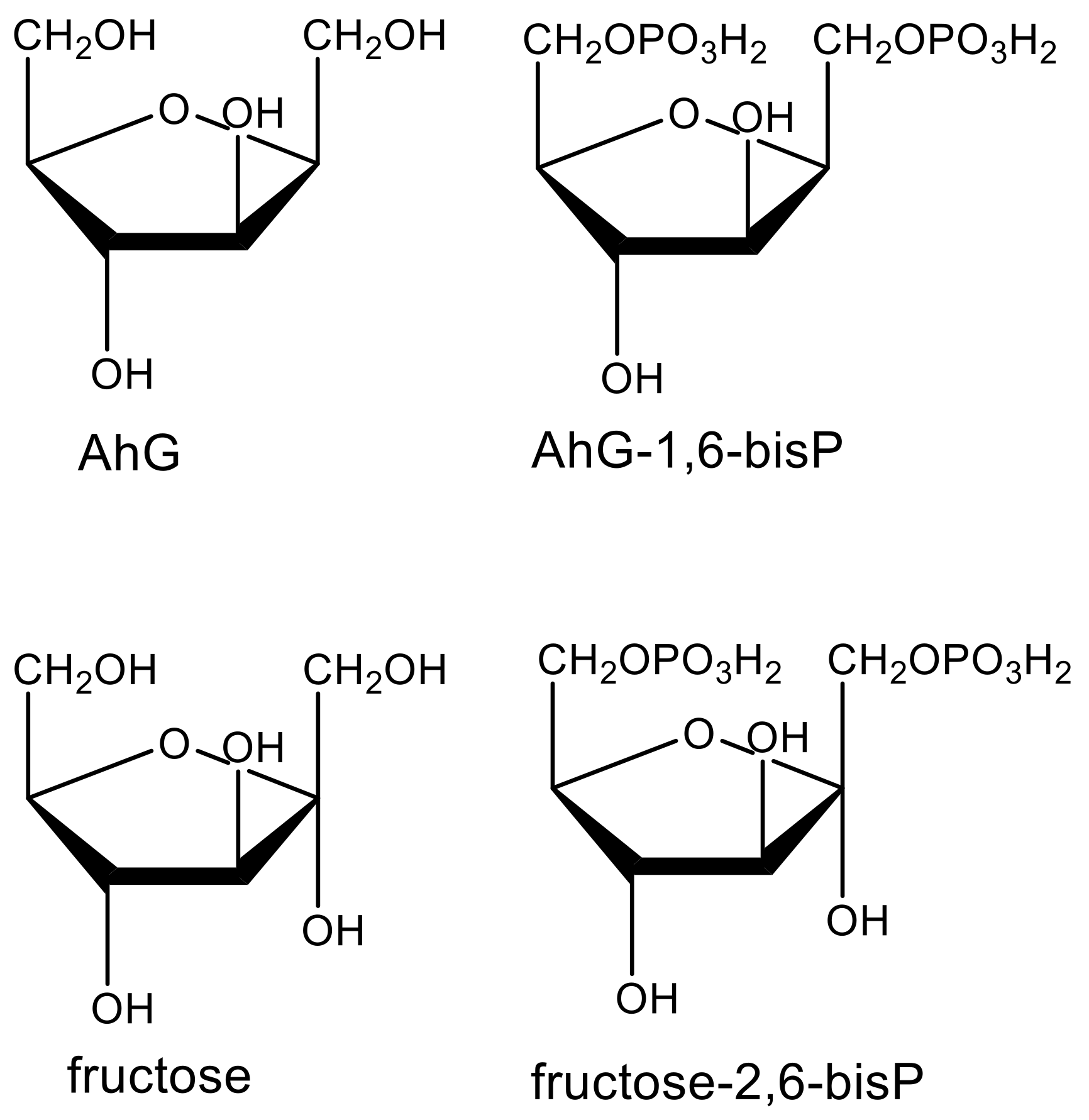
4.8. Complications by Prophytotoxins
5. Genetic Proof of the Target
5.1. Comparing Phenotypes of Known Mutants with That of Treatment with the Phytotoxin
5.2. Generating Genes for Resistant Targets
5.3. Genetics Approach Using Resistant Mutants
6. Artificial Intelligence in MOA Discovery—Prospects
7. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Duke, S.O. Proving allelopathy in crop-weed interactions. Weed Sci. 2015, 63, 121–132. [Google Scholar] [CrossRef]
- Streibig, J.C.; Dayan, F.E.; Rimando, A.M.; Duke, S.O. Joint action of natural and synthetic photosystem II inhibitors. Pestic. Sci. 1999, 55, 137–146. [Google Scholar] [CrossRef]
- Arntzen, C.J.; Ditto, C.L.; Brewer, P.E. Chloroplast membrane alterations in triazine-resistant Amaranthus retroflexus biotypes. Proc. Natl. Acad. Sci. USA 1979, 76, 278–282. [Google Scholar] [CrossRef]
- Dayan, F.E.; Howell, J.; Weidenhamer, J.D. Dynamic root exudation of sorgoleone and its in planta mechanism of action. J. Exp. Bot. 2009, 60, 1032–1039. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: www.weedsci.org (accessed on 9 October 2020).
- Herbicide Resistance Action Committee. Available online: www.hracglobal.com (accessed on 12 October 2020).
- Knudsen, C.G.; Lee, D.L.; Michaely, W.J.; Chin, H.-L.; Nguyen, N.H.; Rusay, R.J.; Cromartie, T.H.; Gray, R.; Lake, B.H.; Fraser, T.E.M.; et al. Discovery of the triketone class of HPPD inhibiting herbicides and their relationship to naturally occurring β-triketones. In Allelopathy in Ecological Agriculture and Forestry; Narwal, S.S., Hoagland, R.E., Dilday, R.H., Reigosa, M.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 101–111. [Google Scholar]
- Duke, S.O. Why have no new herbicide modes of action appeared in recent years? Pest Manag. Sci. 2012, 68, 505–512. [Google Scholar] [CrossRef]
- Dayan, F.E.; Howell, J.L.; Marais, J.M.; Ferreira, D.; Koivunen, M.E. Manuka oil, a natural herbicide with preemergence activity. Weed Sci. 2011, 59, 464–469. [Google Scholar] [CrossRef]
- Owens, D.K.; Nanayakkara, N.P.D.; Dayan, F.E. In planta mechanism of action of leptospermone: Impact of its physico-chemical properties on uptake, translocation, and metabolism. J. Chem. Ecol. 2013, 39, 262–270. [Google Scholar] [CrossRef]
- Dayan, F.E.; Duke, S.O.; Sauldubois, A.; Singh, N.; McCurdy, C.; Cantrell, C.L. p-Hydroxyphenylpyruvate dioxygenase is the herbicidal target site for β-triketones from Leptospermum scoparium. Phytochemistry 2007, 68, 2004–2014. [Google Scholar] [CrossRef]
- Bajsa, J.; Pan, Z.; Dayan, F.E.; Owens, D.K.; Duke, S.O. Validation of serine/threonine protein phosphatase as the herbicide target site of endothall. Pestic. Biochem. Physiol. 2012, 102, 38–44. [Google Scholar] [CrossRef]
- Campe, R.; Hollenbach, E.; Kämmerer, L.; Hendriks, J.; Höffken, H.W.; Kraus, H.; Lerchl, J.; Mietzner, T.; Tresch, S.; Witschel, M.; et al. A new herbicidal site of action: Cinmethylin binds to acyl-ACP thioesterase and inhibits plant fatty acid synthase. Pestic. Bichem. Physiol. 2018, 148, 116–125. [Google Scholar] [CrossRef]
- Jessing, K.; Duke, S.O.; Cedergreen, N. Potential ecological roles of artemisinin produced by Artemisia anna L. J. Chem. Ecol. 2014, 40, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Babula, P.; Vaverkova, V.; Poborilova, Z.; Ballova, L.; Masarik, M.; Provaznik, I. Phyototoxic action of napthoquinone juglone demonstrated on lettuce seedling roots. Plant Physiol. Biochem. 2014, 84, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Koussevitsky, S.; Mittler, R.; Miller, G. ROS and redox signaling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Devine, M.D.; Duke, S.O.; Fedtke, C. Physiology of Herbicide Action; Prentice Hall: Englewood Cliffs, NJ, USA, 1993; pp. 163–166. [Google Scholar]
- Dayan, F.E.; Duke, S.O. Herbicides: Protoporphyrinogen oxidase inhibitors. In Encyclopedia of Agrochemicals; Plimmer, J.R., Gammon, D.W., Ragsdale, N.N., Eds.; John Wiley & Sons: New York, NY, USA, 2003; Volume 2, pp. 850–863. [Google Scholar]
- Yu, J.Q.; Ye, S.F.; Zhang, M.F.; Hu, W.H. Effects of root exudates and aqueous root extracts of cucumber (Cucumis sativus) and allelochemicals, on photosynthesis and antioxidant enzymes in cucumber. Biochem. Syst. Ecol. 2003, 31, 129–139. [Google Scholar] [CrossRef]
- Zhang, H.; Tweel, B.; Tong, L. Molecular basis for the inhibition of the carboxytransferase domain of acetyl-coenzyme-A caroboxylase by haloxyfop and diclofop. Proc. Natl. Acad. Sci. USA 2004, 101, 5910–5915. [Google Scholar] [CrossRef]
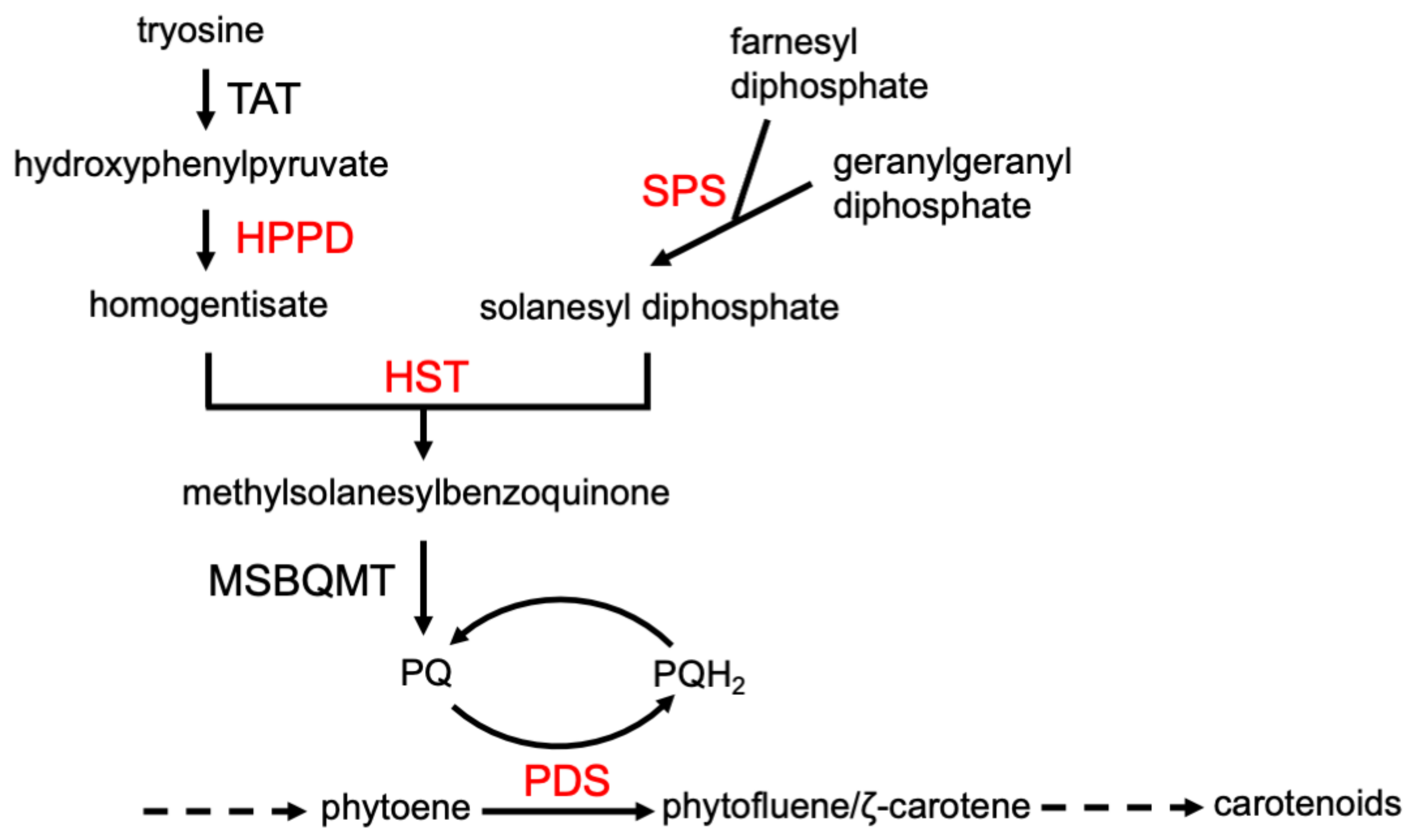
- Kahlau, S.; Schröder, F.; Freigang, J.; Laber, B.; Lange, G.; Passon, D.; Kleeßen, S.; Lohse, M.; Schulz, A.; von Koskull-Döring, P.; et al. Aclonifen targets solanesyl diphosphate synthase, representing a novel mode of action for herbicides. Pest Manag. Sci. 2020, 76, 3377–3388. [Google Scholar] [CrossRef]
- Kunert, K.-J.; Sandmann, G.; Böger, P. Mode of action of diphenyl ethers. Rev. Weed Sci. 1997, 3, 35–55. [Google Scholar]
- Dayan, F.E.; Ferreria, D.; Wang, Y.-H.; Khan, I.A.; McInroy, J.A.; Pan, Z. A pathogenic fungi diphenyl ether phytotoxin targets plant enoyl (acyl carrier protein) reductase. Plant Physiol. 2008, 147, 1062–1071. [Google Scholar] [CrossRef]
- Zhu, S.-S.; Zhang, G.-J.; Liao, H.-B.; Wang, H.-S.; Liang, D. A new chlorinated diphenyl ether and a new sesquilignan from the stems of Mappianthus tomentosa. Phytochem. Lett. 2019, 30, 49–52. [Google Scholar] [CrossRef]
- Abbas, H.K.; Duke, S.O.; Shier, W.T.; Duke, M.V. Inhibition of ceramide synthesis in plants by phytotoxins. In Advances in Microbial Toxin Research and Its Biotechnological Exploitation; Upadhyay, R.K., Ed.; Kluwer Academic/Plenum Publ.: London, UK, 2002; pp. 211–229. [Google Scholar]
- Dayan, F.E.; Rimando, A.M.; Tellez, M.R.; Scheffler, B.E.; Roy, T.; Abbas, H.K.; Duke, S.O. Bioactivation of the fungal phytotoxin 2,4-anhydro-d-glucitol by glycolytic enzymes is an essential component of its mechanism of action. Z. Naturforsch. C 2002, 57, 645–653. [Google Scholar] [CrossRef] [PubMed]
- McRobb, F.M.; Kufareva, I.; Abagyan, R. In silico identification and pharmacological evaluation of novel endocrine disrupting chemicals that act via the ligand-binding domain of the estrogenic receptor α. Toxicol. Sci. 2014, 141, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Graña, E.; Días-Tielas, C.; Sánchez-Moreiras, A.M.; Reigosa, M.J.; Celiero, M.; Abagyan, R.; Teijeira, M.; Pan, Z.; Duke, S.O. Transcriptome and binding data indicate that citral inhibits single strand DNA binding proteins. Physiol. Plant. 2020, 169, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Romagni, J.G.; Duke, S.O. Investigating the mode of action of natural phytotoxins. J. Chem. Ecol. 2000, 26, 2079–2094. [Google Scholar] [CrossRef]
- Michel, A.; Johnson, R.D.; Duke, S.O.; Scheffler, B.E. Dose-response relationships between herbicides with different modes of action and growth of Lemna paucicostata—An improved ecotoxicological method. Environ. Toxicol. Chem. 2004, 23, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Kenyon, W.H. Peroxidizing activity determined by cellular leakage. In Target Assays for Modern Herbicides and Related Phytotoxic Compounds; Böger, P., Sandmann, G., Eds.; CRC Press: Boca Raton, FL, USA, 1993; pp. 61–66. [Google Scholar]
- Cutler, H.G.; Reid, W.W.; Delétang, J. Plant growth inhibiting properties of diterpenes from tobacco. Plant Cell Physiol. 1977, 18, 711–714. [Google Scholar]
- Belz, R.G.; Velini, E.D.; Duke, S.O. Dose/response relationships in allelopathy research. In Allelopathy: New Concepts and Methodology; Fujii, Y., Hiradate, S., Eds.; Science Publishers: Enfield, NH, USA, 2007; pp. 3–29. [Google Scholar]
- Shaner, D.L.; Nadler-Hassar, T.; Henry, W.B.; Koger, C.H. A rapid in vivo shikimate accumulation assay with excised leaf discs. Weed Sci. 2004, 53, 769–774. [Google Scholar] [CrossRef]
- Dayan, F.E.; Duke, S.O. Discovery of new herbicide sites of action by quantification of plant primary metabolite and enzyme pools. Engineering 2020, 6, 509–514. [Google Scholar] [CrossRef]
- Vaughan, M.A.; Vaughn, K.C. Mitotic disruptors from higher plants and their potential uses as herbicides. Weed Technol. 1988, 2, 533–539. [Google Scholar] [CrossRef]
- Grossmann, K. What it takes to get a herbicide’s mode of action. Physionomics, a classical approach in a new complexion. Pest Manag. Sci. 2005, 61, 423–432. [Google Scholar] [CrossRef]
- Killmer, J.; Widholm, J.; Slife, F. Reversal of glyphosate inhibition of carrot cell culture growth by glycolytic intermediates and organic and amino acids. Plant Physiol. 1981, 68, 1299–1302. [Google Scholar] [CrossRef] [PubMed]
- Shaner, D.L.; Anderson, P.A.; Stidham, M.A. Imidazolinones. Potent inhibitors of acetohydroxyacid synthase. Plant Physiol. 1984, 76, 545–546. [Google Scholar] [CrossRef] [PubMed]
- Amagasa, T.; Paul, R.N.; Heitholt, J.J.; Duke, S.O. Biological effects of cornexistin on Lemna pausicostata. Pestic. Biochem. Physiol. 1994, 49, 37–52. [Google Scholar] [CrossRef]
- Siehl, D.L.; Subramanian, M.V.; Walters, E.W.; Lee, S.-F.; Anderson, R.J.; Toshi, A.G. Adenylosuccinate synthetase: Site of action of hydantocidin, a microbial phytotoxin. Plant Phyiol. 1996, 110, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Tielas, C.; Grãna, E.; Sánchez-Moreiras, A.M.; Reigosa, M.J.; Vaughn, J.N.; Pan, Z.; Hirshel-Bajsa, J.; Duke, M.V.; Duke, S.O. Transcriptome responses to the phytotoxin t-chalcone in Arabidopsis thaliana L. Pest Manag. Sci. 2019, 75, 2490–2504. [Google Scholar] [CrossRef] [PubMed]
- Giovanelli, J.; Owens, L.; Mudd, S. Mechanisms of inhibition of β-cystathionase by rhizobitoxine. Biochim. Biophys. Acta 1971, 227, 671–684. [Google Scholar] [CrossRef]
- Killmer, J.L.; Widholm, J.M.; Slife, F.W. Antagonistic effect of p-aminobenzoate or folate on asulam [methyl (4-aminobenzensulphonyl carbonate)] inhibition of carrot suspenstion cultures. Plant Sci. Lett. 1980, 19, 203–208. [Google Scholar] [CrossRef]
- Duke, S.O.; Evidente, A.; Fiore, M.; Rimando, A.M.; Dayan, F.E.; Vurro, M.; Christiansen, N.; Looser, R.; Hutzler, J.; Grossmann, K. Effects of the aglycone of ascaulitoxin on amino acid metabolism in Lemna paucicostata. Pestic. Biochem. Physiol. 2011, 100, 41–50. [Google Scholar] [CrossRef]
- Brilisauer, K.; Rapp, J.; Rath, P.; Schöllhorn, A.; Bleul, L.; Weiß, E.; Stahl, M.; Grond, S.; Forschhammer, K. Cyanobacterial antimetabolite 7-deoxy-sedoheptulose blocks the shikimiate pathway to inhibit the growth of phototrophic organsims. Nat. Commun. 2019, 10, 545. [Google Scholar] [CrossRef]
- Rimando, A.M.; Cuendet, M.; Desmarchelier, C.; Mehta, R.G.; Pezzuto, J.M.; Duke, S.O. Cancer chemopreventative and antioxidant activities of pterostilbene, a naturally occurring analogue of resveratrol. J. Agric. Food. Chem. 2002, 50, 3453–3457. [Google Scholar] [CrossRef]
- Galindo, J.C.G.; Hernández, A.; Dayan, F.E.; Tellez, M.R.; Maciás, F.A.; Paul, F.N.; Duke, S.O. Dehydrozaluzanin C, a natural sesquiterpenolide, causes rapid plasma membrane damage. Phytochemistry 1999, 52, 805–813. [Google Scholar] [CrossRef]
- Lydon, J.; Duke, S.O. Porphyrin synthesis is required for photobleaching activity of the p-nitrosubstituted diphenyl ether herbicides. Pestic. Biochem. Physiol. 1988, 31, 74–83. [Google Scholar] [CrossRef]
- Liu, X.-H.; Yu, W.; Min, L.-J.; Johnson, R.D.; Wedge, D.E.; Tan, C.-X.; Weng, J.-Q.; Wu, H.-K.; Cantrell, C.L.; Bajsa-Hirschel, J.; et al. Synthesis and pesticidal activities of new quinoxalines. J. Agric. Food Chem. 2020, 68, 7324–7332. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Becerril, J.M.; Lydon, J.; Matsumoto, H.; Sherman, T.D. Protoporphyrinogen oxidase-inhibiting herbicides. Weed Sci. 1991, 39, 465–473. [Google Scholar] [CrossRef]
- Grossman, K.; Hutzler, J.; Tresch, S.; Christiansen, N.; Looser, R.; Ehrhardt, T. On the mode of action of herbicides cinmethylin and 5-benzyloxymethyl-1,2-isoxazolines: Putative inhibitors of plant tyrosine aminotransferase. Pest Manag. Sci. 2012, 68, 482–492. [Google Scholar] [CrossRef]
- Duke, S.O. Glyphosate: Uses other than in glyphosate-resistant crops, mode of action, degradation in plants, and effects on non-target plants and agricultural microbes. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 2021; Volume 255, in press. [Google Scholar]
- Duke, S.O.; Bajsa, J.; Pan, Z. Omics methods for probing the mode of action of natural and synthetic phytotoxins. J. Chem. Ecol. 2013, 39, 333–347. [Google Scholar] [CrossRef]
- Duke, S.O.; Pan, Z.; Bajsa-Hirschel, J.; Sánchez-Moreiras, A.M.; Vaughn, J.N. Use of omics methods to determine the mode of action of natural phytotoxins. In Roles of Natural Products for Biorational Pesticides in Agriculture; American Chemical Society: Washington, DC, USA, 2018; Volume 1294, pp. 33–46. [Google Scholar]
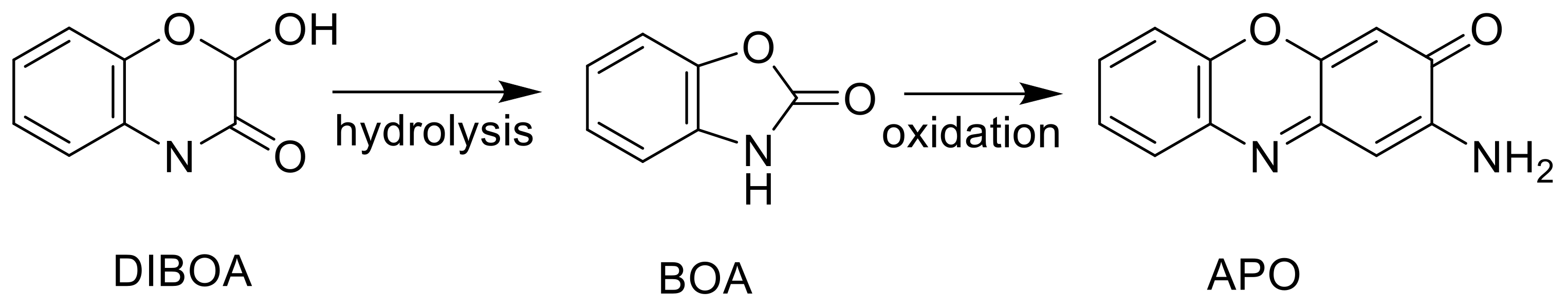
- Baerson, S.R.; Sánchez-Moreiras, A.; Pedrol-Bonjoch, N.; Schulz, M.; Kagan, I.A.; Agarwal, A.K.; Reigosa, M.J.; Duke, S.O. Detoxification and transcriptome response in Arabidopsis seedlings exposed to the allelochemical benzoxazolin-2(3H)-one (BOA). J. Biol. Chem. 2005, 280, 21867–21881. [Google Scholar] [CrossRef]
- Bajsa, J.; Pan, Z.; Duke, S.O. Transcriptional responses to cantharidin, a protein phosphatase inhibitor, in Arabidopsis thaliana reveal the involvement of multiple signal transduction pathways. Physiol. Plant. 2011, 143, 188–205. [Google Scholar] [CrossRef]
- Kagan, I.A.; Michel, A.; Prause, A.; Scheffler, B.E.; Pace, P.; Duke, S.O. Gene transcription profiles of Saccharomyces cerevisiae after treatment with plant protection fungicides that inhibit ergosterol biosynthesis. Pestic. Biochem. Physiol. 2005, 82, 133–153. [Google Scholar] [CrossRef]
- Zhu, J.; Patzoldt, W.L.; Shealy, R.T.; Vodkin, L.O.; Clough, S.J.; Tranel, P.J. Transcriptome response to glyphosate in sensitive and resistant soybean. J. Agric. Food Chem. 2008, 56, 6355–6363. [Google Scholar] [CrossRef]
- Bathke, J.; Konzer, A.; Remes, B.; McIntosh, M.; Klug, G. Comparative analyses of the variation of the transcriptome and proteome of Rhodobacter sphaeroides throughout growth. BMC Genom. 2019, 20, 358. [Google Scholar] [CrossRef] [PubMed]
- Payne, S.H. The utility of protein and mRNA correlation. Trends Biochem. 2015, 40, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Narayananan, R.; Van de Ven, W.J.M. Transcriptome and proteome analysis: A perspective on correlation. MOJ Proteom. Bioinform. 2014, 1, 000227. [Google Scholar] [CrossRef]
- Bajsa, J.; Pan, Z.; Duke, S.O. Cantharidin, a protein phosphatase inhibitor, strongly upregulates detoxification enzymes in the Arabidopsis proteome. J. Plant Physiol. 2015, 173, 33–40. [Google Scholar] [CrossRef]
- Michalski, A.; Damoc, E.; Hauschild, J.P.; Lange, O.; Wieghaus, A.; Makarov, A.; Nagaraj, N.; Cox, J.; Mann, M.; Horning, S. Mass spectrometry-based proteomics using Q Exactive, a high-performance benchtop quadrupole Orbitrap mass spectrometer. Mol. Cell. Proteom. 2011, 10, M111.011015. [Google Scholar] [CrossRef]
- Bantscheff, M.; Drewes, G. Chemoproteomic approaches to drug target identification and drug profiling. Bioorgan. Med. Chem. 2012, 20, 1973–1978. [Google Scholar] [CrossRef]
- Drewes, G.; Knapp, S. Chemoproteomics and chemical probes for target discovery. Trends Biotechnol. 2018, 36, 1275–1286. [Google Scholar] [CrossRef]
- Counihan, J.L.; Dukering, M.; Dalvie, E.; Ku, W.-M.; Bateman, L.A.; Fischer, K.J.; Nomura, D.K. Chemoproteomic profiling of acetanilide herbicides reveals their role in inhibiting fatty acid oxidation. Am. Chem. Soc. Chem. Biol. 2017, 12, 635–642. [Google Scholar] [CrossRef]
- Grossmann, K.; Christiansen, N.; Looser, R.; Tresch, S.; Hutzler, S.; Pollman, S.; Ehrhardt, T. Physionomics and metabolomics—Two key approaches in herbicides discovery mode of action discovery. Pest Manag. Sci. 2012, 68, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Pederson, H.A.; Kudsk, P.; Fomsgaard, I. Metabolic profiling of Arabidopsis thaliana reveals herbicide- and allelochemical-dependent alterations before they become apparent on plant growth. J. Plant Growth Regul. 2015, 34, 95–107. [Google Scholar] [CrossRef]
- Misra, B.B.; Das, V.; Landi, M.; Abenavoli, M.R.; Araniti, F. Short-term effects of the allelochemical umbelliferone on Triticum durum L. metabolism through GC-MS based untargeted metabolomics. Plant Sci. 2020, 298, 110548. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Taconnat, L.; Borrega, N.; Yansouni, J.; Brunaud, V.; Paysant-LeRoux, C.; Dellannoy, E.; Magniette, M.-L.M.; Lepiniec, L.; Faure, J.D.; et al. Combining laser-assisted microdissection (LAM) and RNA-seq allows to perform comprehensive transcriptomic analysis of epidermal cells of Arabidopsis embryo. Plant Methods 2018, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Naylor, A.W. Light effects on phenylalanine ammonia-lyase substrate levels and turnover rate in maize seedlings. Plant Sci. Lett. 1976, 6, 361–367. [Google Scholar] [CrossRef]
- Srivastava, A.; Kowalski, G.M.; Callahan, D.L.; Meikle, P.J.; Creek, D.J. Strategies for extending metabolomics studies with stable isotopes labeling and fluxomics. Metabolites 2016, 6, 32. [Google Scholar] [CrossRef]
- Maroli, A.S.; Nandula, V.K.; Duke, S.O.; Tharayil, N. Stable isotope resolved metabolomics reveals the role of anabolic and catabolic processes in glyphosate-induced amino acid accumulation in Amaranthus palmeri biotypes. J. Agric. Food Chem. 2016, 64, 7040–7048. [Google Scholar] [CrossRef]
- Araniti, F.; Scognamiglio, M.; Chambery, A.; Russo, R.; Esposito, A.; D’Abrosca, B.; Florentino, A.; Lupini, A.; Sunseri, F.; Abenavoli, M.R. Highlighting the effects of coumarin on adult plants of Arabidopsis thaliana (L.) Heynh. by an integrated-omic approach. J. Plant Growth Regul. 2017, 213, 30–41. [Google Scholar] [CrossRef]
- Araniti, R.; Miras-Moreno, B.; Luchini, L.; Landi, M.; Abenavoli, M.R. Metabolomic, proteomic and physiological insights into the potential mode of action of thymol, a phytotoxic natural monoterpenoid phenol. Plant Physiol. Biochem. 2020, 153, 141–153. [Google Scholar] [CrossRef]
- Böger, P.; Sandmann, G. (Eds.) Target Assays for Modern Herbicides and Related Phytotoxic Compounds; CRC Press: Boca Raton, FL, USA, 1993; p. 299. [Google Scholar]
- Dayan, F.E.; Owens, D.K.; Corniani, N.; Silva, F.M.L.; Watson, S.B.; Howell, J.; Shaner, D.L. Biochemical markers and enzyme assays for herbicide mode of action and resistance studies. Weed Sci. 2015, 63, 23–63. [Google Scholar] [CrossRef]
- Dayan, F.E.; Watson, S.B. Plant cell membrane as a marker for light-dependent and light-independent herbicide mechanisms of action. Pestic. Biochem. Physiol. 2011, 101, 182–190. [Google Scholar] [CrossRef]
- Dayan, F.E.; Rimando, A.M.; Pan, Z.; Baerson, S.R.; Gimsing, A.L.; Duke, S.O. Sorgoleone. Phytochemistry 2010, 71, 1032–1039. [Google Scholar] [CrossRef]
- Dayan, F.E.; Owens, D.K.; Watson, S.B.; Asolkar, R.N.; Boddy, L.G. Sarmentine, a natural herbicide from Piper species with multiple mechanisms of action. Front. Plant Sci. 2015, 6, 222. [Google Scholar] [CrossRef] [PubMed]
- Gressel, J. Perspective: Present pesticide discovery paradigms promote the evolution of resistance—Learn from nature and prioritize multi-target site inhibitor design. Pest Manag. Sci. 2020, 76, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P. Propesticides and their use as agrochemicals. Pest Manag. Sci. 2016, 72, 210–225. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Chinchilla, N.; Varela, R.M.; Oliveros-Bastidas, A.; Marín, D.; Molinillo, J.M.G. Structure activity relationship studies of benzoxazinones and related compounds. Phytotoxicity on Echinochloa crus-galli (L.) P. Beauv. J. Agric. Food Chem. 2005, 53, 4373–4380. [Google Scholar] [CrossRef]
- Venturelli, S.; Belz, R.G.; Kämper, A.; Berger, A.; von Horn, K.; Wegner, A.; Böcker, A.; Zabulon, G.; Langennecker, T.; Kohlbacher, O.; et al. Plants release precursors of histone deacetylase inhibitors to suppress growth of competitors. Plant Cell 2015, 27, 3175–3189. [Google Scholar] [CrossRef]
- Vaughn, K.C.; Duke, S.O. Tentoxin-induced loss of plastidic polyphenol oxidase. Physiol. Plant. 1981, 53, 421–428. [Google Scholar] [CrossRef]
- Nandula, V.K.; Reddy, K.N.; Rimando, A.M.; Duke, S.O.; Poston, D.H. Glyphosate-resistant and -susceptible soybean (Glycine fmax) and canola (Brassica napus) dose response and metabolism relationships with glyphosate. J. Agric. Food Chem. 2007, 55, 3540–3545. [Google Scholar] [CrossRef]
- Duke, S.O. Biotechnology: Herbicide-resistant crops. In Encyclopedia of Agriculture and Food Systems; Van Alfen, N., Ed.; Elsevier: San Diego, CA, USA, 2014; Volume 2, pp. 94–116. [Google Scholar]
- Yan, Y.; Liu, Q.; Zang, X.; Yuan, S.; Bat-Erdene, U.; Nguyen, C.; Gan, J.; Zhou, J.; Jacobsen, S.E.; Tang, Y. Resistance-gene-directed discovery of a natural-product herbicide with a new mode of action. Nature 2018, 559, 415–418. [Google Scholar] [CrossRef]
- Heim, D.R.; Roberts, J.L.; Pike, P.D.; Larrinua, I.M. Mutation of a locus of Arabidopsis thaliana confers resistance to the herbicide isoxaben. Plant Physiol. 1989, 90, 146–150. [Google Scholar] [CrossRef]
- Heim, D.R.; Roberts, J.L.; Pike, P.D.; Larrinua, I.M. A second locus, Ixr B1 in Arabidopsis thaliana, that confers resistance to the herbicide isoxaben. Plant Physiol. 1990, 92, 858–861. [Google Scholar] [CrossRef]
- Scheible, W.; Eshed, R.; Richmond, T.; Delmer, D.; Somerville, C. Modifications of cellulose synthase confer resistance to isoxaben and thiazolidinone herbicides in Arabidopsis Ixr1 mutants. Proc. Natl. Acad. Sci. USA 2001, 98, 10079–10084. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, K.J.; Austin, R.S.; Gong, Y.; Zhang, J.; Fung, P.; Wang, P.W.; Guttman, D.S.; Desveaux, D. Forward chemical genetic screens in Arabidopsis identify genes that influence sensitivity to the phytotoxic compound sulfamethoxazole. BMC Plant Biol. 2012, 12, 226. [Google Scholar] [CrossRef] [PubMed]
- Batool, M.; Ahmad, B.; Choi, S. A structure-based drug discovery paradigm. Int. J. Mol. Sci. 2019, 20, 2783. [Google Scholar] [CrossRef] [PubMed]
- Fleming, N. How artificial intelligence is changing drug discovery. Nature 2018, 557, S55–S57. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Du, J.; Fang, J.; Yin, Y.; Xu, G.; Xie, L. DeepScreening: A deep learning-based screening web server for accelerating drug discovery. Database 2019, 2019, baz104. [Google Scholar] [CrossRef]
- Clark, R.D. A perspective on the role of quantitative structure-activity and structure-property relationships in herbicide discovery. Pest Manag. Sci. 2012, 68, 513–518. [Google Scholar] [CrossRef]
- Sparks, T.C.; Crouse, G.D.; Demeter, D.A.; Samaritoni, G.; McLeod, C.L. Discovery of highly insecticidal synthetic mimics—CAMD enabled de novo design simplifying a complex natural product. Pest Manag. Sci. 2019, 75, 309–313. [Google Scholar] [CrossRef]
- Bryce, S.M.; Bernacki, D.T.; Bemis, J.C.; Dertinger, S.D. Genotoxic mode of action predictions from a multiplexed flow cytometric assay and a machine learning approach. Environ. Mol. Mutagen. 2016, 57, 171–189. [Google Scholar] [CrossRef]
- Drakkais, G.; Cortés-Ciriano, I.; Alexander-Dann, B.; Bender, A. Elucidating compound mechanism of action and predicting cytotoxicity using machine learning approaches, taking prediction confidence into account. Curr. Protoc. 2019, 11, e73. [Google Scholar] [CrossRef]
- Kilinc, Ö.; Reynaud, S.; Perez, L.; Tissut, M.; Ravanel, P. Physiological and biochemical modes of action of the diphenylether aclonifen. Pestic. Biochem. Physiol. 2009, 93, 65–71. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytotoxin | Chemical(s) Used for Reversal | Actual or Possible Physiological Process or Enzyme Involved | Ref. |
|---|---|---|---|
| Glyphosate 1 | aromatic amino acids | EPSPS of shikimate pathway | [39] |
| Imidazolinones 1 | branched chain amino acids | acetolactate synthase | [40] |
| Cornexistin 2 | aspartate | aspartate aminotransferase | [41] |
| Hydantocidin 2 | AMP | adenylosuccinate synthetase | [42] |
| t-chalcone 3 | homogentisate | plastoquinone synthesis | [43] |
| Rhizobitoxin 2 | methionine | β-cystathionase | [44] |
| Asulam 1 | folate or p-aminobenzoate | 7,8-dihydropteroate synthetase | [45] |
| auscaulitoxin aglycone 2 | most amino acids | amino acid transporter | [46] |
| 7-deoxy-sedoheptulose 2 | aromatic amino acids | 3-dehydroquinate synthase | [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duke, S.O.; Pan, Z.; Bajsa-Hirschel, J. Proving the Mode of Action of Phytotoxic Phytochemicals. Plants 2020, 9, 1756. https://doi.org/10.3390/plants9121756
Duke SO, Pan Z, Bajsa-Hirschel J. Proving the Mode of Action of Phytotoxic Phytochemicals. Plants. 2020; 9(12):1756. https://doi.org/10.3390/plants9121756
Chicago/Turabian StyleDuke, Stephen O., Zhiqiang Pan, and Joanna Bajsa-Hirschel. 2020. "Proving the Mode of Action of Phytotoxic Phytochemicals" Plants 9, no. 12: 1756. https://doi.org/10.3390/plants9121756
APA StyleDuke, S. O., Pan, Z., & Bajsa-Hirschel, J. (2020). Proving the Mode of Action of Phytotoxic Phytochemicals. Plants, 9(12), 1756. https://doi.org/10.3390/plants9121756