Genome-Wide Identification of the Vacuolar H+-ATPase Gene Family in Five Rosaceae Species and Expression Analysis in Pear (Pyrus bretschneideri)

 ,
,
Abstract
1. Introduction
2. Results
2.1. Identification of V-ATPase (VHA) Genes in Five Rosaceae Species
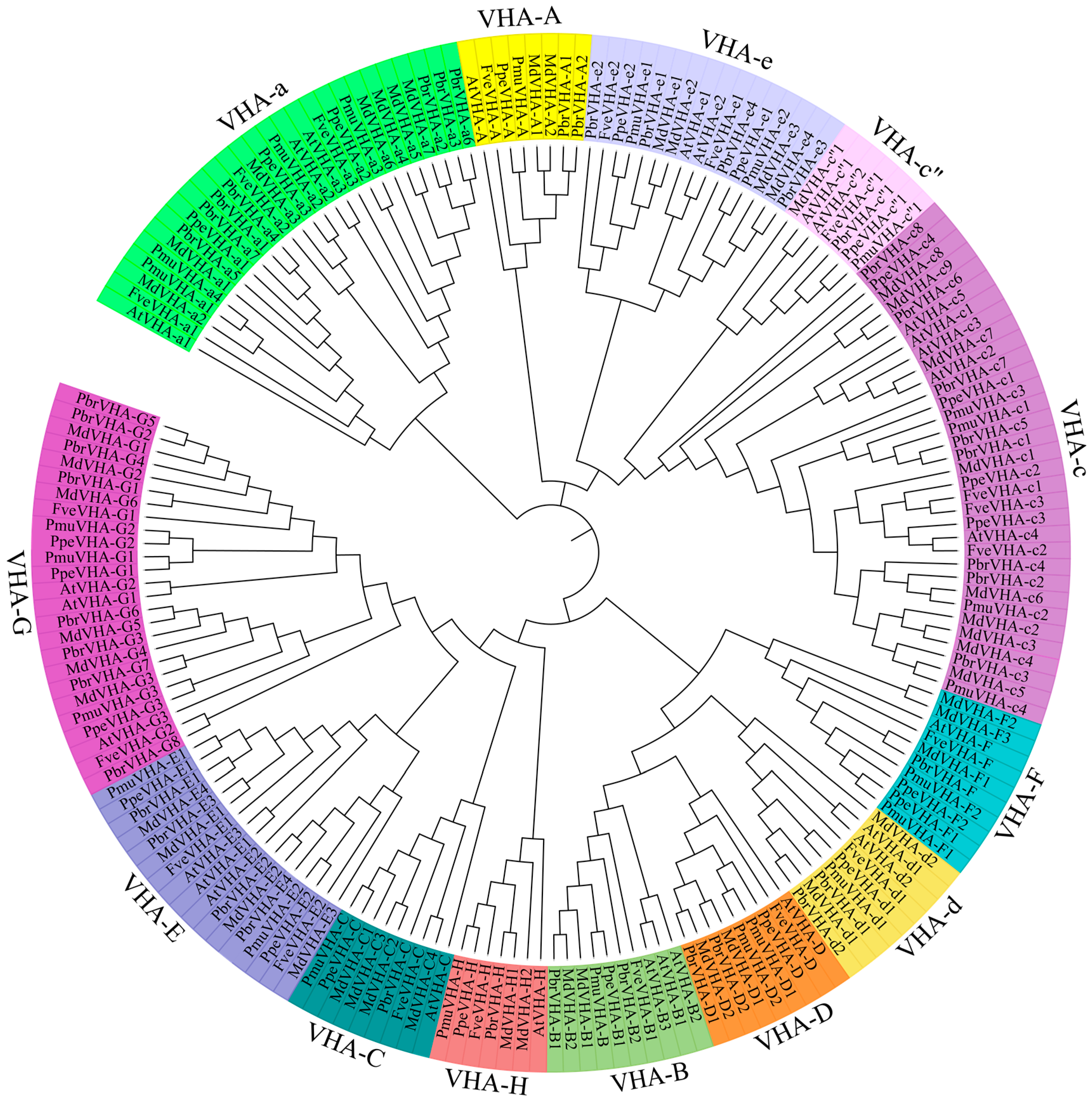
2.2. Phylogenetic Analysis and haracterisation of VHA Genes
2.3. Gene Structure, Conserved Motif, and Phylogenetic Analysis of the PbrVHA Genes
2.4. Chromosomal Distribution and Gene Duplication of PbrVHA Genes
2.5. Gene Ontology (GO) Analysis of PbrVHA Proteins
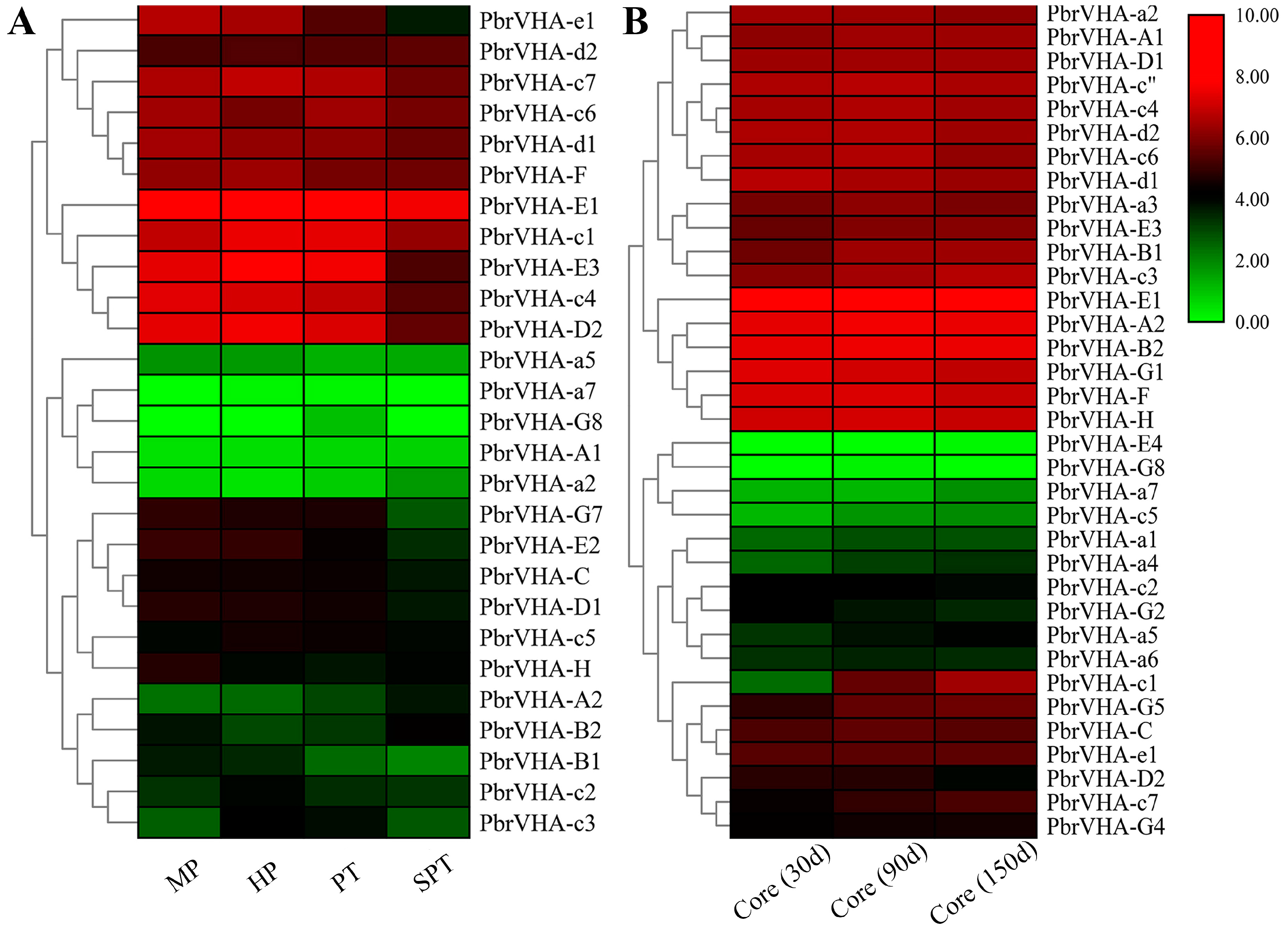
2.6. Gene Expression Patterns of PbrVHA Genes in Different Tissues and During Fruit Development
2.7. Expression Profiles of PbrVHA Genes during Pollen Tube Development and Fruit Senescence
3. Discussion
4. Materials and Methods
4.1. Identification and Characterization of VHA Genes
4.2. Phylogenetic Analysis
4.3. Gene Structure and Conserved Motif Analysis
4.4. Chromosomal Distribution and Synteny Analysis
4.5. Gene ontology (GO) Items and Expression Pattern Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shimada, T.; Takagi, J.; Ichino, T.; Shirakawa, M.; Hara-Nishimura, I. Plant Vacuoles. Annu. Rev. Plant Biol. 2018, 69, 123–145. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Li, K.; Wang, Z.; Zhu, K.; Tan, X.; Cao, J. A Review of Plant Vacuoles: Formation, Located Proteins, and Functions. Plants 2019, 8, 327. [Google Scholar] [CrossRef] [PubMed]
- Krebs, M.; Beyhl, D.; Gorlich, E.; Al-Rasheid, K.A.; Marten, I.; Stierhof, Y.D.; Hedrich, R.; Schumacher, K. Arabidopsis V-ATPase activity at the tonoplast is required for efficient nutrient storage but not for sodium accumulation. Proc. Natl. Acad. Sci. USA 2010, 107, 3251–3256. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.T.; Tang, R.J.; Zhang, Y.J.; Xue, H.W.; Ferjani, A.; Luan, S.; Lin, W.H. Two tonoplast proton pumps function in Arabidopsis embryo development. New Phytol. 2020, 225, 1606–1617. [Google Scholar] [CrossRef]
- Vasanthakumar, T.; Rubinstein, J.L. Structure and roles of V-type ATPases. Trends Biochem. Sci. 2020, 45, 295–307. [Google Scholar] [CrossRef]
- Sze, H.; Schumacher, K.; Müller, M.L.; Padmanaban, S.; Taiz, L. A simple nomenclature for a complex proton pump: VHA genes encode the vacuolar H+-ATPase. Trends Plant Sci. 2002, 7, 157–161. [Google Scholar] [CrossRef]
- Forgac, M. Vacuolar ATPases: Rotary proton pumps in physiology and pathophysiology. Nat. Rev. Mol. Cell Biol. 2007, 8, 917–929. [Google Scholar] [CrossRef]
- Saroussi, S.; Nelson, N. The little we know on the structure and machinery of V-ATPase. J. Exp. Biol. 2009, 212, 1604–1610. [Google Scholar] [CrossRef]
- Schumacher, K.; Krebs, M. V-ATPases: Rotary engines for transport and traffic. In Transporters and Pumps in Plant Signaling. Signaling and Communication in Plants; Springer: Berlin, Germany, 2011; Volume 7, pp. 293–312. [Google Scholar] [CrossRef]
- Hager, A.; Helmle, M. Properties of an ATP-fueled, Cl−1-dependent proton pump localized in membranes of microsomal vesicles from maize coleoptiles. Z. Nat. C 1981, 36, 997–1008. [Google Scholar] [CrossRef]
- Feng, S.; Peng, Y.; Liu, E.; Ma, H.; Qiao, K.; Zhou, A.; Liu, S.; Bu, Y. Arabidopsis V-ATPase d2 Subunit Plays a Role in Plant Responses to Oxidative Stress. Genes 2020, 11, 701. [Google Scholar] [CrossRef]
- Schumacher, K.; Krebs, M. The V-ATPase: Small cargo, large effects. Curr. Opin. Plant Biol. 2010, 13, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Scholl, S.; Doering, A.; Zhang, Y.; Irani, N.G.; Rubbo, S.D.; Neumetzler, L.; Krishnamoorthy, P.; Van Houtte, I.; Mylle, E.; et al. V-ATPase activity in the TGN/EE is required for exocytosis and recycling in Arabidopsis. Nat. Plants 2015, 1, 15094. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Takano, T.; Liu, S. The role of endomembrane-localized VHA-c in plant growth. Plant Signal. Behav. 2018, 13, e1382796. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Liu, E.; Ma, H.; Feng, S.; Gong, S.; Wang, J. NaCl-induced expression of AtVHA-c5 gene in the roots plays a role in response of Arabidopsis to salt stress. Plant Cell Rep. 2018, 37, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ge, Y.; Zhang, W.; Zhao, Y.; Yang, G. The walnut JrVHAG1 gene is involved in cadmium stress response through ABA-signal pathway and MYB transcription regulation. BMC Plant Biol. 2018, 18, 19. [Google Scholar] [CrossRef] [PubMed]
- Kriegel, A.; Andres, Z.; Medzihradszky, A.; Kruger, F.; Scholl, S.; Delang, S.; Patir-Nebioglu, M.G.; Gute, G.; Yang, H.; Murphy, A.S.; et al. Job sharing in the endomembrane system: Vacuolar acidification requires the combined activity of V-ATPase and V-PPase. Plant Cell 2015, 27, 3383–3396. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.Y.; Hussain, S.B.; Guo, L.X.; Yang, H.; Ning, D.Y.; Liu, Y.Z. Genome-wide identification and transcript analysis of vacuolar-ATPase genes in citrus reveal their possible involvement in citrate accumulation. Phytochemistry 2018, 155, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ji, Y.; Zhou, J.; Xing, D. Phosphatidylinositol 3-kinase promotes V-ATPase activation and vacuolar acidification and delays methyl jasmonate-induced leaf senescence. Plant Physiol. 2016. [Google Scholar] [CrossRef]
- Coker, J.S.; Jones, D.; Davies, E. Identification, conservation, and relative expression of V-ATPase cDNAs in tomato plants. Plant Mol. Biol. Rep. 2003, 21, 145–158. [Google Scholar] [CrossRef]
- Axelsen, K.B.; Palmgren, M.G. Inventory of the superfamily of P-type ion pumps in Arabidopsis. Plant Physiol. Biochem. 2001, 126, 696–706. [Google Scholar] [CrossRef]
- Kang, C.; Sun, F.; Yan, L.; Li, R.; Bai, J.; Caetano-Anolles, G. Genome-wide identification and characterization of the vacuolar H+-ATPase subunit H gene family in crop plants. Int. J. Mol. Sci. 2019, 20, 5125. [Google Scholar] [CrossRef] [PubMed]
- Farinati, S.; Rasori, A.; Varotto, S.; Bonghi, C. Rosaceae Fruit Development, Ripening and Post-harvest: An Epigenetic Perspective. Front. Plant Sci. 2017, 8, 1247. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Luo, Z.; Huang, X.; Zhang, L.; Zhao, P.; Ma, H.; Li, X.; Ban, Z.; Liu, X. Label-free quantitative proteomics to investigate strawberry fruit proteome changes under controlled atmosphere and low temperature storage. J. Proteom. 2015, 120, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, Y.; Qin, G.; Tian, S. iTRAQ-based quantitative proteomic analysis reveals the role of the tonoplast in fruit senescence. J. Proteom. 2016, 146, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, Y.; Qin, G.; Tian, S. Molecular basis of 1-methylcyclopropene regulating organic acid metabolism in apple fruit during storage. Postharvest Biol. Technol. 2016, 117, 57–63. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Z.; Shi, Z.; Zhang, S.; Ming, R.; Zhu, S.; Khan, M.A.; Tao, S.; Korban, S.S.; Wang, H. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Yang, W.; Tao, Y.; Wang, J.; Yuan, Z.; Fan, G.; et al. The genome of Prunus mume. Nat. Commun. 2012, 3, 1318. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Zhang, S.; Ma, M.; Zhang, H.; Zhang, S.; Qian, M.; Zhang, Z.; Luo, W.; Fan, J.; Liu, Z.; Wang, L. Genome-wide analysis of polygalacturonase gene family from pear genome and identification of the member involved in pear softening. BMC Plant Biol. 2019, 19, 587. [Google Scholar] [CrossRef]
- Zhang, M.; Xue, C.; Xu, L.; Sun, H.; Qin, M.; Zhang, S.; Wu, J. Distinct transcriptome profiles reveal gene expression patterns during fruit development and maturation in five main cultivated species of pear (Pyrus L.). Sci. Rep. 2016, 6, 28130. [Google Scholar] [CrossRef]
- Zhou, H.; Yin, H.; Chen, J.; Liu, X.; Zhang, S. Gene-expression profile of developing pollen tube of Pyrus bretschneideri. Gene Expr. Patterns 2016, 20, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Tian, M.; Huang, W.; Luo, S.; Hu, H.; Zhang, Y.; Zhang, L.; Li, P. Physiological and transcriptomic analysis of ‘Whangkeumbae’ pear core browning during low-temperature storage. Gene Expr. Patterns 2020, 36, 119113. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Qi, K.; Liu, X.; Yin, H.; Wang, P.; Chen, J.; Wu, J.; Zhang, S. Genome-wide identification and comparative analysis of the cation proton antiporters family in pear and four other Rosaceae species. Mol. Genet. Genom. 2016, 291, 1727–1742. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zeng, Y.; Guan, B.; Zhang, F. Overexpression of a vacuolar H+-pyrophosphatase and a B subunit of H+-ATPase cloned from the halophyte Halostachys caspica improves salt tolerance in Arabidopsis thaliana. Plant Cell Tissue Organ Cult. 2011, 108, 63–71. [Google Scholar] [CrossRef]
- Hu, D.; Sun, M.; Sun, C.; Liu, X.; Zhang, Q.; Zhao, J.; Hao, Y. Conserved vacuolar H+-ATPase subunit B1 improves salt stress tolerance in apple calli and tomato plants. Sci. Hortic. 2015, 197, 107–116. [Google Scholar] [CrossRef]
- Qiao, X.; Li, Q.; Yin, H.; Qi, K.; Li, L.; Wang, R.; Zhang, S.; Paterson, A.H. Gene duplication and evolution in recurring polyploidization–diploidization cycles in plants. Genome Biol. 2019, 20, 38. [Google Scholar] [CrossRef]
- Cao, S.; Wang, Y.; Li, X.; Gao, F.; Feng, J.; Zhou, Y. Characterization of the AP2/ERF transcription factor family and expression profiling of DREB subfamily under cold and osmotic stresses in Ammopiptanthus nanus. Plants 2020, 9, 455. [Google Scholar] [CrossRef]
- Xu, Z.; Zhao, Y.; Ge, Y.; Peng, J.; Dong, M.; Yang, G. Characterization of a vacuolar H+-ATPase G subunit gene from Juglans regia (JrVHAG1) involved in mannitol-induced osmotic stress tolerance. Plant Cell Rep. 2017, 36, 407–418. [Google Scholar] [CrossRef]
- Adem, G.D.; Roy, S.J.; Huang, Y.; Chen, Z.-H.; Wang, F.; Zhou, M.; Bowman, J.P.; Holford, P.; Shabala, S. Expressing Arabidopsis thaliana V-ATPase subunit C in barley (Hordeum vulgare) improves plant performance under saline condition by enabling better osmotic adjustment. Funct. Plant Biol. 2017, 44. [Google Scholar] [CrossRef]
- Ma, B.; Xiang, Y.; An, L. Structural bases of physiological functions and roles of the vacuolar H+-ATPase. Cell. Signal. 2011, 23, 1244–1256. [Google Scholar] [CrossRef]
- Padmanaban, S.; Lin, X.; Perera, I.; Kawamura, Y.; Sze, H. Differential expression of vacuolar H+-ATPase subunit c genes in tissues active in membrane trafficking and their roles in plant growth as revealed by RNAi. Plant Physiol. 2004, 134, 1514–1526. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Bu, Y.; Takano, T.; Zhang, X.; Liu, S. Conserved V-ATPase c subunit plays a role in plant growth by influencing V-ATPase-dependent endosomal trafficking. Plant Biotechnol. J. 2016, 14, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.X.; Shi, C.Y.; Liu, X.; Ning, D.Y.; Jing, L.F.; Yang, H.; Liu, Y.Z. Citrate accumulation-related gene expression and/or enzyme activity analysis combined with metabolomics provide a novel insight for an orange mutant. Sci. Rep. 2016, 6, 29343. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Gong, P.; Li, K.; Huang, F.; Cheng, F.; Pan, G. A single cytosine deletion in the OsPLS1 gene encoding vacuolar-type H+-ATPase subunit A1 leads to premature leaf senescence and seed dormancy in rice. J. Exp. Bot. 2016, 67, 2761–2776. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, H.; Chen, J.; Jiang, X.; Tao, S.; Wu, J.; Zhang, S. Mitochondrial dysfunction mediated by cytoplasmic acidification results in pollen tube growth cessation in Pyrus pyrifolia. Physiol. Plant. 2015, 153, 603–615. [Google Scholar] [CrossRef]
- Wang, W.; Sun, Y.-Q.; Li, G.-L.; Zhang, S.-Y. Genome-wide identification, characterization, and expression patterns of the BZR transcription factor family in sugar beet (Beta vulgaris L.). BMC Plant Biol. 2019, 19. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Walker, J.M., Totowa, N.J., Eds.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Ali, M.; Luo, D.X.; Khan, A.; Haq, S.U.; Gai, W.X.; Zhang, H.X.; Cheng, G.X.; Muhammad, I.; Gong, Z.H. Classification and Genome-Wide Analysis of Chitin-Binding Proteins Gene Family in Pepper (Capsicum annuum L.) and Transcriptional Regulation to Phytophthora capsici, Abiotic Stresses and Hormonal Applications. Int. J. Mol. Sci. 2018, 19, 2216. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.; Frank, M.; He, Y.; Xia, R. Tbtools—An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene Name Prefix | V1 Subunit | V0 Subunit | Total | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | G | H | a | c | c” | d | e | |||
| Arabidopsis thaliana | At | 1 | 3 | 1 | 1 | 3 | 1 | 3 | 1 | 3 | 5 | 2 | 2 | 2 | 28 |
| Fragaria vesca | Fve | 1 | 1 | 1 | 1 | 2 | 1 | 2 | 1 | 3 | 3 | 1 | 1 | 2 | 20 |
| Prunus persica | Ppe | 1 | 1 | 1 | 1 | 2 | 2 | 3 | 1 | 3 | 4 | 1 | 1 | 2 | 23 |
| Prunus mume | Pmu | 1 | 1 | 1 | 2 | 2 | 2 | 3 | 1 | 4 | 4 | 1 | 1 | 2 | 25 |
| Pyrus bretschneideri | Pbr | 2 | 2 | 1 | 2 | 4 | 1 | 8 | 1 | 7 | 8 | 1 | 2 | 4 | 43 |
| Malus x domestica | Md | 2 | 2 | 4 | 2 | 4 | 3 | 6 | 2 | 7 | 9 | 1 | 2 | 4 | 48 |
| Gene 1 | Gene 2 | Ka | Ks | Ka/Ks | Gene Duplication |
|---|---|---|---|---|---|
| PbrVHA-a5 | PbrVHA-a4 | 0.0386 | 0.1449 | 0.2664 | WGD |
| PbrVHA-a7 | PbrVHA-a5 | 0.0389 | 0.1461 | 0.2661 | WGD |
| PbrVHA-a7 | PbrVHA-a1 | 0.0005 | 0.0113 | 0.0479 | WGD |
| PbrVHA-B2 | PbrVHA-B1 | 0.0115 | 0.1826 | 0.0634 | WGD |
| PbrVHA-c5 | PbrVHA-c3 | 0.0054 | 1.1866 | 0.0046 | WGD |
| PbrVHA-c5 | PbrVHA-c1 | 0.0027 | 0.2052 | 0.0133 | WGD |
| PbrVHA-D1 | PbrVHA-D2 | 0.0083 | 0.2743 | 0.0306 | WGD |
| PbrVHA-E1 | PbrVHA-E3 | 0.0481 | 0.2010 | 0.2394 | WGD |
| PbrVHA-G6 | PbrVHA-G7 | 0.0275 | 0.1560 | 0.1764 | WGD |
| PbrVHA-G3 | PbrVHA-G7 | 0.0275 | 0.1560 | 0.1764 | WGD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Huang, W.; Luo, S.; Hu, H.; Zhang, Y.; Zhang, L.; Li, P. Genome-Wide Identification of the Vacuolar H+-ATPase Gene Family in Five Rosaceae Species and Expression Analysis in Pear (Pyrus bretschneideri). Plants 2020, 9, 1661. https://doi.org/10.3390/plants9121661
Zhou H, Huang W, Luo S, Hu H, Zhang Y, Zhang L, Li P. Genome-Wide Identification of the Vacuolar H+-ATPase Gene Family in Five Rosaceae Species and Expression Analysis in Pear (Pyrus bretschneideri). Plants. 2020; 9(12):1661. https://doi.org/10.3390/plants9121661
Chicago/Turabian StyleZhou, Hongsheng, Wen Huang, Shufen Luo, Huali Hu, Yingtong Zhang, Leigang Zhang, and Pengxia Li. 2020. "Genome-Wide Identification of the Vacuolar H+-ATPase Gene Family in Five Rosaceae Species and Expression Analysis in Pear (Pyrus bretschneideri)" Plants 9, no. 12: 1661. https://doi.org/10.3390/plants9121661
APA StyleZhou, H., Huang, W., Luo, S., Hu, H., Zhang, Y., Zhang, L., & Li, P. (2020). Genome-Wide Identification of the Vacuolar H+-ATPase Gene Family in Five Rosaceae Species and Expression Analysis in Pear (Pyrus bretschneideri). Plants, 9(12), 1661. https://doi.org/10.3390/plants9121661