Potassium Application Improves Grain Yield and Alleviates Drought Susceptibility in Diverse Maize Hybrids

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site and Treatments
2.2. Data Collection
3. Statistical Analysis
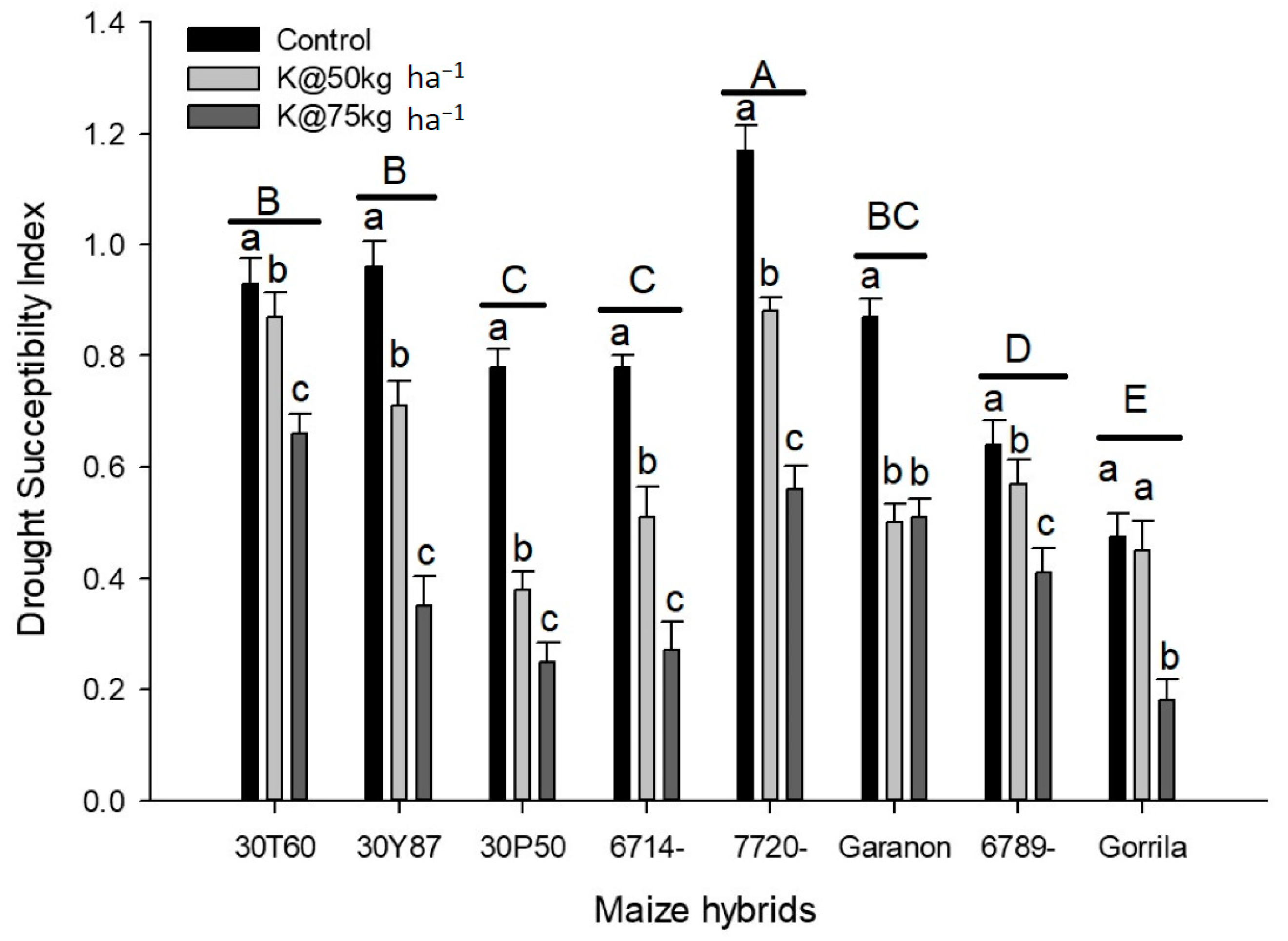
4. Results
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops that feed the world 6. Past successes and future challenges to the role played by maize in global food security. Food Secur. 2011, 3, 307. [Google Scholar] [CrossRef]
- Tesfaye, K.; Gbegbelegbe, S.; Cairns, J.E.; Shiferaw, B.; Prasanna, B.M.; Sonder, K.; Boote, K.; Makumbi, D.; Robertson, R. Maize systems under climate change in sub-Saharan Africa: Potential impacts on production and food security. Int. J. Clim. Chang. Strat. Manag. 2015, 7, 247–271. [Google Scholar] [CrossRef]
- Jin, C.; Luo, X.; Xiao, X.; Dong, J.; Li, X.; Yang, J.; Zhao, D. The 2012 Flash Drought Threatened US Midwest Agroecosystems. Chin. Geogr. Sci. 2019, 1–15. [Google Scholar] [CrossRef]
- Jin, Z.; Zhuang, Q.; Tan, Z.; Dukes, J.S.; Zheng, B.; Melillo, J.M. Do maize models capture the impacts of heat and drought stresses on yield? Using algorithm ensembles to identify successful approaches. Glob. Chang. Biol. 2016, 22, 3112–3126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hu, Z. Assessment of drought during corn growing season in Northeast China. Theor. Appl. Climatol. 2018, 133, 1315–1321. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, Z. Impact of Extreme Heat on Corn Yield in Main Summer Corn Cultivating Area of China at Present and Under Future Climate Change. Int. J. Plant Prod. 2019, 1–8. [Google Scholar] [CrossRef]
- Ul-Allah, S.; Khan, A.A.; Burkert, A.; Wachendorf, M. Socio-economic aspects of fodder production in urban and peri-urban areas of Faisalabad. Pak. J. Agric. Sci. 2014, 51, 483–490. [Google Scholar]
- Samarah, N.H.; Alqudah, A.M.; Amayreh, J.A.; McAndrews, G.M. The effect of late-terminal drought stress on yield components of four barley cultivars. J. Agron. Crop Sci. 2009, 195, 427–441. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M. Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: Growth, photosynthesis, water relations and oxidative defence mechanism. J. Agron. Crop Sci. 2011, 197, 258–271. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef]
- Ge, T.; Sui, F.; Bai, L.; Tong, C.; Sun, N. Effects of water stress on growth, biomass partitioning, and water-use efficiency in summer maize (Zea mays L.) throughout the growth cycle. Acta Physiol. Plant. 2012, 34, 1043–1053. [Google Scholar] [CrossRef]
- Aslam, M.; Zamir, M.S.I.; Afzal, I.; Yaseen, M.; Mubeen, M.; Shoaib, A. Drought stress, its effect on maize production and development of drought tolerance through potassium application. Cercet. Agron. Mold. 2013, 46, 99–114. [Google Scholar]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y. Role of mineral nutrition in alleviation of drought stress in plants. Aust. J. Crop Sci. 2011, 5, 764. [Google Scholar]
- Abbasi, H.; Jamil, M.; Haq, A.; Ali, S.; Ahmad, R.; Malik, Z. Salt stress manifestation on plants, mechanism of salt tolerance and potassium role in alleviating it: A review. Zemdirb. Agric. 2016, 103, 229–238. [Google Scholar] [CrossRef]
- Soleimani, B. Genetic Regulation of Juvenile Plant Growth under Contrasting Levels of Phosphorus, Potassium and Carbon Dioxide Nutrition in the Wild Barley Introgression Library S42IL; Martin-Luther-University: Halle-Wittenberg, Germany, 2018. [Google Scholar]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- Parker, B.; Gaines, T.P.; Gascho, G.J. Chloride effects on corn. Commun. Soil Sci. Plant Anal. 2008, 16, 1319–1333. [Google Scholar] [CrossRef]
- Christensen, N.W.; Roseberg, R.J.; Brett, M.; Jackson, T.L. Chloride inhibition of nitrification as related to take-all disease of wheat. In Special Bulletin on Chloride and Crop Production, 2nd ed.; Jackson, T.L., Ed.; Potash & Phosphate Institute: Atlanta, Georgia, 1986; pp. 22–39. [Google Scholar]
- Du, Z.; Zhou, J.; Wang, H.; Chen, X.; Wang, Q. Soil pH changes from fertilizer site as affected by application of monocalcium phosphate and potassium chloride. Commun. Soil Sci. Plant Anal. 2010, 41, 1779–1788. [Google Scholar] [CrossRef]
- Hussain, F.; Malik, A.U.; Haji, M.A.; Malghani, A.L. Growth and yield response of two cultivars of mungbean (Vigna radiata L.) to different potassium levels. J. Anim. Plant Sci. 2011, 21, 622–625. [Google Scholar]
- Anser, A.M.; Hussain, S. Nutritional and physiological significance of potassium application in maize hybrid crop production. Pak. J. Nutr. 2012, 11, 187–202. [Google Scholar]
- Singh, H.; Verma, A.; Ansari, M.W.; Shukla, A. Physiological response of rice (Oryza sativa L.) genotypes to elevated nitrogen applied under field conditions. Plant Sign. Behav. 2014, 9, e29015. [Google Scholar] [CrossRef]
- Amanullah; Iqbal, A.; Irfanullah; Hidayat, Z. Potassium management for improving growth and grain yield of maize (Zea mays L.) under moisture stress condition. Sci. Rep. 2016, 6, 34627. [Google Scholar] [CrossRef] [PubMed]
- Grzesiak, M.T.; Marcińska, I.; Janowiak, F.; Rzepka, A.; Hura, T. The relationship between seedling growth and grain yield under drought conditions in maize and triticale genotypes. Acta Physiol. Plant. 2012, 34, 1757–1764. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics: A Biological Approach; McGraw-Hill: New York, NY, USA, 1997. [Google Scholar]
- Nawaz, A.; Farooq, M.; Cheema, S.A.; Yasmeen, A.; Wahid, A. Stay green character at grain filling ensures resistance against terminal drought in wheat. Int. J. Agric. Biol. 2013, 15, 1272–1276. [Google Scholar]
- Lv, X.; Li, T.; Wen, X.; Liao, Y.; Liu, Y. Effect of potassium foliage application post-anthesis on grain filling of wheat under drought stress. Field Crops Res. 2017, 206, 95–105. [Google Scholar] [CrossRef]
- Zahoor, R.; Dong, H.; Abid, M.; Zhao, W.; Wang, Y.; Zhou, Z. Potassium fertilizer improves drought stress alleviation potential in cotton by enhancing photosynthesis and carbohydrate metabolism. Environ. Exp. Bot. 2017, 137, 73–83. [Google Scholar] [CrossRef]
- Athar, H.R.; Ashraf, M. Photosynthesis under drought stress. In Handbook of Photosynthesis, 2nd ed.; Pessarakli, M., Marcel, D., Eds.; Taylor and Francis, Inc.: New York, NY, USA, 2005; pp. 793–809. [Google Scholar]
- Negin, B.; Moshelion, M. The evolution of the role of ABA in the regulation of water-use efficiency: From biochemical mechanisms to stomatal conductance. Plant Sci. 2016, 251, 82–89. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Wheeler, R.M.; Stutte, G.W.; Levine, L.H. Low potassium enhances sodium uptake in red-beet under moderate saline conditions. J. Plant Nutr. 2000, 23, 1449–1470. [Google Scholar] [CrossRef][Green Version]
- Cerda, A.; Pardines, J.; Botella, M.A.; Martinez, V. Effect of potassium on growth, water relations, and the inorganic and organic solute contents for two maize cultivars grown under saline conditions. J. Plant Nutr. 1995, 18, 839–851. [Google Scholar] [CrossRef]
- Hussain, R.A.; Ahmad, R.; Nawaz, F.; Ashraf, M.Y.; Waraich, E.A. Foliar NK application mitigates drought effects in sunflower (Helianthus annuus L.). Acta Physiol. Plant. 2016, 38, 83. [Google Scholar] [CrossRef]
- Jan, A.U.; Hadi, F.; Nawaz, M.A.; Rahman, K. Potassium and zinc increase tolerance to salt stress in wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2017, 116, 139–149. [Google Scholar] [CrossRef]
- Zheng, Y.; Jia, A.; Ning, T.; Xu, J.; Li, Z.; Jiang, G. Potassium nitrate application alleviates sodium chloride stress in winter wheat cultivars differing in salt tolerance. J. Plant Physiol. 2008, 165, 1455–1465. [Google Scholar] [CrossRef]
- Soleimanzadeh, H.; Habibi, D.; Ardakani, M.R.; Paknejad, F.; Rejali, F. Effect of potassium levels on antioxidant enzymes and malondialdehyde content under drought stress in sunflower (Helianthus annuus L.). Am. J. Agric. Biol. Sci. 2010, 5, 56–61. [Google Scholar] [CrossRef]
- Martineau, E.; Domec, J.C.; Bosc, A.; Dannoura, M.; Gibon, Y.; Bénard, C.; Jordan-Meille, L. The role of potassium on maize leaf carbon exportation under drought condition. Acta Physiol. Plant. 2017, 39, 219. [Google Scholar] [CrossRef]
- Huber, S.C. Biochemical basis for effects of K-deficiency on assimilate export rate and accumulation of soluble sugars in soybean leaves. Plant Physiol. 1984, 76, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Hengeler, C.; Marschner, H. Changes in phloem export of sucrose in leaves in response to phosphorus, potassium and magnesium deficiency in bean plants. J. Exp. Bot. 1994, 45, 1251–1257. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Bush, D.R. Carbohydrate export from the leaf: A highly regulated process and target to enhance photosynthesis and productivity. Plant Physiol. 2011, 155, 64–69. [Google Scholar] [CrossRef]
- Lv, X.; Han, J.; Liao, Y.; Liu, Y. Effect of phosphorus and potassium foliage application post-anthesis on grain filling and hormonal changes of wheat. Field Crops Res. 2017, 214, 83–93. [Google Scholar] [CrossRef]
- White, A.C.; Rogers, A.; Rees, M.; Osborne, C.P. How can we make plants grow faster? A source–sink perspective on growth rate. J. Exp. Bot. 2015, 67, 31–45. [Google Scholar] [CrossRef]
- Ahmad, M.; Riaz, A.; Ishaque, M.; Malik, A.U. Response of maize hybrids to varying potassium application in Pakistan. Pak. J. Agric. Sci. 2009, 46, 179–184. [Google Scholar]
- Jan, M.F.; Khan, A.A.; Wahab, F.; Liaqat, W.; Ahmad, H.; Rehan, W. Phenology and productivity response of maize (Zea mays L.) hybrids to different levels of mineral potassium under semiarid climate. Middle East J. Agric. Res. 2018, 7, 287–291. [Google Scholar]
- Aslam, M.; Zamir, I.; Shahid, M.; Afzal, I.; Yaseen, M. Morphological and physiological response of maize hybrids to potassium application under drought stress. J. Agric. Res. 2013, 51, 443–454. [Google Scholar]
- Sadiq, G.; Khan, A.A.; Inamullah, A.R.; Fayyaz, H.; Naz, G.; Nawaz, H.; Ali, I.; Raza, H.; Amin, J.; Ali, S.; et al. Impact of phosphorus and potassium levels on yield and yield components of maize. Pure Appl. Biol. 2017, 6, 1071–1078. [Google Scholar] [CrossRef]
- Niu, J.; Zhang, W.; Chen, X.; Li, C.; Zhang, F.; Jiang, L.; Liu, Z.; Xiao, K.; Assaraf, M.; Imas, P. Potassium fertilization on maize under different production practices in the North China Plain. Agron. J. 2011, 103, 822–829. [Google Scholar] [CrossRef]
- Wu, L.Q.; Ma, W.Q.; Zhang, C.C.; Wu, L.; Zhang, W.F.; Jiang, R.F.; Zhang, F.S.; Cui, Z.L.; Chen, X.P. Current potassium-management status and grain-yield response of Chinese maize to potassium application. J. Plant Nutr. Soil Sci. 2013, 176, 441–449. [Google Scholar] [CrossRef]
- Wu, L.; Cui, Z.; Chen, X.; Zhao, R.; Si, D.; Sun, Y.; Yue, S. High-yield maize production in relation to potassium uptake requirements in China. Agron. J. 2014, 106, 1153–1158. [Google Scholar] [CrossRef]
- Qiu, S.; Xie, J.; Zhao, S.; Xu, X.; Hou, Y.; Wang, X.; Zhou, W.; He, P.; Johnston, A.M.; Christie, P.; et al. Long-term effects of potassium fertilization on yield, efficiency, and soil fertility status in a rain-fed maize system in northeast China. Field Crops Res. 2014, 163, 1–9. [Google Scholar] [CrossRef]
- Mallarino, A.P.; Webb, J.R.; Blackmer, A.M. Corn and soybean yields during 11 years of phosphorus and potassium fertilization on a high-testing soil. J. Prod. Agric. 1991, 4, 312-NP. [Google Scholar] [CrossRef]
- Menezes, C.B.; Ticona-Benavente, C.A.; Tardin, F.D.; Cardoso, M.J.; Bastos, E.A.; Nogueira, D.W.; Portugal, A.F.; Santos, C.V.; Schaffert, R.E. Selection indices to identify drought-tolerant grain sorghum cultivars. Genet. Mol. Res. 2014, 13, 9817–9827. [Google Scholar] [CrossRef]


{kind=link}
| Parameter | Unit | Value | Status | |
|---|---|---|---|---|
| 2017 | 2018 | |||
| Physical Parameters | ||||
| Sand | % | 61.6 | 61.4 | |
| Silt | % | 21.9 | 21.3 | |
| Clay | % | 16.5 | 16.3 | |
| Bulk Density | g cm−3 | 1.28 | 1.28 | |
| Texture Class | Sandy Loam | Sandy Loam | ||
| Chemical Parameters of Soil | ||||
| pH | 8.1 | 8.1 | ||
| EC | µS/cm | 1.9 | 1.8 | |
| Total Nitrogen | mg kg−1 | 432 | 421 | Very Low |
| Available Phosphorus | mg kg−1 | 6.9 | 6.8 | Low |
| Available Potassium | mg kg−1 | 124 | 118 | Adequate |
| Chemical Parameters of Water | ||||
| EC | µS/cm | 309 | 321 | |
| Ca+Mg | meq L−1 | 2.12 | 2.08 | |
| Na | meq L−1 | 1.21 | 1.09 | |
| HCO−3 | meq L−1 | 1.88 | 1.82 | |
| SAR | Cmolc L−1 | 1.01 | 0.96 | |
| Maize Hybrids | Source of Seed | Characteristics |
|---|---|---|
| 30Y87 | Pioneer Pakistan | Stay green character, single cross, adopted to all types of weather conditions and a yield potential of 10.4 t ha−1 |
| 30T60 | Pioneer Pakistan | Strong stem and root, single cross, reddish seeds and a yield potential of 10.9 t ha−1 |
| 30R50 | Pioneer Pakistan | Appropriate only for average environments |
| S-7720 | Syngenta Pakistan | Single cross, well suited product for high density planting environments. responds very well to high management conditions with a yield potential of 9.8 t ha−1 |
| DK-6714 | Monsanto Pakistan | Single cross, flex ear with 117 days relative maturity and a yield potential of 11.4 t ha−1 |
| DK-6789 | Monsanto Pakistan | Single cross and a yield potential of 10.4 t ha−1 |
| DK-Garanon | Monsanto Pakistan | White hybrid well adapted to diverse climatic conditions |
| Gorrila | Monsanto Pakistan | White seeded maize hybrid |
| Well-Watered | Drought | |||||||
|---|---|---|---|---|---|---|---|---|
| Hybrids | K0 | K1 | K2 | Mean (G) | K0 | K1 | K2 | Mean (G) |
| 1000-grain weight (g) | ||||||||
| 30T60 | 192.0 | 207.0 | 201.3 | 200.1 | 175.3 | 190.0 | 199.7 | 188.3 |
| 30Y87 | 181.7 | 194.7 | 207.0 | 194.5 | 186.0 | 194.0 | 196.3 | 192.1 |
| 30R50 | 185.0 | 188.3 | 196.3 | 189.9 | 173.0 | 180.7 | 165.3 | 173.0 |
| DK-6714 | 196.7 | 196.3 | 204.0 | 199.0 | 184.7 | 185.7 | 195.7 | 188.7 |
| S-7720 | 190.3 | 197.3 | 195.3 | 194.3 | 188.3 | 194.0 | 193.3 | 191.9 |
| DK-Garanon | 194.3 | 200.5 | 192.7 | 195.8 | 175.0 | 183.0 | 186.5 | 181.5 |
| DK-6789 | 187.5 | 202.5 | 190.7 | 193.6 | 182.5 | 185.0 | 174.3 | 180.6 |
| Gorrila | 187.0 | 187.5 | 202.5 | 192.3 | 181.0 | 184.5 | 193.0 | 186.2 |
| Mean (K) | 189.3B | 196.8A | 198.7A | 180.7 | 187.1 | 188.0 | ||
| HSD (p ≤ 0.05) | K = 6.7 | K = 9.7 | ||||||
| Biological Yield (t ha−1) | ||||||||
| 30T60 | 12.3bc | 12.3bc | 13.4a | 12.7A | 9.8gh | 10.3e–h | 11.2a–d | 10.4BC |
| 30Y87 | 10.5g–j | 10.7d–i | 11.6c–f | 10.9C | 8.5i | 10.3d–h | 11.4ab | 10.1CD |
| 30R50 | 10.0h–j | 10.8d–h | 10.5g–j | 10.4CD | 8.6i | 10.1f–h | 10.3d–h | 9.7D |
| DK-6714 | 13.1ab | 12.2bc | 12.4a–c | 12.6A | 10.3c–g | 11.3a–c | 11.7a | 11.1A |
| S-7720 | 11.7cd | 11.6c–e | 11.5c–g | 11.6B | 9.4hi | 10.2f–h | 10.6b–g | 10.1CD |
| DK-Garanon | 11.1d–h | 11.5c–g | 12.3bc | 11.6B | 9.9f–h | 10.7b–g | 11.2a–e | 10.6B |
| DK-6789 | 10.4h–j | 11.1d–h | 10.5f–j | 10.7C | 10.0f–h | 10.4b–g | 10.3e–h | 10.2BC |
| Gorrila | 9.7ij | 10.67e–i | 9.5j | 10.0D | 8.6i | 10.2f–h | 9.4hi | 10.1CD |
| Mean (K) | 11.1B | 11.4A | 11.5A | 9.4C | 10.4B | 10.8A | ||
| HSD (p ≤ 0.05) | G = 0.5; K = 0.3; G × K = 1.03 | G = 0.5; K = 0.20; G × K = 0.90 | ||||||
| Grain Yield (t ha−1) | ||||||||
| 30T60 | 4.9d | 5.5b | 5.8a | 5.4A | 3.0hi | 3.5d–g | 4.2b | 3.6B |
| 30Y87 | 4.5h | 4.7fg | 4.8ef | 4.7E | 2.7ij | 3.3f–h | 4.1b | 3.3CD |
| 30R50 | 3.7l | 3.8k | 3.9k | 3.8G | 2.5jk | 3.2gh | 3.5e–g | 3.1E |
| DK-6714 | 5.2c | 5.2c | 5.4b | 5.3B | 3.5e–g | 4.1b | 4.8a | 4.1A |
| S-7720 | 4.3i | 4.9de | 5.1c | 4.8D | 2.2k | 3.1hi | 3.9bc | 3.1E |
| DK-Garanon | 4.7fg | 4.8de | 5.2c | 4.9C | 3.0hi | 3.8b-e | 4.1b | 3.7B |
| DK-6789 | 4.1j | 4.6g | 4.7fg | 4.5F | 3.0hi | 3.5efg | 3.9b–d | 3.5BC |
| Gorrila | 3.5m | 3.81k | 3.9k | 3.7G | 2.9h–j | 3.1gh | 3.6c–f | 3.2DE |
| Mean (K) | 4.3C | 4.7B | 4.9A | 2.9C | 3.5B | 4.0A | ||
| HSD (p ≤ 0.05) | G = 0.06; K = 0.02; G × K = 0.13 | G = 0.19; K = 0.09; G × K = 0.40 | ||||||
| Water Productivity (kg ha−1 mm−1) | ||||||||
| 30T60 | 8.2d | 9.2b | 9.7a | 9.0A | 8.6hi | 10.0d–f | 12.0b | 10.2B |
| 30Y87 | 7.5h | 7.8fg | 8.0ef | 7.8C | 7.7ij | 9.4fgh | 11.7b | 9.6D |
| 30R50 | 6.2l | 6.3k | 6.5k | 6.3E | 7.1jk | 9.1gh | 10.0e–g | 8.8F |
| DK-6714 | 8.7c | 8.7c | 9.0b | 8.8A | 10.0e–g | 11.7b | 13.7a | 11.8A |
| S-7720 | 7.2i | 8.2de | 8.5c | 7.9C | 6.3k | 8.9hi | 11.1bc | 8.8F |
| DK-Garanon | 7.8fg | 8.0de | 8.7c | 8.2B | 8.6hi | 10.9b–d | 11.7b | 10.4B |
| DK-6789 | 6.8j | 7.7g | 7.8fg | 7.4D | 8.6hi | 10.0e–g | 11.1bc | 9.9C |
| Gorrila | 5.8m | 6.4k | 6.5k | 6.2E | 8.3h–j | 8.9gh | 10.3c–e | 9.1E |
| Mean (K) | 7.3C | 7.8B | 8.1A | 8.1C | 9.9B | 11.5A | ||
| HSD (p ≤ 0.05) | G = 0.18; K = 0.15 | G = 0.19; K = 0.17 | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ul-Allah, S.; Ijaz, M.; Nawaz, A.; Sattar, A.; Sher, A.; Naeem, M.; Shahzad, U.; Farooq, U.; Nawaz, F.; Mahmood, K. Potassium Application Improves Grain Yield and Alleviates Drought Susceptibility in Diverse Maize Hybrids. Plants 2020, 9, 75. https://doi.org/10.3390/plants9010075
Ul-Allah S, Ijaz M, Nawaz A, Sattar A, Sher A, Naeem M, Shahzad U, Farooq U, Nawaz F, Mahmood K. Potassium Application Improves Grain Yield and Alleviates Drought Susceptibility in Diverse Maize Hybrids. Plants. 2020; 9(1):75. https://doi.org/10.3390/plants9010075
Chicago/Turabian StyleUl-Allah, Sami, Muhammad Ijaz, Ahmad Nawaz, Abdul Sattar, Ahmad Sher, Muhammad Naeem, Umbreen Shahzad, Umar Farooq, Farukh Nawaz, and Khalid Mahmood. 2020. "Potassium Application Improves Grain Yield and Alleviates Drought Susceptibility in Diverse Maize Hybrids" Plants 9, no. 1: 75. https://doi.org/10.3390/plants9010075
APA StyleUl-Allah, S., Ijaz, M., Nawaz, A., Sattar, A., Sher, A., Naeem, M., Shahzad, U., Farooq, U., Nawaz, F., & Mahmood, K. (2020). Potassium Application Improves Grain Yield and Alleviates Drought Susceptibility in Diverse Maize Hybrids. Plants, 9(1), 75. https://doi.org/10.3390/plants9010075