Selenium in Germinated Chickpea (Cicer arietinum L.) Increases the Stability of Its Oil Fraction

 and
and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Physical and Chemical Properties of Oils Obtained from Chickpea Seeds and Seeds Germinated with Selenium
2.2. Fatty Acid Profiles (FAMEs) of Oils Obtained from Chickpea Seeds and Seeds Germinated with Selenium
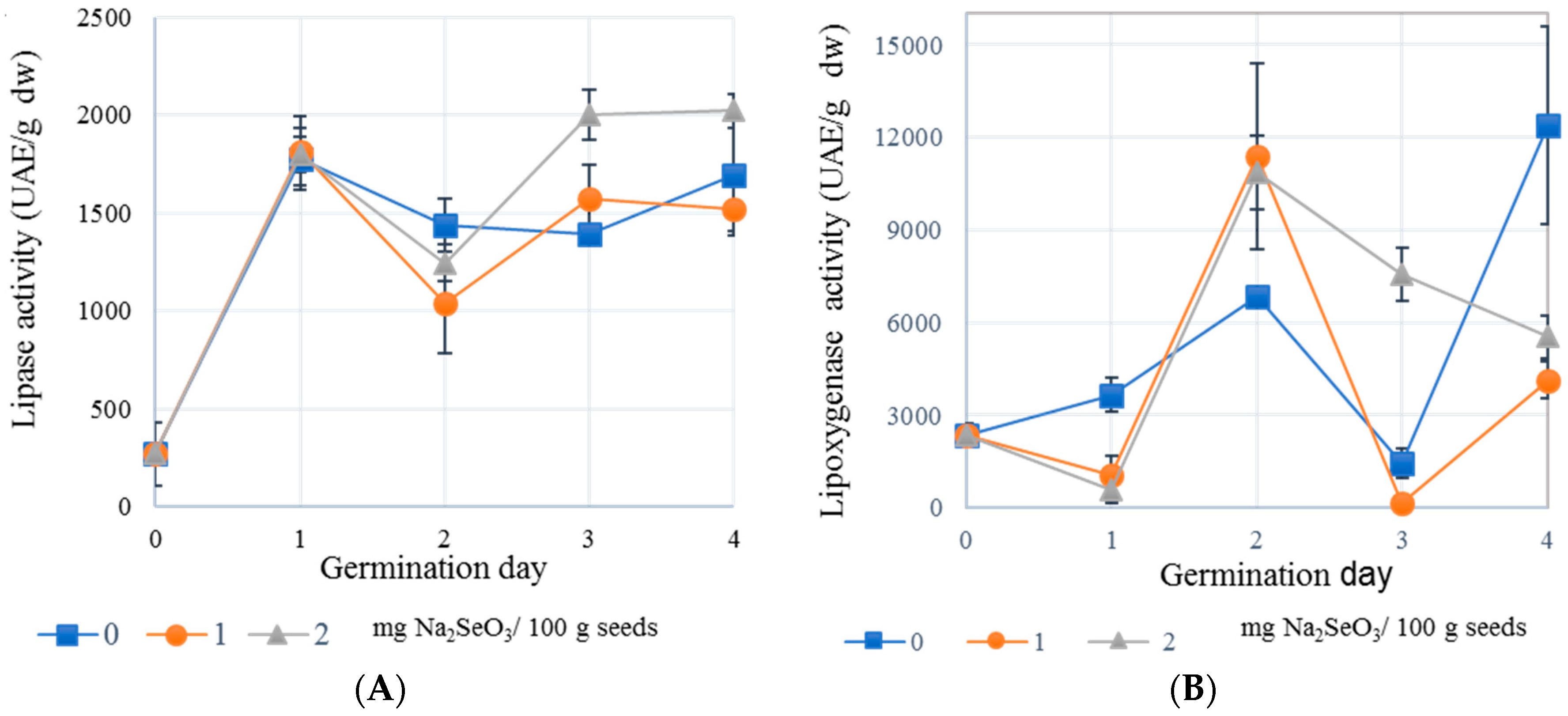
2.3. Effect of Selenium on Lipase and LOX Activities of Chickpea Sprouts
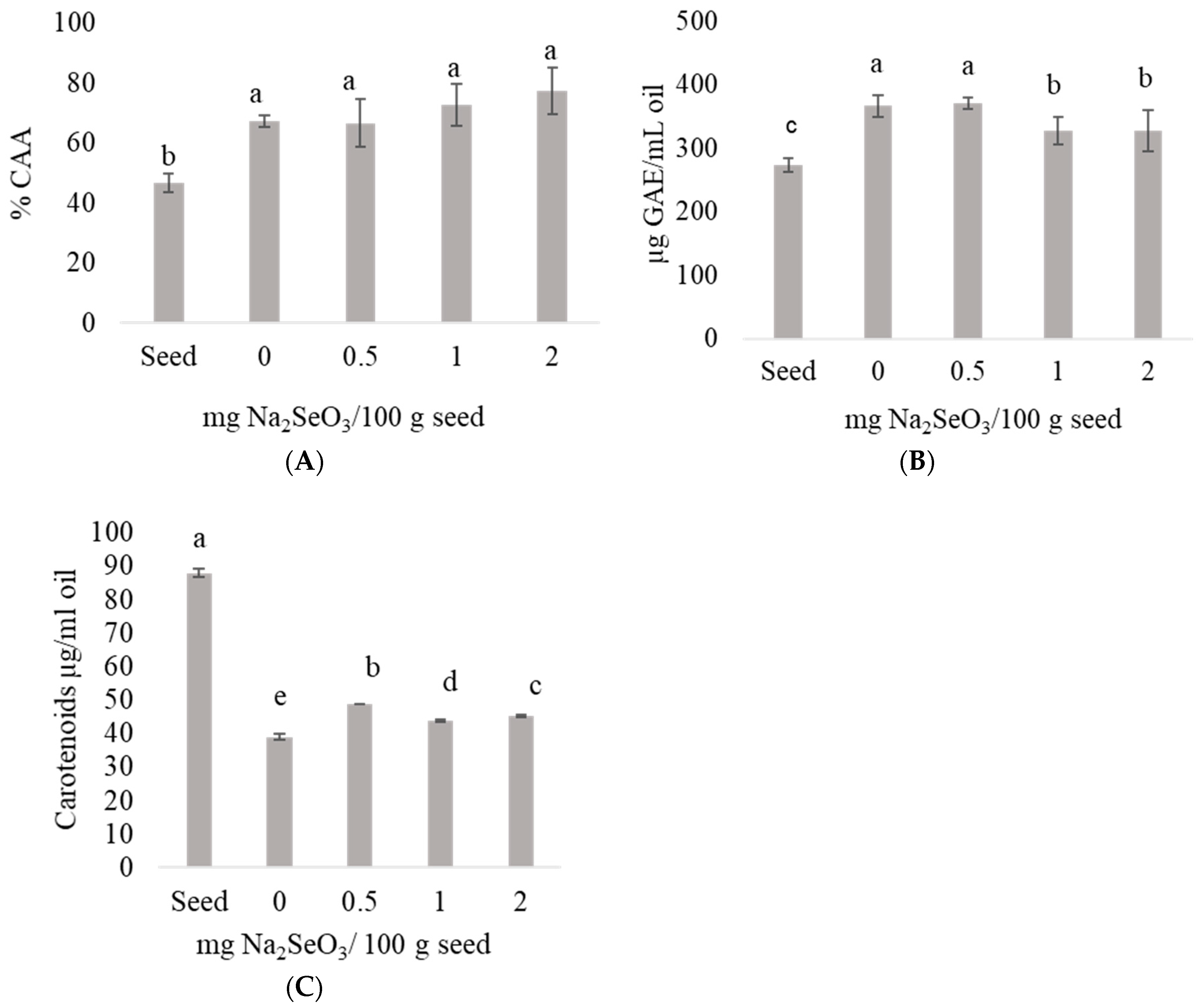
2.4. Cellular Antioxidant Activity (CAA), β-Carotenoids and Phenolic Compounds of Oils Obtained from Chickpea Seeds and Seeds Germinated with Selenium
3. Materials and Methods
3.1. Production of Chickpea Sprouts and Selenium Determination
3.2. Physical and Chemical Properties of Oils Extracted from Chickpea Seeds and Seeds Germinated with Selenium
3.3. Fatty Acid Profiles (FAMEs) of Oils Obtained from Chickpea Seeds and Seeds Germinated with Selenium
3.4. Lipase and Lipoxygenase (LOX) Activities of Chickpea Seeds and Seeds Germinated with Selenium
3.5. Cellular Antioxidant Activity (CAA) of Oils Obtained from Chickpea Seeds and Seeds Germinated with Selenium
3.6. Total Carotenoids of Oil from Chickpea Seeds and Seeds Germinated with Selenium
3.7. Total Phenolic Compounds of Oil from Chickpea Seeds and Seeds Germinated with Selenium
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Lazo-Vélez, M.A.; Chávez-Santoscoy, A.; Serna-Saldivar, S.O. Selenium-enriched breads and their benefits in human nutrition and health as affected by agronomic, milling, and baking factors. Cereal Chem. J. 2015, 92, 134–144. [Google Scholar] [CrossRef]
- Guardado-Félix, D.; Serna-Saldivar, S.O.; Cuevas-Rodríguez, E.O.; Jacobo-Velázquez, D.A.; Gutiérrez-Uribe, J.A. Effect of sodium selenite on isoflavonoid contents and antioxidant capacity of chickpea (Cicer arietinum L.) sprouts. Food Chem. 2017, 226, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Guardado-Félix, D.; Antunes-Ricardo, M.; Rocha-Pizaña, M.R.; Martínez-Torres, A.-C.; Gutiérrez-Uribe, J.A.; Serna Saldivar, S.O. Chickpea (Cicer arietinum L.) sprouts containing supranutritional levels of selenium decrease tumor growth of colon cancer cells xenografted in immune-suppressed mice. J. Funct. Foods 2019, 53, 76–84. [Google Scholar] [CrossRef]
- Frias, J.; Gulewicz, P.; Martínez-Villaluenga, C.; Pilarski, R.; Blazquez, E.; Jiménez, B.; Gulewicz, K.; Vidal-Valverde, C. Influence of germination with different selenium solutions on nutritional value and cytotoxicity of lupin seeds. J. Agric. Food Chem. 2009, 57, 1319–1325. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Mleczek, M.; Niedzielski, P.; Siwulski, M.; Gąsecka, M. Potential of Cultivated Ganoderma lucidum Mushrooms for the Production of Supplements Enriched with Essential Elements. J. Food Sci. 2016, 81, C587–C592. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Chen, X.; Luo, P.; Pei, F.; Kimatu, B.M.; Liu, K.; Du, M.; Qiu, W.; Hu, Q. The correlation between in vitro antioxidant activity and immunomodulatory activity of enzymatic hydrolysates from selenium-enriched rice protein. J. Food Sci. 2017, 82, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Lazo-Vélez, M.A.; Guajardo-Flores, D.; Mata-Ramírez, D.; Gutiérrez-Uribe, J.A.; Serna-Saldivar, S.O. Characterization and quantitation of triterpenoid saponins in raw and sprouted Chenopodium berlandieri spp. (Huauzontle) grains subjected to germination with or without selenium stress Conditions. J. Food Sci. 2016, 81, C19–C26. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, R.; De Feudis, M.; Hasuoka, P.E.; Regni, L.; Pacheco, P.H.; Onofri, A.; Businelli, D.; Proietti, P. The selenium supplementation influences olive tree production and oil stability against oxidation and can alleviate the water deficiency effects. Front. Plant Sci. 2018, 9, 1191. [Google Scholar] [CrossRef]
- Wang, N.; Daun, J. The Chemical Composition and Nutritive Value of Canadian Pulses; Laboratory, G.R., Ed.; Canadian Grain Commission: Winnipeg, MB, Canada, 2004. [Google Scholar]
- Serna-Saldívar, S.O. Cereal Grains: Properties, Processing and Nutritional Attributes, 1st ed.; CRC Press (Taylor & Francis Group): Boca Raton, FL, USA, 2010. [Google Scholar]
- Filek, M.; Keskinen, R.; Hartikainen, H.; Szarejko, I.; Janiak, A.; Miszalski, Z.; Golda, A. The protective role of selenium in rape seedlings subjected to cadmium stress. J. Plant Physiol. 2008, 165, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Jukanti, A.K.; Gaur, P.M.; Gowda, C.L.L.; Chibbar, R.N. Nutritional quality and health benefits of chickpea (Cicer arietinum L.): A review. Br. J. Nutr. 2012, 108, S11–S26. [Google Scholar] [CrossRef] [PubMed]
- Vasishtha, H.; Srivastava, R. Changes in lipids and fatty acids during soaking and germination of chickpea (Cicer arietinum). Indian Soc. Agric. Biochem. 2012, 25, 14–19. [Google Scholar]
- Khalil, A.W.; Zeb, A.; Mahmood, F.; Tariq, S.; Khattak, A.B.; Shah, H. Comparison of sprout quality characteristics of desi and kabuli type chickpea cultivars (Cicer arietinum L.). LWT Food Sci. Technol. 2007, 40, 937–945. [Google Scholar] [CrossRef]
- Duperon, P.; Delage, H.; Duperson, R. Changes in the levels of phospholipids, sterols and total fatty acids during germination of Phaseolus vulgaris seeds in the dark; relation to growth. C. R. Soc. Biol. 1968, 1792–1796. [Google Scholar]
- Mostafa, M.M.; Rahma, E.H.; Rady, A.H. Chemical and nutritional changes in soybean during germination. Food Chem. 1987, 23, 257–275. [Google Scholar] [CrossRef]
- Godswill, A.C.; Amagwula, I.O.; Victory, I.S.; Gonzaga, A.I. Effects of repeated deep frying on refractive index and peroxide value of selected vegetable oils. Int. J. Adv. Acad. Res. 2018, 4, 106–119. [Google Scholar]
- Zia-Ul-Haq, M.; Ahmad, M.; Iqbal, S.; Ahmad, S.; Ali, H. Characterization and compositional studies of oil from seeds of desi chickpea (Cicer arietinum L.) cultivars grown in Pakistan. J. Am. Oil Chem. Soc. 2007, 84, 1143–1148. [Google Scholar] [CrossRef]
- Arya, S.S.; Ramanujam, S.; Vijayaraghavan, P.K. Refractive index as an objective method for evaluation of rancidity in edible oils and fats. J. Am. Oil Chem. Soc. 1969, 46, 28–30. [Google Scholar] [CrossRef]
- Katkade, M.; Syed, H.; Andhale, R.; Sontakke, M. Fatty acid profile and quality assessment of safflower (Carthamus tinctorius) oil. J. Pharmacogn. Phytochem. 2018, 7, 3581–3585. [Google Scholar]
- Li, Y.; Liu, K.; Chen, F. Effect of selenium enrichment on the quality of germinated brown rice during storage. Food Chem. 2016, 207, 20–26. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, R.; Proietti, P.; Nasini, L.; Del Buono, D.; Tedeschini, E.; Businelli, D. Increase in the selenium content of extra virgin olive oil: quantitative and qualitative implications. Grasas y Aceites 2014, 65, 1–9. [Google Scholar] [CrossRef]
- Rahman, M.S.; Sana, N.K.; Hasan, M.M.; Huque, M.E.; Shaha, R.K. Enzyme activities and degradation of nutrients in chickpea (Cicer arietinum L.) seeds during germination. J. Bio Sci. 2008, 16, 29–34. [Google Scholar] [CrossRef][Green Version]
- Wanasundara, P.; Wanasundara, U.; Shahidi, F. Lipolytic activity of enzymes from germinating seeds of sesame (Sesamum indicum L.). J. Food Lipids 2001, 8, 75–84. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, D.; Huang, Y.; Liu, X. Effects of seed soaking with La3+ on seed germination and seedling growth of rice. Chin. J. Ecol. 2005, 24, 893–896. [Google Scholar]
- Zheng-qiu, Z. Effect INTECof heavy metals Cr and Cu on seeds germination and lipase activity of peanut. J. Anhui Agric. Sci. 2009, 2009. [Google Scholar]
- Deshpande, T.; Padma, A.S.; Sukumaran, M.K.; Rajani, D. Enzymatic activity levels of lipoxygenase (LOX) in germinating green gram seeds (Vigna radiate). Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 621–628. [Google Scholar]
- Chedea, V.S.; Jisaka, M. Inhibition of soybean lipoxygenases—Structural and activity models for the lipoxygenase isoenzymes family. In Recent Trends for Enhancing the Diversity and Quality of Soybean Products; Krezhova, D., Ed.; IntechOpen Limited: London, UK, 2011; pp. 109–130. [Google Scholar]
- Rock, C.; Moos, P.J. Selenoprotein P protects cells from lipid hydroperoxides generated by 15-LOX-1. Prostaglandins Leukot. Essent. Fat. Acids 2010, 83, 203–210. [Google Scholar] [CrossRef] [PubMed]
- da Marineli, R.S.; Moraes, É.A.; Lenquiste, S.A.; Godoy, A.T.; Eberlin, M.N.; Maróstica, M.R., Jr. Chemical characterization and antioxidant potential of Chilean chia seeds and oil (Salvia hispanica L.). LWT Food Sci. Technol. 2014, 59, 1304–1310. [Google Scholar] [CrossRef]
- Siger, A.; Nogala-Kalucka, M.; Lampart-Szczapa, E. The content and antioxidant activity of phenolic compounds in cold-pressed plant oils. J. Food Lipids 2008, 15, 137–149. [Google Scholar] [CrossRef]
- Perera, C.O.; Yen, G.M. Functional properties of carotenoids in human health. Int. J. Food Prop. 2007, 10, 201–230. [Google Scholar] [CrossRef]
- Boon, C.S.; McClements, D.J.; Weiss, J.; Decker, E.A. Factors influencing the chemical stability of carotenoids in foods. Crit. Rev. Food Sci. Nutr. 2010, 50, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Liu, P. Inhibition of angiotensin I―converting enzymes by enzymatic hydrolysates from chicken blood. J. Food Drug Anal. 2010, 10, 458–463. [Google Scholar]
- Crude Fat in Grain and Stock Feeds. In Approved Methods of Analysis, 11th ed.; AACCI Method 30-20.01; AACCI: St. Paul, MN, USA, 1999.
- Paquot, C. Standard Methods for the Analysis of Oils, Fats and Derivatives, 6th ed.; Elsevier Ltd: Amsterdam, The Netherlands, 1979. [Google Scholar]
- AOCS. Official Methods and Recommended Practices of the AOCS, 5th ed.; Firestone, D., Ed.; AOCS Press: Champaign, IL, USA, 1999. [Google Scholar]
- Esquivel-Hernández, D.A.; López, V.H.; Rodríguez-Rodríguez, J.; Alemán-Nava, G.S.; Cuéllar-Bermúdez, S.P.; Rostro-Alanis, M.; Parra-Saldívar, R. Supercritical carbon dioxide and microwave-assisted extraction of functional lipophilic compounds from Arthrospira platensis. Int. J. Mol. Sci. 2016, 17, 658. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Glatz, C.E. Enzymatic assay method for evaluating the lipase activity in complex extracts from transgenic corn seed. J. Agric. Food Chem. 2006, 54, 3181–3185. [Google Scholar] [CrossRef] [PubMed]
- Sosulski, F.W.; Gadan, H.M. Variations in lipid composition among chickpea cultivars. J. Am. Oil Chem. Soc. 1988, 65, 369–372. [Google Scholar] [CrossRef]
- Wan, H.; Liu, D.; Yu, X.; Sun, H.; Li, Y. A Caco-2 cell-based quantitative antioxidant activity assay for antioxidants. Food Chem. 2015, 175, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Ranjith, A.; Kumar, K.S.; Venugopalan, V.V.; Arumughan, C.; Sawhney, R.C.; Singh, V. Fatty acids, tocols, and carotenoids in pulp oil of three sea buckthorn species (Hippophae rhamnoides, H. salicifolia, and H. tibetana) grown in the Indian Himalayas. J. Am. Oil Chem. Soc. 2006, 83, 359–364. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M.B.T.-M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Oxidants and Antioxidants Part A; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]



{kind=link}
{kind=link}
| mg Na2SeO3/100 g Seeds | Oil Content (%) | Refractive Index (RI, 20 °C) | Acid Value (AV, % as Linoleic Acid) | Oxidative Stability Index (OSI, h) 2 |
|---|---|---|---|---|
| 0.0 | 6.9 ± 0.19 a | 1.468 ± 0.0002 a | 12.4 ± 0.06 b | 11.4 ± 1.52 b |
| 0.5 | 6.7 ± 0.02 b | 1.467 ± 0.0002 b | 15.6 ± 0.24 a | 14.6 ± 1.42 b |
| 1.0 | 6.8 ± 0.01 ab | 1.466 ± 0.0002 b | 12.3 ± 0.28 b | 16.7 ± 3.05 b |
| 2.0 | 6.8 ± 0.04 ab | 1.466 ± 0.0003 b | 12.0 ± 0.01 b | 13.1 ± 1.90 b |
| Seed | 5.5 ± 0.29 c | 1.467 ± 0.0007 ab | 5.0 ± 0.44 c | 31.8 ± 1.88 a |
| mg Na2SeO3/100 g Seeds | Fatty Acids, % | |||
|---|---|---|---|---|
| Palmitic C16:0 | Stearic C18:0 | Oleic C18:1 | Linoleic C18:2 | |
| 0.0 | 8.1 | 7.8 | 25.5 | 58.6 |
| 0.5 | 8.3 | 7.7 | 25.0 | 59.0 |
| 1.0 | 8.3 | 7.6 | 25.4 | 58.7 |
| 2.0 | 8.5 | 7.7 | 25.5 | 58.3 |
| Seed | 8.4 | 7.9 | 26.5 | 57.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guardado-Félix, D.; Serna-Saldivar, S.O.; Gutiérrez-Uribe, J.A.; Chuck-Hernández, C. Selenium in Germinated Chickpea (Cicer arietinum L.) Increases the Stability of Its Oil Fraction. Plants 2019, 8, 113. https://doi.org/10.3390/plants8050113
Guardado-Félix D, Serna-Saldivar SO, Gutiérrez-Uribe JA, Chuck-Hernández C. Selenium in Germinated Chickpea (Cicer arietinum L.) Increases the Stability of Its Oil Fraction. Plants. 2019; 8(5):113. https://doi.org/10.3390/plants8050113
Chicago/Turabian StyleGuardado-Félix, Daniela, Sergio O. Serna-Saldivar, Janet A. Gutiérrez-Uribe, and Cristina Chuck-Hernández. 2019. "Selenium in Germinated Chickpea (Cicer arietinum L.) Increases the Stability of Its Oil Fraction" Plants 8, no. 5: 113. https://doi.org/10.3390/plants8050113
APA StyleGuardado-Félix, D., Serna-Saldivar, S. O., Gutiérrez-Uribe, J. A., & Chuck-Hernández, C. (2019). Selenium in Germinated Chickpea (Cicer arietinum L.) Increases the Stability of Its Oil Fraction. Plants, 8(5), 113. https://doi.org/10.3390/plants8050113