1. Introduction
The air pollutant ozone (O
3) reduces yield in many crops including wheat, rice, and soybean [
1,
2]. Ozone at the ground level forms from precursor gases—chiefly NO
x and volatile organic compounds (VOCs)—in chemical reactions catalysed by sunlight and heat [
3]. Concentrations over much of the Earth’s land surface have approximately doubled since pre-industrial times, mainly due to anthropogenic emissions from vehicles, industry, and agriculture [
4,
5,
6]. Annual mean surface O
3 concentrations have largely stabilized in Europe since the year 2000 as a result of emission control policies [
7,
8], but continued increase to 2050 is likely across South and East Asia [
9,
10]. The pattern of O
3 exposure across regions is also expected to change over coming decades. Short peak ‘episodes’ of very high concentrations are predicted to become more frequent in India and China [
10,
11], while in Europe and North America a decline in peak episode frequency, alongside steadily increasing annual mean O
3 concentrations, was observed between 1990 and 2010 [
12]. Modelling suggests this decline in peak episode frequency in Europe and North America is likely to continue to 2050 [
10].
Model-based estimates of yield loss under future climate and air pollution scenarios represent a powerful way of highlighting yield benefits that could come from reduced surface O
3 [
13,
14,
15,
16]. Global O
3-induced wheat yield loss for the year 2000 has been estimated as ranging from 5% to 26%, with potential additional losses of 1.5% to 10% predicted for 2030 [
13,
14]. However, nearly all large-scale assessments of O
3-induced yield loss for wheat published to date have followed an empirical approach, where O
3 concentration is simulated using a chemical transport model (CTM), concentration is linked to yield loss using response functions, and response is scaled up using crop production maps and agricultural statistics. An alternative is to use a process-based approach that could potentially produce more robust estimates of future yield, through inclusion of interactive effects between O
3, CO
2, and climate variables [
17].
This process-based crop modelling approach builds on the development of methods for modelling O
3 flux into leaves that have been made over recent decades [
18,
19]. These methods provide an hourly estimate of O
3 dose reaching sites of damage in the leaf, creating the potential for O
3 effects to be integrated into crop simulation models in a dynamic way. Studies applying O
3 flux modelling have generally either used a multiplicative stomatal conductance (g
sto) algorithm—described in prior research [
20,
21,
22]—or followed a semi-mechanistic approach where g
sto is estimated empirically from photosynthetic rate, which in turn is modelled using the biochemical model of Farquhar et al. [
23], also described in [
24,
25]. Since most crop models simulate growth responses at daily (and less frequently, hourly) timesteps and can respond to a changing environment [
26], integration of O
3 effects into crop models is feasible, if plant response to O
3 can be represented in the model formulation. Attempts have been made to integrate O
3 effects and crop modelling [
25,
27], but few estimates of O
3-induced crop yield loss using a dynamic approach have been published to date [
28,
29]. Reasons for slow progress include the challenge of upscaling responses from the leaf to the canopy, the need for species and cultivar-specific model parameters, and the incomplete understanding of physiological mechanisms driving O
3-induced yield reduction [
17]. This paper goes some way to addressing these issues and identifying future research direction that would benefit from empirical investigation targeted towards developing the models that are currently in development.
It is well established that O
3 exposure can reduce yield in wheat [
30,
31,
32] and can cause foliar injury, impaired photosynthesis, altered carbon translocation, and accelerated senescence [
33,
34]. However, the processes linking O
3 uptake to these responses are not fully understood and it is not clear which are most important in driving ultimate yield loss. Once O
3 has been taken up through stomata, reactions in the plant apoplast lead to the formation of reactive oxygen species (ROS), which can then react with and damage membranes and proteins [
35]. Most plants have in-built defence mechanisms and can up-regulate antioxidants to detoxify ROS, but this comes at a carbon cost, meaning O
3 damage to productivity often occurs before visible symptoms appear [
35,
36]. An O
3-induced reduction in photosynthetic rate has been widely reported [
32,
37,
38], but quantifying the extent to which this is a direct effect of O
3 on the photosynthetic mechanism, or indirect via changes to leaf pigmentation or g
sto, has been a challenge for experimentalists. Disentangling direct O
3 impacts on photosynthesis from the accelerated senescence response is also difficult. Some studies have observed reduced activity of the carbon-fixing enzyme ribulose-1,5-biphosphate carboxylase/oxygenase (rubisco) in response to O
3 [
39,
40,
41], leading to the hypothesis that ‘instantaneous’ effects of O
3 on photosynthesis act via effects on this enzyme. The physiological mechanism underpinning the often-observed accelerated senescence response to O
3 [
42,
43,
44,
45,
46] is also unknown, although it has been hypothesized that it relates to long-term respiratory costs associated with detoxification and repair [
25].
Several approaches for modelling O
3 effects on photosynthesis and senescence have been published. In an early paper, Reich et al. proposed that ozone effects on a number of plant groups could be expressed via a linear relationship between exposure and growth [
47]. Subsequent published approaches have attempted to model the separate effects of O
3 on productivity and senescence and have tried to account for differential sensitivity across species. A function for modelling ‘instantaneous’ suppression of photosynthesis was proposed by Martin et al. [
27], who simulated a linear reduction in carboxylation capacity of rubisco (the parameter V
cmax in the model of Farquhar et al. [
23]), above a threshold hourly flux value representative of the species or cultivar-specific detoxification capacity. A similar approach of O
3 acting on V
cmax was also used by Deckmyn et al. in their O
3 damage module for forest trees, alongside an overnight repair mechanism and a parameter representing the respiratory cost of detoxification [
48]. Ewert and Porter [
25] applied a version of the Martin et al. ‘short-term’ function alongside a ‘long-term’ algorithm for modelling O
3-induced senescence and assumed that ‘short-term’ photosynthetic suppression by O
3 occurs throughout the leaf lifespan. Their senescence function assumes a linear reduction in mature leaf lifespan as accumulated O
3 flux increases, and senescence comprises the final third of the mature leaf lifespan, during which time V
cmax is assumed to decline linearly. In this function, onset and completion of leaf senescence therefore move progressively earlier and closer together as accumulated O
3 flux increases. An alternative approach for modelling O
3-induced senescence is applied in the multiplicative DO
3SE model (Deposition of ozone for stomatal exchange), a g
sto model which estimates accumulated O
3 flux—known as the Phytotoxic Ozone Dose (POD)—to vegetation [
18]. In this model, leaf senescence is induced by a threshold POD, which triggers curvilinear decline in leaf g
sto with a fixed shape but variable decline rate [
20,
49,
50]. The POD ‘trigger’ can be parameterized according to the sensitivity of the cultivar or species.
The integration of O
3 damage functions, such as those described above, into crop models could improve yield estimates under O
3 stress. Model development must however be guided by experimental evidence that identifies damage mechanisms and their relative importance, informs parameterization, and indicates likely degree of error. Models must also be able to replicate the response to different patterns of O
3 exposure. For example, acute peaks in concentration have been observed to induce greater yield loss than consistent, moderate levels with the same 24-h mean exposure [
51], and modelling methods need to be able to capture these nuances.
This study combines data from two independent O3 exposure experiments on European wheat that took place at the same experimental facility in 2015 and 2016 and used consistent protocols for data collection. We model the accumulated O3 flux to wheat flag leaves across different O3 treatments, using the multiplicative DO3SE model, to derive the POD0SPEC metric of O3 flux exposure (mmol m−2 PLA day−1). Exposure to O3 flux is then considered alongside leaf chlorophyll, gsto, and photosynthesis responses in order to test key assumptions underpinning published O3 effect model functions. Firstly, with regards to O3 effects on leaf senescence, we (i) examine whether inter-cultivar differences in response are captured by current senescence functions and (ii) whether leaf senescence begins at an accumulated O3 flux ‘trigger’ value. Secondly, we examine whether O3 reduces Vcmax before—and therefore independent of—onset of O3-induced leaf senescence. Thirdly, we investigate if flux is a better predictor of the physiological response to O3 than concentration-based metrics and whether flux can account for differences in the pattern of O3 exposure (i.e., peak vs. background).
Our results reveal several insights about physiological responses to O3 that can add to the evidence base for designing O3 effect model functions. The two cultivars of European wheat studied here showed substantially different sensitivity to O3 in terms of their senescence response, indicating the importance of cultivar-specific parameterization in senescence functions. The study also finds that O3 effects on the photosynthetic mechanism are not observed in young flag leaves and are only seen following the onset of leaf senescence, suggesting that O3-induced accelerated senescence is more important than direct effects on photosynthesis in determining final yield.
3. Discussion
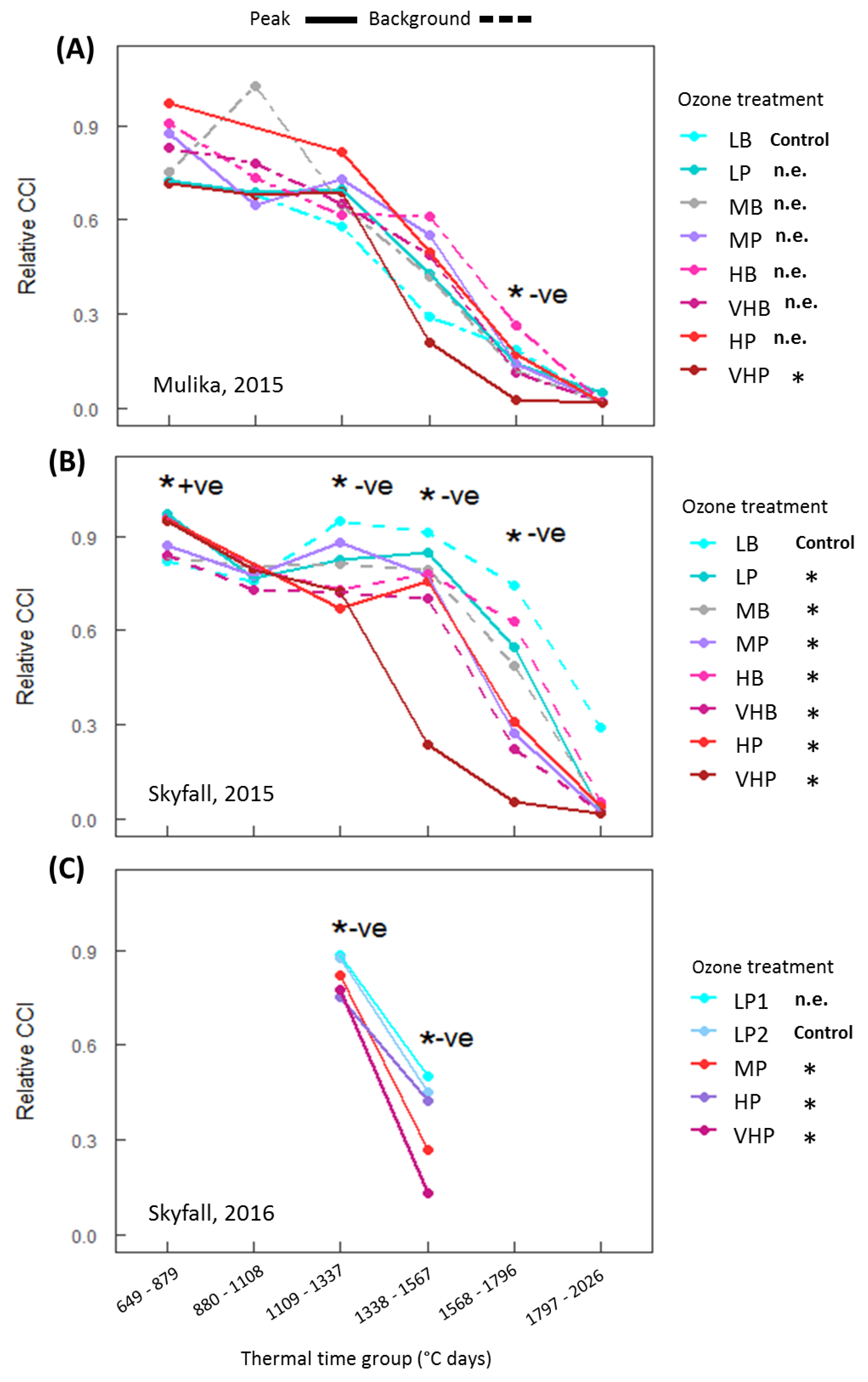
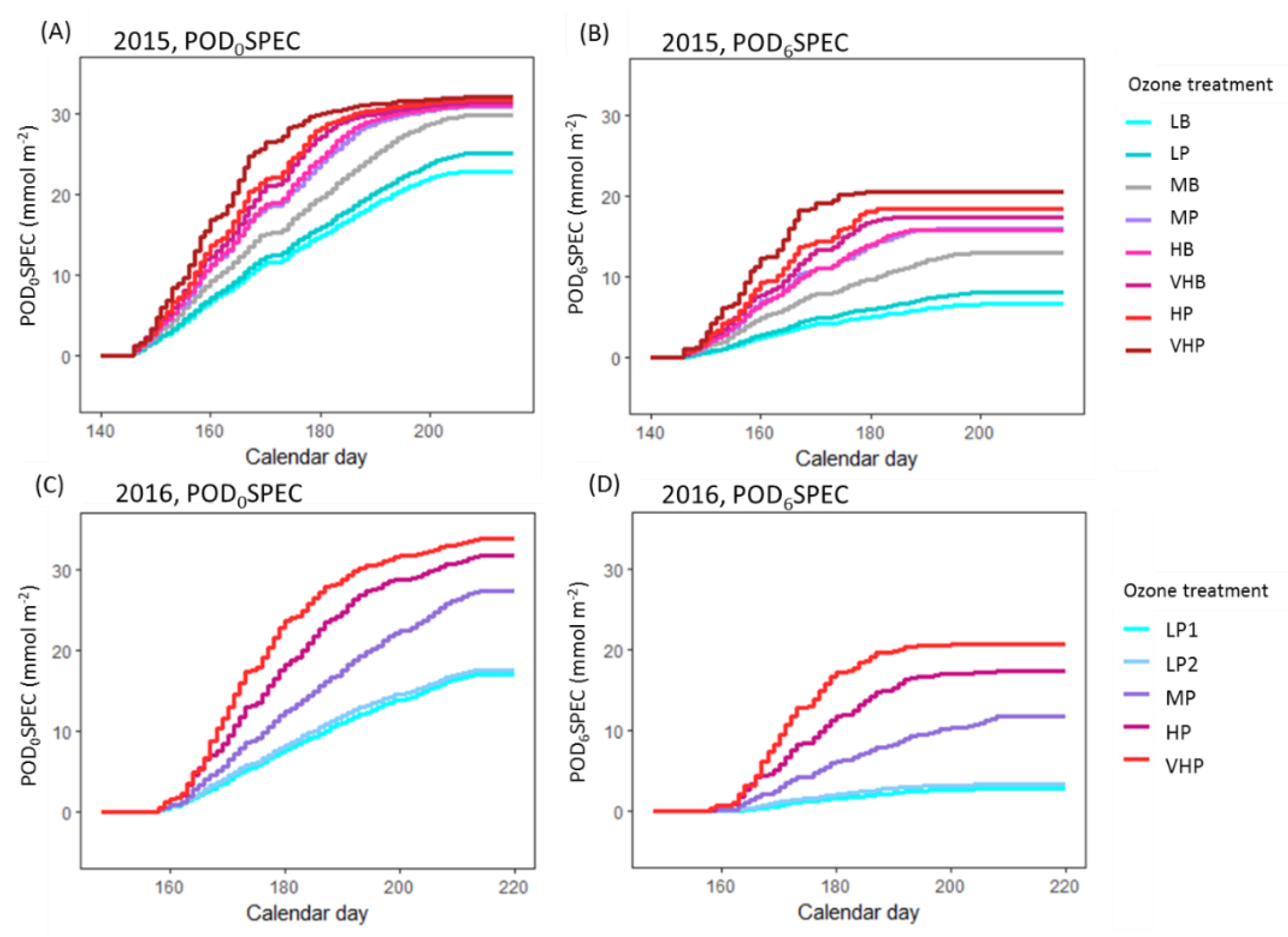
The first aim of the analysis presented here was to assess whether published approaches for modelling O
3-induced senescence can account for inter-cultivar variation in response. Both cultivars exhibited accelerated senescence in response to O
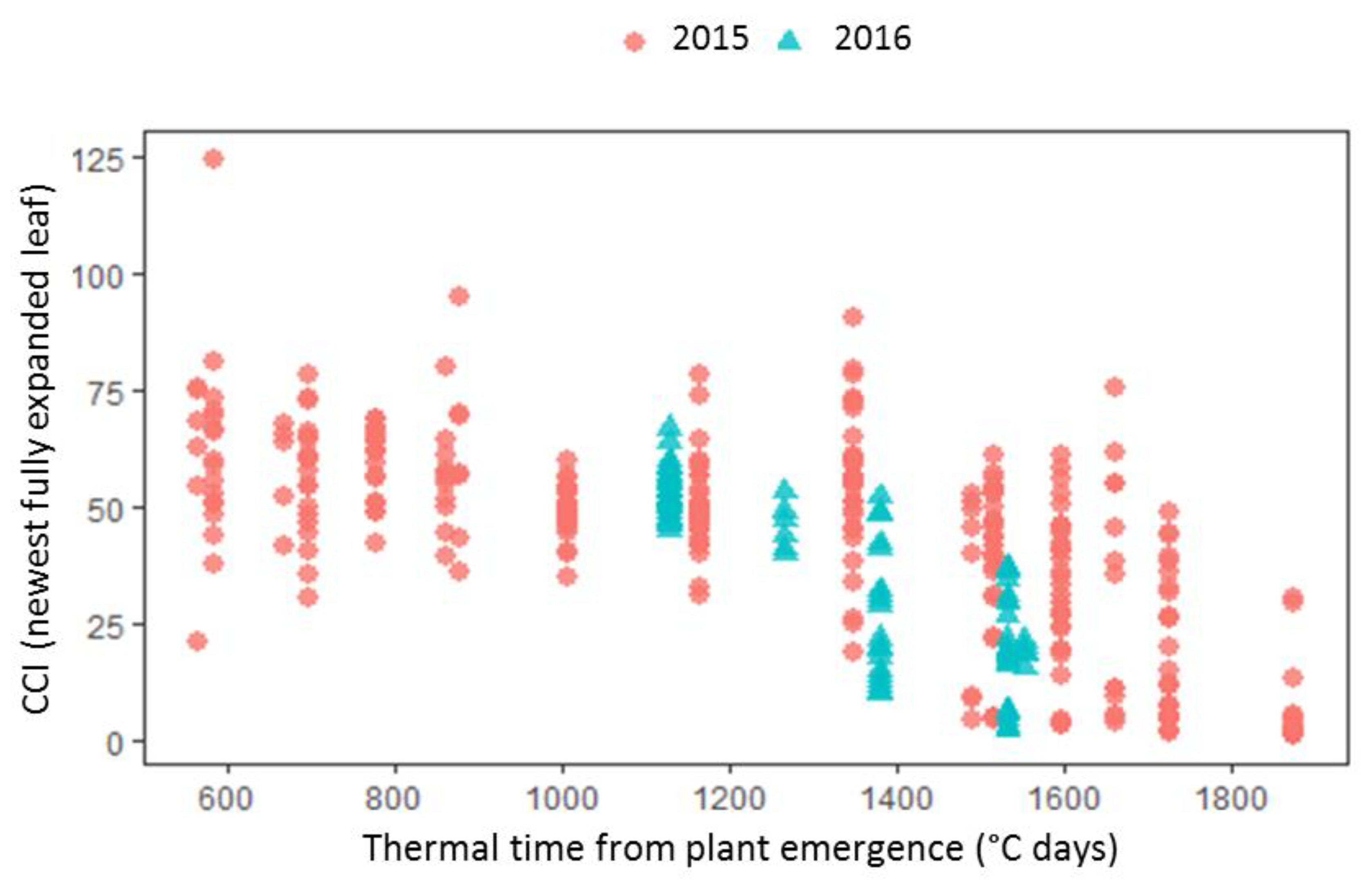
3, but the pattern of response differed according to cultivar. In 2015, significant accelerated senescence was observed in seven O
3 treatments for Skyfall, but only in the highest treatment for Mulika, suggesting higher O
3 tolerance in Mulika (
Figure 1). This differential tolerance is also indicated by the earlier appearance of significant O
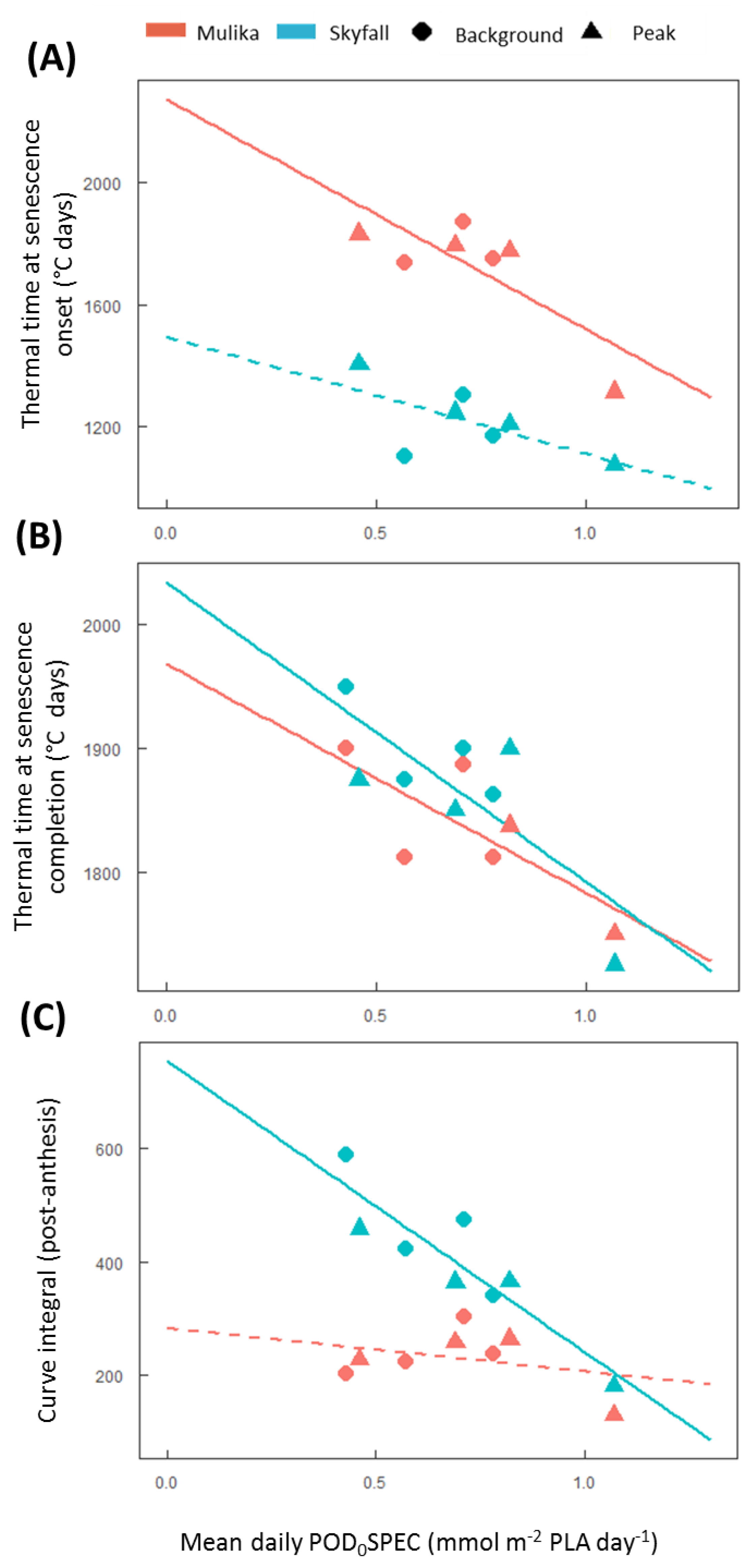
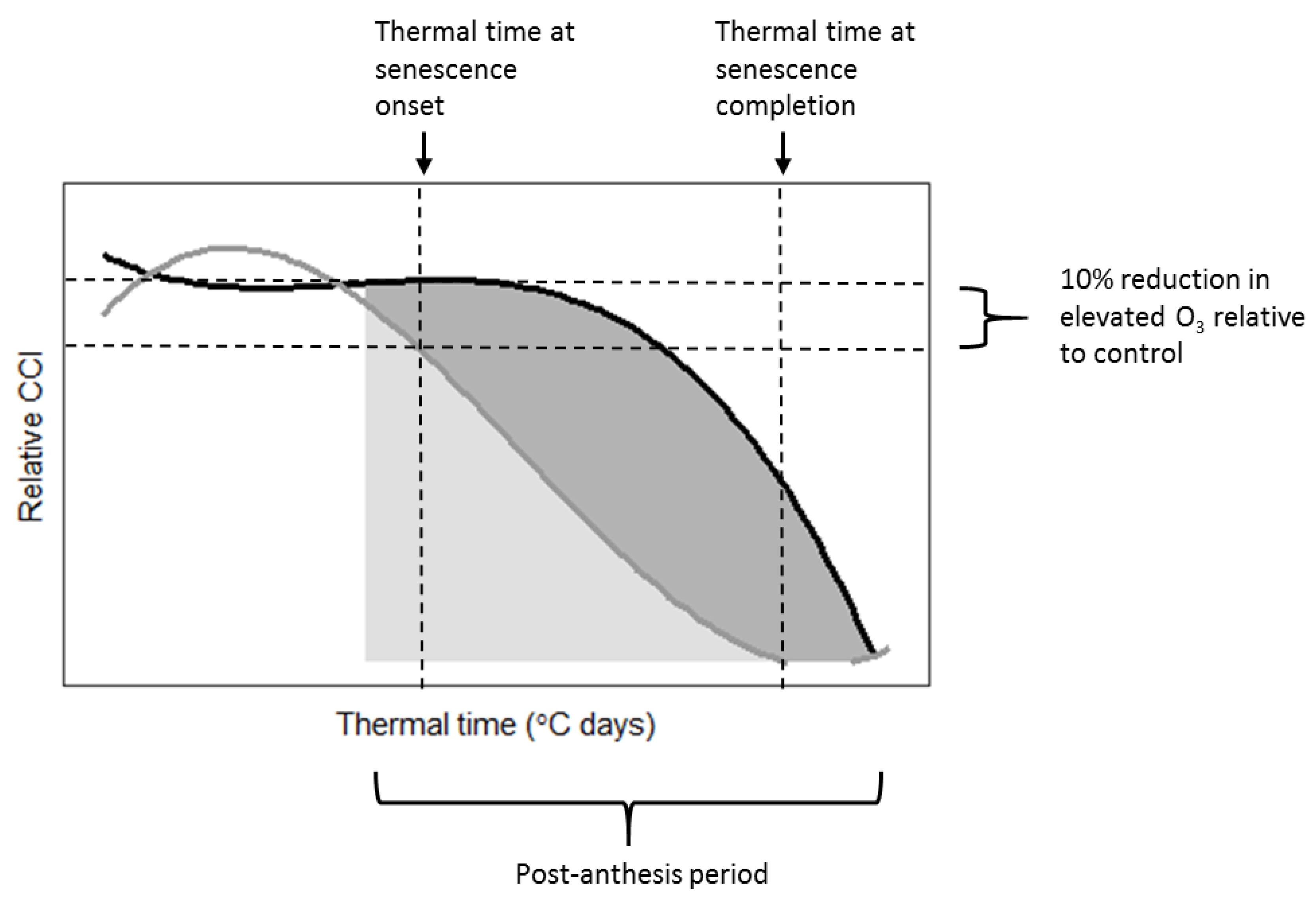
3 effects on leaf CCI across all treatments for Skyfall compared to Mulika. Senescence completion occurred progressively earlier, hence total leaf duration became progressively shorter, in both cultivars as average O
3 flux (mean daily POD
0SPEC) in the treatment increased (
Figure 2B), according to a linear relationship. Completion of leaf senescence occurred at a similar thermal time in both cultivars (Mulika = 1841 °C days, Skyfall = 1867 °C days), meaning that the total senescence duration was longer for Skyfall. While O
3-induced senescence in Mulika was characterised by a sudden drop in leaf CCI in the late-season, Skyfall exhibited a more gradual O
3-induced decline in CCI.
The linear relationship between mean flux and total leaf duration observed in this study for both cultivars gives support to the senescence function of Ewert and Porter [
25], which assumes a linear decline in mature leaf lifespan as O
3 exposure increases. However, evidence presented here suggests that the duration of leaf senescence is likely to vary with O
3 exposure. Additionally, the differential senescence duration in the two cultivars suggests that a key assumption of the Ewert and Porter function—that leaf senescence comprises the final third of the mature leaf lifespan—may not hold true for all cultivars. For example, in 2015 for Skyfall, leaf senescence in the highest O
3 treatment comprised 76.7% of the total flag leaf lifespan (flag leaf emergence = 877 °C days, leaf senescence onset = 1075 °C days, senescence completion = 1725 °C days)—substantially greater than one third. The inter-cultivar variation in senescence response observed in this study would therefore only be captured by a model function that allows for the proportion of leaf lifespan comprising leaf senescence to be parameterised according to cultivar and ozone exposure. Our results suggest that to effectively model variation in the pattern of O
3-induced senescence, the timing of senescence onset, and the rate (or duration) of senescence, models need to be calibrated for particular species and cultivars.
However, the analysis in this paper highlights the uncertainty associated with the approach of Danielsson et al. [
20] for modelling onset of O
3-induced senescence using a threshold of accumulated flux. Following this approach, senescence onset may occur at different points in time at different levels of mean exposure but should occur at approximately the same value of accumulated flux. This method was designed in the absence of a known mechanism for induction of senescence by O
3 but could be interpreted mechanistically if accumulated O
3 flux is assumed to be proportional to increased respiratory effort accumulated over the season, which has been proposed as a potential trigger for O
3-induced senescence [
25]. For Skyfall, across the five highest O
3 treatments in 2015, onset occurred across a POD
0SPEC range of 22.0–25.7 mmol m
−2. Given the limitations associated with the method used to identify senescence onset—arbitrarily defined as a 10% reduction in leaf CCI relative to the control—as well as the inherent variation that exists between seedlings, this flux range can be considered relatively narrow. However, when all treatments that exhibited a significant O
3 effect on senescence are considered for Skyfall in 2015, the range of flux at senescence onset is considerably wider (17.8–25.7 mmol m
−2 POD
0SPEC). These results provide an estimate of the degree of error potentially associated with applying this approach in models and suggest that the accumulation of respiratory effort does not fully explain the triggering of O
3-induced senescence.
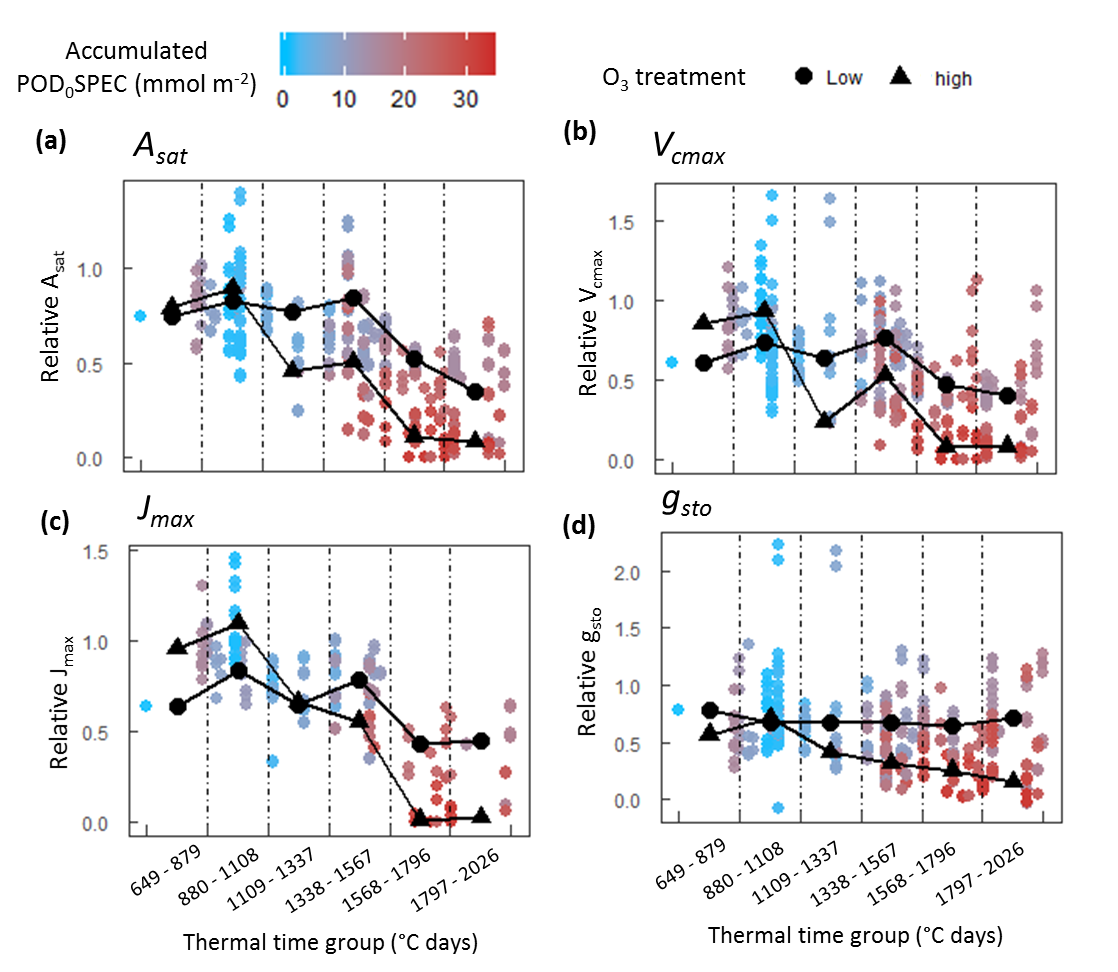
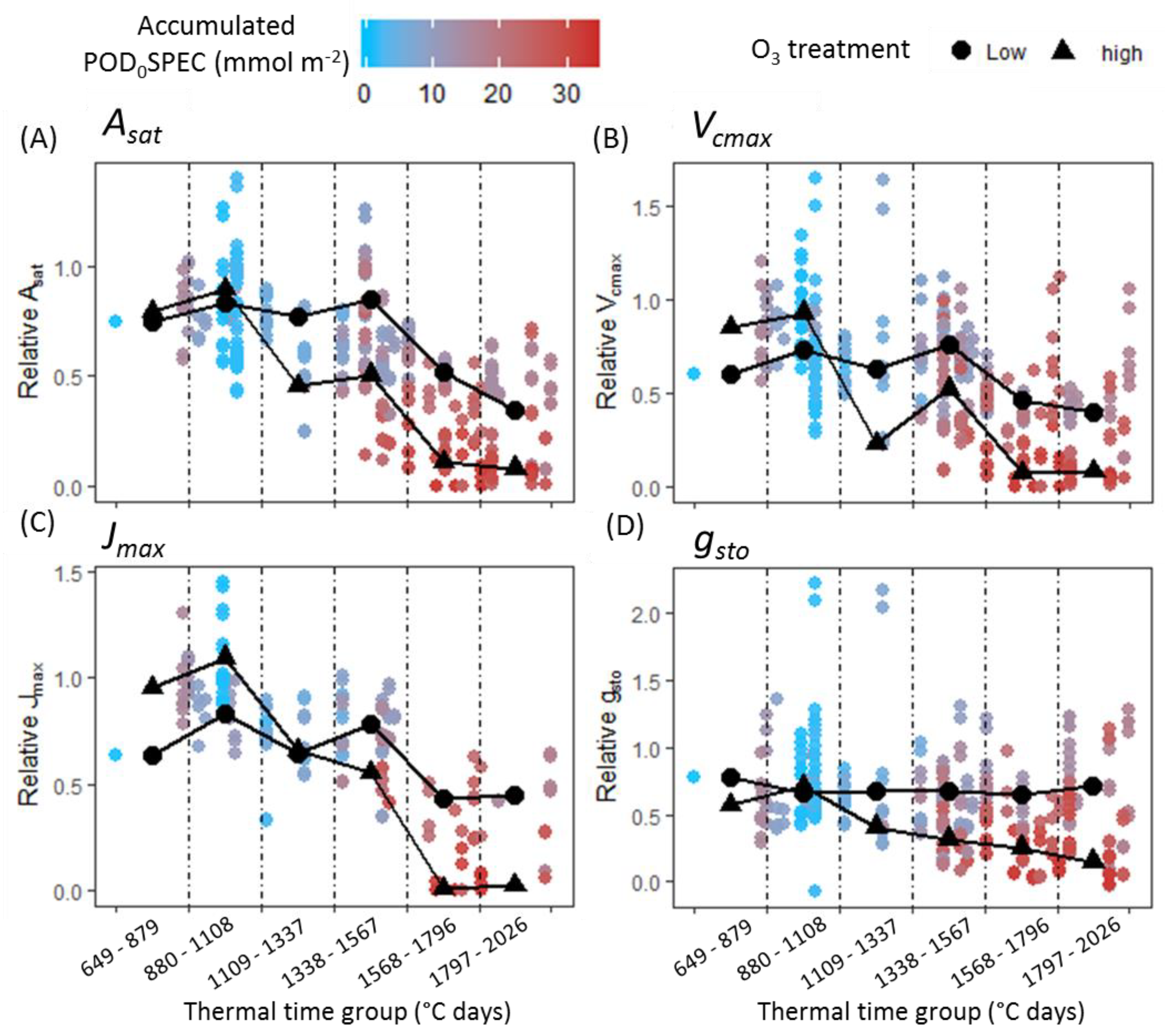
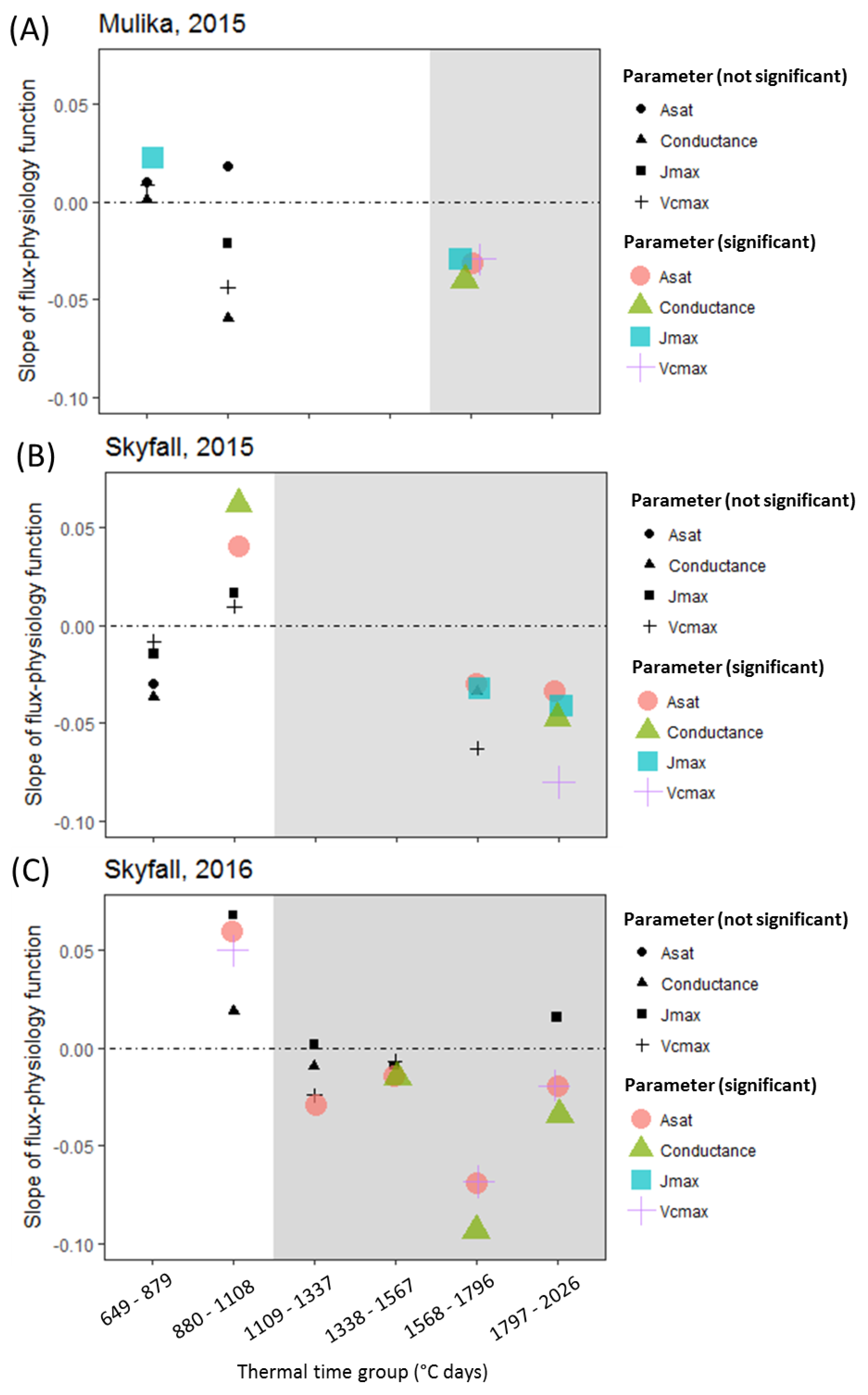
A second objective of this study was to test the hypothesis that O
3 reduces photosynthetic rate in the short term by reducing carboxylation capacity of rubisco (
Vcmax). The assumption that O
3 reduces
Vcmax is central to the ‘instantaneous’ O
3 effect function of Martin et al. [
27]. A version of this function is also applied by Ewert and Porter [
25], where O
3 reduces photosynthesis in a short-term and reversible way, in addition to and independent of the O
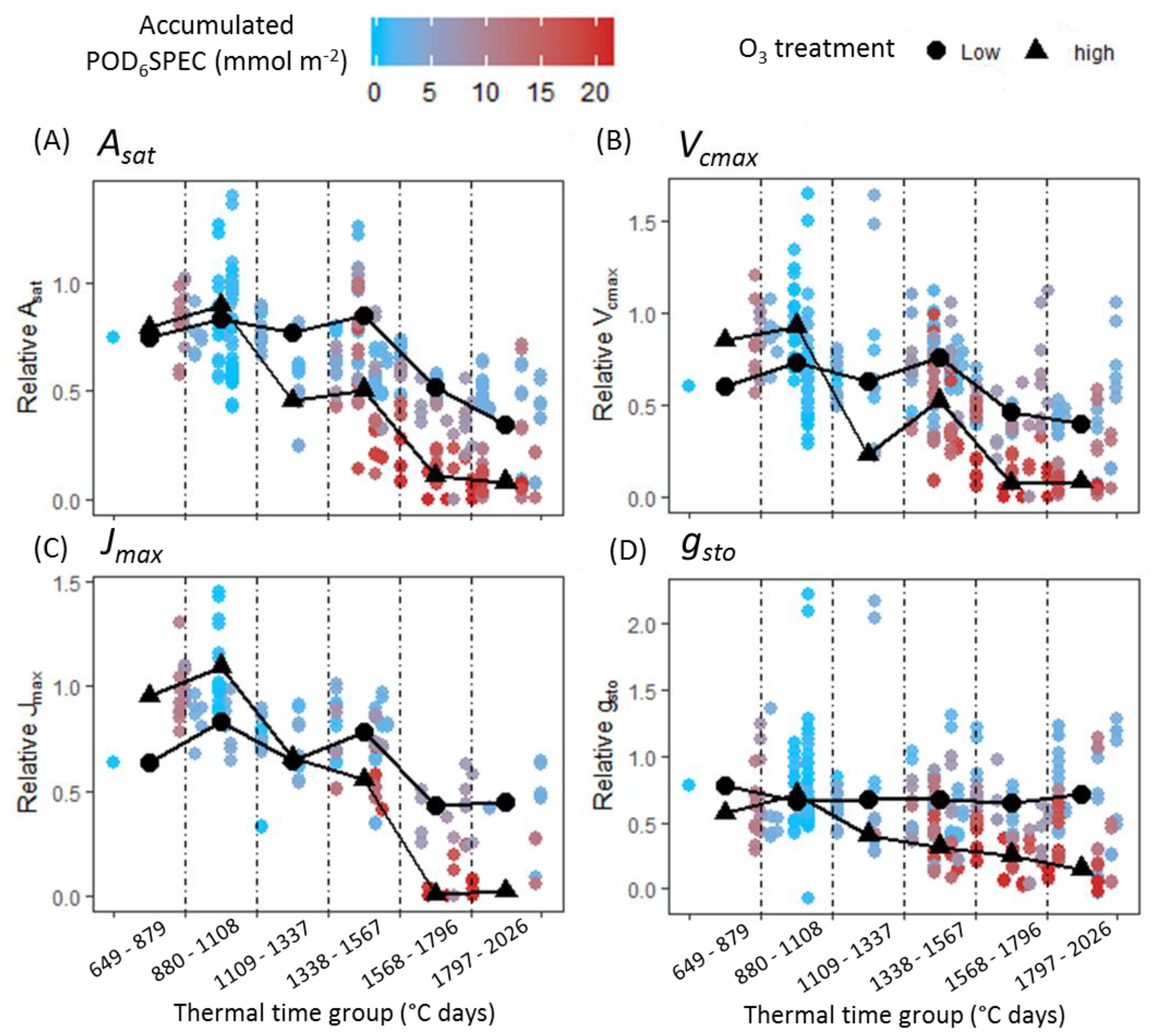
3 senescence effect. The analysis presented here found that a significant negative effect of O
3 on photosynthesis and
gsto was only observed concurrent with O
3-induced leaf senescence (
Figure 4). This result was consistent across all cultivar–year combinations. For Skyfall in 2015 and 2016, significant negative effects of O
3 on
Asat were observed
before a negative association between O
3 and
Vcmax, suggesting that reduced carboxylation capacity is not responsible for the initial reduction in photosynthetic capacity observed in these experiments. There was therefore no evidence of an ‘instantaneous’ effect of O
3 on the photosynthetic mechanism, in the period preceding leaf senescence.
These results contradict several studies which observed short-term reduction in photosynthetic rate in response to O
3 [
39,
40,
41]. One possible explanation for this contradiction is that instantaneous reduction of carboxylation capacity by O
3 is only relevant at acute concentrations. The reduced carboxylation efficiency reported by Farage et al. [
41] was observed following 4–16 h of exposure at unrealistically high O
3 concentrations of 200–400 ppb, considerably higher hourly concentrations compared to those used in the experiments in this study, which more closely mimic ambient conditions (maximum hourly O
3 exposure of 117 ppb). The results presented here therefore indicate that accelerated senescence is likely to be more important than direct effects on photosynthesis for determining crop yield loss in most agricultural landscapes, where O
3 concentrations are typically moderate (ranging from ~20 to ~45 ppb) with occasional peaks in concentration [
5,
52]. Understanding and simulating the early senescence response to O
3 should therefore be the priority for O
3 experimentalists and modellers.
Alternatively, our results could be explained by a differential response to O
3 in younger and older leaves. Bernacchi et al. [
53] and Morgan et al. [
54] observed in field experiments with soybean that O
3 effects on photosynthesis and
gsto were not apparent in new fully expanded leaves and Reichenauer et al. [
55] saw similar results in three wheat cultivars. Younger leaves may have a higher tolerance to O
3, or alternatively the O
3 effect on photosynthesis may be associated with a cumulative build-up of O
3 damage in leaves or leaf age. Either way, the age-dependency of O
3 effects is an important consideration in O
3 effects modelling. The function described by Ewert and Porter [
25] for modelling short-term effects of O
3 on photosynthesis allows for leaf age to influence the rate of overnight recovery from O
3 damage but not the threshold for damage. The role of leaf age in determining O
3 flux thresholds would benefit from further investigation.
A surprising result from the data analysis is that O
3 had a significant positive effect on some physiological parameters early in the season. CCI,
Asat,
Vcmax, and
gsto all exhibited a positive association with O
3 exposure for one or more of the cultivar–year combinations, in either the first or second thermal time group (up to 32 days following beginning of exposure). Stimulation of growth at low doses of a toxin is known as hormesis, and this phenomenon has been observed previously in a number of plants in response to exposure to low concentrations of O
3 [
56]. Stimulation of photosynthesis and
gsto in wheat during the first few weeks of O
3 exposure was also observed by Mulholland et al. [
57] in their open-top chamber experiment, although generally there are few reported cases of this phenomenon in crop species. Ozone-induced physiological stimulation could be an adaptive response associated with plant defence responses, for example, heightened
gsto and photosynthesis may enable the upregulation of antioxidant synthesis. Observations that yield can be stimulated at low O
3 exposure concentrations have also given rise to the theory that free radicals, at low concentrations, can act as growth promotors in plants [
58]. An alternative hypothesis is that the observed early-season physiological boost in this study is related to disruption of stomatal control by O
3, as has been observed in some grassland species [
59,
60], leading to heightened
gsto and an associated boost in other physiological parameters. Our results suggest that a bi-phase hermetic dose-response curve, such as proposed by Agathokleous et al. [
56], could be considered in ozone effect functions. More experimental data is needed in order to establish if the early-season physiological boost induced by O
3 in this study is consistent across other plant species and environments.
The third aim of this study was to test whether O
3 flux would be a better predictor of physiological response than concentration-based exposure (AOT40 and 24-h mean). Flux was superior at predicting the response to O
3 of five physiological variables (CCI,
Asat,
Vcmax,
Jmax,
gsto) in regression models, for both cultivars (
Table 3). Previous studies have reported that flux is better than AOT40 at predicting the spatial distribution of O
3 injury [
61] and is a better predictor of the response of wheat yield, poplar biomass, and assimilation rate in urban trees to O
3 exposure [
62,
63,
64]. However, few have compared the association between leaf-level physiology and different exposure metrics in crop species. Our results align with the general consensus in the O
3 research community that O
3 flux represents a more biologically relevant metric of O
3 exposure than ambient concentration [
61,
65,
66,
67] and indicate that O
3 flux should be the preferred metric of exposure in O
3 effect model functions. More surprising is the fact that the flux metric without an accumulation threshold, POD
0SPEC, was a better or equal predictor of physiological response compared to POD
6SPEC, which employs an accumulation threshold of six. POD
6SPEC produced the closest correlation between flux and yield of wheat in a previous analysis testing different flux accumulation thresholds [
50] and has been applied in several assessments of O
3 impacts in wheat [
22,
68]. More research is therefore needed to establish how much the capacity to detoxify O
3 varies between cultivars and why the threshold flux required to induce leaf-level physiological changes appears to differ from the threshold required to reduce yield. This result also highlights the limitations of an empirical rather than a mechanistic approach to modelling ozone tolerance and detoxification capacity. The development and integration of mechanistic ozone detoxification modules in models, such as that proposed by Plöchl et al., could potentially lead to improved model accuracy [
69].
The view that O
3 flux should be the metric of exposure in O
3 effect modelling is also supported by the fact that O
3 flux accounted for the different levels of exposure in treatments dominated by peaks in concentration, versus those characterised by a consistent background level, for the majority of physiological parameters. Flux is therefore likely to perform well as a predictor of physiological response across different world regions which are currently experiencing divergent trends in the pattern of O
3 exposure [
10,
11,
12].
Limitations of this study need to be considered when interpreting and applying the results. Calculated values of O
3 flux were not verified by leaf-level measurements of gas flux through stomata. However, the decision to apply the multiplicative DO
3SE model in this study was based on the fact that fluxes produced by this model have previously been evaluated in several independent studies which have demonstrated the model’s predictive capability [
19,
21,
22,
70,
71,
72]. A further limitation is that estimates of the onset of leaf senescence were based on leaf chlorophyll content, which would have represented both the chlorophyll loss resulting from leaf injury, as well as chlorophyll loss relating to senescence. In addition, the analysis is based on only one crop species and two cultivars. As the variation in yield response to O
3 exhibited by different crops, and different cultivars within the same crop species, is well established [
2,
73], caution must be used when extrapolating results presented here to other wheat cultivars and crops. It should however be noted that the observation in this study that no O
3 effect on photosynthesis could be observed in young wheat leaves—indicating the senescence response is more important than direct effects on photosynthesis—is supported by previous work in other wheat cultivars [
55] and by other experimental work in soybean [
53,
54]. Considerations when applying the results presented in this study, particularly when attempting to up-scale modelled responses from the leaf to canopy level, include the fact that the response observed in the wheat flag leaf may differ from the responses of lower-canopy leaves, and exposure to O
3 during early seedling and leaf development may also alter the sensitivity to O
3 observed in the flag leaf.
In conclusion, this study has shown that current approaches for modelling O3 effects on leaf longevity and photosynthesis in crops have some limitations and are not fully supported by the experimental data presented here. Model functions representing O3-induced senescence must allow for parameterisation of the timing of senescence onset and rate of senescence, if inter-cultivar variation in response is to be accurately simulated. Further research aimed at understanding the mechanistic ‘trigger’ of O3-induced senescence should be a priority, as this understanding may allow for the development of a more effective mechanism in models for inducing the senescence response. The results also suggest an age-dependency in the response of photosynthesis to O3—which is not currently fully considered in modelling methods—and indicate that acceleration of senescence is more important than direct effects of O3 on photosynthesis in determining final O3-induced yield loss, at O3 concentrations that crops are likely to be exposed to on a day-to-day basis. Building functions that can accurately represent the O3-induced senescence effect in crops should therefore be the priority for O3 effect modellers.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










