Transcriptomic Analysis of Orange Fruit Treated with Pomegranate Peel Extract (PGE)



 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sampling
2.2. Data Analysis
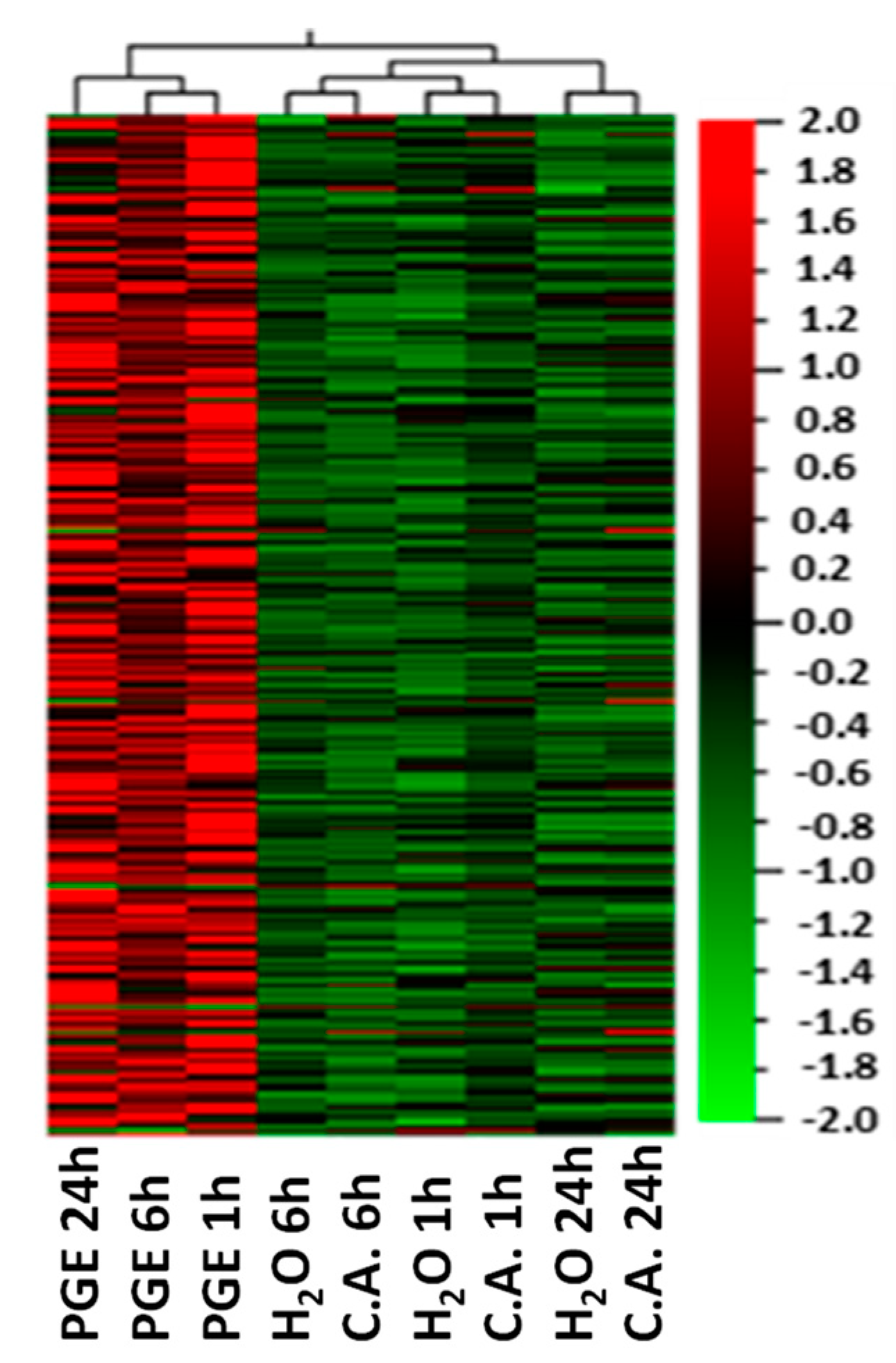
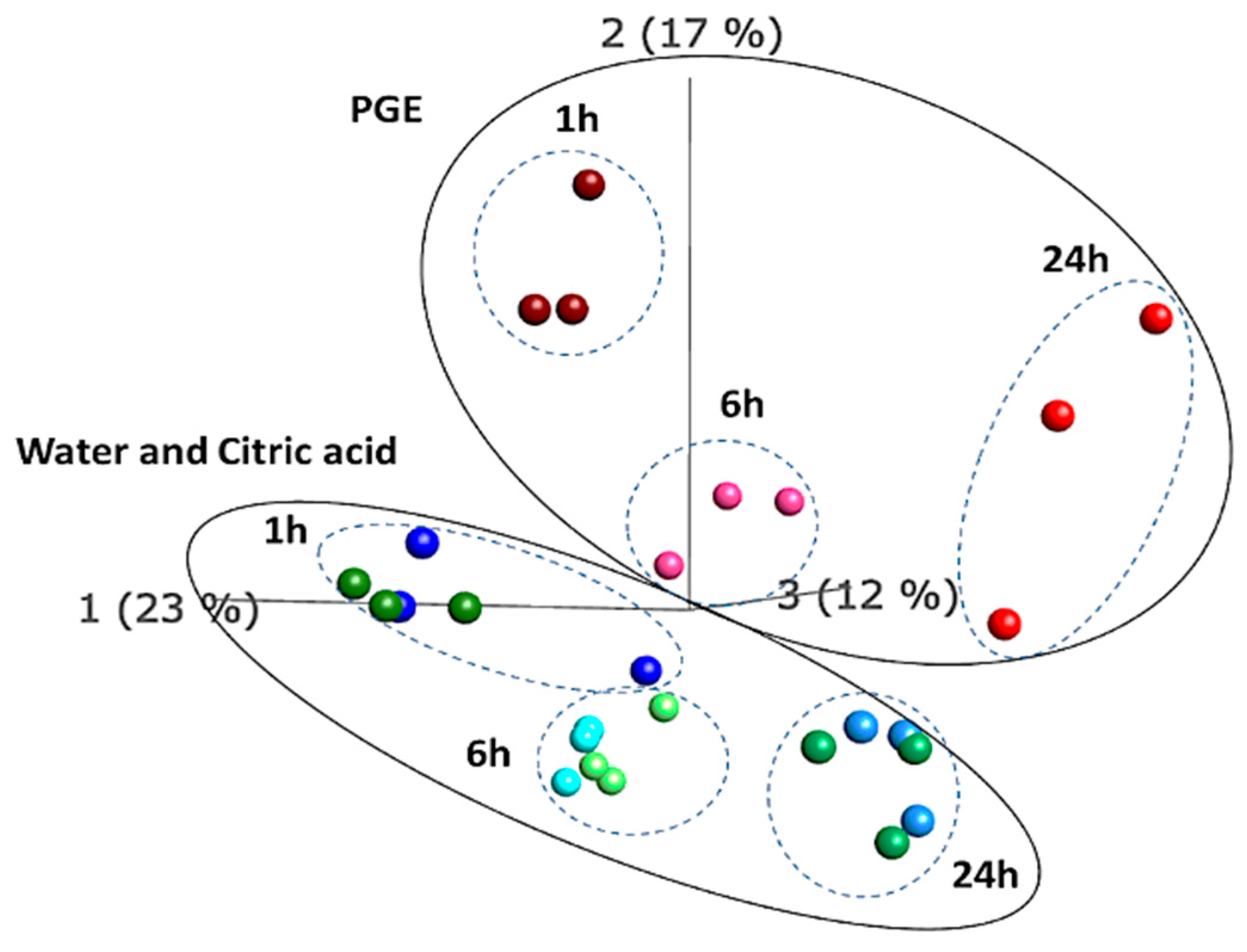
3. Results
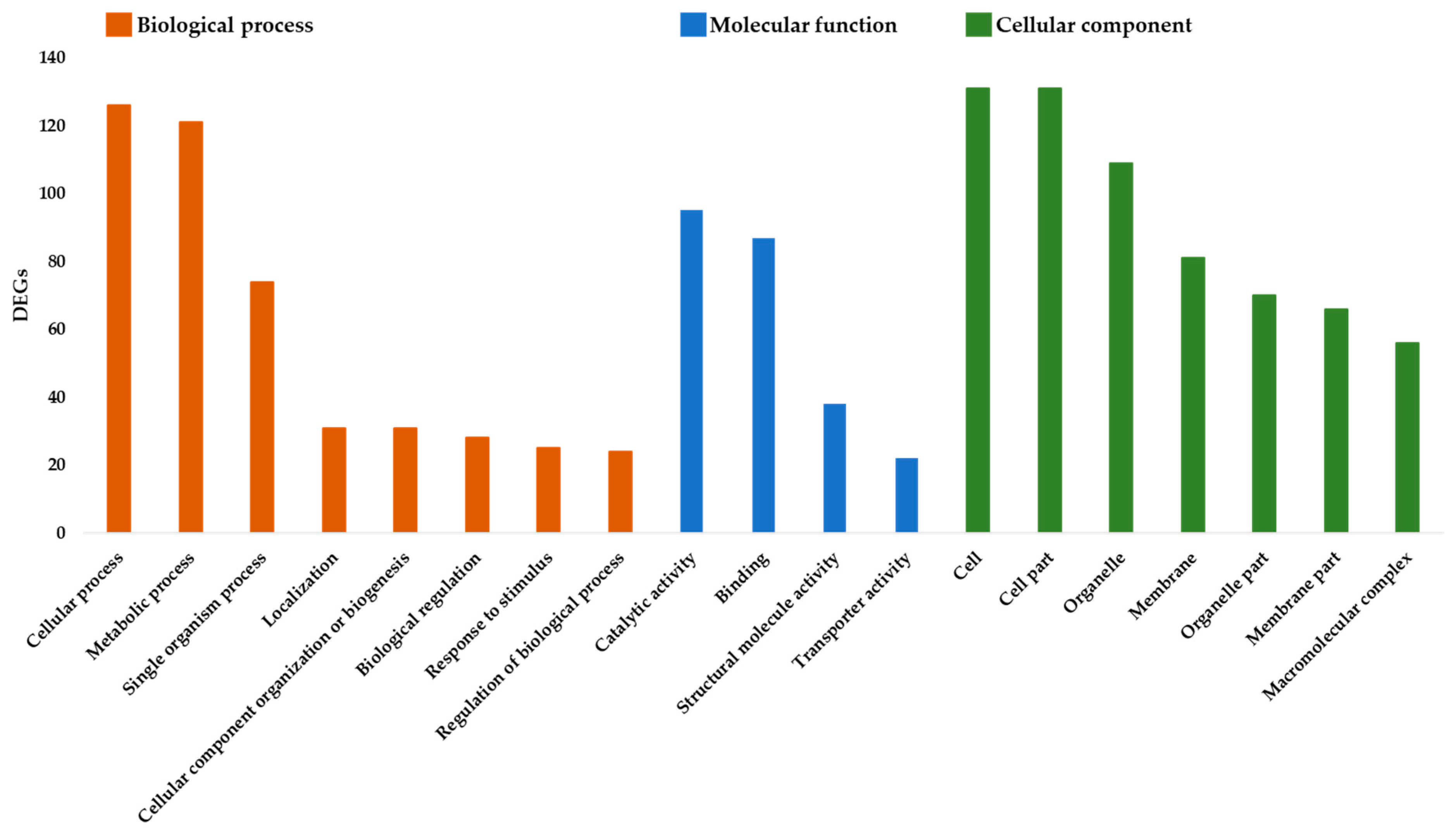
3.1. Gene Ontology Enrichment
3.2. KEGG Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Venkataramanamma, D.; Aruna, P.; Singh, R. Standardization of the conditions for extraction of polyphenols from pomegranate peel. J. Food Sci. Technol. 2016, 53, 2497–2503. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.A.; Carle, R.; Kammerer, D.R. Identification and quantification of phenolic compounds from pomegranate (Punica granatum L.) peel, mesocarp, aril and differently produced juices by HPLC-DAD–ESI/MSn. Food Chem. 2011, 127, 807–821. [Google Scholar] [CrossRef] [PubMed]
- Tehranifar, A.; Selahvarzi, Y.; Kharrazi, M.; Bakhsh, V.J. High potential of agro-industrial by-products of pomegranate (Punica granatum L.) as the powerful antifungal and antioxidant substances. Ind. Crops Prod. 2011, 34, 1523–1527. [Google Scholar] [CrossRef]
- Zahin, M.; Aqil, F.; Ahmad, I. Broad spectrum antimutagenic activity of antioxidant active fraction of Punica granatum L. peel extracts. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2010, 703, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Al-Zoreky, N. Antimicrobial activity of pomegranate (Punica granatum L.) fruit peels. Int. J. Food Microbiol. 2009, 134, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Gullon, B.; Pintado, M.E.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Assessment of polyphenolic profile and antibacterial activity of pomegranate peel (Punica granatum) flour obtained from co-product of juice extraction. Food Control 2016, 59, 94–98. [Google Scholar] [CrossRef]
- Romeo, F.V.; Ballistreri, G.; Fabroni, S.; Pangallo, S.; Li Destri Nicosia, M.G.; Schena, L.; Rapisarda, P. Chemical characterization of different sumac and pomegranate extracts effective against Botrytis cinerea rots. Molecules 2015, 20, 11941–11958. [Google Scholar] [CrossRef] [PubMed]
- Li Destri Nicosia, M.G.; Pangallo, S.; Raphael, G.; Romeo, F.V.; Strano, M.C.; Rapisarda, P.; Droby, S.; Schena, L. Control of postharvest fungal rots on citrus fruit and sweet cherries using a pomegranate peel extract. Postharvest Biol. Technol. 2016, 114, 54–61. [Google Scholar] [CrossRef]
- Pangallo, S.; Li Destri Nicosia, M.G.; Raphael, G.; Levin, E.; Ballistreri, G.; Cacciola, S.; Rapisarda, P.; Droby, S.; Schena, L. Elicitation of resistance responses in grapefruit and lemon fruits treated with a pomegranate peel extract. Plant Pathol. 2017, 66, 633–640. [Google Scholar] [CrossRef]
- Pangallo, S.; Li Destri Nicosia, M.G.; Agosteo, G.E.; Abdelfattah, A.; Romeo, F.V.; Cacciola, S.O.; Rapisarda, P.; Schena, L. Evaluation of a pomegranate peel extract as an alternative means to control olive anthracnose. Phytopathology 2017, 107, 1462–1467. [Google Scholar] [CrossRef]
- Alsaggaf, M.S.; Moussa, S.H.; Tayel, A.A. Application of fungal chitosan incorporated with pomegranate peel extract as edible coating for microbiological, chemical and sensorial quality enhancement of Nile tilapia fillets. Int. J. Biol. Macromol. 2017, 99, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Sanzani, S.M.; Schena, L.; De Girolamo, A.; Ippolito, A.; González-Candelas, L. Characterization of genes associated with induced resistance against Penicillium expansum in apple fruit treated with quercetin. Postharvest Biol. Technol. 2010, 56, 1–11. [Google Scholar] [CrossRef]
- Jia, Z.; Zou, B.; Wang, X.; Qiu, J.; Ma, H.; Gou, Z.; Song, S.; Dong, H. Quercetin-induced H 2 O 2 mediates the pathogen resistance against Pseudomonas Syringae pv. Tomato DC3000 in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2010, 396, 522–527. [Google Scholar] [CrossRef]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: the importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Chen, L.-L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.-B.; Hao, B.-H.; Lyon, M.P. The draft genome of sweet orange (Citrus Sinensis). Nat. Genet. 2013, 45, 59. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Rojas, C.M.; Senthil-Kumar, M.; Tzin, V.; Mysore, K. Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front. Plant Sci. 2014, 5, 17. [Google Scholar] [CrossRef]
- Bin, P.; Huang, R.; Zhou, X. Oxidation resistance of the sulfur amino acids: Methionine and cysteine. BioMed Res. Int. 2017, 2017. [Google Scholar] [CrossRef]
- Bolton, M.D. Primary metabolism and plant defense—Fuel for the fire. Mol. Plant Microbe Interact. 2009, 22, 487–497. [Google Scholar] [CrossRef]
- Dean, J.; Goodwin, P.; Hsiang, T. Induction of glutathione S-transferase genes of Nicotiana benthamiana following infection by Colletotrichum destructivum and C. orbiculare and involvement of one in resistance. J. Exp. Bot. 2005, 56, 1525–1533. [Google Scholar] [CrossRef]
- Ghanta, S.; Bhattacharyya, D.; Chattopadhyay, S. Glutathione signaling acts through NPR1-dependent SA-mediated pathway to mitigate biotic stress. Plant Signal. Behav. 2011, 6, 607–609. [Google Scholar] [CrossRef]
- Guerriero, G.; Berni, R.; Muñoz-Sanchez, J.; Apone, F.; Abdel-Salam, E.; Qahtan, A.; Alatar, A.; Cantini, C.; Cai, G.; Hausman, J.-F. Production of plant secondary metabolites: Examples, tips and suggestions for biotechnologists. Genes 2018, 9, 309. [Google Scholar] [CrossRef]
- Couée, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef]
- Qi, H.; Jiang, Z.; Zhang, K.; Yang, S.; He, F.; Zhang, Z. PlaD: A transcriptomics database for plant defense responses to pathogens, providing new insights into plant immune system. Genomics Proteomics Bioinform. 2018, 16, 283–293. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.; Wang, L. The phenylpropanoid pathway and plant defence—A genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Wink, M.; Schimmer, O. Molecular modes of action of defensive secondary metabolites. Funct. Biotechnol. Plant Second. Metab. 2010, 39, 21–161. [Google Scholar]
- Dugo, G.; Di Giacomo, A. Citrus: The Genus Citrus, Medicinal and Aromatic Plants—Industrial Profiles; Taylor & Francis: New York, NY, USA, 2002; Volume 26, pp. 195–201. [Google Scholar]
- Rodríguez, A.; Shimada, T.; Cervera, M.; Alquézar, B.; Gadea, J.; Gómez-Cadenas, A.; De Ollas, C.J.; Rodrigo, M.J.; Zacarías, L.; Peña, L. Terpene down-regulation triggers defense responses in transgenic orange leading to resistance against fungal pathogens. Plant Physiol. 2014, 164, 321–339. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.; Lattanzio, V.M.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. Phytochem. Adv. Res. 2006, 661, 23–67. [Google Scholar]
- De Oliveira, M.R.; Nabavi, S.M.; Braidy, N.; Setzer, W.N.; Ahmed, T.; Nabavi, S.F. Quercetin and the mitochondria: A mechanistic view. Biotechnol. Adv. 2016, 34, 532–549. [Google Scholar] [CrossRef] [PubMed]
- Sandermann, H., Jr. Plant metabolism of xenobiotics. Trends Biochem. Sci. 1992, 17, 82–84. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Replicates | Sampling Time (hpt) | Read 1 | Read 2 |
|---|---|---|---|---|
| Citric acid | R1 | 1 | 16509545 | 16509545 |
| R2 | 1 | 10819465 | 10819465 | |
| R3 | 1 | 12792832 | 12792832 | |
| R1 | 6 | 11027909 | 11027909 | |
| R2 | 6 | 14228323 | 14228323 | |
| R3 | 6 | 8719836 | 8719836 | |
| R1 | 24 | 15550388 | 15550388 | |
| R2 | 24 | 24863143 | 24863143 | |
| R3 | 24 | 11287417 | 11287417 | |
| H2O | R1 | 1 | 13733858 | 13733858 |
| R2 | 1 | 16898374 | 16898374 | |
| R3 | 1 | 9761332 | 9761332 | |
| R1 | 6 | 22501980 | 22501980 | |
| R2 | 6 | 20726432 | 20726432 | |
| R3 | 6 | 20954767 | 20954767 | |
| R1 | 24 | 15517302 | 15517302 | |
| R2 | 24 | 18455542 | 18455542 | |
| R3 | 24 | 20239137 | 20239137 | |
| PGE | R1 | 1 | 9514582 | 9514582 |
| R2 | 1 | 8451256 | 8451256 | |
| R3 | 1 | 12624217 | 12624217 | |
| R1 | 6 | 11596104 | 11596104 | |
| R2 | 6 | 14399695 | 14399695 | |
| R3 | 6 | 10650424 | 10650424 | |
| R1 | 24 | 13176031 | 13176031 | |
| R2 | 24 | 9556111 | 9556111 | |
| R3 | 24 | 9187532 | 9187532 |
| Category | Pathway | Number of Genes | Enzymes in Pathway |
|---|---|---|---|
| Carbohydrate and Energy metabolism | Glycolysis/Gluconeogenesis | 3 | ec:5.3.1.1, ec:4.1.2.13, ec:2.7.2.3 |
| Pyruvate metabolism | 3 | ec:1.1.1.37, ec:4.4.1.5, ec:3.1.2.6 | |
| Pentose phosphate pathway | 3 | ec:2.7.1.15, ec:2.2.1.2, ec:4.1.2.13 | |
| Glyoxylate and dicarboxylate metabolism | 2 | ec:1.1.1.37, ec:1.1.3.15 | |
| Fructose and mannose metabolism | 2 | ec:5.3.1.1, ec:4.1.2.13 | |
| Pentose and glucuronate interconversions | 2 | ec:1.1.1.22, ec:4.2.2.2 | |
| Amino sugar and nucleotide sugar metabolism | 1 | ec:1.1.1.22 | |
| Inositol phosphate metabolism | 1 | ec:5.3.1.1 | |
| Ascorbate and aldarate metabolism | 1 | ec:1.1.1.22 | |
| Citrate cycle (TCA cycle) | 1 | ec:1.1.1.37 | |
| Oxidative phosphorylation | 6 | ec:1.10.2.2, ec:1.9.3.1, ec:1.6.5.3 | |
| Carbon fixation pathways in prokaryotes | 1 | ec:1.1.1.37 | |
| Carbon fixation in photosynthetic organisms | 4 | ec:1.1.1.37, ec:5.3.1.1, ec:4.1.2.13, ec:2.7.2.3 | |
| Methane metabolism | 2 | ec:1.1.1.37, ec:4.1.2.13 | |
| Nitrogen metabolism | 1 | ec:1.7.1.1 | |
| Sulfur metabolism | 5 | ec:3.6.2.1, ec:2.5.1.48, ec:2.7.7.4, ec:2.7.1.25 | |
| Lipid metabolism | Glycerolipid metabolism | 1 | ec:3.1.1.3 |
| Nucleotide metabolism | Purine metabolism | 5 | ec:3.6.1.3, ec:2.7.7.4, ec:2.7.4.6, ec:2.4.2.7, ec:2.7.1.25 |
| Pyrimidine metabolism | 1 | ec:2.7.4.6 | |
| Amino acid metabolism | Cysteine and methionine metabolism | 6 | ec:1.1.1.37, ec:2.5.1.6, ec:2.1.1.14, ec:1.13.11.54, ec:3.3.1.1, ec:2.5.1.48 |
| Phenylalanine metabolism | 1 | ec:2.1.1.104 | |
| Selenocompound metabolism | 4 | ec:2.1.1.14, ec:2.5.1.48, ec:2.7.7.4 | |
| Glutathione metabolism | 4 | ec:2.5.1.18, ec:1.11.1.15 | |
| Biosynthesis of secondary metabolites | Antibiotic biosynthesis | 10 | ec:1.1.1.37, ec:1.1.3.15 ec:2.5.1.48, ec:2.2.1.2, ec:5.3.1.1, ec:2.7.7.4, ec:4.1.2.13, ec:2.7.4.6, ec:2.7.2.3 |
| Monoterpenoid biosynthesis | 2 | ec:4.2.3.20 | |
| Phenylpropanoid biosynthesis | 4 | ec:1.11.1.7, ec:2.1.1.104 | |
| Flavonoid biosynthesis | 1 | ec:2.1.1.104 | |
| Monobactam biosynthesis | 2 | ec:2.7.7.4 | |
| Stilbenoid, diarylheptanoid and gingerol biosynthesis | 1 | ec:2.1.1.104 | |
| Xenobiotics biodegradation and metabolism | Fluorobenzoate degradation | 1 | ec:3.1.1.45 |
| Toluene degradation | 1 | ec:3.1.1.45 | |
| Metabolism of xenobiotics by cytochrome P450 | 3 | ec:2.5.1.18 | |
| Drug metabolism - cytochrome P450 | 3 | ec:2.5.1.18 | |
| Drug metabolism - other enzymes | 2 | ec:3.1.1.1 | |
| Chlorocyclohexane and chlorobenzene degradation | 1 | ec:3.1.1.45 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belgacem, I.; Pangallo, S.; Abdelfattah, A.; Romeo, F.V.; Cacciola, S.O.; Li Destri Nicosia, M.G.; Ballistreri, G.; Schena, L. Transcriptomic Analysis of Orange Fruit Treated with Pomegranate Peel Extract (PGE). Plants 2019, 8, 101. https://doi.org/10.3390/plants8040101
Belgacem I, Pangallo S, Abdelfattah A, Romeo FV, Cacciola SO, Li Destri Nicosia MG, Ballistreri G, Schena L. Transcriptomic Analysis of Orange Fruit Treated with Pomegranate Peel Extract (PGE). Plants. 2019; 8(4):101. https://doi.org/10.3390/plants8040101
Chicago/Turabian StyleBelgacem, Imen, Sonia Pangallo, Ahmed Abdelfattah, Flora V. Romeo, Santa O. Cacciola, Maria G. Li Destri Nicosia, Gabriele Ballistreri, and Leonardo Schena. 2019. "Transcriptomic Analysis of Orange Fruit Treated with Pomegranate Peel Extract (PGE)" Plants 8, no. 4: 101. https://doi.org/10.3390/plants8040101
APA StyleBelgacem, I., Pangallo, S., Abdelfattah, A., Romeo, F. V., Cacciola, S. O., Li Destri Nicosia, M. G., Ballistreri, G., & Schena, L. (2019). Transcriptomic Analysis of Orange Fruit Treated with Pomegranate Peel Extract (PGE). Plants, 8(4), 101. https://doi.org/10.3390/plants8040101