Abstract
Efficient transformation of numerous important crops remains a challenge, due predominantly to our inability to stimulate growth of transgenic cells capable of producing plants. For years, this difficulty has been partially addressed by tissue culture strategies that improve regeneration either through somatic embryogenesis or meristem formation. Identification of genes involved in these developmental processes, designated here as morphogenic genes, provides useful tools in transformation research. In species from eudicots and cereals to gymnosperms, ectopic overexpression of genes involved in either embryo or meristem development has been used to stimulate growth of transgenic plants. However, many of these genes produce pleiotropic deleterious phenotypes. To mitigate this, research has been focusing on ways to take advantage of growth-stimulating morphogenic genes while later restricting or eliminating their expression in the plant. Methods of controlling ectopic overexpression include the use of transient expression, inducible promoters, tissue-specific promoters, and excision of the morphogenic genes. These methods of controlling morphogenic gene expression have been demonstrated in a variety of important crops. Here, we provide a review that highlights how ectopic overexpression of genes involved in morphogenesis has been used to improve transformation efficiencies, which is facilitating transformation of numerous recalcitrant crops. The use of morphogenic genes may help to alleviate one of the bottlenecks currently slowing progress in plant genome modification.
1. Introduction
Despite progress in crop transformation over the past several decades, efficient production of transgenic plants remains one of the major barriers to crop improvement [1]. There are two components to producing transgenic plants. The first is the ability to introduce and express transgenes (transformation), and the second is the ability to form tissue (typically de novo embryos or shoots) capable of regenerating into a fertile plant. Many plant species (or genotypes) remain difficult to transform and regenerate. Such varieties are referred to as being recalcitrant to transformation and plant regeneration. One of the promising tools helping to reduce this recalcitrance (and thus alleviate the bottleneck) is the use of genes involved in controlling plant growth and development.
Morphogenesis, or the organized spatial development of embryos, tissues, and organs, is a tightly controlled process involving networks of genes acting sequentially or in concert. Within this broad context, the concept of trying to use genes involved in either embryogenesis or meristem maintenance has attracted the attention of plant transformation researchers for many years. Basic research within these two well-defined areas has contributed an ever-expanding number of genes and gene networks involved in embryo development [2] and meristem development [3,4,5,6] that we will not attempt to cover here. De novo regeneration of plants typically occurs through either somatic embryogenesis or organogenesis (de novo formation of new meristems or through rearrangement of pre-existing meristems), traditionally manipulated by adjusting auxin/cytokinin ratios in the medium [6,7,8]. In addition to exogenous hormone manipulation, ectopic overexpression of plant genes that control growth and development has also proven to be useful.
Of course, there are numerous reports where non-plant genes have been used to improve transformation frequencies and/or plant regeneration. Examples include tumor-inducing genes from Agrobacterium, such as the isopentyl transferase or ipt gene [9,10], rolC [11,12], rolB, 6B [13], and tzs [14,15], and viral genes that stimulate the plant cell cycle [16]. Further details for non-plant genes are beyond the scope of this review. Instead, we will focus on research that has demonstrated a potentially useful morphogenic growth response due to ectopic overexpression of a plant embryo or meristem gene (morphogenic genes), or has demonstrated a practical benefit for plant transformation or regeneration. Plant transformation and regeneration improvements can be further distinguished by their impact on either improving transformation efficiencies, or improving the regeneration process to recover transgenic plants. Finally, we are making a distinction between reports focused solely on observations of morphogenic responses, and those studies that describe practical methods to improve transformation and/or regeneration. Both types of results are important in terms of their contribution to the field of transformation research, and are described below.
Characterizing morphogenic genes involves phenotypic analysis of knockout mutants and/or transgenic experiments typically involving either ectopic overexpression or downregulation in plants. Often, the two strategies are combined, where introduction of an expression cassette is used to complement a mutant phenotype. Morphogenic responses have been observed using transient expression, constitutive expression, or induction of gene expression by exposure of a stable transgenic event to a chemical ligand. Using a plant morphogenic gene to improve transformation, on the other hand, almost invariably involves limiting the length of time that expression occurs to avoid later pleiotropic effects in regenerated T0 plants and subsequent progeny generations (Figure 1).
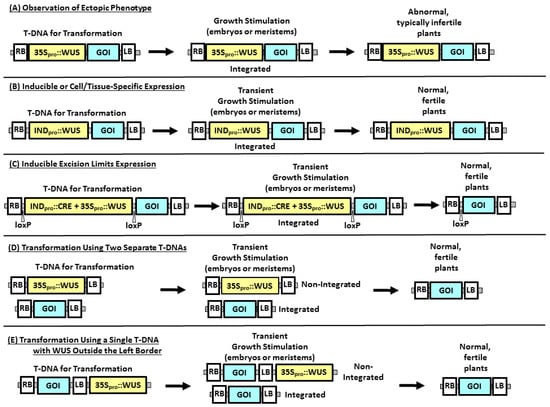
Figure 1.
Methods for expression of morphogenic genes in plant transformation. WUS is used to exemplify the morphogenic gene expression cassettes, which could be designed for overexpression of the gene, downregulation of a gene, or combinations of genes, while the box labeled “GOI” (Genes Of Interest) represents trait gene expression cassettes. (A) Using a constitutive promoter, such as CaMV35S, to drive expression of a WUS gene results in growth stimulation of cells transformed with the T-DNA, either through somatic embryogenesis or through meristem proliferation. (B) Using an inducible promoter to drive expression of WUS will result in growth stimulation only when the plant tissue is exposed to the inducing stimulus (typically a chemical ligand). (C) Growth stimulation can also be effectively controlled by using a combination of constitutive expression of WUS and inducible expression of CRE recombinase to remove the WUS expression cassette. (D) Transforming the same plant cell with a T-DNA containing the WUS expression cassette and a second T-DNA containing the trait expression cassette will also provide transient growth stimulation sufficient to recover regenerable tissues, such as somatic embryos, without the integration of the morphogenic gene. (E) Using a single T-DNA containing the trait, with the WUS expression cassette outside the T-DNA Left Border sequence, higher numbers of the trait-containing T-DNA are introduced relative to the low numbers of “read-through” sequences containing the WUS gene, providing transient growth stimulation without WUS integration. INDpro is used to represent chemically inducible promoters, such as the estradiol-, glucocorticoid-, or tetracycline-responsive promoters in 1-B. Promoters that are induced by physical conditions, such as desiccation (e.g., the RAB17 promoter in [17]), are used to control recombinase-mediated excision (as in 1-C). CRE represents the CRE recombinase expression cassette and loxP are the CRE-recombinase target sites. RB and LB represent the right and left T-DNA border sequences, respectively.
2. Phenotypic Responses from Ectopic Overexpression of Morphogenic Genes
The genes involved in embryogenesis, meristem maintenance, and hormone metabolism are numerous and have been studied for many years [2,3,4,5,6]. Within this large and ever-expanding body of literature, there has been a steady stream of reports demonstrating morphogenesis in response to altered expression of these genes. These observations provide the groundwork that inspires new strategies for transformation research, and are discussed below.
Numerous genes mentioned in this review are typically categorized as genes involved in embryogenesis, meristem function, or hormone pathways. However, we have chosen to group these genes based on their practical benefit (or potential) when used for transformation. Therefore, morphogenic genes that have stimulated an embryogenic or meristematic response when overexpressed were grouped into two categories based on the observed growth response: A) those that enhance a pre-existing embryogenic response under conditions (media composition, exogenous hormones, or even the tissue type) that already elicit the growth response, and B) those that produce ectopic somatic embryos or meristems under conditions where such a response is typically not observed (see Table 1 for a list of genes).

Table 1.
Strategies to improve transformation using morphogenic genes.
2.1. Enhancing the Somatic Embryogenic Response
In this category, overexpression of the plant gene results in enhanced formation of somatic embryos under in vitro culture conditions in which somatic embryogenesis already occurs (see Table 1, Strategy A). This includes the observation that when SOMATIC EMBRYOGENESIS RECEPTOR KINASE1 (SERK1), a gene normally associated with anther and pollen development, was overexpressed using the Cauliflower Mosaic Virus 35S promoter (CaMV 35S) in Arabidopsis thaliana, no changes in plant phenotype were observed, but the embryogenic callus response was improved 3-4-fold over wild-type [18]. This demonstrated that SERK1 stimulated an enhanced somatic embryo response from germinating seedlings placed on media capable of eliciting this response already. Similarly, overexpression of the Coffea canephora SERK1 gene during the in vitro somatic embryogenesis process enhanced the production of somatic embryos by 2-fold, while silencing the gene dramatically reduced the somatic embryogenesis response [19]. Similar conclusions have been reached in reports in which the AGAMOUS-LIKE15 (AGL15) gene was overexpressed, enhancing the formation of secondary somatic embryos from cultured zygotic embryos in Arabidopsis [20], increasing the number of somatic embryos in soybean cultures [21], and again enhancing production of embryogenic callus in cotton [22]. The similarities between SERK1 and AGL15 overexpression are not surprising, since AGL15 is part of the SERK1 protein complex [58]. Interestingly, SERK1 and SERK3 have been shown to be co-receptors, along with BRASSINOSTEROID INSENSITIVE 1 protein (BRI1), of the brassinosteroid class of plant growth regulators [59,60], and the SERK proteins and BRI1 phosphorylate one another upon brassinosteroid sensing. Based on protein structure, the SERK proteins appear to mediate brassinosteroid signaling across the plasma membrane [61]. This observation makes an intriguing connection between brassinosteroid response and embryogenesis.
Increased embryogenic responses have also been reported using genes more typically associated with meristem formation, as with A. thaliana WUSCHEL (AtWUS), a key regulator of meristem cell fate [62], or SHOOT MERISTEMLESS (STM), which is required for proper meristem formation [63]. In transgenic Coffea canephora containing an estradiol-inducible AtWUS construct, leaf discs placed on estradiol increased somatic embryo formation from a control level of one somatic embryo per leaf segment (non-treated), up to a level of 3–5 somatic embryos per transgenic leaf segment after estradiol exposure [23]. Shortly after, it was reported that constitutive overexpression (using the CaMV 35S promoter) of the Brassica napus, Brassica oleracea, or Brassica rapa homologs of STM in Arabidopsis thaliana cotyledons placed on auxin-containing medium resulted in an approximately two-fold increase of somatic embryo formation relative to the wild-type control. In transgenic B. napus containing the 35S::BnSTM construct, a similar two-fold increase was observed in microspore-derived embryogenesis [24]. Similarly, in experiments with the objective of improving transformation methods in cotton (Gossypium hirsutum L.), the 35S::AtWUS cassette was introduced into hypocotyl segments and a three-fold increase in the formation of somatic embryos was observed [25]. Further, the somatic embryos derived from the WUS treatment (when the WUS gene is being overexpressed) produced leaf-like structures but failed to regenerate into plants, likely due to the deleterious effect of WUS ectopic overexpression on subsequent regeneration.
2.2. Ectopic Formation of Somatic Embryos or Meristems
In the second category, overexpression of the plant gene results in direct ectopic formation or spontaneous formation/acquisition of structures resembling embryos (often with embryo characteristics, such as increased oil levels) or meristems in the absence of inductive conditions (see Table 1, Strategy B). In 2002, two important milestone research articles were published that characterized embryonic morphogenesis as a result of ectopic overexpression of either the Brassica napus BABY BOOM (BnBBM) gene [26] or the AtWUS gene [38]. We will first review BBM and other genes involved in embryogenesis, and then later turn our attention to genes involved in meristem function. The BBM gene, a member of the AP2/ERF superfamily of transcription factors [64], has generated great interest among transformation researchers from the first publication [26]. In these experiments, it was observed that constitutive expression of the Brassica BBM gene in Arabidopsis resulted in ectopic somatic embryo formation in vegetative portions of progeny plants, for example in the shoot apex and leaves, and these ectopic somatic embryos could in turn produce plants in the absence of hormones. These results stimulated further research aimed at harnessing BBM-induced somatic embryogenesis to aid in the recovery of transgenic T0 plants.
This first publication was followed by reports in other plant species or using orthologs of the BnBBM gene, providing additional insights into how BBM worked. Srinivasan et al. [27] investigated the ability of various BBM orthologs to induce embryogenic responses in a less-related species. Spontaneous somatic embryogenesis was not observed using 35S::AtBBM in Nicotiana tabacum [27]. However, when these authors used a steroid-inducible, post-translationally controlled AtBBM fusion protein (AtBBM~GR) regulated by the 35S promoter to create stable lines and evaluated progeny, spontaneous ectopic shoot and root formation was observed upon addition of the inducing ligand dexamethasone (DEX). Further, when hypocotyls were exposed to DEX, somatic embryos could be induced when the growth medium contained either zeatin or benzylaminopurine. While the authors attributed the difference observed between Arabidopsis and tobacco to varying competence in response to the BBM signal, they also pointed out that expression of BBM from either Arabidopsis or Brassica in tobacco could produce developmental responses that differ from those observed using the endogenous tobacco BBM gene.
In another example of expressing an orthologous BBM gene, a constitutively expressed soybean gene (35S::GmBBM) was transformed into Arabidopsis [28], and ectopic somatic embryos were observed growing from the cotyledons, the shoot apical meristem, and the hypocotyls of stably transformed plants. Again, differences were observed in the pattern of somatic embryo formation, but, in general, the three studies provide strong evidence that constitutive expression of BBM can result in ectopic somatic embryo formation. Using the genomic clone of Theobroma cacao BBM (TcBBM) under the control of the 35S promoter, Florez et al. [29] demonstrated that it phenocopied the effects of AtBBM in Arabidopsis and stimulated the formation of somatic embryos from Theobroma cotyledons cultured on hormone-free media. Although somatic embryos were formed in cacao using TcBBM, constitutive expression prevented normal plant regeneration.
For many species, recovery of transgenic events is not the bottleneck, but instead regeneration of viable T0 plants is inefficient and rate-limiting. For example, when a BBM ortholog from oil palm (Elaeis guineensis) was cloned into an expression cassette behind the CaMV 35S promoter and then transformed into Arabidopsis, it was observed that cotyledon, leaf, or root segments from stable transgenic events exhibited enhanced rates of shoot formation relative to the wild-type controls [30]. These results are consistent with earlier observations where regeneration was improved through ectopic overexpression of BBM [26]. However, in all these reports, the 35S promoter was used to drive constitutive expression of the transgene and, as a result, no data on recovery of mature fertile plants were presented.
Tsuwamoto et al. [31] using Arabidopsis EMBRYOMAKER (AtEMK), a gene related to BBM (both within the AP2/ERF superfamily) driven by the CaMV 35S promoter in Arabidopsis, produced transgenic progeny that could be phenotypically evaluated. In these experiments, ectopic overexpression of AtEMK produced light-green embryo-like structures (possessing morphological and/or biochemical characteristics normally observed in zygotic embryos but lacking the full functionality of being able to develop into a plant) at the tip of cotyledons in 23% of the seedlings. While several of the embryo-like structures developed small features resembling roots and leaves, these outgrowths did not continue to develop. As ectopic overexpression of AtEMK resulted in pleiotropic effects, it may be necessary to express AtEMK under a regulated system for obtaining normal plantlets from somatic embryos and the embryo-like structures.
Another gene observed to function during early embryo development in Arabidopsis is RKD4, a member of the RWP-RK transcription factor family essential for the first asymmetrical division of the zygote to form the two cells that will give rise to the embryo and suspensor [65,66]. Following up on these characterizations of RKD4 function, Mursyanti et al. [32] demonstrated in orchid (the hybrid Phalaenopsis “Solo Vivien”) that chemical induction of transgenic RKD4 in leaf tissue resulted in ectopic somatic embryogenesis, a very exciting observation in a species normally reluctant to produce direct somatic embryos (de novo embryos that arise from somatic cells, having the capacity to develop into plants).
Genes normally involved in embryo maturation, such as LEAFY COTYLEDON1 (LEC1), LEAFY COTYLEDON2 (LEC2), and FUSCA3 (FUS3), produce similar morphogenic responses when overexpressed. The first of these genes characterized in Arabidopsis was LEC1 by Lotan et al. [33], in which a 35S::AtLEC1 cassette was introduced into Arabidopsis using Agrobacterium. Progeny seed were germinated and embryo-like structures were observed in many germinating plantlets. For example, cotyledon-like structures were observed to replace what should have been the first true leaves of the seedling. The embryo-like nature of these tissues was corroborated by other embryo characteristics, such as accumulation of cruciferin-A storage protein and oleosin RNAs. However, despite forming embryo-like structures, no functional ectopic somatic embryos were observed (i.e., embryo formation was incomplete).
In a similar study, a Citrus sinensis LEC1 paralog called L1L (LEC1-Like) was constitutively overexpressed in a 35S::CsL1L cassette after transformation of “Olinda” sweet orange or “Guoqing No. 1” Satsuma mandarin epicotyls using Agrobacterium [34]. In these experiments, the authors observed that the normally recalcitrant epicotyls formed some embryo-like structures after one month, and after another two months on elongation medium formed shoots with aberrant leaves. This suggests that in Citrus, L1L overexpression is sufficient to produce functional somatic embryos.
Uddenberg et al. [35] observed that overexpression of the PaHAP3A (a LEC1/L1L gene from Norway spruce, based on sequence information) did not result in ectopic somatic embryo formation in vegetative tissues. However, when expression was induced during zygotic embryo maturation, ectopic somatic embryos formed on the surface of the zygotic embryos. As noted by Srinivasan et al. [27] for BBM, this suggests that certain cell types may be more receptive to inductive signals, such as that being provided by the PaHAP3A protein [35].
In a variation on the general theme of using the CaMV 35S promoter to drive constitutive expression, Gazzarrini et al. [36] used an epidermal-specific promoter Meristem Layer1 (ML1) from Arabidopsis to drive expression of the Arabidopsis FUSCA gene (AtFUS3). Consistent with previous LEC1 results, ectopic overexpression of AtFUS3 resulted in the formation of cotyledon-like leaves that accumulated storage protein bodies (similar to the cells in an embryo). This observation is similar to phenotypes observed with LEC1 overexpression.
In experiments similar to those described above, stable transgenic lines were produced by introducing 35S::AtLEC2 into lec2-1 and lec2-5 mutant lines of Arabidopsis, and ectopic somatic embryos formed that were competent to germinate and produce plants [37]. However, the resultant plant phenotypes were aberrant and the authors did not comment on fertility. Nonetheless, it appears that LEC2 might result in more complete somatic embryo formation, compared to either LEC1, L1L, or FUS3. This is consistent with evidence indicating that LEC2 functions upstream of and activates both LEC1 and FUS3 [67], in which case the penetrance of the somatic embryo phenotype might be stronger in plants overexpressing LEC2.
Similar to observations in which overexpression of genes involved in meristem initiation and maintenance have increased a pre-existing embryogenic response [19,20,21,22,24,25,26], their ectopic overexpression has also been reported to stimulate the formation of somatic embryos where they would otherwise not be observed.
The first report of a “meristem” gene stimulating embryo formation was by Zuo et al. [38], who obtained an estradiol-induced activation-tagged pga6-1 mutant line in Arabidopsis that formed somatic embryos from root tips. This was confirmed to be AtWUS, which phenocopied the original activation-tagged mutant when expressed using either the estradiol-inducible or 35S promoters. Somatic embryo formation was also observed from a variety of tissues with de novo embryos from root tips being the most common observation. As with the observations by Boutilier et al. [26] with BBM, these results clearly demonstrate that WUS can stimulate the vegetative-to-embryonic transition.
By 2002 it was well-established that the proper expression of numerous genes was essential for morphogenesis. In keeping with this concept, Gallois et al. [39] analyzed the impact that two such genes might have on ectopic meristem initiation, using two different methods to control expression of STM and WUS in Arabidopsis (chemical induction and heat shock, respectively). Based on their observations of treated leaf tissue, the authors hypothesized that STM and WUS expression would produce clusters of cells adjacent to the WUS foci that represented incipient meristems (confirmed by a meristem-specific biomarker). These young ectopic meristems were initiated, but self-perpetuating meristems were not established.
In another interesting paper by Gallois et al. [40], stable transgenic lines were produced in Arabidopsis where WUS expression was activated through either HSP::CRE-mediated excision, or a GAL4-VP16 activation system. Upon either type of WUS activation, unique phenotypes were observed in root tips, and the type of response was dependent on other variables, such as hormone regime or co-expression of another morphogenic transcription factor. For example, when WUS was expressed alone in hormone-free medium, ectopic shoots and leaves were observed. When WUS was expressed in the presence of exogenous auxin (2,4-D), ectopic somatic embryos were formed. Finally, floral structures were observed when WUS was induced along with a constitutively expressed LEAFY gene, a master regulator of floral development [68].
Despite somatic embryogenesis from root tips being consistently observed when WUS expression was induced in Arabidopsis, estradiol induction of AtWUS in Nicotiana tabacum resulted in a direct organogenic response, where the root tips became swollen and developed green shoots rather than somatic embryos [41]. When AtWOX5 (a member of the WUS/WOX gene family expressed in the root tip) was substituted in these experiments, root tip swelling and green shoot formation were again observed [42].
While Gallois et al. [39] focused on the interaction of WUS and STM in Arabidopsis, the impact of the maize STM ortholog KNOTTED1 (KN1) has also been reported. When the maize KN1 gene was constitutively overexpressed (35S::ZmKN1) in Nicotiana tabacum, it resulted in a 3-fold increase in shoot organogenesis, relative to the NPTII-only control [43]. The increase in shoots in the KN1 treatment was obtained with no antibiotic or herbicide selection and no exogenous hormones in the media, and the resultant plants were bushy, with altered leaf morphology and underdeveloped roots.
The overexpression of KN1 (using the CaMV 35S promoter) can also bypass an intermediate callus phase, as reported by Nishimura et al. [44]. Constitutive expression of tobacco KN1 orthologs in Nicotiana tabacum resulted in a range of pleiotropic phenotypes. Transgenic plants containing the NTH20 (a knotted-like homeobox gene) expression cassette occasionally produced ectopic shoot meristems that would develop into small shoots with leaves emerging from the original leaf surface.
Other genes that play a role in shoot meristem formation are the CUP-SHAPED COTYLEDON genes CUC1 and CUC2. Using the Arabidopsis CUC1 and CUC2 genes, Daimon et al. [45] showed that overexpression of these genes under a strong promoter (CaMV 35S) led to the rapid production of adventitious shoots in transgenic calli derived from Arabidopsis hypocotyls. CUC1- and CUC2- overexpressing calli produced an average of 4.8 and 3.3 adventitious shoots per callus, respectively, while the controls produced 0.5 shoots that developed more slowly. In the absence of the phytohormones, no adventitious shoots were formed, indicating that CUC1 and CUC2 function was hormone-dependent.
In the context of using plant genes to improve plant transformation, genes involved in hormone signal transduction (whether receptors or downstream targets) also fall under our ’morphogenic’ classification. Two such candidates are the Arabidopsis ENHANCER OF SHOOT REGENERATION genes (ESR1 and ESR2), both identified through mutant screening and demonstrated to be involved in the cytokinin response pathway. As part of this characterization, it was demonstrated that overexpression of both ESR1 and ESR2 conferred cytokinin-independent shoot formation [47,48]. The Arabidopsis auxin-response gene MONOPTEROS (MP) is also of interest in this respect. In experiments focusing on the role of the MP gene on shoot formation, it was observed that using the endogenous promoter to drive expression of a C-terminally deleted gene referred to as MPΔ (lacking the domain involved in auxin/IAA interactions) increased the formation of shoot apical meristems from callus [49].
In addition to genes involved in the hormone signaling pathway, levels of hormones can affect the phenotypic response of genes involved in morphogenesis. Transgenic Arabidopsis explants overexpressing LEC2 under a DEX-inducible system produced somatic embryos in the presence of low auxin concentrations, while increasing the concentration resulted in the production of calli [46].
3. Strategies to Improve Transformation Using Morphogenic Genes
Given the focus of this review, it is easy to see why researchers in plant transformation would view the genes described above as potential tools in the transformation process. However, constitutive and strong expression of these morphogenic genes often caused undesired pleiotropic effects, including reduced fertility. In the examples discussed in this section, constitutive expression of the morphogenic genes (Figure 1A) or inducible expression in stably transformed plants (Figure 1B) were the predominant strategies. To render true utility, an additional step is required: combining optimized expression of the plant morphogenic genes being used with a robust method to limit expression after plant transformation/regeneration has occurred and their utility has expired. Such methods typically demonstrate improved transformation efficiency, enhanced regeneration, or both in a manner that produces healthy, fertile T0 plants. It should also be emphasized that to characterize gain-of-function phenotypes, as in many of the examples above, stable transgenic germplasm was produced using conventional selection methods (for example, after floral-dip transformation in Arabidopsis). As a result, the ectopic phenotype was evaluated in homogeneously transgenic tissues in seed-derived plants. This contrasts with expressing a morphogenic gene in a single cell surrounded by wild-type tissue, as when trying to use a morphogenic gene to recover transgenic events. In this situation, differential expression of the morphogenic gene provides a positive growth advantage or an identifiable phenotype (relative to wild-type cells) that can be used for selection. This is the case in the examples described below (see Table 1, Strategy C).
There are several reports that describe ectopic overexpression of morphogenic genes; however, deleterious pleiotropic phenotypes were observed in plants when constitutively expressed. Studies on how to control the timing and level of expression for these genes through downregulation or elimination have lagged. To date, we can identify four approaches to address this problem in the literature: (i) stimulating the morphogenic growth response through inducible expression of the morphogenic gene (Figure 1B) followed by removal of the inducing ligand to turn off expression, (ii) excision of the plant morphogenic gene (Figure 1C) when no longer needed, (iii) use of a plant promoter that turns off when no longer needed to permit normal growth and reproduction in transgenic plants, and (iv) using Agrobacterium-mediated delivery in a manner that favors transient expression of the morphogenic genes (Figure 1D,E).
Inducible expression has provided a robust method for using morphogenic genes to recover fertile transgenic plants. There are few reports on the transformation of recalcitrant species, and thus the first case presented here of using an inducible morphogenic gene for improving transformation and regeneration deserves emphasis. Pepper varieties (Capsicum annuum) have very poor transformation efficiencies and regenerative capacity. Heidmann et al. [50] transformed cotyledon explants of two sweet pepper varieties with a 35S::BnBBM~GR construct and cultured the explants on media supplemented with thidiazuron and DEX for 2 months. Emerging shoots were transferred to DEX-free elongation medium for 4 weeks and then pre-rooting medium for a month. Transformation efficiency with the regulated BBM expression was >1% compared to 0% with a 35S::GUS (β-glucuronidase) construct. This is an important step forward in what has historically been a very recalcitrant crop.
Also using the DEX-inducible AtBBM~GR system, Lutz and colleagues [51] described a method to obtain fertile, transgenic Arabidopsis plants from leaf cultures. In the absence of auxins, transgenic leaf explants produced prolific shoots in around 4 weeks in the presence of DEX, while explants on medium without the ligand did not produce shoots, and explants from wild-type plants became necrotic, irrespective of whether DEX was added or not. Recovered shoots were further cut into smaller segments and regenerated on medium containing DEX for 3 months. Removing the shoots from the DEX-containing medium allowed for regeneration of plantlets with normal flowering and seed formation over 3 months’ time. Fertile transgenic progeny were produced from the collected seeds.
Using the estradiol-inducible system [38], Wang et al. [52] identified PLANT GROWTH ACTIVATION genes, such as PGA37, which resulted in a vegetative-to-embryogenic transition when overexpressed in Arabidopsis. While the downstream targets have yet to be characterized, PGA37 was determined to encode a MYB118 transcription factor based on structural similarities within the DNA-binding domain. When PGA37 was expressed under inducible control, somatic embryos developed from root explants and was associated with increased LEC1 expression. Estradiol-induced expression of PGA37 in the presence of auxin produced green-yellowish embryonic calli in 7–10 days and generated somatic embryos upon culturing for 3–5 weeks. Upon removal of estradiol from the medium (thus downregulating PGA37 expression), the somatic embryos developed into healthy, fertile plantlets. Overexpression of a closely related homolog, MYB115, under the estradiol inducible system also led to the formation of somatic embryos from root explants.
When transforming recalcitrant species, such as trees, that in addition have maturation periods running into years, direct production of somatic embryos offers a relatively fast way for genetic manipulation [69]. One such example is use of the morphogenic gene LEC2 to generate transgenic plants in Theobroma cacao, as described by Shires and colleagues [53]. They identified the TcLEC2 transcription factor sequence and cloned it as a translational fusion with GR (the Glucocorticoid Receptor) driven by a CaMV 35S promoter. This DEX-inducible construct was used to transform cotyledon tissue of the variety Scavina-6 and cultured for about 6 months to screen for transgenic somatic embryos. Consistent with the transformation recalcitrance in T. cacao, only one transgenic embryo was recovered. This was proliferated by segmenting the cotyledons into several pieces to produce clonal-transformed somatic embryos. When tissue from these secondary embryos was placed on hormone-free media supplemented with DEX, multiple embryos were formed in 6 days. Levels of DEX up to 50 µM produced the most embryos (403) per 100 explants over a period of 4 months. In this experiment, a single somatic embryo was converted to a transgenic plant that developed normally. Young leaf tissue from the transgenic 35S::TcLEC2:GR plant was capable of prolific somatic embryo formation in the presence of DEX after 3 months, providing a promising method to regenerate secondary transgenics from leaf material.
In a recent publication using the estradiol-inducible system to control WOX gene expression, two combinations of Arabidopsis-derived WOX genes (WOX2 + WOX8 or WOX2 + WOX9) were evaluated in the presence of 1 μM 2,4-D for 10 days. Both combinations resulted in substantial plantlet regeneration from Nicotiana tabacum leaf pieces, in contrast to the wild-type control where no plantlet regeneration was observed [54].
Excision-based strategies to control morphogenic gene expression are another alternative. The first demonstration of this concept in a dicot was reported in Populus tomentosa using BBM and FLP-recombinase for excision [55]. These authors designed a T-DNA construct consisting of a single pair of FRT (FLP Recombination Target Sites) flanking both a heat-shock inducible promoter driving FLP recombinase expression and a CaMV 35S promoter driving expression of the Brassica campestris BBM gene cassette. Using Agrobacterium-mediated transformation, 21 callus cultures of Chinese white poplar were transformed with this T-DNA and cultured on hormone-free medium. Six of the 21 calli developed a total of 12 somatic embryos approximately four weeks after transformation, and half of the somatic embryos germinated to form plantlets that had a dwarf phenotype with small wrinkled leaves when cultured for 60 days. Heat shock treatment at 42 °C for 2 hours led to excision of both the FLP recombinase and BBM cassettes in four of the six plants, resulting in reappearance of the normal phenotype in regenerated plants.
The first demonstration of using morphogenic genes followed by excision to improve monocot transformation has only recently been reported by Lowe et al. [17]. After Agrobacterium-mediated transformation of immature embryos of a normally recalcitrant Pioneer maize inbred, a strongly expressed ZmBBM (using the Zea mays UBIQUITIN promoter) plus a weakly expressed ZmWUS2 (using the Agrobacterium NOPALINE SYNTHASE, or NOS, promoter) resulted in the stimulation of embryogenic callus. This combination of a weakly expressed WUS2 gene and a strongly expressed BBM gene resulted in high transformation frequencies when immature embryos of the maize inbred were transformed. The callus was placed on dry filter paper for 3 days to stimulate a desiccation-induced maize promoter from an ABA-responsive gene (RAB17) driving CRE recombinase, which then efficiently excised all three expression cassettes. After excision of the CRE, WUS2, and BBM transgenes, all that remained in the integrated T-DNA locus were the genes of interest (for example, herbicide resistance and/or a visual marker gene). This also improved transformation efficiency in many other difficult or recalcitrant genotypes (difficult corn inbreds, sorghum, sugarcane, and Indica rice), and permitted transformation of previously non-transformable explants, such as mature embryo sections (starting with mature, dry seed) or leaf segments from 7–14-day-old seedlings. This method is beginning to facilitate enhanced transformation of previously recalcitrant public cereal varieties, such as the maize inbred B73 and the sorghum variety P898012 [56].
More recently, Lowe et al. [57] described an improved transformation system for maize using two new promoters: the maize AXIG1 promoter (auxin-inducible) driving WUS2 and the maize PLTP promoter (PhosphoLipid Transfer Protein promoter) driving BBM. This new configuration of expression cassettes resulted in rapid formation of somatic embryos within 7 days, with germination of these newly formed somatic embryos producing plantlets ready for transplantation into soil and growth in the greenhouse within 21–30 days. Expression of both promoters was so low or confined to specific cell types (or tissues) in the plant that, even without excision of the PLTP::BBM and AXIG1::WUS2 cassettes, the resultant T0 plants were all robust and fertile. This new rapid transformation system has worked in all Pioneer and public inbreds tested, as well as in recalcitrant sorghum and wheat varieties.
A recurring theme in all the transformation methods described above is the necessity of controlling expression of the morphogenic genes in order to recover normal-phenotype plants, through either inducible expression, developmentally regulated expression, or excision. In the case of inducible or developmentally regulated expression of BBM and WUS2, the morphogenic genes and trait genes are linked within the same construct. Alternatively, constructs designed for excision are larger and more complex, but the morphogenic genes are removed prior to plant regeneration.
However, two additional alternatives exist that avoid both pitfalls. In the first alternative, two T-DNAs are introduced from the same Agrobacterium (containing both plasmids). In the second alternative, two separate Agrobacteria are used to introduce different T-DNA plasmids. Using either strategy, the two transgenic loci are later segregated away from each other [70]. The basis for the second approach was illuminated in a report by Florez et al. [29], who demonstrated that transient expression of TcBBM delivered by Agrobacterium was sufficient to stimulate somatic embryo formation, with no indication of the transgenic BBM cassette being detected in the somatic embryos. The authors then speculated that a co-transfection technique could be used to obtain transgenic plants by mixing an Agrobacterium strain containing a BBM expression cassette along with a trait-containing strain.
One final variation on using Agrobacterium to deliver transient morphogenic gene activity relies on a commonly observed characteristic of T-DNA delivery, with many labs over the years reporting on the phenomenon of T-DNA border read-through [71,72,73,74]. As the name implies, when the T-DNA is processed in the Agrobacterium before delivery, inefficient T-strand processing will produce some percentage of T-DNA molecules that are not terminated precisely at the Left Border (LB) but continue to include sequence beyond the LB. As a means of reducing this type of unwanted read-through, researchers have employed negative selectable markers [72] positioned beyond the LB (outside the T-DNA) to eliminate plant cells that had integrated these sequences. Others have tried to exploit inefficient T-strand processing by positioning a positive marker gene beyond the LB that could be used transiently, producing selectable marker-free transgenic plants [75]. By placing an Agrobacterium-derived IPT gene outside the LB, many cells would receive a mixture of T-DNAs, with the majority of the T-strands processed properly (only containing the trait), but with a smaller percentage of T-strands not terminated properly at the LB thus containing the flanking IPT gene. In these cells, transient IPT expression would stimulate cytokinin production and shoot proliferation, and when the processed T-strand integrated and the IPT-containing T-strand did not, this would result in the recovery of trait-containing transgenic plants that contain no selectable marker.
Since it has not been reported that shoot proliferation in response to IPT expression occurs in maize and other cereal crops, we tested WUS2 alone or WUS2 + BBM for transient somatic embryo formation and subsequent germination to produce T0 plants by positioning these cassettes outside the LB [76]. This resulted in a simple non-excision method for using WUS2 and BBM for transformation of recalcitrant maize genotypes that exploits inefficiencies of both T-strand processing and integration to allow for rapid transformation of maize while enriching for events with no WUS2 or BBM integration.
For example, Agrobacterium-mediated T-DNA delivery into the Pioneer maize inbred PH1V69 is very efficient; however, without BBM and WUS2 expression cassettes present in the T-DNA, it has so far proven impossible to recover transgenic T0 plants. Using inbred PH1V69, transformation was performed using an Agrobacterium strain LBA4404 that contained a single T-DNA plasmid [76]. Two versions were tested, with both containing two ‘Mock Trait’ expression cassettes within the T-DNA, a Setaria italica-derived UBIQUITIN promoter driving expression of a green fluorescent protein and an herbicide resistance cassette containing a Sorghum bicolor Acetolactate Synthase (ALS) promoter driving expression of HRA (a mutant maize ALS gene that confers resistance to sulfonylurea herbicides). For the first treatment, two expression cassettes were also positioned outside the LB of the T-DNA, with a PLTP promoter driving WUS2, and a PLTP promoter driving expression of BBM (WUS2/BBM). In a second treatment, only the PLTP::WUS2 was placed outside the LB (WUS2 only). Agrobacterium-infection, resting, somatic embryo maturation, and regeneration were performed as described in Lowe et al. [57] and T0 plants were analyzed for the presence of the marker genes plus WUS2 and BBM (when applicable) using qPCR. Starting with 196 immature embryos in each treatment, it was observed after analysis that the frequency of T0 plants (relative to the number of starting immature embryos) that were single-copy for marker genes and were negative for WUS2 and BBM (when applicable) was 12.2% and 10.2% for the ‘WUS2 only’ and ‘WUS2/BBM’ treatments, respectively. The percentage of T0 plants that contained either BBM/WUS2 or WUS2 alone (depending on the treatment) was 49% and 38%, respectively [76]. This suggests that while ‘read-through’ copies of WUS2 or WUS2/BBM were clearly having a positive impact in terms of stimulating somatic embryo formation, there was also some unavoidable integration of T-DNA sequences that also carried along the flanking sequence (‘backbone’ from the Agrobacterium T-DNA-containing plasmid). This is expected and, consequently, the method requires PCR screening to identify perfect, single-copy T-DNA events. However, it should also be noted that the frequency of recovering perfect, single-copy events was comparable to that observed for the excision method for this inbred [17]. This represents a viable alternative to excision as a means of creating high-quality transgenic events in recalcitrant monocot crops that do not contain helper genes (in this case WUS2 and BBM).
4. Conclusions
There has been meteoric progress in plant genome modification engendered by CRISPR/CAS9 (CLUSTERED REGULARLY-INTERSPERSED SHORT PALINDROMIC REPEATS and the CRISPR-ASSOCIATED 9 gene) over the past half-decade. This explosion has also brought into sharp focus the impediment presented by the state of transformation technology for many crops [1]. For maize, use of the morphogenic genes WUS2 and BBM has mitigated this bottleneck and has been used in-house for several years for all aspects of our genome modification programs. These include particle-gun-mediated creation of mini-chromosomes [77], CRISPR/CAS9-mediated mutagenesis or editing [78,79], and, of course, random Agrobacterium-mediated transformation [17,57]. From our experience with other cereal crops [17] and the progress by Mookkan and colleagues in recalcitrant public lines [56], we feel this technology should make all aspects of genome modification accessible to all cereals, with future enhancements continuing to simplify and improve this approach. Similarly, based on observed morphogenic responses in eudicots and gymnosperms, broadening these methods to include more plant species will hopefully continue to erode the barriers that make so many crops inaccessible for genome editing.
In the foreseeable future, however, finding a single solution that works across all crops is unlikely. Different species and even different varieties within the same species will require new combinations of morphogenic triggers (new combinations of genes or varied expression patterns) to produce either somatic embryos or new apical meristems for rapid production of genetically modified plants. Basic research over the past three decades has provided us with a detailed understanding of the genes that control morphogenesis, and the signaling networks that are so critical to meristem and embryo development, with new insights constantly being discovered. These insights will continue to provide the inspiration for testing morphogenic genes (or combinations of genes) and, along with new strategies to control or limit expression, will result in continued improvements that expand the range of plant species amenable to transformation. Hopefully, this will make plant transformation much more efficient, routine, and accessible for all crops of interest, and will alleviate this key bottleneck to crop improvement, enabling CRISPR/CAS-mediated genome modification in many important crops.
Author Contributions
Writing Review & Editing, N.S.; M.A.; K.L.; G.H.; S.B.; T.J.; B.G.-K.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Altpeter, F.; Springer, N.M.; Bartley, L.E.; Blechl, A.E.; Brutnell, T.P.; Citovsky, V.; Conrad, L.J.; Gelvin, S.B.; Jackson, D.P.; Kausch, A.P.; et al. Advancing Crop Transformation in the Era of Genome Editing. Plant Cell 2016, 28, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Smertenko, A.; Bozhkov, P.V. Somatic embryogenesis: Life and death processes during apical-basal patterning. J. Exp. Bot. 2014, 65, 1343–1360. [Google Scholar] [CrossRef] [PubMed]
- Snipes, S.A.; Rodriguez, K.; DeVries, A.E.; Miyawaki, K.N.; Perales, M.; Xie, M.; Reddy, G.V. Cytokinin stabilizes WUSCHEL by acting on the protein domains required for nuclear enrichment and transcription. PLOS Genet. 2018, 14, e1007351. [Google Scholar] [CrossRef] [PubMed]
- Grienenberger, E.; Fletcher, J.C. Polypeptide signaling molecules in plant development. Curr. Opin. Plant Biol. 2015, 23, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.K. Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Dev. Biol. 2010, 341, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef]
- Skoog, F.; Miller, C.O. Chemical regulation of growth and organ formation in plant tissues cultured. In Vitro Symp. Soc. Exp. Biol. 1957, 11, 118–130. [Google Scholar]
- Murashige, T. Plant propagation through tissue cultures. Ann. Rev. Plant Physiol. 1974, 25, 135–166. [Google Scholar] [CrossRef]
- Smigocki, A.C.; Owens, L.D. Cytokinin gene fused with a strong promoter enhances shoot organogenesis and zeatin levels in transformed plant cells. Proc. Natl. Acad. Sci. USA 1998, 85, 5131–5135. [Google Scholar] [CrossRef]
- Ondřej, M.; Macháčková, I.; Čatský, J.; Eder, J.; Hrouda, M.; Pospíšilová, J.; Synková, H. Potato transformation by T-DNA cytokinin synthesis gene. Biol. Plant. 1990, 32, 401–406. [Google Scholar] [CrossRef]
- Casanova, E.; Valdes, A.E.; Zuker, A.; Frenandez, B.; Vainstein, A.; Trillas, M.I. rolC-transgenic carnation plants: Adventitious organogenesis and levels of endogenous auxin and cytokinins. Plant Sci. 2004, 167, 551–560. [Google Scholar] [CrossRef]
- Gorpenchenko, T.Y.; Kiselev, K.V.; Bulgakov, V.P.; Tchernoded, G.K.; Bragina, E.A.; Khodakovskaya, M.V.; Koren, O.G.; Batygina, T.B.; Zhuravlev, Y.N. The Agrobacterium rhizogenes rolC-gene-induced somatic embryogenesis and shoot organogenesis in Panax ginseng transformed calluses. Planta 2006, 223, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Otten, L. The Agrobacterium Phenotypic Plasticity (Plast) Genes. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Roeckel, P.; Oancia, T.; Drevet, J.R. Phenotypic alterations and component analysis of seed yield in transgenic Brassica napus plants expressing the tzs gene. Physiol. Plant. 1998, 102, 243–249. [Google Scholar] [CrossRef]
- Choi, Y.I.; Noh, E.W.; Choi, K.S. Low level expression of prokaryotic tzs gene enhances growth performance of transgenic poplars. Trees 2009, 23, 7441–7750. [Google Scholar] [CrossRef]
- Gordon-Kamm, W.; Dilkes, B.P.; Lowe, K.; Hoerster, G.; Sun, X.; Ross, M.; Church, L.; Bunde, C.; Farrell, J.; Hill, P.; et al. Stimulation of the cell cycle and maize transformation by disruption of the plant retinoblastoma pathway. Proc. Nat. Acad. Sci. USA 2002, 99, 11975–11980. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic Regulators Baby boom and Wuschel Improve Monocot Transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.; Velle-Calzada, J.P.; Hartog, M.V.; Schmidt, E.D.; Boutilier, K.; Grossniklaus, U.; de Vries, S.C. The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE 1 gene is expressed in developing ovules and embryos and enhances embryogenic competence in culture. Plant Physiol. 2001, 127, 803–816. [Google Scholar] [CrossRef]
- Pérez-Pascual, D.; Jiménez-Guillen, D.; Villanueva-Alonzo, H.; Souza-Perera, R.; Godoy-Hernández, G.; Zúñiga-Aguilar, J.J. Ectopic expression of the Coffea canephora SERK1 homolog-induced differential transcription of genes involved in auxin metabolism and in the developmental control of embryogenesis. Physiol. Plant. 2018, 163, 530–551. [Google Scholar] [CrossRef]
- Harding, E.W.; Tang, W.; Nichols, K.W.; Fernandez, D.E.; Perry, S.E. Expression and maintenance of embryogenic potential is enhanced through constitutive expression of AGAMOUS-LIKE15. Plant Physiol. 2003, 133, 653–663. [Google Scholar] [CrossRef]
- Thakare, D.; Tang, W.; Hill, K.; Perry, S.E. The MADS-domain transcriptional regulator AGAMOUS-LIKE15 promotes somatic embryo development in Arabidopsis and soybean. Plant Physiol. 2008, 146, 1663–1672. [Google Scholar] [CrossRef]
- Yang, Z.; Li, C.; Wang, Y.; Zhang, C.; Wu, Z.; Zhang, X.; Liu, C.; Li, F. GhAGL15s, preferentially expressed during somatic embryogenesis, promote embryogenic callus formation in cotton (Gossypium hirsutum L.). Mol. Genet. Genom. 2014, 289, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Herrera, A.; Gonzalez, A.K.; Moo, R.C.; Quiroz-Figueroa, F.R.; Loyola-Vargas, V.M.; Rodriguez-Zapata, L.C.; Burgeff D′Hondt, C.; Suárez-Solís, V.M.; Castaño, E. Expression of WUSCHEL in Coffea canephora causes ectopic morphogenesis and increases somatic embryogenesis. Plant Cell Tissue Organ Cult. 2008, 94, 171–180. [Google Scholar] [CrossRef]
- Elhiti, M.; Tahir, M.; Gulden, R.H.; Khamiss, K.; Stasolla, C. Modulation of embryo-forming capacity in culture through the expression of Brassica genes involved in the regulation of the shoot apical meristem. J. Exp. Bot. 2010, 61, 4069–4085. [Google Scholar] [CrossRef] [PubMed]
- Bouchabké-Coussa, O.; Obellianne, M.; Linderme, D.; Montes, E.; Maia-Grondard, A.; Vilaine, F.; Pannetier, C. Wuschel overexpression promotes somatic embryogenesis and induces organogenesis in cotton (Gossypium hirsutum L.) tissues cultured in vitro. Plant Cell Rep. 2013, 32, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.M.; van Lammeren, A.A.; Miki, B.L.; et al. Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, C.; Liu, Z.; Heidmann, I.; Supena, E.D.; Fukuoka, H.; Joosen, R.; Lambalk, J.; Angenent, G.; Scorza, R.; Custers, J.B.; et al. Heterologous expression of the BABY BOOM AP2/ERF transcription factor enhances the regeneration capacity of tobacco (Nicotiana tabacum L.). Planta 2007, 225, 341–351. [Google Scholar] [CrossRef]
- El Ouakfaoui, S.; Schnell, J.; Abdeen, A.; Colville, A.; Labbé, H.; Han, S.; Baum, B.; Laberge, S.; Miki, B. Control of somatic embryogenesis and embryo development by AP2 transcription factors. Plant Mol. Biol. 2010, 74, 313–326. [Google Scholar] [CrossRef]
- 29 Florez, S.L.; Erwin, R.L.; Maximova, S.N.; Guiltinan, M.J.; Curtis, W.R. Enhanced somatic embryogenesis in Theobroma cacao using the homologous BABY BOOM transcription factor. BMC Plant Biol. 2015, 16, 121. [Google Scholar] [CrossRef]
- Morcillo, F.; Gallard, A.; Pillot, M.; Jouannic, S.; Aberlenc-Bertossi, F.; Collin, M.; Verdeil, J.L.; Tregear, J.W. EgAP2-1, an AINTEGUMENTA-like (AIL) gene expressed in meristematic and proliferating tissues of embryos in oil palm. Planta 2007, 226, 1353–1362. [Google Scholar] [CrossRef]
- Tsuwamoto, R.; Yokoi, S.; Takahata, Y. Arabidopsis EMBRYOMAKER encoding an AP2 domain transcription factor plays a key role in developmental change from vegetative to embryonic phase. Plant Mol. Biol. 2010, 73, 481–492. [Google Scholar] [CrossRef]
- Mursyanti, E.; Purwantoro, A.; Moeljopawiro, S.; Semiarti, E. Induction of Somatic Embryogenesis through Overexpression of ATRKD4 Genes in Phalaenopsis “Sogo Vivien”. Indones. J. Biotechnol. 2015, 20, 42–53. [Google Scholar] [CrossRef]
- Lotan, T.; Ohto, M.; Yee, K.M.; West, M.A.; Lo, R.; Kwong, R.W.; Yamagishi, K.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells. Cell 1998, 93, 1195–1205. [Google Scholar] [CrossRef]
- Zhu, S.-P.; Wang, J.; Ye, J.-L.; Zhu, A.-D.; Guo, W.-W.; Deng, X.-X. Isolation and characterization of LEAFY COTYLEDON 1-LIKE gene related to embryogenic competence in Citrus sinensis. Plant Cell Tiss. Organ Cult. 2014, 119, 1–13. [Google Scholar] [CrossRef]
- Uddenberg, D.; Abrahamsson, M.; von Arnold, S. Overexpression of PaHAP3A stimulates differentiation of ectopic embryos from maturing somatic embryos of Norway spruce. Tree Genet. Genomes 2016, 12, 18. [Google Scholar] [CrossRef]
- Gazzarrini, S.; Tsuchiya, Y.; Lumba, S.; Okamoto, M.; McCourt, P. The transcription factor FUSCA3 controls developmental timing in Arabidopsis through the hormones gibberellin and abscisic acid. Dev. Cell 2004, 7, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Kwong, L.W.; Yee, K.M.; Pelletier, J.; Lepiniec, L.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON2 encodes a B3 domain transcription factor that induces embryo development. Proc. Natl. Acad. Sci. USA 2001, 98, 11806–11811. [Google Scholar] [CrossRef]
- Zuo, J.; Niu, Q.W.; Frugis, G.; Chua, N.H. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. Plant J. 2002, 30, 349–359. [Google Scholar] [CrossRef]
- Gallois, J.L.; Woodward, C.; Reddy, G.V.; Sablowski, R. Combined SHOOT MERISTEMLESS and WUSCHEL trigger ectopic organogenesis in Arabidopsis. Development 2002, 129, 3207–3217. [Google Scholar]
- Gallois, J.L.; Nora, F.R.; Mizukami, Y.; Sablowski, R. WUSCHEL induces shoot stem cell activity and developmental plasticity in the root meristem. Genes Dev. 2004, 18, 375–380. [Google Scholar] [CrossRef]
- Rashid, S.Z.; Yamaji, N.; Kyo, M. Shoot formation from root tip region: A developmental alteration by WUS in transgenic tobacco. Plant Cell Rep. 2007, 26, 1449–1455. [Google Scholar] [CrossRef]
- Rashid, S.Z.; Kyo, M. Ectopic Expression of WOX5 Dramatically Alters Root-tip Morphology in Transgenic Tobacco. Transgenic Plant J. 2009, 3, 92–96. [Google Scholar]
- Luo, K.; Zheng, X.; Chen, Y.; Xiao, Y.; Zhao, D.; McAvoy, R.; Pei, Y.; Li, Y. The maize Knotted1 gene is an effective positive selectable marker gene for Agrobacterium-mediated tobacco transformation. Plant Cell Rep. 2006, 25, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Tamaoki, M.; Sakamoto, T.; Matsuoka, M. Over-Expression of Tobacco knotted 1-Type Classl Homeobox Genes Alters Various Leaf Morphology. Plant Cell Physiol. 2000, 41, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Daimon, Y.; Takabe, K.; Tasaka, M. The CUP-SHAPED COTYLEDON genes promote adventitious shoot formation on calli. Plant Cell. Physiol. 2003, 44, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Wójcikowska, B.; Jaskóła, K.; Gąsiorek, P.; Meus, M.; Nowak, K.; Gaj, M.D. LEAFY COTYLEDON2 (LEC2) promotes embryogenic induction in somatic tissues of Arabidopsis, via YUCCA-mediated auxin biosynthesis. Planta 2013, 238, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Banno, H.; Ikeda, Y.; Niu, Q.W.; Chua, N.H. Overexpression of Arabidopsis ESR1 induces initiation of shoot regeneration. Plant Cell 2001, 13, 2609–2618. [Google Scholar] [CrossRef]
- Ikeda, Y.; Banno, H.; Niu, Q.W.; Howell, S.H.; Chua, N.H. The ENHANCER OF SHOOT REGENERATION 2 gene in Arabidopsis regulates CUP-SHAPED COTYLEDON 1 at the transcriptional level and controls cotyledon development. Plant Cell Physiol. 2006, 47, 1443–1456. [Google Scholar] [CrossRef]
- Ckurshumova, W.; Smirnova, T.; Marcos, D.; Zayed, Y.; Berleth, T. Irrepressible MONOPTEROS/ARF5 promotes de novo shoot formation. New Phytol. 2014, 204, 556–566. [Google Scholar] [CrossRef]
- Heidmann, I.; de Lange, B.; Lambalk, J.; Angenent, G.C.; Boutilier, K. Efficient sweet pepper transformation mediated by the BABY BOOM transcription factor. Plant Cell Rep. 2011, 30, 1107–1115. [Google Scholar] [CrossRef]
- Lutz, K.A.; Martin, C.; Khairzada, S.; Maliga, P. Steroid-inducible BABY BOOM system for development of fertile Arabidopsis thaliana plants after prolonged tissue culture. Plant Cell Rep. 2015, 34, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Niu, Q.-W.; Teng, C.; Li, C.; Mu, J.; Chua, N.-H.; Zuo, J. Overexpression of PGA37/MYB118 and MYB115 promotes vegetative-to-embryonic transition in Arabidopsis. Cell Res. 2009, 19, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Shires, M.E.; Florez, S.L.; Lai, T.S.; Curtis, W.R. Inducible somatic embryogenesis in Theobroma cacao achieved using the DEX-activatable transcription factor-glucocorticoid receptor fusion. Biotechnol. Lett. 2017, 39, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Kyo, M.; Maida, K.; Nishioka, Y.; Matsui, K. Coexpression of WUSCHEL related homeobox (WOX) 2 with WOX8 or WOX9 promotes regeneration from leaf segments and free cells in Nicotiana tabacum L. Plant Biotechnol. 2018, 35, 23–30. [Google Scholar] [CrossRef]
- Deng, W.; Li, Z.; Luo, K.; Yang, Y. A novel method for induction of plant regeneration via somatic embryogenesis. Plant Sci. 2009, 177, 43–48. [Google Scholar] [CrossRef]
- Mookkan, M.; Nelson-Vasilchik, K.; Hague, J.; Zhang, Z.J.; Kausch, A.P. Selectable marker independent transformation of recalcitrant maize inbred B73 and sorghum P898012 mediated by morphogenic regulators BABY BOOM and WUSCHEL2. Plant Cell Rep. 2017, 36, 1477–1491. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; La Rota, M.; Hoerster, G.; Hastings, C.; Wang, N.; Chamberlin, M.; Wu, E.; Jones, T.; Gordon-Kamm, W. Rapid genotype “independent” Zea mays L. (maize) transformation via direct somatic embryogenesis. In Vitro Cell Dev. Biol. Plant. 2018, 54, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Karlova, R.; Boeren, S.; Russinova, E.; Aker, J.; Vervoort, J.; de Vries, S. The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE1 protein complex includes brassinosteroid-insensitive1. Plant Cell 2006, 18, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Gou, X.; Yin, H.; He, K.; Du, J.; Yi, J.; Xu, S.; Lin, H.; Clouse, S.D. Genetic Evidence for an Indispensable Role of Somatic Embryogenesis Receptor Kinases in Brassinosteroid Signaling. PLOS Genet. 2012, 8, e1002452. [Google Scholar] [CrossRef]
- Santiago, J.; Henzler, C.; Hothorn, M. Molecular mechanism for plant steroid receptor activation by somatic embryogenesis co-receptor kinases. Science 2013, 341, 889–892. [Google Scholar] [CrossRef]
- Hohmann, U.; Lau, K.; Hothorn, M. The structural basis of ligand perception and signal activation by receptor kinases. Ann. Rev. Plant Biol. 2017, 68, 109–137. [Google Scholar] [CrossRef]
- Laux, T.; Mayer, K.F.; Berger, J.; Jurgens, G. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar] [PubMed]
- Long, J.A.; Moan, E.I.; Medford, J.I.; Barton, M.K. A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA 2/Ethylene Responsive Factor (AP 2/ERF) transcription factors: Mediators of stress responses and developmental programs. New Phytol. 2013, 199, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Waki, T.; Kiki, T.; Watanabe, R. The Arabidopsis RWP-RK Protein RKD4 Triggers Gene Expression and Pattern Formation in Early Embryogenesis. Curr. Biol. 2011, 21, 1277–1281. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Palmer, T.M.; Lukowitz, W. The RWP-RK Factor GROUNDED Promotes Embryonic Polarity by Facilitating YODA MAP Kinase Signaling. Curr. Biol. 2011, 21, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Santos Mendoza, M.; Dubreucq, B.; Miquel, M.; Caboche, M.; Lepiniec, L. LEAFY COTYLEDON 2 activation is sufficient to trigger the accumulation of oil and seed specific mRNAs in Arabidopsis leaves. FEBS Lett. 2005, 579, 4666–4670. [Google Scholar] [CrossRef] [PubMed]
- Weigel, D.; Alvarez, J.; Smyth, D.R.; Yanofsky, M.F.; Meyerowitz, E.M. LEAFY controls floral meristem identity in Arabidopsis. Cell 1992, 69, 843–859. [Google Scholar] [CrossRef]
- Nagle, M.; Déjardin, A.; Pilate, G.; Strauss, S.H. Opportunities for innovation in genetic transformation of forest trees. Front. Plant Sci. 2018, 9, 1443. [Google Scholar] [CrossRef]
- Miller, M.; Tagliani, L.; Wang, N.; Berka, B.; Bidney, D.; Zhao, Z.-Y. High Efficiency Transgene Segregation in Co-Transformed Maize Plants using an Agrobacterium Tumefaciens 2 T-DNA Binary System. Transgenic Res. 2002, 11, 381–396. [Google Scholar] [CrossRef]
- Kononov, M.E.; Bassuner, B.; Gelvin, S.B. Integration of T-DNA binary vector ‘backbone’ sequences into the tobacco genome: Evidence for multiple complex patterns of integration. Plant J. 1997, 11, 945–957. [Google Scholar] [CrossRef]
- Hanson, B.; Engler, D.; Moy, Y.; Newman, B.; Ralston, E.; Gutterson, N. A simple method to enrich an Agrobacterium-transformed population for plants containing only T-DNA sequences. Plant J. 1999, 19, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Lee, J.; Jun, S.H.; Park, S.; Kang, H.G.; Kwon, S.; An, G. Transgene structures in TDNA-inserted rice plants. Plant Mol. Biol. 2003, 52, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Podevin, N.; De Buck, S.; De Wilde, C.; Depicker, A. Insights into recognition of the T-DNA border repeats as termination sites for T-strand synthesis by Agrobacterium tumefaciens. Transgenic Res. 2006, 15, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Richael, C.; Kalyeava, M.; Chretien, R.C.; Rommens, C.M. Cytokinin vectors mediate marker-free and backbone-free plant transformation. Transgenic Res. 2008, 17, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; Hoerster, G.; Anand, A.; Arling, M.; Wang, N.; McBride, K.; Gordon-Kamm, W. Transient expression of morphogenic genes positioned outside the T-DNA borders results in rapid formation of somatic embryos and fertile transgenic cereal plants. Unpublished; manuscript in preparation.
- Ananiev, E.V.; Wu, C.; Chamberlin, M.A.; Svitashev, S.; Schwartz, C.; Gordon-Kamm, W.; Tingey, S. Artificial chromosome formation in maize (Zea mays L.). Chromosoma 2009, 118, 157–177. [Google Scholar] [CrossRef] [PubMed]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted Mutagenesis, Precise Gene Editing, and Site-Specific Gene Insertion in Maize Using Cas9 and Guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Cigan, A.M. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).