Abstract
Brassicaceae species employ both self-compatibility and self-incompatibility systems to regulate post-pollination events. Arabidopsis halleri is strictly self-incompatible, while the closely related Arabidopsis thaliana has transitioned to self-compatibility with the loss of functional S-locus genes during evolution. The downstream signaling protein, ARC1, is also required for the self-incompatibility response in some Arabidopsis and Brassica species, and its gene is deleted in the A. thaliana genome. In this study, we attempted to reconstitute the SCR-SRK-ARC1 signaling pathway to restore self-incompatibility in A. thaliana using genes from A. halleri and B. napus, respectively. Several of the transgenic A. thaliana lines expressing the A. halleri SCR13-SRK13-ARC1 transgenes displayed self-incompatibility, while all the transgenic A. thaliana lines expressing the B. napus SCR1-SRK1-ARC1 transgenes failed to show any self-pollen rejection. Furthermore, our results showed that the intensity of the self-incompatibility response in transgenic A. thaliana plants was not associated with the expression levels of the transgenes. Thus, this suggests that there are differences between the Arabidopsis and Brassica self-incompatibility signaling pathways, which perhaps points to the existence of other factors downstream of B. napus SRK that are absent in Arabidopsis species.
1. Introduction
Self-incompatibility is one of the most important breeding systems present in many hermaphroditic flowering plants as it causes self-pollen rejection to promote out-crossing. Self-incompatibility systems under investigation include species in Brassicaceae, Solanaceae, Rosaceae, Scrophulariaceae and Papaveraceae [1,2,3,4]. In the Brassicaceae (crucifer family), self-incompatibility is controlled by tightly linked multi-allelic genes at the S-locus region, which rarely recombine and so have collectively been named S-haplotypes [4]. Three S-locus-linked genes have been identified in Brassica species. The S-locus glycoprotein (SLG) gene encodes an abundant, secreted glycoprotein that is located in the cell wall of stigma papillae, and SLG has been reported to be connected to pollen adhesion on stigmatic surfaces [5,6]. The S-locus receptor kinase (SRK) is the female determinant in stigma, and mediates the self-incompatibility signaling pathway [7,8,9]. Finally, the S-locus Protein 11/S-locus Cysteine-Rich (SP11/SCR) gene encodes the male determinant and is expressed in the anther tapetum and pollen [10,11,12]. SP11/SCR is recognized by SRK as a ligand in an S-haplotype-specific manner, and SRK is then autophosphorylated to induce various signaling cascades [13,14].
Models of the downstream self-incompatibility signaling pathways have been proposed in Brassica species [15,16]. The M locus protein kinase (MLPK) was identified as a positive regulator of self-incompatible response, since the B. rapa mlpk mutant lost its self-incompatibility phenotype and knocking out the B. napus MLPK genes by CRISPR/Cas9 completely knocked out the self-incompatibility response [17,18]. MLPK is tethered on the plasma membrane and can be phosphorylated by SRK by forming a complex with SRK to mediate signal transduction in self-incompatibility response [19,20]. ARC1 (Armed Repeat Containing 1) and THL1/2 (thioredoxin-h like 1/2) were isolated for interacting with the kinase domain of SRK in yeast two-hybrid analysis [21,22]. THL1/2 were believed to regulate self-incompatibility response negatively [21,23]. Down-regulation of ARC1 expression in the stigma caused a breakdown of self-incompatibility in the B. napus line ‘W1′ [22,24]. ARC1 is an E3 ubiquitin ligase that can direct the degradation of target proteins required for compatible pollen responses [25]. One target is EXO70A1, which directly interacts with ARC1 and can be ubiquitinated by ARC1 in vitro. This action is proposed to block the transfer of secretory vesicles to the pollen-stigma contact site resulting in pollen rejection [26]. GLO1 (glyoxalase I), a key enzyme involved in detoxification of methylglyoxal, and has been identified as another ARC1 target. ARC1 was proposed to negatively regulate GLO1 in self-pollen rejection, leading to the accumulation of the methylglyoxal toxin in papillar cells [27].
In addition to Brassica species, Arabidopsis species have been used for self-incompatibility studies in Brassicaceae. By analyzing the sequences of Arabidopsis S-locus genes, naturally self-incompatible species, such as Arabidopsis lyrata and Arabidopsis halleri, were shown to have intact SRK and SCR/SP11 genes. Arabidopsis thaliana ecotypes are self-compatible and do not carry functional SRK or SCR/SP11 genes [28,29,30,31,32]. Efforts have been made to reconstruct the self-incompatibility response in A. thaliana by introducing functional SCR and SRK genes, and significant differences were observed in the stability and strength of the self-incompatibility traits among various ecotypes tested [31,33,34,35,36,37]. When B. napus self-incompatibility stigma genes (SLG, SRK and ARC1 genes) were transformed into A. thaliana, the transgenic plants still produced seeds, and their stigmas failed to reject the corresponding self-incompatible B. napus W1 pollen [38]. Similarly, transgenic A. thaliana ecotype Col-0 plants expressing the A. lyrata SCRb-SRKb genes were fully self-fertile [33,35,39]. However, when the same experiments were conducted with the A. thaliana C24 ecotype, the transformation of the AlSCRb-AlSRKb genes resulted in transgenic plants showing a strong self-incompatibility response [33,35,40]. Self-incompatible A. thaliana C24 lines were also created using the A. halleri AhSCR1 and AhSRK1 genes of the most recessive S1 haplotype [41]. Finally, ARC1 was found to be a pseudogene in A. thaliana, and a homolog of MLPK, AtAPK1b, did not appear to be involved in regulating self-incompatibility in transgenic A. thaliana, suggesting that SRK-mediated signaling in transgenic A. thaliana might be different from the Brassica models [37,42]. This is also supported by differences in the post-pollination cellular responses of Brassica and Arabidopsis stigmas [15,16]. Nevertheless, stable self-incompatible A. thaliana were finally generated in the Col-0 ecotype when AlARC1 or BnARC1 were expressed along with the AlSCRb-AlSRKb transgenes [43].
Here, we further examine the question of whether Brassica and Arabidopsis self-incompatibility genes can share a common self-incompatibility signaling pathway in transgenic A. thaliana using a new set of self-incompatibility genes from B. napus and A. halleri. The B. napus S1-haplotype SCR/SP11 and SRK genes [44,45,46] along with BnARC1 were transformed into two A. thaliana accessions, Col-0 and C24. As both the pollen (BnSCR/SP111) and stigma (BnSRK1, BnARC1) factors were included for this study, this differs from a previous study [38] where only stigma B. napus factors were transformed. A new set of Arabidopsis self-incompatibility genes from A. halleri S13-haplotype SCR/SP11 and SRK genes [47], along with AhARC1, were also tested into Col-0 as a comparison. The S1- and S13-haplotypes were previously found to be the most dominant S-haplotypes in B. napus and A. halleri, respectively [41,44,45]. Our results show that self-incompatibility genes from B. napus were not sufficient to restore the self-incompatibility in both the Col-0 and C24 ecotypes, while self-incompatibility was successfully transmitted into Col-0 with the A. halleri self-incompatibility genes.
2. Results
2.1. Reconstruction of ARC1-Mediated Self-Incompatibility in A. thaliana Col-0 using A. halleri Self-Incompatibility Genes
To examine if the SCR-SRK-ARC1 model is conserved using genes from another naturally self-incompatible Arabidopsis species, A. halleri SCR13-SRK13 genes were introduced into A. thaliana Col-0 plants in the presence or absence of AhARC1, with all three transgenes in a single construct. Thirty and 34 independent T1 Col-0 lines were generated with the addition of AhSCR13-AhSRK13 and AhSCR13-AhSRK13-AhARC1, respectively (Table 1). The self-incompatibility phenotype of the transgenic plants was examined by staining their stigmas with aniline blue to visualize the pollen grains and the pollen tubes growing into papillar cells. Pollinated pistils from the different transgenic lines showed variations in the levels of rejection/acceptance of self-pollen, which was also reported by Indriolo et al. [43]. Accordingly, the phenotypes were defined as self-compatible (similar to Col-0), moderately self-incompatible (>5 pollen tubes/pistil, but visibly reduced compared to Col-0) or strongly self-incompatible (<5 pollen tubes/pistil).

Table 1.
Phenotypes of transgenic Arabidopsis thaliana Col-0 plants.
All 30 independent transgenic T1 plants with AhSCR13-AhSRK13 were found to be fully self-compatible, as they could accept self-pollen similarly to that observed for wild-type self-pollinated Col-0 plants in stage-14 stigmas (Table 1 and Figure 1A–D). In contrast, four out of 34 transgenic T1 plants carrying AhARC1 along with AhSCR13-AhSRK13 showed strong self-incompatibility (Table 1 and Figure 1E,F). To ensure that the strong self-incompatibility phenotype was not caused by other reproduction factors, reciprocal crosses were performed between the transgenic lines and wild-type Col-0 plants, leading to complete self-compatibility (Supplementary Figure S1). In addition, 16 of the 34 AhSCR13-AhSRK13-AhARC1 transgenic T1 plants displayed moderate self-incompatibility, and the final 14 AhSCR13-AhSRK13-AhARC1 transgenic T1 plants were fully self-compatible (Table 1 and Figure 1G–J). The fact that only the combination of AhSCR13-AhSRK13-AhARC1 transgenes could reconstruct self-incompatibility in the A. thaliana Col-0 ecotype confirms a role for ARC1 in this self-pollen rejection signaling pathway, as previously reported by Indriolo et al. [43].
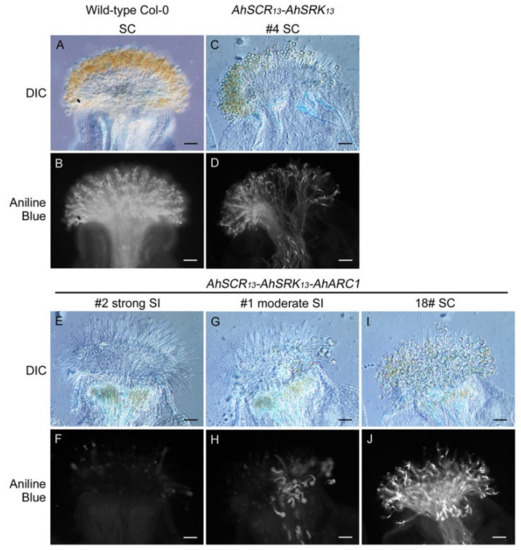
Figure 1.
Pollen tube germination and growth in transgenic A. thaliana Col-0 plants. (A,B) Wild-type A. thaliana Col-0 self-pollinated stigma. (C,D) Transgenic A. thaliana Col-0 AhSCR13-AhSRK13 line-4 self-pollinated stigma. (E–J) Transgenic A. thaliana Col-0 AhSCR13-AhSRK13-AhARC1 line-2, -1 and -18 self-pollinated stigmas. Differential interference contrast (DIC) and aniline blue-stained images are shown for each sample. SC: self-compatible; SI: self-incompatible. Bars = 50 μm.
2.2. Self-Incompatibility Related Genes of B. napus are Not Sufficient to Restore Self-Incompatibility in both A. thaliana Col-0 and C24 Ecotypes
To test whether the B. napus SCR1-SRK1 genes were sufficient to restore self-incompatibility in A. thaliana plants, and to test if BnARC1 was also required, the BnSCR1-BnSRK1 genes were transformed with or without BnARC1 into the A. thaliana Col-0 and C24 ecotypes. Independent transgenic plants were obtained for each transformation event, and the pollination phenotypes were noted (Table 1 and Table 2). Twenty-one independent T1 Col-0 plants were obtained with the transformation of BnSCR1-BnSRK1 and all of them exhibited a self-compatible phenotype in stage-14 self-pollinated pistils stained with aniline blue (Table 1 and Figure 2A–D). Similarly, 13 independent T1 C24 lines were created with the BnSCR1-BnSRK1 transgenes and showed a similar level of self-pollen acceptance to wild-type C24 plants at developmental stage 14 (Table 2 and Figure 2G–J). Previous studies have reported that A. thaliana Col-0 plants carrying the A. lyrata SCRb-SRKb transgenes were fully self-compatible, while for A. thaliana C24 plants transformed with the SCRb-SRKb transgenes, self-incompatibility was successfully reinstated [33,35,40]. However, the introduction of B. napus BnSCR1-BnSRK1 into C24 plants failed to restore self-incompatibility, indicating that BnSCR1-BnSRK1 may need other unknown factors in the self-incompatibility pathway that do not exist in the C24 ecotype.
Table 2.
Phenotypes of transgenic A. thaliana C24 plants.
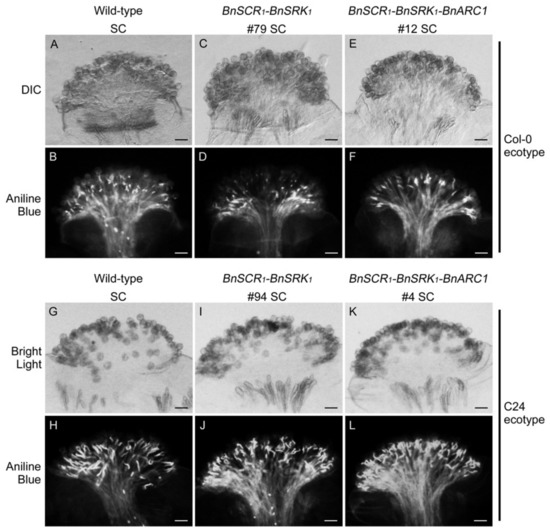
Figure 2.
Pollen tube germination and growth in transgenic A. thaliana Col-0 and C24 plants. (A,B) Wild-type A. thaliana Col-0 self-pollinated stigma. (C,D) Transgenic A. thaliana Col-0 BnSCR1-BnSRK1 line-79 self-pollinated stigma. (E,F) Transgenic A. thaliana Col-0 BnSCR1-BnSRK1 -BnARC1 line-12 self-pollinated stigma. (G,H) Wild-type A. thaliana C24 self-pollinated stigma. (I,J) Transgenic A. thaliana C24 BnSCR1-BnSRK1 line-94 self-pollinated stigma. (K,L) Transgenic A. thaliana C24 BnSCR1-BnSRK1 -BnARC1 line-4 self-pollinated stigma. Differential interference contrast (DIC) (or regular bright light) and aniline blue-stained images are shown for each sample. SC: self-compatible. Bars = 50 μm.
To examine if the transformation of BnARC1 along with BnSCR1-BnSRK1 could produce a functional self-incompatible pathway in Col-0 and C24 plants, 12 and 9 independent T1 Col-0 and C24 plants were generated, respectively. The phenotypes of these transgenic lines were again scored for self-pollen rejection by aniline blue staining. Unexpectedly and in contrast to the AhSCR13-AhSRK13-AhARC1 transgenic Col-0 plants, all Col-0 lines carrying BnSCR1-BnSRK1-BnARC1 genes showed self-compatible phenotypes (Table 1 and Figure 2E,F). In addition, no self-incompatible or moderately self-incompatible phenotypes were observed in the BnSCR1-BnSRK1-BnARC1 transgenic C24 lines (Table 2 and Figure 2K,L). These results indicate that the addition of BnARC1 with BnSCR1-BnSRK1 also does not work to restore the self-incompatibility signaling pathway in the A. thaliana Col-0 and C24 ecotypes.
2.3. Seed Production is Significantly Reduced in Col-0 Lines Expressing AhSCR13-AhSRK13-AhARC1
As the transformation of SCRb-SRKb-ARC1 was previously found to lead to an approach herkogamous phenotype in A. thaliana Col-0 and Sha plants [43], we examined whether the transgenic SCR-SRK-ARC1 Col-0 and C24 lines displayed any changes in their floral morphology. The approach herkogamous phenotype was only exhibited in the strongly self-incompatible Col-0 plants carrying the AhSCR13-AhSRK13-AhARC1 transgenes (Figure 3J,K and Supplementary Figure S2). The physical stigma-anther separation (SAS) was measured in flowers from wild-type and representative self-incompatible Col-0 plants. A positive SAS value was indicative of approach herkogamy which is consistent with the self-incompatible phenotype in the AhSCR13-AhSRK13-AhARC1 plants (Figure 3L). However, all the self-compatible transgenic lines showed a normal flower trait with the anthers positioned above the stigmas, similar to wild-type Col-0 and C24 plants, which ensures that the released pollen falls onto the stigmas (Supplementary Figure S2). Therefore, the approach herkogamous phenotype correlates with the degree of self-incompatibility in the transgenic A. thaliana plants.
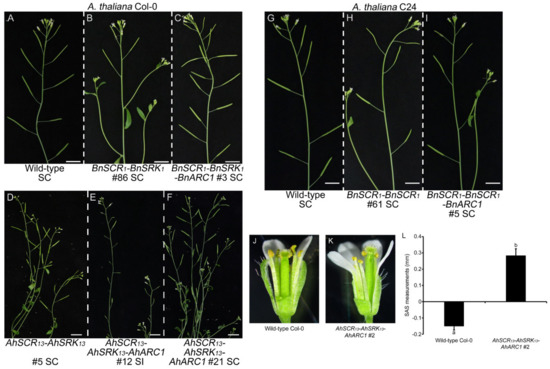
Figure 3.
Branches with siliques from transgenic A. thaliana Col-0 and C24 plants following self-pollination and the approach herkogamy phenotype in AhSCR13-AhSRK13-AhARC1 line 2. (A,G) Wild-type Col-0 and C24 branches with well-developed siliques. (B–F) Branches with siliques from transgenic A. thaliana Col-0 lines: BnSCR1-BnSRK1 line 86, BnSCR1-BnSRK1-BnARC1 line 3, AhSCR13-AhSRK13 line 5, AhSCR13-AhSRK13-AhARC1 lines 12 and 21. (H,I) Branches with siliques from transgenic A. thaliana C24 lines: BnSCR1-BnSRK1 line 61 and BnSCR1-BnSRK1-BnARC1 line 5. SC: self-compatible; SI: self-incompatible. Bars = 1 cm. (J) Wild-type Col-0 and (K) AhSCR13-AhSRK13-AhARC1 line-2 flowers. Petals and sepals have been removed to allow a clearer view of the anther positions relative to the stigma. (L) Stigma-anther separation (SAS) measurements. Col-0 flowers typically have a reverse herkogamy phenotype, which is represented by a negative SAS value. In contrast, the positive SAS value for AhSCR13-AhSRK13-AhARC1 line-2 flowers indicates an approach herkogamy phenotype. SAS measurements were taken as described in Luo and Widmer 2013 [48] and Indriolo et al. 2014 [43]. n = 10 flowers.
Since self-incompatibility has been defined as a reproductive mechanism that prevents self-fertilization, seed production of representative transgenic Col-0 and C24 lines was examined. The plants were allowed to self-fertilize naturally, and photographs of branches with siliques from plants with each combination of transgenes are shown in Figure 3. The wild-type A. thaliana Col-0 and C24 plants are self-compatible and should produce well-developed siliques full of seeds with self-pollination (Figure 3A,G). The transgenic Col-0 plants with AhSCR13-AhSRK13, AhSCR13-AhSRK13-AhARC1, BnSCR1-BnSRK1 or BnSCR1-BnSRK1-BnARC1 that did not display self-incompatibility produced siliques with similar sizes to the wild-type Col-0 plants (Figure 3B–D,F). Regular-sized siliques were also observed in the self-compatible transgenic C24 plants carrying the BnSCR1-BnSRK1 or BnSCR1-BnSRK1-BnARC1 transgenes (Figure 3H,I). However, the strongly self-incompatible transgenic Col-0 plants with the AhSCR13-AhSRK13-AhARC1 transgenes displayed much smaller silique sizes, as a result of very reduced seed production with self-pollination (Figure 3E).
Fully developed siliques from each of these lines were harvested and dissected to record the number of seeds per silique (Figure 4). Siliques from wild-type A. thaliana Col-0 plants produced an average of 44.7 seeds/silique via self-pollination (Figure 4A). Similarly, the siliques from the three self-compatible AhSCR13-AhSRK13 transgenic Col-0 plants contained 40.4, 42.5 and 46.1 seeds/silique on average, respectively, showing no significant difference with wild-type Col-0 (Figure 4A). The siliques of the three self-compatible BnSCR1-BnSRK1 and BnSCR1-BnSRK1-BnARC1 transgenic Col-0 plants generated similar averages of 48.5, 47, 47.3, and 47.2, 47.3, 48.5 seeds/silique, respectively (Figure 4B). Thus, these self-compatible transgenic Col-0 lines displayed an equivalent ability to accept self-pollen compared to the wild-type Col-0 plants. The addition of BnARC1 with BnSCR1-BnSRK1 did not promote the self-pollen rejection in A. thaliana Col-0. In contrast, some reduction in the number of seeds/silique was observed for the three moderately self-incompatible AhSCR13-AhSRK13-AhARC1 transgenic Col-0 lines, with averages of 28.6, 23.8, and 26.5 seeds/silique, respectively (Figure 4A). In addition, significant reduction in seed production was observed for the three strong self-incompatible AhSCR13-AhSRK13-AhARC1 transgenic Col-0 lines following self-pollination, with the lowest value of 1.5 seeds/silique scored for AhSCR13-AhSRK13-AhARC1 line 2 (Figure 4A). Therefore, the expression of AhARC1 along with AhSCR13-AhSRK13 in A. thaliana Col-0 induced a strong self-incompatibility response resulting in very reduced seed production.
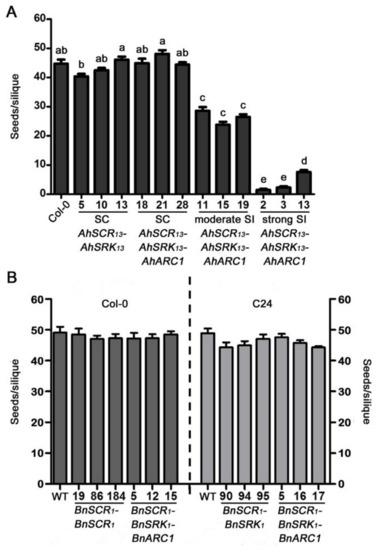
Figure 4.
Seed set of transgenic A. thaliana Col-0 and C24 plants following self-pollination. (A) Mean seeds/silique of self-pollinated wild-type A. thaliana Col-0 and transgenic Col-0 AhSCR13-AhSRK13 and AhSCR13-AhSRK13-AhARC1 lines. Means with significant differences at p < 0.05 are shown with different letters (one-way ANOVA with Tukey-HSD post-hoc tests). n = 10 siliques. (B) Mean seeds/silique of self-pollinated wild-type A. thaliana Col-0 and C24 plants, as well as transgenic Col-0 and C24 BnSCR1-BnSRK1 and BnSCR1-BnSRK1-BnARC1 lines. Significant difference at p < 0.05 calculated by one-way ANOVA with Tukey-HSD post-hoc tests. n = 10 siliques. Error bars indicate SE. SC: self-compatible; SI: self-incompatible.
Mature siliques of natural self-pollination transgenic A. thaliana C24 lines were also examined for seed set and compared to wild-type A. thaliana C24 plants, which produced an average of 48.9 seeds/silique (Figure 4B). All the self-compatible BnSCR1-BnSRK1 and BnSCR1-BnSRK1-BnARC1 transgenic C24 lines showed similarly high numbers of seeds/silique following self-pollination, which ranged from 44.3 to 47.5 seeds/silique (Figure 4B). Accordingly, the B. napus SCR1-SRK1 transgenes in the A. thaliana C24 ecotype could not elicit a self-incompatibility response, and the addition of BnARC1 with BnSCR1-BnSRK1 did not result in any reduction of seed set in the C24 ecotype.
2.4. The Expression Levels of the SCR-SRK-ARC1 Transgenes are Not Associated with the Intensity of Self-Incompatibility Response in A. thaliana
Given the differences observed in the ability of the AhSCR13-AhSRK13-AhARC1 and BnSCR1-BnSRK1-BnARC1 transgenes to confer self-incompatibility in A. thaliana, the transgene expression levels were investigated to reveal if they correlated with the observed phenotypes. Mature buds from representative transgenic lines were harvested to test the relative expression levels of each transgene using quantitative RT-PCR (qRT-PCR). Overall, the transgene expression levels were quite variable, and there was no correlation between a particular transgene expression level and the self-incompatibility trait (Figure 5). For example, the strongly self-incompatible AhSCR13-AhSRK13-AhARC1 transgenic Col-0 plants tended to show lower levels of relative expression for all three transgenes (Figure 5A–C). In fact, the highest relative expression level for each transgene was detected in the fully self-compatible AhSCR13-AhSRK13-AhARC1 transgenic lines: line 18 (AhSRK13), line 21 (AhSCR13) and line 28 (AhARC1) (Figure 5A–C). Thus, the expression level of these transgenes did not account for the intensity of self-incompatibility in transgenic A. thaliana Col-0 lines.
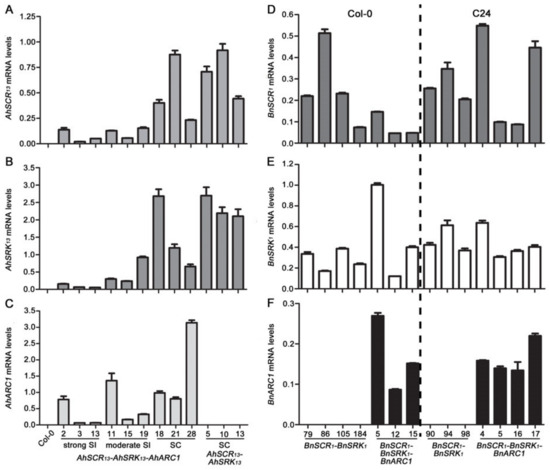
Figure 5.
Relative expression levels of AhSCR13, AhSRK13, AhARC1, BnSCR1, BnSRK1, and BnARC1 in transgenic A. thaliana Col-0 and C24 plants. (A) Relative mRNA levels of AhSCR13 in mature buds from different lines. (B) Relative mRNA levels of AhSRK13 in mature buds from different lines. (C) Relative mRNA levels of AhARC1 in mature buds from different lines. (D) Relative mRNA levels of BnSCR1 in mature buds from different lines. (E) Relative mRNA levels of BnSRK1 in mature buds from different lines. (F) Relative mRNA levels of BnARC1 in mature buds from different lines. The relative expression levels of all the genes were normalized to the expression of Actin2 and TUB4. The wild-type Col-0 bud samples were used as a negative control for AhSCR13, AhSRK13, and AhARC1 genes. Primers for qRT-PCR were designed to span introns where possible. Means of two biological replicates (three technical replicates each) are shown. Error bars indicate SE. SC: self-compatible; SI: self-incompatible.
Although the addition of the BnSCR1-BnSRK1 and BnSCR1-BnSRK1-BnARC1 transgenes did not restore the self-incompatibility in the A. thaliana Col-0 and C24 ecotypes, the expression levels of these transgenes were also quantified to confirm that they were successfully being expressed in the transgenic lines (Figure 5D–F). The BnSCR1 transgene tended to show quite high relative expression levels, particularly in the C24 ecotype (Figure 5D), and the relative levels in both Col-0 and C24 showed similar ranges to that observed for AhSCR13 (Figure 5A). Conversely, the BnSRK1 relative expression levels were generally lower than those observed for several of the transgenic lines carrying the AhSRK13 transgene (Figure 5B,E). However, the BnSCR1-BnSRK1-BnARC1 transgenic lines did have BnSRK1 expression levels that were comparable to the AhSRK13 transgene in the strongly self-incompatible AhSCR13-AhSRK13-AhARC1 transgenic lines (Figure 5B,E). Finally, the BnARC1 expression levels were generally higher than the observed relative expression values for the AhARC1 (Figure 5C,F). Thus, no trend was seen in the various expression levels for the BnSCR1-BnSRK1-BnARC1 transgenes that could account for lack of self-pollen rejection in both the A. thaliana Col-0 and C24 transgenic plants. This suggests that the B. napus self-incompatibility signaling pathway cannot be reconstructed in the A. thaliana Col-0 and C24 ecotypes.
3. Discussion
The transition to selfing has made the model plant A. thaliana an ideal system for the reconstruction of self-incompatibility. It is believed that the loss-of-function mutations in SRK and SCR genes has led to self-fertility in all A. thaliana ecotypes [28,30,31,32,35,49,50,51]. Self-incompatibility has been restored in some A. thaliana ecotypes by introducing functional A. lyrata SCR and SRK genes, while several other ecotypes have remained self-fertile [31,33,35,40,43]. Notably, transgenic A. thaliana expressing A. lyrata SCRb-SRKb in the C24 ecotype were self-incompatible and produced very few seeds, while the same combination in the Col-0 ecotype resulted in full seed production with selfing [33,35,40]. On the other hand, the addition of ARC1 with A. lyrata SCRb-SRKb in the Col-0 ecotype resulted in self-incompatible plants with reduced seed set [43].
In this study, we tested the reconstitution of the SCR-SRK-ARC1 signaling pathway in self-compatible A. thaliana using new sets of self-incompatibility genes from two other species, self-incompatible B. napus and A. halleri. Our results showed that self-pollen rejection was achieved only in the A. thaliana AhSCR13-AhSRK13-AhARC1 Col-0 lines. The other combination of the AhSCR13-AhSRK13 transgenes was not sufficient to restore self-incompatibility in A. thaliana Col-0 ecotypes, similar to that previously observed [43]. Interestingly, only flowers from the AhSCR13-AhSRK13-AhARC1 strongly self-incompatible Col-0 plants displayed an approach herkogamous phenotype, which had been previously detected with the SCRb-SRKb-ARC1 transgene combination [43]. The self-compatible A. thaliana Col-0 AhSCR13-AhSRK13-AhARC1 plants maintained the same floral phenotype as wild-type Col-0 plants, indicating that the approach herkogamous phenotype is associated with the intensity of self-incompatibility of the transgenic plants. The expression of SRKb was previously found to enhance pistil elongation and stigma exertion in the rdr6 mutant background [52]. Thus, the self-incompatibility signaling transduction mediated by AhSCR13-AhSRK13-AhARC1 might also activate downstream components that are regulating pistil development.
As nearly half of the A. thaliana Col-0 AhSCR13-AhSRK13-AhARC1 plants still exhibited a self-compatible phenotype, quantitative RT-PCR was conducted to test if the expression levels of the transgenes were correlated with the ability of self-pollen rejection. However, there was no clear relationship between the relative expression levels of the three transgenes and the intensity of self-incompatibility response. Thus, the trigger for self-pollen rejection in the transgenic A. thaliana Col-0 AhSCR13-AhSRK13-AhARC1 lines did not appear to be dependent on the expression levels of the transgenes. Unexpectedly, no self-incompatible lines were generated in both the A. thaliana Col-0 and C24 ecotypes with the transformation of BnSCR1-BnSRK1 or BnSCR1-BnSRK1-BnARC1 genes. These B. napus transgenes displayed similar expression patterns to the A. halleri transgenes, and so their relative expression levels did not appear to account for the lack of a self-incompatibility phenotype. A potential reason may be the design of these transgenes, where only coding sequences were cloned and driven by promoters from other genes, potentially impacting their expression. As the BnARC1 transgene was previously shown to be functional in the A. thaliana Col-0 background [43], it is more likely that BnSCR1 or BnSRK1 are not functional in A. thaliana. Similar, the BnSCR1 construct was found to be functional in B. napus [46], and so BnSRK1 would be the only previously untested construct. One other possible reason is that Arabidopsis and Brassica species may not completely share the same SRK-mediated self-incompatibility signaling pathway. These differences may potentially be due to B. napus SRK1 activity, as B. napus ARC1 was previously shown to interact with A. lyrata SRK, and the A. lyrata SCRb-SRKb and B. napus ARC1 transgenes produced self-incompatible Col-0 transgenic plants [43,53]. Ideally, the potential activity of the B. napus transgenes could be tested by reciprocal pollinations between the A. thaliana Col-0 BnSCR1-BnSRK1-BnARC1 lines and the B. napus S1 haplotype transgenic line [46]. However, large physical differences in pollen and pistil sizes between these species (Supplementary Figure S2, [38]) as well as potential complications from interspecies crosses may obscure any self-incompatibility reactions.
There are several pieces of evidence that would support the idea of differences in the Brassica and Arabidopsis SRK-mediated signaling pathways. A. thaliana diverged from A. lyrata about 10 MYA and diverged from Brassica approximately 24 MYA [54,55,56,57,58]. As would be expected within the Arabidopsis genus, the inactivated SCR and SRK alleles in A. thaliana share high sequence similarity with the functional S-haplotype genes in A. lyrata and A. halleri [28,30,31,34,35,50,59,60]. In contrast, there is high variation in their sequences when compared to Brassica SP11/SCR and SRK alleles, and so this variation could lead to differences in SRK interactions with other downstream components (the ARC1 interaction is conserved). The ability of the SCRb-SRKb transgenes to restore self-incompatibility in the C24 ecotype (i.e., no ARC1) supports the idea that C24 plants might employ other unknown self-incompatibility signaling players; for example, a strong calcium flux mediated by glutamate receptor-like channels has been shown to occur in self-incompatible C24 stigmas [33,35,40]. In addition, while MLPK is required for self-incompatibility both in B. rapa and B. napus [17,18], an A. thaliana homolog of MLPK (APK1b) was found to be unfunctional in regulating the transient self-incompatibility response observed in SCRb-SRKb Col-0 lines [42]. Finally, there are differences in the cellular responses that take place in the stigmatic papillae of Brassica and Arabidopsis species, such as autophagy and the trafficking of vesicles and multivesicular bodies [16,61]. The potential redundancy shaped by genome triplication and the formation of distinct subgenomes in Brassica species may have increased robustness in the self-incompatibility system, allowing further evolution of the underlying signaling pathways and distinction from Arabidopsis species [62,63].
In conclusion, we have demonstrated that the previously untested A. halleri S13-haplotype genes, along with AhARC1, can restore the self-incompatibility phenotype in A. thaliana Col-0 plants, and that AhARC1 is required along with AhSCR13-AhSRK13 for self-pollen rejection in the Col-0 ecotype. The generation of these self-incompatible A. thaliana AhSCR13-AhSRK13-AhARC1 Col-0 plants will be a very useful resource for further studies on sporophytic self-incompatibility in A. thaliana. Our results also show that the equivalent set of self-incompatibility genes from Brassica (BnSRK1, BnSCR1 and BnARC1) was not sufficient to establish self-incompatibility in both the A. thaliana Col-0 and C24 ecotypes. These observations indicate that the transfer of self-incompatibility into A. thaliana is based on the phylogenetic relationships with the transgene donor, and it would be good to verify this by testing other combinations of SP11/SCR and SRK alleles from different Brassicaceae species. Since the Brassica-specific MLPK gene has been proven to be necessary for self-incompatibility in Brassica species [17,18], it would be worth transforming B. napus SCR-SRK-MLPK-ARC1 all together into A. thaliana to determine if MLPK is the missing component for reconstructing the Brassica self-incompatibility pathway. As well, the reciprocal experiment of transferring Arabidopsis SCR and SRK genes into self-compatible B. napus would be quite interesting in order to see if the Arabidopsis SCR and SRK genes can restore self-incompatibility in a Brassica species, or if the diversification of the self-incompatibility pathway extends in both directions.
4. Materials and Methods
4.1. Plant Materials
The wild-type A. thaliana Col-0 and C24 plants and the transgenic lines were grown under long-day conditions in a greenhouse with a 16-h light/8-h dark photoperiod at 22 °C. The transgenic T1 plants were selected from seeds collected following floral dipping by being plated on 1/2 MS medium containing kanamycin.
4.2. Vector Construction and Plant Transformation
The functional promoter and CDS of BrSP11-47 (the CDS sequence is the same as BnSCR1) was amplified from the pCAMBIA2301-1+4 vector [46]. And the Nopaline synthase polyadenylation signal (NosT) was amplified from the pCAMBIA2301 vector used as the terminator of BnSCR1 [64]. These two fragments were subcloned into the Bsp120I and Psp1406I sites of the pORE_O4 binary vector, respectively [65]. The SLR1 promoter is expressed in particular in stigma of Arabidopsis, and was amplified from B. napus ‘Westar’ genome DNA using forward and reverse primers with KpnI and XbaI sites [66,67]. The full-length CDS of BnSRK1 was cloned from ‘Westar’ cDNA with primers carrying XbaI and BstEII sites. The SLR1 promoter and BnSRK1 CDS were joined using the XbaI site by being cloned into the pCAMBIA2301 vector, then the SLR1pro-BnSRK1 fragment was amplified from the medium vector with primers with Cfr9I and NotI sites for directional cloning into the pORE_O4 vector. A 989 bp-terminator of BnSRK1 was obtained from ‘Westar’ genome DNA and was cloned into the pORE_O4 vector at the NotI site. For the reconstitution of the BnSCR1-BnSRK1-BnARC1 construct, the full-length CDS of BnARC1 was amplified from ‘Westar’ cDNA with primers with SalI and KpnI restriction sites. BnARC1 was also driven by the SLR1 promoter, which was amplified using primers with HindIII and SalI sites that were then cloned into the pCAMBIA2301 vector to be joined. The SLR1pro-BnARC1 fragment was further cloned by primers carrying the KpnI site and ligated into the pORE_O4 vector.
All promoters and genes from the A. halleri S13 haplotype [47] and AhARC1 together with its promoter were synthesized by Invitrogen GeneArt (Thermo Fisher Scientific, Waltham, MA, USA). It should be noted that AhSRK13 was driven by AhSRK20 promoter, since AhSRK13 and AhSRK20 were codominant in A. halleri [41]. AhSRK20 promoter, AhARC1pro-AhARC1, AhSCR13pro-AhSCR13 and AhSRK13 were ligated into the pORE_O4 vector using the Cfr9I-NotI, XbaI, Bsp120I and NotI-Acc65I sites or pairs of sites in the order they were mentioned (see Supplementary File S1 for further details and sequences). AhARC1 was not cloned into the pORE_O4 vector for the combination of AhSCR13-AhSRK13 transgenes. All PCR products were amplified with PrimeSTAR HS DNA polymerase (Takara, Kusatsu, Shiga, Japan), and the PCR products were then ligated to pGEM-T Easy (Promega, Madison, WI, USA) and sequenced. These constructs were introduced into Agrobacterium tumefaciens GV3101 host cells. PCR primers used in vector construction steps are listed in Supplementary Table S1. The plant transformation was performed following the floral-dip method [49,68].
4.3. Pollination Assays
All the floral buds of the A. thaliana Col-0 and C24 transgenic plants were emasculated and covered with a paper bag one day before anthesis to avoid pollen contamination. Pollinations were performed the next day (day of anthesis). The pistil was cut at the peduncle 4 h after pollination, then fixed for 2 h in ethanol–acetic acid (3:1), softened in 1 N NaOH at 60 °C for 1 h and stained with 0.01% (w/v) decolorized aniline blue for 2.5 h in 2% (w/v) K3PO4. Pistils were gently squashed onto a microscopic slide glass by placing the cover glass over the pistils. Samples were examined under a fluorescence microscope (Eclipse 80i, Nikon, Minato, Tokyo, Japan).
4.4. Quantitative RT-PCR Assays
Total RNAs were extracted using the SV Total RNA Isolation System (Promega Madison, WI, USA). The RNA samples were quantified using a NanoDrop Spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA), and 1 µg RNA of each sample was used to synthesize the first-strand cDNA with a Thermo RT kit (Thermo Fisher, Waltham, MA, USA). The cDNA samples were used as templates of the qRT-PCR assays. The qRT-PCRs on the transgenic lines were performed using cDNA of the mature buds and 2× Power SYBR green (Toyobo, Osaka, Osaka Prefecture, Japan). qRT-PCRs were then conducted on the CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA), and the conditions used were a pre-denaturation at 95 °C for 3 min, followed by a three-step cycle of 10 s denaturation at 95 °C, 10 s annealing at 65 °C and a 30 s extension at 72 °C for 47 cycles with a melt curve. The results were calculated with CFX Manager Software (Bio-Rad Hercules, CA, USA). Primers used in qRT-PCR analysis are shown in Supplementary Table S2.
4.5. Accession Numbers
BnSCR1 gene (Genbank accession AB270773), BnSRK1 gene (Genbank accession AB270771), BnARC1 gene (Genbank accession AF024625). Sequence data and accession numbers of B. napus can be found in http://www.genoscope.cns.fr/brassicanapus/. Sequence and accession numbers information for A. thaliana and A. halleri genes used in this study were obtained from TAIR and Phytozome.
Supplementary Materials
The following are available online at https://www.mdpi.com/2223-7747/8/12/570/s1. Figure S1. Pollen tube germination and growth on stigmas of wild-type A. thaliana Col-0 and transgenic Col-0 plants following reciprocal crossings. (A,B) Wild-type A. thaliana Col-0 stigma pollinated with pollen from transgenic strong self-incompatible Col-0 AhSCR13-AhSRK13-AhARC1 line 2. (C,D) Transgenic strong self-incompatible Col-0 AhSCR13-AhSRK13-AhARC1 line 2 stigma pollinated with pollen from wild-type A. thaliana Col-0 plant. Differential interference contrast (DIC) and aniline blue-stained images are shown for each sample. SC: self-compatible; SI: self-incompatible. Bars = 100 μm. Figure S2. Flowers from wild-type B. napus ‘Westar’, A. thaliana Col-0 and C24, and transgenic A. thaliana Col-0 and C24 plants. (A,B,H) Flowers of wild-type B. napus ‘Westar’, A. thaliana Col-0 and C24 plants. (C–G) Flowers of transgenic A. thaliana Col-0 lines: BnSCR1-BnSRK1 line 12, BnSCR1-BnSRK1-BnARC1 line 9, AhSCR13-AhSRK13 line 5, and AhSCR13-AhSRK13-AhARC1 lines 2 and 21. (I,J) flowers of transgenic A. thaliana C24 lines: BnSCR1-BnSRK1 line 90, and BnSCR1-BnSRK1-BnARC1 line 16. SC: self-compatible; SI: self-incompatible. Bar = 1 mm in (A) and bars = 200 μm in (B–J). Table S1. Primers used in vector construction. Table S2. Primers used in qRT-PCR assay. Supplementary File S1. Map and sequences for AhSCR13-AhSRK13-AhARC1.
Author Contributions
T.Z. and G.Z. designed and performed the research, analyzed data and wrote the manuscript. D.R.G. designed the AhSCR13-AhSRK13-AhARC1 experiment and edited the manuscript. X.L. took part in investigation. S.M. participated in investigation and formal analysis. C.D. guided the vector construction of BnSCR1-BnSRK1 and BnSCR1-BnSRK1-BnARC1 genes. J.W., B.Y., J.S., J.T. and T.F. supervised the research. C.M. supervised the research, acquired funding and wrote the original draft.
Funding
This research was funded by grants from National Science Foundation of China (31571706) and Outstanding Agricultural Scientific Research Talents and Innovative Teams to C.M. T.Z.’s research in the Goring lab was supported by a scholarship from the China Scholarship Council, and a grant from Natural Sciences and Engineering Research Council of Canada to D.R.G.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Franklin-Tong, V.E.; Holdaway-Clarke, T.L.; Straatman, K.R.; Kunkel, J.G.; Hepler, P.K. Involvement of extracellular calcium influx in the self-incompatibility response of Papaver rhoeas. Plant J. 2002, 29, 333–345. [Google Scholar] [CrossRef]
- Franklin-Tong, N.V.; Franklin, F.C.H. Gametophytic self-incompatibility inhibits pollen tube growth using different mechanisms. Trends Plant Sci. 2003, 8, 598–605. [Google Scholar] [CrossRef]
- Kao, T.H.; Tsukamoto, T. The molecular and genetic bases of S-RNase-based self-incompatibility. Plant Cell 2004, 16, S72–S83. [Google Scholar] [CrossRef]
- Takayama, S.; Isogai, A. Self-incompatibility in plants. Annu. Rev. Plant Biol. 2005, 56, 467–489. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, J.B.; Nasrallah, M.E. Pollen-stigma signaling in the sporophytic self-incompatibility response. Plant Cell 1993, 5, 1325. [Google Scholar] [CrossRef] [PubMed]
- Luu, D.T.; Marty-Mazars, D.; Trick, M.; Dumas, C.; Heizmann, P. Pollen-stigma adhesion in Brassica spp involves SLG and SLR1 glycoproteins. Plant Cell 1999, 11, 251–262. [Google Scholar] [PubMed]
- Giranton, J.L.; Dumas, C.; Cock, J.M.; Gaude, T. The integral membrane S-locus receptor kinase of Brassica has serine/threonine kinase activity in a membranous environment and spontaneously forms oligomers in planta. Proc. Natl. Acad. Sci. USA 2000, 97, 3759–3764. [Google Scholar] [CrossRef]
- Takasaki, T.; Hatakeyama, K.; Suzuki, G.; Watanabe, M.; Isogai, A.; Hinata, K. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature 2000, 403, 913–916. [Google Scholar] [CrossRef]
- Silva, N.F.; Stone, S.L.; Christie, L.N.; Sulaman, W.; Nazarian, K.A.; Burnett, L.A.; Arnoldo, M.A.; Rothstein, S.J.; Goring, D.R. Expression of the S receptor kinase in self-compatible Brassica napus cv. Westar leads to the allele-specific rejection of self-incompatible Brassica napus pollen. Mol. Genet. Genom. 2001, 265, 552–559. [Google Scholar]
- Schopfer, C.R.; Nasrallah, M.E.; Nasrallah, J.B. The male determinant of self-incompatibility in Brassica. Science 1999, 286, 1697–1700. [Google Scholar] [CrossRef]
- Suzuki, G.; Kai, N.; Hirose, T.; Fukui, K.; Nishio, T.; Takayama, S.; Isogai, A.; Watanabe, M.; Hinata, K. Genomic organization of the S locus: Identification and characterization of genes in SLG/SRK region of S9 haplotype of Brassica campestris (syn Rapa). Genetics 1999, 153, 391–400. [Google Scholar]
- Shiba, H.; Iwano, M.; Entani, T.; Ishimoto, K.; Shimosato, H.; Che, F.S.; Satta, Y.; Ito, A.; Takada, Y.; Watanabe, M.; et al. The dominance of alleles controlling self-incompatibility in Brassica pollen is regulated at the RNA level. Plant Cell 2002, 14, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.; Schopfer, C.R.; Nasrallah, M.E.; Nasrallah, J.B. Allele-specific receptor-ligand interactions in Brassica self-incompatibility. Science 2001, 293, 1824–1826. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Shimosato, H.; Shiba, H.; Funato, M.; Che, F.S.; Watanabe, M.; Iwano, M.; Isogai, A. Direct ligand-receptor complex interaction controls Brassica self-incompatibility. Nature 2001, 413, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Doucet, J.; Lee, H.K.; Goring, D.R. Pollen acceptance or rejection: A tale of two pathways. Trends Plant Sci. 2016, 21, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Goring, D.R. Exocyst, exosomes, and autophagy in the regulation of Brassicaceae pollen-stigma interactions. J. Exp. Bot. 2017, 69, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Murase, K.; Shiba, H.; Iwano, M.; Che, F.S.; Watanabe, M.; Isogai, A.; Takayama, S. A membrane-anchored protein kinase involved in Brassica self-incompatibility signaling. Science 2004, 303, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yang, Y.; Li, B.; Liu, Z.; Khan, F.; Zhang, T.; Zhou, G.; Tu, J.; Shen, J.; Yi, B.; et al. Functional analysis of M-locus protein kinase revealed a novel regulatory mechanism of self-incompatibility in Brassica napus L. Int. J. Mol. Sci. 2019, 20, 3303. [Google Scholar] [CrossRef]
- Kakita, M.; Murase, K.; Iwano, M.; Matsumoto, T.; Watanabe, M.; Shiba, H.; Isogai, A.; Takayama, S. Two distinct forms of M-locus protein kinase localize to the plasma membrane and interact directly with S-locus receptor kinase to transduce self-incompatibility signaling in Brassica rapa. Plant Cell 2007, 19, 3961–3973. [Google Scholar] [CrossRef]
- Kakita, M.; Shimosato, H.; Murase, K.; Isogai, A.; Takayama, S. Direct interaction between S-locus receptor kinase and M-locus protein kinase involved in Brassica self-incompatibility signaling. Plant Biotechnol. 2007, 24, 185–190. [Google Scholar] [CrossRef]
- Bower, M.S.; Matias, D.D.; Fernandes-Carvalho, E.; Mazzurco, M.; Gu, T.; Rothstein, S.J.; Goring, D.R. Two members of the thioridoxin-h family interact with the kinase domain of a Brassica S locus receptor kinase. Plant Cell 1996, 8, 1641–1650. [Google Scholar] [PubMed]
- Gu, T.; Mazzurco, M.; Sulaman, W.; Matias, D.D.; Goring, D.R. Binding of an arm repeat protein to the kinase domain of the S-locus receptor kinase. Proc. Natl. Acad. Sci. USA 1998, 95, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Cabrillac, D.; Cock, J.M.; Dumas, C.; Gaude, T. The S-locus receptor kinase is inhibited by thioredoxins and activated by pollen coat proteins. Nature 2001, 410, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Arnoldo, M.; Goring, D.R. A breakdown of Brassica self-incompatibility in ARC1 antisense transgenic plants. Science 1999, 286, 1729–1731. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Anderson, E.M.; Mullen, R.T.; Goring, D.R. ARC1 is an E3 ubiquitin ligase and promotes the ubiquitination of proteins during the rejection of self-incompatible Brassica pollen. Plant Cell 2003, 15, 885–898. [Google Scholar] [CrossRef]
- Samuel, M.A.; Chong, Y.T.; Haasen, K.E.; Aldea-Brydges, M.G.; Stone, S.L.; Goring, D.R. Cellular pathways regulating responses to compatible and self-incompatible pollen in Brassica and Arabidopsis stigmas intersect at Exo70A1,a putative component of the exocyst complex. Plant Cell 2009, 21, 2655–2671. [Google Scholar] [CrossRef]
- Sankaranarayanan, S.; Jamshed, M.; Samuel, M.A. Degradation of glyoxalase I in Brassica napus stigma leads to self-incompatibility response. Nat. Plants 2015, 1, 15185. [Google Scholar] [CrossRef]
- Kusaba, M.; Dwyer, K.; Hendershot, J.; Vrebalov, J.; Nasrallah, J.B.; Nasrallah, M.E. Self-incompatibility in the genus Arabidopsis: Characterization of the S locus in the outcrossing A. lyrata and its autogamous relative A. thaliana. Plant Cell 2001, 13, 627–643. [Google Scholar] [CrossRef]
- Schierup, M.H.; Mable, B.K.; Awadalla, P.; Charlesworth, D. Identification and characterization of a polymorphic receptor kinase gene linked to the self-incompatibility locus of Arabidopsis lyrata. Genetics 2001, 158, 387–399. [Google Scholar]
- Tang, C.; Toomajian, C.; Sherman-Broyles, S.; Plagnol, V.; Guo, Y.L.; Hu, T.T.; Clark, R.M.; Nasrallah, J.B.; Weigel, D.; Nordborg, M. The evolution of selfing in Arabidopsis thaliana. Science 2007, 317, 1070–1072. [Google Scholar] [CrossRef]
- Tsuchimatsu, T.; Suwabe, K.; Shimizu-Inatsugi, R.; Isokawa, S.; Pavlidis, P.; Städler, T.; Suzuki, G.; Takayama, S.; Watanabe, M.; Shimizu, K.K. Evolution of self-compatibility in Arabidopsis by a mutation in the male specificity gene. Nature 2010, 464, 1342–1346. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.L.; Zhao, X.; Lanz, C.; Weigel, D. Evolution of the S-locus region in Arabidopsis relatives. Plant Physiol. 2011, 157, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, M.E.; Liu, P.; Sherman-Broyles, S.; Boggs, N.A.; Nasrallah, J.B. Natural variation in expression of self-incompatibility in Arabidopsis thaliana: Implications for the evolution of selfing. Proc. Natl. Acad. Sci. USA 2004, 101, 16070–16074. [Google Scholar] [CrossRef] [PubMed]
- Sherman-Broyles, S.; Boggs, N.; Farkas, A.; Liu, P.; Vrebalov, J.; Nasrallah, M.E.; Nasrallah, J.B. S locus genes and the evolution of self-fertility in Arabidopsis thaliana. Plant Cell 2007, 19, 94–106. [Google Scholar] [CrossRef]
- Boggs, N.A.; Nasrallah, J.B.; Nasrallah, M.E. Independent S-locus mutations caused self-fertility in Arabidopsis thaliana. PLoS Genet. 2009, 5, e1000426. [Google Scholar] [CrossRef]
- Boggs, N.A.; Dwyer, K.G.; Shah, P.; McCulloch, A.A.; Bechsgaard, J.; Schierup, M.H.; Nasrallah, M.E.; Nasrallah, J.B. Expression of distinct self-incompatibility specificities in Arabidopsis thaliana. Genetics 2009, 182, 1313–1321. [Google Scholar] [CrossRef]
- Rea, A.C.; Liu, P.; Nasrallah, J.B. A transgenic self-incompatible Arabidopsis thaliana model for evolutionary and mechanistic studies of crucifer self-incompatibility. J. Exp. Bot. 2010, 61, 1897–1906. [Google Scholar] [CrossRef]
- Bi, Y.M.; Brugiere, N.; Cui, Y.; Goring, D.; Rothstein, S. Transformation of Arabidopsis with a Brassica SLG/SRK region and ARC1 gene is not sufficient to transfer the self-incompatibility phenotype. Mol. General Genet. MGG 2000, 263, 648–654. [Google Scholar] [CrossRef]
- Nasrallah, M.E.; Liu, P.; Nasrallah, J.B. Generation of self-incompatible Arabidopsis thaliana by transfer of two S locus genes from A. lyrata. Science 2002, 297, 247–249. [Google Scholar] [CrossRef]
- Iwano, M.; Ito, K.; Fujii, S.; Kakita, M.; Asano-Shimosato, H.; Igarashi, M.; Kaothien-Nakayama, P.; Entani, T.; Kanatani, A.; Takehisa, M.; et al. Calcium signalling mediates self-incompatibility response in the Brassicaceae. Nat Plants 2015, 1, 15128. [Google Scholar] [CrossRef]
- Durand, E.; Meheust, R.; Soucaze, M.; Goubet, P.M.; Gallina, S.; Poux, C.; Fobis-Loisy, I.; Guillon, E.; Gaude, T.; Sarazin, A.; et al. Dominance hierarchy arising from the evolution of a complex small RNA regulatory network. Science 2014, 346, 1200–1205. [Google Scholar] [CrossRef] [PubMed]
- Kitashiba, H.; Liu, P.; Nishio, T.; Nasrallah, J.B.; Nasrallah, M.E. Functional test of Brassica self-incompatibility modifiers in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2011, 108, 18173–18178. [Google Scholar] [CrossRef] [PubMed]
- Indriolo, E.; Safavian, D.; Goring, D.R. The ARC1 E3 ligase promotes two different self-pollen avoidance traits in Arabidopsis. Plant Cell 2014, 26, 1525–1543. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, S.; Odashima, M.; Fujimoto, R.; Sato, Y.; Kitashiba, H.; Nishio, T. Self-compatibility in Brassica napus is caused by independent mutations in S-locus genes. Plant J. 2007, 50, 391–400. [Google Scholar] [CrossRef]
- Tochigi, T.; Udagawa, H.; Li, F.; Kitashiba, H.; Nishio, T. The self-compatibility mechanism in Brassica napus L. is applicable to F1 hybrid breeding. Theor. Appl. Genet. 2011, 123, 475–482. [Google Scholar] [CrossRef]
- Gao, C.B.; Zhou, G.L.; Ma, C.Z.; Zhai, W.; Zhang, T.; Liu, Z.; Yang, Y.; Wu, M.; Yue, Y.; Duan, Z.; et al. Helitron-like transposons contributed to the mating system transition from out-crossing to self-fertilizing in polyploid Brassica napus L. Sci. Rep. 2016, 6, 33785. [Google Scholar] [CrossRef]
- Goubet, P.M.; Berges, H.; Bellec, A.; Prat, E.; Helmstetter, N.; Mangenot, S.; Gallina, S.; Holl, A.C.; Fobis-Loisy, I.; Vekemans, X.; et al. Contrasted patterns of molecular evolution in dominant and recessive self-incompatibility haplotypes in Arabidopsis. PLoS Genet. 2012, 8, e1002495. [Google Scholar] [CrossRef]
- Luo, Y.; Widmer, A. Herkogamy and its effects on mating patterns in Arabidopsis thaliana. PLoS ONE 2013, 8, e57902. [Google Scholar] [CrossRef]
- Xia, S.Q.; Wang, Z.X.; Zhang, H.Y.; Hu, K.; Zhang, Z.; Qin, M.; Dun, X.; Yi, B.; Wen, J.; Ma, C.; et al. Altered transcription and neofunctionalization of duplicated genes rescue the harmful effects of a chimeric gene in Brassica napus. Plant Cell 2016, 28, 2060–2078. [Google Scholar] [CrossRef]
- Shimizu, K.K.; Shimizu-Inatsugi, R.; Tsuchimatsu, T.; Purugganan, M.D. Independent origins of self-compatibility in Arabidopsis thaliana. Mol. Ecol. 2008, 17, 704–714. [Google Scholar] [CrossRef]
- Tsuchimatsu, T.; Goubet, P.M.; Gallina, S.; Holl, A.C.; Fobis-Loisy, I.; Bergès, H.; Marande, W.; Prat, E.; Meng, D.; Long, Q.; et al. Patterns of polymorphism at the self-incompatibility locus in 1083 Arabidopsis thaliana genomes. Mol. Biol. Evol. 2017, 34, 1878–1889. [Google Scholar] [CrossRef] [PubMed]
- Tantikanjana, T.; Rizvi, N.; Nasrallah, M.E.; Nasrallah, J.B. A dual role for the s-locus receptor kinase in self-incompatibility and pistil development revealed by an Arabidopsis rdr6 mutation. Plant Cell 2009, 21, 2642–2654. [Google Scholar] [CrossRef] [PubMed]
- Indriolo, E.; Goring, D.R. Yeast two-hybrid interactions between Arabidopsis lyrata S Receptor Kinase and the ARC1 E3 ligase. Plant Signal. Behav. 2016, 11, e1188233. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Koch, M.A.; Haubold, B.; Mitchell-Olds, T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis, and related genera Brassicaceae. Mol. Biol. Evol. 2000, 17, 1483–1498. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.I.; Lauga, B.; Charlesworth, D. Rates and patterns of molecular evolution in inbred and outbred Arabidopsis. Mol. Biol. Evol. 2002, 19, 1407–1420. [Google Scholar] [CrossRef] [PubMed]
- Beilstein, M.A.; Nagalingum, N.S.; Clements, M.D.; Manchester, S.R.; Mathews, S. Dated molecular phylogenies indicate a Miocene origin for Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 18724–18728. [Google Scholar] [CrossRef]
- Ossowski, S.; Schneeberger, K.; Lucas-Lledo, J.I.; Warthmann, N.; Clark, R.M.; Shaw, R.G.; Weigel, D.; Lynch, M. The rate and molecular spectrum of spontaneous mutations in Arabidopsis thaliana. Science 2010, 327, 92–94. [Google Scholar] [CrossRef]
- Hu, T.T.; Pattyn, P.; Bakker, E.G.; Cao, J.; Cheng, J.F.; Clark, R.M.; Fahlgren, N.; Fawcett, J.A.; Grimwood, J.; Gundlach, H.; et al. The Arabidopsis lyrata genome sequence and the basis of rapid genome size change. Nat. Genet. 2011, 43, 476–481. [Google Scholar] [CrossRef]
- Bechsgaard, J.S.; Castric, V.; Charlesworth, D.; Vekemans, X.; Schierup, M.H. The transition to self-compatibility in Arabidopsis thaliana and evolution within S-haplotypes over 10 Myr. Mol. Biol. Evol. 2006, 23, 1741–1750. [Google Scholar] [CrossRef]
- Dwyer, K.G.; Berger, M.T.; Ahmed, R.; Hritzo, M.K.; McCulloch, A.A.; Price, M.J.; Serniak, N.J.; Walsh, L.T.; Nasrallah, J.B.; Nasrallah, M.E. Molecular characterization and evolution of self-incompatibility genes in Arabidopsis thaliana: The case of the Sc haplotype. Genetics 2013, 193, 985–994. [Google Scholar] [CrossRef]
- Safavian, D.; Goring, D.R. Secretory activity is rapidly induced in stigmatic papillae by compatible pollen, but inhibited for self-incompatible pollen in the Brassicaceae. PLoS ONE 2013, 8, e84286. [Google Scholar] [CrossRef] [PubMed]
- Lachowiec, J.; Queitsch, C.; Kliebenstein, D.J. Molecular mechanisms governing differential robustness of development and environmental responses in plants. Ann. Bot. 2016, 117, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Schnable, J.C. Functional divergence between subgenomes and gene pairs after whole genome duplications. Mol. Plant 2018, 11, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Hajdukiewicz, P.; Svab, Z.; Maliga, P. The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Mol. Biol. 1994, 25, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Coutu, C.; Brandle, J.; Brown, D.; Brown, K.; Miki, B.; Simmonds, J.; Hegedus, D.D. pORE: A modular binary vector series suited for both monocot and dicot plant transformation. Transgenic Res. 2007, 16, 771–781. [Google Scholar] [CrossRef]
- Foster, E.; Levesque-Lemay, M.; Schneiderman, D.; Albani, D.; Schernthaner, J.; Routly, E.; Robert, L.S. Characterization of a gene highly expressed in the Brassica napus pistil that encodes a novel proline-rich protein. Sex. Plant Reprod. 2005, 17, 261–267. [Google Scholar] [CrossRef]
- Fobis-Loisy, I.; Chambrier, P.; Gaude, T. Genetic transformation of Arabidopsis lyrata: Specific expression of the green fluorescent protein GFP in pistil tissues. Plant Cell Rep. 2007, 26, 745–753. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).