Valuable Fatty Acids in Bryophytes—Production, Biosynthesis, Analysis and Applications



Abstract
1. Introduction
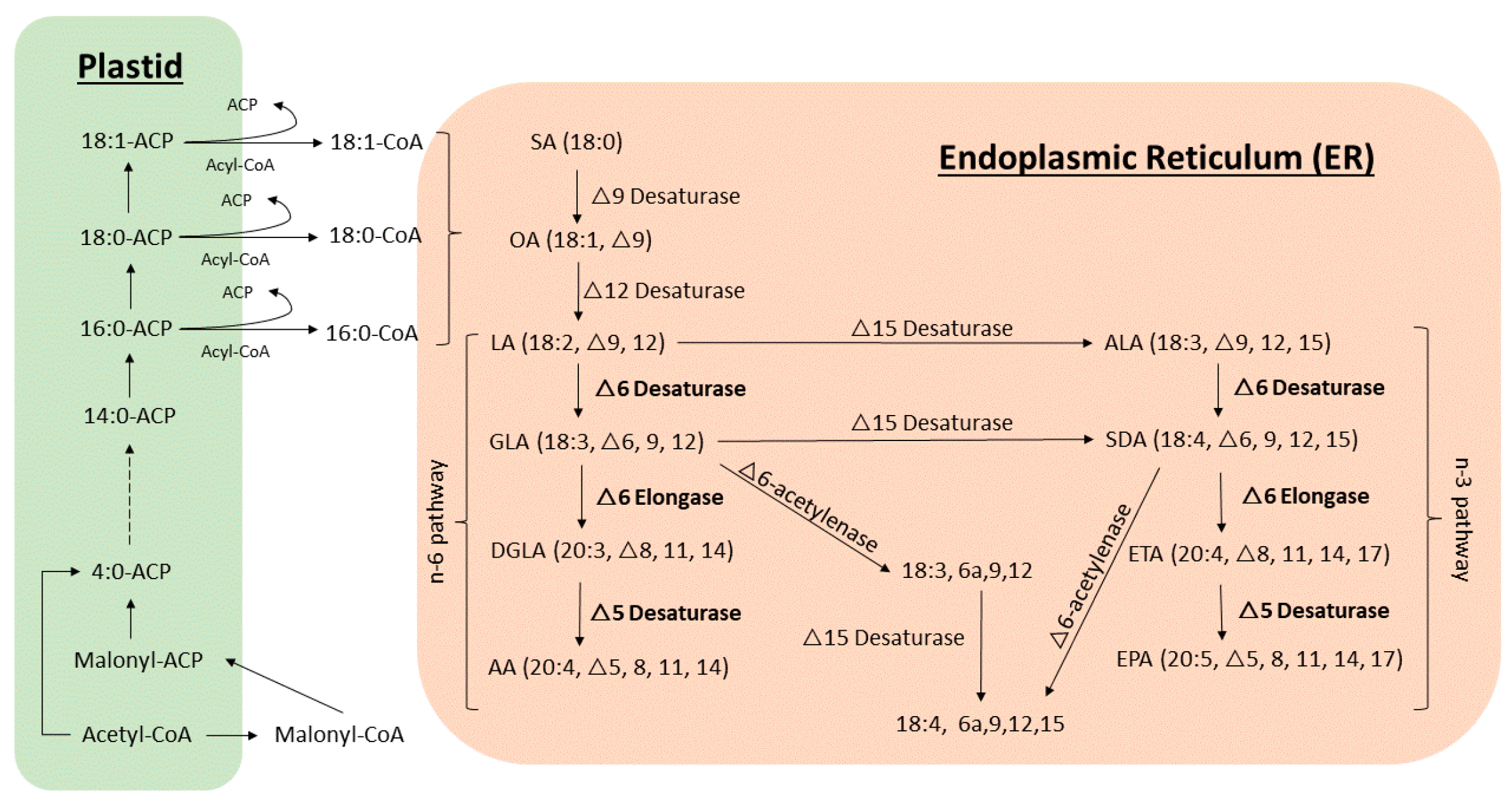
2. Fatty Acid Biosynthetic Pathways
3. Analysis of Fatty Acids in Bryophytes
4. Fatty Acids Present in Bryophytes
5. Arachidonic Acid (AA) and Eicosapentaenoic Acid (EPA)
6. Strategies of Enhancing the Production of vl-PUFAs
6.1. Developmental Stages
6.2. Environmental Stressors
6.3. Genetic Transformation
7. Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AA | arachidonic acid |
| ACPs | acyl carrier proteins |
| AFAs | acetylenic fatty acids |
| ALA | α-linolenic acid |
| APCI | atmospheric pressure chemical ionization |
| DGLA | di-homo γ-linolenic acid |
| EPA | eicosapentaenoic acid |
| ER | endoplasmic reticulum |
| ESI-MS/MS | electrospray ionization triple quadrupole mass spectrometry |
| ETA | eicosatetraenoic acid |
| FAs | fatty acids |
| GC-FID | gas chromatography-flame ionization detector |
| GC-MS | gas chromatography-mass spectrometry |
| GL | glycolipid |
| GLA | γ-linolenic acid |
| HPLC | high-performance liquid chromatography |
| LA | linoleic acid |
| LC-MS | liquid chromatography–mass spectrometry |
| MALDI-TOF-MS | matrix-assisted laser desorption/ionization time-of-flight mass spectrometry |
| MGDG | monogalactosyldiacylglycerol |
| MUFAs | monounsaturated fatty acids |
| NL | neutral lipid |
| NMR | nuclear magnetic resonance |
| OA | oleic acid |
| PC | phosphatidycholine |
| PL | polar lipid |
| PUFAs | polyunsaturated fatty acids |
| SA | stearic acid |
| SDA | stearidonic acid |
| SFAs | saturated fatty acids |
| SPE | solid phase extraction |
| TAGs | triacylglycerides |
| TL | total lipid |
| TLC | thin layer chromatography |
| vl-PUFAs | very long chain polyunsaturated fatty acids |
References
- Asakawa, Y.; Ludwiczuk, A.; Nagashima, F. Chemical Constituents of Bryophytes; Progress in the Chemistry of Organic Natural Products; Springer: Vienna, Austria, 2013; Volume 95, ISBN 978-3-7091-1083-6. [Google Scholar]
- Australian National Botanic Gardens Habitats-Ecology-Bryophytes. Available online: https://www.anbg.gov.au/bryophyte/ecology-habitats.html (accessed on 23 October 2018).
- Qiu, Y.-L.; Palmer, J.D. Phylogeny of early land plants: Insights from genes and genomes. Trends Plant Sci. 1999, 4, 26–30. [Google Scholar] [CrossRef]
- Sheffield, L.; Rowntree, J. Bryophyte Biology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2009; Volume 104. [Google Scholar]
- Vesty, E.F.; Saidi, Y.; Moody, L.A.; Holloway, D.; Whitbread, A.; Needs, S.; Choudhary, A.; Burns, B.; McLeod, D.; Bradshaw, S.J.; et al. The decision to germinate is regulated by divergent molecular networks in spores and seeds. New Phytol. 2016, 211, 952–966. [Google Scholar] [CrossRef]
- He, X.; Sun, Y.; Zhu, R.L.R.-L. The Oil Bodies of Liverworts: Unique and Important Organelles in Land Plants. CRC Crit. Rev. Plant Sci. 2013, 32, 293–302. [Google Scholar] [CrossRef]
- Klavina, L. Composition of Mosses, Their Metabolites and Environmental Stress Impacts. Ph.D. Thesis, University of Latvia, Riga, Latvia, 2018. [Google Scholar]
- Roberts, A.W.; Roberts, E.M.; Haigler, C.H. Moss cell walls: Structure and biosynthesis. Front. Plant Sci. 2012, 3, 166. [Google Scholar] [CrossRef]
- Haines, W.P.; Renwick, J.A.A. Bryophytes as food: Comparative consumption and utilization of mosses by a generalist insect herbivore. Entomol. Exp. Appl. 2009, 133, 296–306. [Google Scholar] [CrossRef]
- Sabovljević, M.S.; Sabovljević, A.D.; Ikram, N.K.K.; Peramuna, A.; Bae, H.; Simonsen, H.T. Bryophytes—An emerging source for herbal remedies and chemical production. Plant Genet. Resour. 2016, 14, 314–327. [Google Scholar] [CrossRef]
- Reski, R. Quantitative moss cell biology. Curr. Opin. Plant Biol. 2018, 46, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Klavina, L.; Bikovens, O.; Steinberga, I.; Maksimova, V.; Eglīte, L. Characterization of chemical composition of some bryophytes common in Latvia. Environ. Exp. Biol. 2012, 10, 27–34. [Google Scholar]
- Beike, A.K.; Jaeger, C.; Zink, F.; Decker, E.L.; Reski, R. High contents of very long-chain polyunsaturated fatty acids in different moss species. Plant Cell Rep. 2014, 33, 245–254. [Google Scholar] [CrossRef]
- Shinmen, Y.; Katoh, K.; Shimizu, S.; Jareonkitmongkol, S.; Yamada, H. Production of arachidonic acid and eicosapentaenoic acids by Marchantia polymorpha in cell culture. Phytochemistry 1991, 30, 3255–3260. [Google Scholar] [CrossRef]
- Baas, M.; Pancost, R.; van Geel, B.; Sinninghe Damsté, J.S. A comparative study of lipids in Sphagnum species. Org. Geochem. 2000, 31, 535–541. [Google Scholar] [CrossRef]
- Tosun, A.; Nagashima, F.; Asakawa, Y. Terpenoid and Steroid Components of Selected Liverworts. Chem. Nat. Compd. 2015, 51, 387–391. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Sim-Sim, M.; Costa, M.M.; Barroso, J.G.; Pedro, L.G.; Esquível, M.G.; Gutierres, F.; Lobo, C.; Fontinha, S. Comparison of the essential oil composition of four Plagiochila species: P. bifaria, P. maderensis, P. retrorsa and P. stricta. Flavour Fragr. J. 2005, 20, 703–709. [Google Scholar] [CrossRef]
- Klavina, L.; Springe, G.; Nikolajeva, V.; Martsinkevich, I.; Nakurte, I.; Dzabijeva, D.; Steinberga, I. Chemical Composition Analysis, Antimicrobial Activity and Cytotoxicity Screening of Moss Extracts (Moss Phytochemistry). Molecules 2015, 20, 17221–17243. [Google Scholar] [CrossRef]
- Asakawa, Y.; Ludwiczuk, A. Bryophytes: Liverworts, Mosses, and Hornworts: Extraction and Isolation Procedures. In Metabolomics Tools for Natural Product Discovery: Methods and Protocols; Roessner, U., Dias, D.A., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 1–20. ISBN 978-1-62703-577-4. [Google Scholar]
- Duckett, J.G.; Burch, J.; Fletcher, P.W.; Matcham, H.W.; Read, D.J.; Russell, A.J.; Pressel, S. In vitro cultivation of bryophytes: A review of practicalities, problems, progress and promise. J. Bryol. 2004, 26, 3–20. [Google Scholar]
- Hohe, A.; Reski, R. From axenic spore germination to molecular farming. Plant Cell Rep. 2005, 23, 513–521. [Google Scholar] [CrossRef]
- Christie, W.W.; Han, X. Lipids: Their structures and occurrence. In Lipid Analysis; Elsevier: Amsterdam, The Netherlands, 2012; pp. 3–19. ISBN 978-0-9552512-4-5. [Google Scholar]
- Dembitsky, V.M. Lipids of bryophytes. Prog. Lipid Res. 1993, 32, 281–356. [Google Scholar] [CrossRef]
- Harris, E.S.J. Ethnobryology: Traditional uses and folk classification of bryophytes. Bryologist 2008, 111, 169–217. [Google Scholar] [CrossRef]
- Chandra, S.; Chandra, D.; Barh, A.; Pankaj; Pandey, R.K.; Sharma, I.P. Bryophytes: Hoard of remedies, an ethno-medicinal review. J. Tradit. Complement. Med. 2017, 7, 94–98. [Google Scholar] [CrossRef]
- Glime, J.M. Volume 5, Chapter 2-1: Medical Uses: Medical Conditions. Available online: http://digitalcommons.mtu.edu/bryophyte-ecology/ (accessed on 12 October 2018).
- Büttner-Mainik, A.; Parsons, J.; Jérôme, H.; Hartmann, A.; Lamer, S.; Schaaf, A.; Schlosser, A.; Zipfel, P.F.; Reski, R.; Decker, E.L. Production of biologically active recombinant human factor H in Physcomitrella. Plant Biotechnol. J. 2011, 9, 373–383. [Google Scholar] [CrossRef]
- Reski, R.; Parsons, J.; Decker, E.L. Moss-made pharmaceuticals: From bench to bedside. Plant Biotechnol. J. 2015, 13, 1191–1198. [Google Scholar] [CrossRef] [PubMed]
- Frahm, J.P.; Kirchhoff, K. Antifeeding effects of bryophyte extracts from Neckera crispa and Porella obtusata against the slug Arion lusitanicus. Cryptogam. Bryol. 2002, 23, 271–275. [Google Scholar]
- Frahm, J.-P.P. Recent Developments of Commercial Products from Bryophytes. Bryologist 2004, 107, 277–283. [Google Scholar] [CrossRef]
- Asakawa, Y. Bryophytes: Chemical diversity, synthesis and biotechnology. A review. Flavour Fragr. J. 2011, 26, 318–320. [Google Scholar] [CrossRef]
- Asakawa, Y.; Ludwiczuk, A.; Nagashima, F. Phytochemical and biological studies of bryophytes. Phytochemistry 2013, 91, 52–80. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Ludwiczuk, A.; Wei, G.; Chen, X.; Crandall-Stotler, B.; Bowman, J.L. Terpenoid Secondary Metabolites in Bryophytes: Chemical Diversity, Biosynthesis and Biological Functions. CRC Crit. Rev. Plant Sci. 2018, 37, 210–231. [Google Scholar] [CrossRef]
- Calder, P.C.; Yaqoob, P. Omega-3 polyunsaturated fatty acids and human health outcomes. BioFactors 2009, 35, 266–272. [Google Scholar] [CrossRef]
- Bhardwaj, K.; Verma, N.; Trivedi, R.K.; Bhardwaj, S.; Shukla, N. Significance of Ratio of Omega-3 and Omega-6 in Human Health with Special Reference to Flaxseed Oil. Int. J. Biol. Chem. 2016, 10, 1–6. [Google Scholar] [CrossRef]
- Resemann, H.C.; Lewandowska, M.; Gömann, J.; Feussner, I. Membrane Lipids, Waxes and Oxylipins in the Moss Model Organism Physcomitrella patens. Plant Cell Physiol. 2019, 60, 1166–1175. [Google Scholar] [CrossRef]
- Jiao, J.; Zhang, Y. Transgenic Biosynthesis of Polyunsaturated Fatty Acids: A Sustainable Biochemical Engineering Approach for Making Essential Fatty Acids in Plants and Animals. Chem. Rev. 2013, 113, 3799–3814. [Google Scholar] [CrossRef]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.-F.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella Genome Reveals Evolutionary Insights into the Conquest of Land by Plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Kohchi, T.; Yamato, K.T.; Jenkins, J.; Shu, S.; Ishizaki, K.; Yamaoka, S.; Nishihama, R.; Nakamura, Y.; Berger, F.; et al. Insights into Land Plant Evolution Garnered from the Marchantia polymorpha Genome. Cell 2017, 171, 287–304.e15. [Google Scholar] [CrossRef] [PubMed]
- Kajikawa, M.; Matsui, K.; Ochiai, M.; Tanaka, Y.; Kita, Y.; Ishimoto, M.; Kohzu, Y.; Shoji, S.; Yamato, K.T.; Ohyama, K.; et al. Production of Arachidonic and Eicosapentaenoic Acids in Plants Using Bryophyte Fatty Acid Δ6-Desaturase, Δ6-Elongase, and Δ5-Desaturase Genes. Biosci. Biotechnol. Biochem. 2008, 72, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Řezanka, T.; Sigler, K. Odd-numbered very-long-chain fatty acids from the microbial, animal and plant kingdoms. Prog. Lipid Res. 2009, 48, 206–238. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Salem, N. Separation of lipid classes by solid phase extraction. J. Lipid Res. 1990, 31, 2285–2289. [Google Scholar]
- Klavina, L.; Kviesis, J. Solid Phase Extraction of Bryophyte Lipids. Mater. Sci. Appl. Chem. 2015, 32, 58–67. [Google Scholar]
- Yang, K.; Zhao, Z.; Gross, R.W.; Han, X. Identification and Quantitation of Unsaturated Fatty Acid Isomers by Electrospray Ionization Tandem Mass Spectrometry: A Shotgun Lipidomics Approach. Anal. Chem. 2011, 83, 4243–4250. [Google Scholar] [CrossRef]
- Ahlstrand, N.I.; Reghev, N.H.; Markussen, B.; Hansen, H.C.B.; Eiriksson, F.F.; Thorsteinsdóttir, M.; Rønsted, N.; Barnes, C.J. Untargeted metabolic profiling reveals geography as the strongest predictor of metabolic phenotypes of a cosmopolitan weed. Ecol. Evol. 2018, 8, 6812–6826. [Google Scholar] [CrossRef]
- Welti, R.; Li, W.; Li, M.; Sang, Y.; Biesiada, H.; Zhou, H.-E.; Rajashekar, C.B.; Williams, T.D.; Wang, X. Profiling Membrane Lipids in Plant Stress Responses. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef]
- Qin, L.; Zhang, Y.; Liu, Y.; He, H.; Han, M.; Li, Y.; Zeng, M.; Wang, X. Recent advances in matrix-assisted laser desorption/ionisation mass spectrometry imaging (MALDI-MSI) for in situ analysis of endogenous molecules in plants. Phytochem. Anal. 2018, 29, 351–364. [Google Scholar] [CrossRef]
- Podolskaya, E.P.; Gladchuk, A.S.; Keltsieva, O.A.; Dubakova, P.S.; Silyavka, E.S.; Lukasheva, E.; Zhukov, V.; Lapina, N.; Makhmadalieva, M.R.; Gzgzyan, A.M.; et al. Thin Film Chemical Deposition Techniques as a Tool for Fingerprinting of Free Fatty Acids by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Anal. Chem. 2019, 91, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Kamide, Y.; Hirai, M.Y.Y.; Saito, K. Plant lipidomics based on hydrophilic interaction chromatography coupled to ion trap time-of-flight mass spectrometry. Metabolomics 2013, 9, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Wei, D.; Chen, F.; Yang, S.-T.T. Lipidomic profiling and discovery of lipid biomarkers in snow alga Chlamydomonas nivalis under salt stress. Eur. J. Lipid Sci. Technol. 2012, 114, 253–265. [Google Scholar] [CrossRef]
- Bromke, M.A.; Sabir, J.S.; Alfassi, F.A.; Hajarah, N.H.; Kabli, S.A.; Al-Malki, A.L.; Ashworth, M.P.; Méret, M.; Jansen, R.K.; Willmitzer, L. Metabolomic Profiling of 13 Diatom Cultures and Their Adaptation to Nitrate-Limited Growth Conditions. PLoS ONE 2015, 10, e0138965. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, X.; Wang, T.; Zhu, P.; Yang, L. The major changes in lipid composition of Sargassum horneri during different growth phases. J. Appl. Phycol. 2018, 30, 517–523. [Google Scholar] [CrossRef]
- Anderson, W.H.; Gellerman, J.L.; Schlenk, H. Arachidonic and eicosapentaenoic acids in developing gametophores and sporophytes of the moss, Mnium cuspidatum. Lipids 1972, 7, 710–714. [Google Scholar] [CrossRef]
- Åndersson, B.; Anderson, W.H.; Chipault, J.R.; Ellison, E.C.; Fenton, S.W.; Gellerman, J.L.; Hawkins, J.M.; Schlenk, H. 9,12,15-octadecatrien-6-ynoic acid, new acetylenic acid from mosses. Lipids 1974, 9, 506–511. [Google Scholar] [CrossRef]
- Kohn, G.; Demmerle, S.S.; Vandekerkhove, O.; Hartmann, E.; Beutelmann, P.; Vandekerkhove, C.; Hartmann, E.; Beutelmann, P.; Vandekerkhove, O.; Hartmann, E.; et al. Distribution and chemotaxonomic significance of acetylenic fatty acids in mosses of the dicranales. Phytochemistry 1987, 26, 2271–2275. [Google Scholar] [CrossRef]
- Vierengel, A.; Kohn, G.; Vandekerkhove, O.; Hartmann, E. 9-octadecen-6-ynoic acid from Riccia fluitans. Phytochemistry 1987, 26, 2101–2102. [Google Scholar] [CrossRef]
- Kohn, G.; Vandekerkhove, O.; Hartmann, E.; Beutelmann, P. Acetylenic fatty acids in the ricciaceae (hepaticae). Phytochemistry 1988, 27, 1049–1051. [Google Scholar] [CrossRef]
- Hansen, C.E.; Rossi, P. Arachidonic and eicosapentaenoic acids in brachytheciaceae and hypnaceae moss species. Phytochemistry 1990, 29, 3749–3754. [Google Scholar] [CrossRef]
- Hansen, C.E.; Rossi, P. Effects of culture conditions on accumulation of arachidonic and eicosapentaenoic acids in cultured cells of Rhytidiadelphus squarrosus and Eurhynchium striatum. Phytochemistry 1991, 30, 1837–1841. [Google Scholar] [CrossRef]
- Sewón, P. Fatty acyl composition of monogalactosyldiacyl glycerols in Bryophyta. Phytochemistry 1992, 31, 3461–3465. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rezanka, T.; Bychek, I.A.; Afonina, O.M. Polar lipid and fatty acid composition of some bryophytes. Phytochemistry 1993, 33, 1009–1014. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rezanka, T. Distribution of diacylglycerylhomoserines, phospholipids and fatty acids in thirteen moss species from Southwestern Siberia. Biochem. Syst. Ecol. 1995, 23, 71–78. [Google Scholar] [CrossRef]
- Saruwatari, M.; Takio, S.; Ono, K. Low temperature-induced accumulation of eicosapentaenoic acids in Marchantia polymorpha cells. Phytochemistry 1999, 52, 367–372. [Google Scholar] [CrossRef]
- Chiou, S.-Y.; Su, W.-W.; Su, Y.-C. Optimizing production of polyunsaturated fatty acids in Marchantia polymorpha cell suspension culture. J. Biotechnol. 2001, 85, 247–257. [Google Scholar] [CrossRef]
- Takemura, M.; Okimura, Y.; Kida, H.; Hamada, T.; Ohyama, K. Blue light enhances the accumulation of eicosapentaenoic acid in a liverwort, Marchantia polymorpha L. Plant Biotechnol. 2011, 28, 489–492. [Google Scholar] [CrossRef]
- Chodok, P.; Kanjana-Opas, A.; Kaewsuwan, S. The Plackett–Burman Design for Evaluating the Production of Polyunsaturated Fatty Acids by Physcomitrella patens. J. Am. Oil Chem. Soc. 2010, 87, 521–529. [Google Scholar] [CrossRef]
- You, J.; Zhao, X.; Suo, Y.; Li, Y.; Wang, H.; Chen, G. Determination of long-chain fatty acids in bryophyte plants extracts by HPLC with fluorescence detection and identification with MS. J. Chromatogr. B 2007, 848, 283–291. [Google Scholar] [CrossRef]
- Pejin, B.; Bianco, A.; Newmaster, S.; Sabovljević, M.; Vujisić, L.; Tešević, V.; Vajs, V.; De Rosa, S. Fatty acids of Rhodobryum ontariense (Bryaceae). Nat. Prod. Res. 2012, 26, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Pejin, B.; Vujisić, L.; Sabovljević, M.; Tešević, V.; Vajs, V. Fatty acid chemistry of Atrichum undulatum and Hypnum andoi. Hem. Ind. 2012, 66, 207–209. [Google Scholar] [CrossRef]
- Mitra, S. High Content of Dicranin in Anisothecium spirale (Mitt.) Broth., a Moss from Eastern Himalayas and Its Chemotaxonomic Significance. Lipids 2017, 52, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, D. Production of Unusual Fatty Acids in Plants. Available online: http://lipidlibrary.aocs.org/Biochemistry/content.cfm?ItemNumber=40317 (accessed on 12 March 2019).
- Prins, H.H.T. Why Are Mosses Eaten in Cold Environments Only? Oikos 1982, 38, 374. [Google Scholar] [CrossRef]
- Glime, J.M. Volume 1, Chapter 10-2: Temperature: Cold. Available online: http://digitalcommons.mtu.edu/bryophyte-ecology/ (accessed on 13 November 2018).
- Chen, Y.; Meesapyodsuk, D.; Qiu, X. Transgenic production of omega-3 very long chain polyunsaturated fatty acids in plants: Accomplishment and challenge. Biocatal. Agric. Biotechnol. 2014, 3, 38–43. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rezanka, T. Acetylenic fatty acids of the dicranaceae. Phytochemistry 1994, 36, 685–689. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rezanka, T.; Bychek, I.A.; Afonina, O.M. Acetylenic acids and lipid compositions of some mosses from Russia. Phytochemistry 1993, 33, 1021–1027. [Google Scholar] [CrossRef]
- Croisier, E.; Rempt, M.; Pohnert, G. Survey of volatile oxylipins and their biosynthetic precursors in bryophytes. Phytochemistry 2010, 71, 574–580. [Google Scholar] [CrossRef]
- Pejin, B.; Vujisić, L.; Sabovljević, M.; Tešević, V.; Vajs, V.; Vujisic, L.; Sabovljevic, M.; Tesevic, V.; Vajs, V. The moss Mnium hornum, a promising source of arachidonic acid. Chem. Nat. Compd. 2012, 48, 120–121. [Google Scholar] [CrossRef]
- Pejin, B.; Vujisic, L.; Sabovljevic, A.; Sabovljevic, M.; Tesevic, V.; Vajs, V. Fatty acids of some moss species from Germany. Asian J. Chem. 2011, 23, 5187–5188. [Google Scholar]
- Koskimies, K.; Simola, L.K. The fatty acid composition of some Sphagnum species. Can. J. Bot. 1980, 58, 259–263. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rezanka, T. Distribution of acetylenic acids and polar lipids in some aquatic bryophytes. Phytochemistry 1995, 40, 93–97. [Google Scholar] [CrossRef]
- Shanab, S.M.M.; Hafez, R.M.; Fouad, A.S. A review on algae and plants as potential source of arachidonic acid. J. Adv. Res. 2018, 11, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Kihara, H.; Tanaka, M.; Yamato, K.T.; Horibata, A.; Yamada, A.; Kita, S.; Ishizaki, K.; Kajikawa, M.; Fukuzawa, H.; Kohchi, T.; et al. Arachidonic acid-dependent carbon-eight volatile synthesis from wounded liverwort (Marchantia polymorpha). Phytochemistry 2014, 107, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, E.; Beutelmann, P.; Vandekerkhove, O.; Euler, R.; Kohn, G. Moss cell cultures as sources of arachidonic and eicosapentaenoic acids. FEBS Lett. 1986, 198, 51–55. [Google Scholar] [CrossRef]
- Gellerman, J.L.; Anderson, W.H.; Richardson, D.G.; Schlenk, H. Distribution of arachidonic and eicosapentaenoic acids in the lipids of mosses. Biochim. Biophys. Acta Lipids Lipid Metab. 1975, 388, 277–290. [Google Scholar] [CrossRef]
- Pejin, B.; Vujisić, L.; Sabovljević, A.; Sabovljević, M.; Tešević, V.; Vajs, V. An insight into fatty acid chemistry of Rhytididelphus squarrosus (Hedw.) Warnst. Bot. Serbia 2011, 35, 99–101. [Google Scholar]
- Abbadi, A.; Domergue, F.; Bauer, J.; Napier, J.A.; Welti, R.; Zähringer, U.; Cirpus, P.; Heinz, E. Biosynthesis of Very-Long-Chain Polyunsaturated Fatty Acids in Transgenic Oilseeds: Constraints on Their Accumulation. Plant Cell 2004, 16, 2734–2748. [Google Scholar] [CrossRef]
- Takemura, M.; Hamada, T.; Kida, H.; Ohyma, K. Cold-Induced Accumulation of ω-3 Polyunsaturated Fatty Acid in a Liverwort, Marchantia polymorpha L. Biosci. Biotechnol. Biochem. 2012, 76, 785–790. [Google Scholar] [CrossRef][Green Version]
- Al-Hasan, R.H.; Ka’wash, H.H.; Radwan, S.S. Enrichment of Mosses with Lipids and Polyunsaturated Fatty Acids by Nitrogen Starvation. Bryologist 1991, 94, 196. [Google Scholar] [CrossRef]
- Kaewsuwan, S.; Bunyapraphatsara, N.; Cove, D.J.; Quatrano, R.S.; Chodok, P. High level production of adrenic acid in Physcomitrella patens using the algae Pavlova sp. Δ5-elongase gene. Bioresour. Technol. 2010, 101, 4081–4088. [Google Scholar] [CrossRef] [PubMed]
- Zank, T.K.; Zähringer, U.; Beckmann, C.; Pohnert, G.; Boland, W.; Holtorf, H.; Reski, R.; Lerchl, J.; Heinz, E. Cloning and functional characterisation of an enzyme involved in the elongation of Δ6-polyunsaturated fatty acids from the moss Physcomitrella patens. Plant J. 2002, 31, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Domergue, F.; Abbadi, A.; Zähringer, U.; Moreau, H.; Heinz, E. In vivo characterization of the first acyl-CoA Δ6-desaturase from a member of the plant kingdom, the microalga Ostreococcus tauri. Biochem. J. 2005, 389, 483–490. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Lipid Types | Methods | Derivatization | References |
|---|---|---|---|---|
| Mnium cuspidotum | FAs | TLC for purification, GC for quantification | Alkylation (diazomethane) | [53] |
| Ceratodon purpureus | FAs | GC for purification, Mass spectrometry for molecular weight, UV and IR for structural confirmation | Alkylation (diazomethane) | [54] |
| 38 moss species | FAs | TLC for purification and identification. GC-MS for FAs analysis | Alkylation (methylation) | [55] |
| Riccia fluitans | FAs | GC for purification, FAs analysis by GC-MS, NMR | Alkylation (methylation) | [56] |
| Twelve liverworts in genus Riccia | FAs | TLC for purification and identification, GC-MS for analysis | Alkylation (methylation) | [57] |
| Eurhynchium striatum, Brachythecium rutabulum, Brachythecium salebrosum, Scleropodium purum, Rhytidiadelphus squarrosus and Rhytidiadelphus triquetrus | FAs (from NL, GL, PL and TL) | HPTLC for NL, PL and GL separation, GC-FID for FA analysis | Acylation (acetyl chloride) | [58] |
| Rhytidiadelphus squurrosus and Eurhynchium striatum | FAs | GC-FID | Trimethylsilylation (Triphenylmethanol) | [59] |
| 55 species of Bryophyta | FAs on MGDG | TLC for separation, GC for FAs analysis | - | [60] |
| Marchantia polymorpha, Atrichum angustatum and Brachythecium sp. | FAs (from TL, NL, PL, GL) | TLC for separation, GC-MS for FAs analysis | Silylation | [61] |
| Calliergon cordifolium, Drepanocladus lycopodioides, Fontinalis antipyretica and Riccia fluitans | FA (fromTL, TAG, PL, GL) | TLC for separation, GC-MS for FAs analysis | Silylation | [62] |
| Marchantia polymorpha | FAs (from NL, GL, PL) | TLC for separation, HPLC with fluorescence detection | No derivatization | [63] |
| Marchantia polymorpha | FAs | GC-FID | alkylation (BF3-methanol | [64] |
| Marchantia polymorpha | FAs | GC-MS | 10% methanolic HCl) | [40] |
| Marchantia polymorpha | FAs | GC | Alkylation | [65] |
| Physcomitrella patens | FAs | GC-FID | Alkylation (methylation) | [66] |
| Homomallium connexum, Actinothuidium hookeri and Neckera pennata | FAs (only carbon number) | HPLC with fluorescence detection, APCI/MS for determination | Alkylation (N,N-dimethyl-formamide) | [67] |
| Rhodobryum ontariense | FAs | GC-MS, NMR | Alkylation (methylation) | [68] |
| Atrichum undulatum and Hypnum andoi | FAs | GC-FID and GC-MS | Alkylation (methylation) | [69] |
| Physcomitrella patens, Encalypta streptocarpa, Pottia lanceolata, Plagiomnium undulatum, Atrichum undulatum, Brachythecium rutabulum and Rhynchostegium murale | FAs | GC-MS | Alkylation (methylation) | [13] |
| Polytrichum commune and Dicranum polysetum | FAs and other hydrophobic compounds | SPE for separation, GC-MS for identification | Silylation (N,O-Bis(trimethylsilyl)-trifluoroacetamide) | [43] |
| Anisothecium spirale | Free FAs, TAG | HP-TLC for lipids separation, GC-FID and GC-MS for quantification | Alkylation (methylation) | [70] |
| Saturated Fatty Acids (SFAs) | ||||||||||||||||||||||||||||||||
| Species | Position | 12:0 | 14:0 | 15:0 | 16:0 | 17:0 | 18:0 | 20:0 | 21:0 | 22:0 | 23:0 | 24:0 | 25:0 | 26:0 | References | |||||||||||||||||
| Marchantia polymorpha | 24.8 * | 1.2 * | [40] | |||||||||||||||||||||||||||||
| Fontinalis antipyretica | TL | 1.5 | 0.4 | 1.2 | 13.6 | 1.3 | 5.6 | 1.0 | [62] | |||||||||||||||||||||||
| TAG | 0.3 | 0.7 | 0.2 | 4.7 | 1.8 | 0.4 | ||||||||||||||||||||||||||
| Riccia fluitans | TL | 0.2 | 0.1 | 0.3 | 8.5 | 0.3 | 1.6 | 0.2 | ||||||||||||||||||||||||
| TAG | 0.5 | 0.2 | 9.3 | 3.0 | 0.3 | |||||||||||||||||||||||||||
| Pellia neesiana | TL | 0.5 | 0.3 | 0.3 | 12.0 | 0.5 | 4.1 | 0.4 | ||||||||||||||||||||||||
| TAG | 0.6 | 0.8 | 0.5 | 6.9 | 0.3 | 5.0 | 1.3 | |||||||||||||||||||||||||
| Calliergon cordifoliu | TL | 0.7 | 0.1 | 0.6 | 10.2 | 0.6 | 3.4 | 0.4 | ||||||||||||||||||||||||
| TAG | 1.3 | 0.1 | 0.4 | 12.7 | 0.3 | 4.7 | 2.0 | |||||||||||||||||||||||||
| Drepanocladus lycopodioides | TL | 0.8 | 0.5 | 0.5 | 11.0 | 0.7 | 6.1 | 0.6 | ||||||||||||||||||||||||
| TAG | 0.9 | 0.7 | 0.6 | 18.3 | 0.4 | 7.2 | 1.1 | |||||||||||||||||||||||||
| Anisothecium spirale | PL | 2.51 * | 16.43 * | 2.36 * | 44.49 * | 1.51 * | 14.19 * | 1.38 * | 1.37 * | [70] | ||||||||||||||||||||||
| NL | 0.14 * | 0.56 * | 0.45 * | 6.70 * | 0.49 * | 0.11 * | 1.29 * | 0.35 * | ||||||||||||||||||||||||
| Phycomitrella patens | 0.47 | 0.47 | 25.67 | 0.30 | 4.24 | 1.43 | 0.28 | 1.36 | 0.32 | 0.98 | 0.12 | 0.10 | [13] | |||||||||||||||||||
| Encalypta vulgaris | 0.60 | 0.55 | 29.97 | 0.25 | 1.27 | 0.78 | 0.13 | 0.67 | 0.34 | 0.49 | 0.05 | 0.04 | ||||||||||||||||||||
| Pottia lanceolata | 0.52 | 0.43 | 30.40 | 0.53 | 2.05 | 1.38 | 0.20 | 0.93 | 0.51 | 2.04 | 0.26 | 0.25 | ||||||||||||||||||||
| Plagiomnium undulatum | 0.26 | 0.24 | 29.79 | 0.60 | 2.70 | 1.62 | 0.49 | 1.52 | 0.41 | 0.73 | 0.04 | 0.10 | ||||||||||||||||||||
| Brachythecium rutabulum | 0.39 | 0.33 | 30.68 | 0.49 | 1.11 | 0.16 | 0.13 | 1.15 | 0.16 | 0.24 | ||||||||||||||||||||||
| Rhynchostegium murale | 0.21 | 0.16 | 23.94 | 0.36 | 1.13 | 0.10 | 0.08 | 0.99 | 0.19 | 0.12 | ||||||||||||||||||||||
| Atrichum undulatum | 0.62 | 0.27 | 24.80 | 0.83 | 1.60 | 1.66 | 0.08 | 0.79 | 0.20 | 1.34 | 0.12 | 0.09 | ||||||||||||||||||||
| Atrichum undulatum | 22.17 | 3.34 | 1.01 | [78] | ||||||||||||||||||||||||||||
| Hypnum andoi | 63.48 | 8.08 | 4.64 | 6.26 | 5.16 | |||||||||||||||||||||||||||
| Mnium hornum | 25.25 | 2.74 | ||||||||||||||||||||||||||||||
| Rhodobryum ontariense | 14.31 | 1.14 | ||||||||||||||||||||||||||||||
| Rhytididelphus squarrosus | 11.90 | 1.00 | [79] | |||||||||||||||||||||||||||||
| Sphagnum fimbriatum | 0.90 | 12.80 | 0.80 | [80] | ||||||||||||||||||||||||||||
| Sphagnum magellanicum | 1.10 | 14.70 | 1.60 | |||||||||||||||||||||||||||||
| Sphagnum najus | 1.10 | 10.70 | 0.90 | |||||||||||||||||||||||||||||
| Sphagnum nemoreum | 0.70 | 11.40 | 2.80 | |||||||||||||||||||||||||||||
| Monounsaturated Fatty Acids (MUFAs) | ||||||||||||||||||||||||||||||||
| Species | Position | 13:1 | 15:1 | 16:1 Δ | 16:1, n-9 | 16:1, n-7 | 17:1, n-9 | 18:1 Δ | 18:1, n-11 | 18:1, n-9 | 18:1, n-7 | C19:1 | C22:1 | References | ||||||||||||||||||
| Fontinalis antipyretica | TL | 1.5 | 0.8 | 2.5 | 1.7 | 0.6 | 1.3 | 13.4 | 0.4 | [62] | ||||||||||||||||||||||
| TAG | 1.1 | 0.6 | 0.3 | 10.2 | 2.2 | |||||||||||||||||||||||||||
| Riccia fluitans | TL | 0.7 | 0.7 | 1.1 | 1.5 | 0.7 | 0.9 | 3.2 | 0.9 | |||||||||||||||||||||||
| TAG | 0.4 | 0.3 | 0.1 | 0.5 | 0.2 | |||||||||||||||||||||||||||
| Pellia neesiana | TL | 0.5 | 0.4 | 3.8 | 2.0 | 0.6 | 0.1 | 10.0 | 0.9 | |||||||||||||||||||||||
| TAG | 0.1 | 2.3 | 0.6 | 0.1 | 0.4 | 8.4 | 0.3 | |||||||||||||||||||||||||
| Calliergon cordifoliu | TL | 0.7 | 0.6 | 3.2 | 1.8 | 0.6 | 0.8 | 6.1 | 0.8 | |||||||||||||||||||||||
| TAG | 0.1 | 0.2 | 2.6 | 1.1 | 0.2 | 1.3 | 10.3 | 1.0 | ||||||||||||||||||||||||
| Drepanocladus lycopodioides | TL | 0.5 | 0.4 | 5.9 | 2.5 | 0.6 | 1.4 | 10.7 | 1.0 | |||||||||||||||||||||||
| TAG | 0.1 | 0.1 | 3.9 | 1.2 | 0.1 | 3.2 | 14.3 | 0.2 | ||||||||||||||||||||||||
| Anisothecium spirale | PL | 6.03 * | [70] | |||||||||||||||||||||||||||||
| NL | 0.65 * | 0.63 * | ||||||||||||||||||||||||||||||
| Phycomitrella patens | 0.32 | 0.48 | 0.06 | 0.06 | [13] | |||||||||||||||||||||||||||
| Encalypta vulgaris | 0.60 | 3.61 | 0.05 | |||||||||||||||||||||||||||||
| Pottia lanceolata | 0.43 | 1.11 | 0.08 | |||||||||||||||||||||||||||||
| Plagiomnium undulatum | 0.38 | 3.03 | 0.10 | |||||||||||||||||||||||||||||
| Brachythecium rutabulum | 0.17 | 4.04 | 0.16 | |||||||||||||||||||||||||||||
| Rhynchostegium murale | 0.12 | 4.85 | 0.06 | |||||||||||||||||||||||||||||
| Atrichum undulatum | 0.46 | 1.06 | 1.84 | 0.08 | ||||||||||||||||||||||||||||
| Atrichum undulatum | 18.49 | [78] | ||||||||||||||||||||||||||||||
| Hypnum andoi | 12.38 | |||||||||||||||||||||||||||||||
| Mnium hornum | 5.13 | |||||||||||||||||||||||||||||||
| Rhodobryum ontariense | 2.47 | |||||||||||||||||||||||||||||||
| Rhytididelphus squarrosus | 2.30 | [79] | ||||||||||||||||||||||||||||||
| Sphagnum fimbriatum | 0.70 | 5.70 | [80] | |||||||||||||||||||||||||||||
| Sphagnum magellanicum | 0.70 | 5.90 | ||||||||||||||||||||||||||||||
| Sphagnum najus | 2.30 | 7.70 | ||||||||||||||||||||||||||||||
| Sphagnum nemoreum | 1.80 | 7.80 | ||||||||||||||||||||||||||||||
| Polyunsaturated Fatty Acids (PUFAs) | ||||||||||||||||||||||||||||||||
| Species | Position | 16:2 | 16:3 Δ | 16:3, n-6 | 16:3, n-3 | 16:4, n-3 | 18:2 Δ | 18:2, n-6 (LA) | 18:2, n-3 | 18:3, n-6 (GLA) | 18:3 Δ | 18:3, n-3 (ALA) | 18:4, n-3 | References | ||||||||||||||||||
| Marchantia polymorpha | 18.4 * | 6.4 * | 0.7 * | 38.9 * | 0.5 * | [40] | ||||||||||||||||||||||||||
| Fontinalis antipyretica | TL | 7.7 | 3.8 | 9.0 | [62] | |||||||||||||||||||||||||||
| TAG | 1.3 | 5.0 | 6.1 | |||||||||||||||||||||||||||||
| Riccia fluitans | TL | 0.6 | 1.7 | 2.3 | 9.1 | 1.2 | 5.6 | 6.2 | 3.1 | |||||||||||||||||||||||
| TAG | 0.1 | 0.3 | 0.6 | 0.2 | 1.2 | 2.2 | ||||||||||||||||||||||||||
| Pellia neesiana | TL | 0.9 | 0.9 | 1.4 | 6.9 | 0.9 | 4.4 | 9.3 | 2.6 | |||||||||||||||||||||||
| TAG | 0.1 | 0.2 | 3.1 | 2.3 | 1.3 | 3.9 | ||||||||||||||||||||||||||
| Calliergon cordifoliu | TL | 0.6 | 1.0 | 1.1 | 8.5 | 0.8 | 10.9 | 14.7 | 3.2 | |||||||||||||||||||||||
| TAG | 0.2 | 0.6 | 1.4 | 9.8 | 0.1 | 11.2 | 19.7 | 5.0 | ||||||||||||||||||||||||
| Drepanocladus lycopodioides | TL | 0.5 | 0.5 | 1.0 | 9.0 | 0.9 | 2.9 | 11.0 | 2.2 | |||||||||||||||||||||||
| TAG | 0.3 | 0.2 | 0.2 | 5.6 | 1.4 | 16.0 | 0.6 | |||||||||||||||||||||||||
| Anisothecium spirale | PL | 0.08 * | 9.63 * | [70] | ||||||||||||||||||||||||||||
| NL | 5.47 * | 1 * | ||||||||||||||||||||||||||||||
| Phycomitrella patens | 4.29 | 2.27 | 24.25 | 13.27 | [13] | |||||||||||||||||||||||||||
| Encalypta vulgaris | 3.66 | 0.34 | 32.02 | 7.66 | ||||||||||||||||||||||||||||
| Pottia lanceolata | 3.76 | 0.17 | 42.88 | 4.68 | ||||||||||||||||||||||||||||
| Plagiomnium undulatum | 1.34 | 0.46 | 24.25 | 11.32 | ||||||||||||||||||||||||||||
| Brachythecium rutabulum | 0.12 | 0.13 | 20.51 | 5.68 | ||||||||||||||||||||||||||||
| Rhynchostegiummurale | 0.22 | 0.00 | 27.51 | 8.42 | ||||||||||||||||||||||||||||
| Atrichum undulatum | 4.61 | 0.76 | 31.49 | 15.31 | ||||||||||||||||||||||||||||
| Atrichum undulatum | 26.80 | 20.50 | [78] | |||||||||||||||||||||||||||||
| Mnium hornum | 11.76 | 19.65 | ||||||||||||||||||||||||||||||
| Rhodobryum ontariense | 5.25 | 20.32 | ||||||||||||||||||||||||||||||
| Rhytididelphus squarrosus | 15.10 | 1.40 | 19.10 | [79] | ||||||||||||||||||||||||||||
| Sphagnum fimbriatum | 22.10 | 38.20 | [80] | |||||||||||||||||||||||||||||
| Sphagnum magellanicum | 22.00 | 34.00 | ||||||||||||||||||||||||||||||
| Sphagnum najus | 25.90 | 33.70 | ||||||||||||||||||||||||||||||
| Sphagnum nemoreum | 30.30 | 29.20 | ||||||||||||||||||||||||||||||
| Acetylenic Acids (AFAs) | ||||||||||||||||||||||||||||||||
| Species | Position | 18:1, 6a | 18:1, 9a | 18:1, 12a | 18:2, 6a, 9 | 18:2, 9, 12a | 18:2, 9a, 12 | 18:3, 6a, 9, 12 | 18:4, 6a, 9, 12, 15 | 20:3, 8a, 11, 14 | 20:4, 5a, 8, 11, 14 | References | ||||||||||||||||||||
| Fontinalis antipyretica | TL | 0.6 | 2.5 | 1.5 | 3.2 | 2.5 | 0.1 | 0.1 | 3.1 | 0.1 | 0.5 | [81] | ||||||||||||||||||||
| TAG | 2.0 | 9.4 | 4.4 | 16.3 | 9.0 | 1.9 | 2.2 | 15.2 | 1.4 | 1.2 | ||||||||||||||||||||||
| Riccia fluitans | TL | 0.8 | 1.6 | 1.5 | 2.9 | 1.1 | 0.9 | 5.3 | 1.0 | 1.4 | 2.6 | |||||||||||||||||||||
| TAG | 1.4 | 7.0 | 6.7 | 9.9 | 4.7 | 3.4 | 24.2 | 5.1 | 6.3 | 11.5 | ||||||||||||||||||||||
| Pellia neesiana | TL | 0.1 | 0.6 | 0.5 | 0.4 | 0.4 | 0.4 | 3.1 | 1.4 | 0.5 | 0.4 | |||||||||||||||||||||
| TAG | 2.7 | 5.8 | 5.4 | 2.6 | 4.1 | 4.1 | 14.1 | 9.2 | 5.2 | 3.8 | ||||||||||||||||||||||
| Calliergon cordifoliu | TL | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.3 | 0.1 | 0.1 | |||||||||||||||||||||
| TAG | 0.1 | 0.1 | 0.2 | 0.3 | 0.6 | 0.8 | 0.3 | 3.3 | 0.5 | 0.4 | ||||||||||||||||||||||
| Drepanocladus lycopodioides | TL | 0.1 | 0.2 | 0.3 | 0.1 | 0.2 | 0.2 | 1.2 | 0.6 | 0.1 | 0.2 | |||||||||||||||||||||
| TAG | 1.1 | 1.2 | 2.1 | 0.8 | 1.2 | 1.1 | 6.3 | 3.1 | 1.0 | 1.1 | ||||||||||||||||||||||
| Anisothecium spirale | PL | 0.5 * | [70] | |||||||||||||||||||||||||||||
| NL | 1.9 * | 72.19 * | ||||||||||||||||||||||||||||||
| Rhodobryum ontariense | 13.3 | 42.3 | [68] | |||||||||||||||||||||||||||||
| Species | Tissues | Regions | AA Contents * | EPA Contents * | References |
|---|---|---|---|---|---|
| Eurhynchium striatum | Gametophyte | Switzerland | 36.7 | 10.8 | [58] |
| Brachythecium rutabulum | 23.5 | 23.4 | |||
| Brachythecium salebrosum | 20.9 | 15.0 | |||
| Scleropodium purum | 29.0 | 8.6 | |||
| Rhytidiadelphus squarrosus | 24.0 | 14.9 | |||
| Rhytidiadelphus triquetrus | 24.6 | 9.5 | |||
| Eurhynchium striatum | Protonema | Collected in Switzerland then keep in cell culture | 33.6 | 2.7 | |
| Brachythecium rutabulum | 40.1 | 4.7 | |||
| Brachythecium salebrosum | 32.2 | 4.9 | |||
| Scleropodium purum | 41.5 | 2.6 | |||
| Rhytidiadelphus squarrosus | 32.4 | 5.2 | |||
| Rhytidiadelphus triquetrus | 20.0 | 1.7 | |||
| Marchantia polymorpha | Cell culture | 11 | 3 | [63] | |
| Marchantia polymorpha | Cell culture | 2.2 + | 2.6 + | [14] | |
| Marchantia polymorhpa | Agar plate, WT | 3.1 # | 5.9 # | [40] | |
| Leptobryum pyriforme | Protonema | Cell culture | 20 | 7 | [84] |
| Physcomitrella patens | Gametophores | Cell culture | 18.7 | 1.5 | [13] |
| Protonema | 15.9 | 6.8 | |||
| Rhynchostegium murale | Gametophores | Cell culture | 26.4 | 3.5 | |
| Mnium cuspidatum | Gametophores | Minnesota | 11.4 | 8.9 | [85] |
| Mnium medium | Gametophores | Minnesota | 23.0 | 19.0 | |
| Hylocomium splendens | Gametophores | Alaska | 12.9 | 18.3 | |
| Pleurozium schreberi | Gametophores | Alaska | 29.0 | 11.0 | |
| Rhytididelphus squarrosus | Gametophores | Germany | 30.7 | 14.4 | [86] |
| Atrichum undulatum | Germany | 6.21 | 1.52 | [69] | |
| Mnium hornum | 26.03 | 9.44 | [78] | ||
| Anisothecium spirale | Gametophyte | Eastern Himalayas | 1.09 # | 0.27 # | [70] |
| Environmental Stresses | |||||||
| Species | Growtd Conditions | Environmental Factors | Variables | Biomass Growtd (mg/plant) | AA * | EPA * | References |
| Rhytidiadelphus squarrosus | MS medium | Temperature | 5 °C | 18.6 | 9.4 | [59] | |
| 10 °C | 23.8 | 13.7 | |||||
| 15 °C | 24.6 | 11.7 | |||||
| 20 °C | 32.7 | 5.7 | |||||
| 25 °C | 26.8 | 6.3 | |||||
| 30 °C | 23.0 | 2.4 | |||||
| Eurhynchium striatum | MS medium | Temperature | 5 °C | 28.4 | 5.6 | ||
| 10 °C | 32.5 | 4.9 | |||||
| 15 °C | 30.5 | 7.5 | |||||
| 20 °C | 34.1 | 2.9 | |||||
| 25 °C | 32.9 | 3.0 | |||||
| 30 °C | 31.7 | 2.4 | |||||
| Rhytidiadelphus squarrosus | MS medium | pH | 5.8 | 13.2 | |||
| 6.5 | 9.5 | ||||||
| Marchantia polymorpha | MS medium | Temperature | 15 °C | 12.1 | 3.6 | [63] | |
| 25 °C | 11 | 3 | |||||
| Marchantia polymorpha | M51C solid medium | Temperature | 5 °C | 2 | 14 | [88] | |
| 25 °C | 3.5 | 5 | |||||
| Ctenidium molluscum | Nitrogen (g/L) | 0 | 22.9 | 5.3 | [89] | ||
| 0.04 | 14.7 | 2.6 | |||||
| 0.4 | 17.5 | 5.0 | |||||
| Pogonatum urnigerum | 0 | 2.8 | 0.4 | ||||
| 0.04 | 3.0 | 3.9 | |||||
| 0.4 | 1.7 | 1.1 | |||||
| Dichodontium pellucidum | 0 | 1.7 | 1.1 | ||||
| 0.04 | 3.4 | 2.2 | |||||
| 0.4 | 3.3 | 1.9 | |||||
| Tortella tortuosa | 0 | 5.1 | 1.8 | ||||
| 0.04 | 5.3 | 1.5 | |||||
| 0.4 | 5.8 | 1.9 | |||||
| Marchantia polymorpha | Photon flux density (umol/m2/s) | 3 | 1028.2 + | 2.8 + | 2.6 + | [64] | |
| 9 | 979.0 + | 2.9 + | 2.8 + | ||||
| 20 | 976.8 + | 3.7 + | 3.4 + | ||||
| 32 | 987.0 + | 3.3 + | 3.1 + | ||||
| Osmolarity (NaCl %) | 0 | 576.5 + | 3.0 + (C20 PUFA) | ||||
| 0.2 | 651.6 + | 3.0 + (C20 PUFA) | |||||
| 0.5 | 62.9 + | 0.2 + (C20 PUFA) | |||||
| Marchantia polymorpha | Light quality | White | 543 | Not reported | [65] | ||
| Blue | 454 | Varied slightly | 1.5-fold higher than under white light | ||||
| Light intensity (umol/m2/s) | 40 | 696 | 5.7 | 2.5 | |||
| 60 | 1075 | 5.7 | 3.0 | ||||
| 80 | 1088 | 5.7 | 4.5 | ||||
| Genetic Transformation | |||||||
| Species | Types | AA * | EPA * | Reference | |||
| Marchantia polymorpha | WT | 3.1 | 5.9 | [40] | |||
| DEOE-34 | 5.0 | 12.1 | |||||
| DEDOE-58 | 11.4 | 8.9 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Eiriksson, F.F.; Thorsteinsdóttir, M.; Simonsen, H.T. Valuable Fatty Acids in Bryophytes—Production, Biosynthesis, Analysis and Applications. Plants 2019, 8, 524. https://doi.org/10.3390/plants8110524
Lu Y, Eiriksson FF, Thorsteinsdóttir M, Simonsen HT. Valuable Fatty Acids in Bryophytes—Production, Biosynthesis, Analysis and Applications. Plants. 2019; 8(11):524. https://doi.org/10.3390/plants8110524
Chicago/Turabian StyleLu, Yi, Finnur Freyr Eiriksson, Margrét Thorsteinsdóttir, and Henrik Toft Simonsen. 2019. "Valuable Fatty Acids in Bryophytes—Production, Biosynthesis, Analysis and Applications" Plants 8, no. 11: 524. https://doi.org/10.3390/plants8110524
APA StyleLu, Y., Eiriksson, F. F., Thorsteinsdóttir, M., & Simonsen, H. T. (2019). Valuable Fatty Acids in Bryophytes—Production, Biosynthesis, Analysis and Applications. Plants, 8(11), 524. https://doi.org/10.3390/plants8110524