Seedling Emergence from Seed Banks in Ludwigia hexapetala-Invaded Wetlands: Implications for Restoration



Abstract
:1. Introduction
2. Results
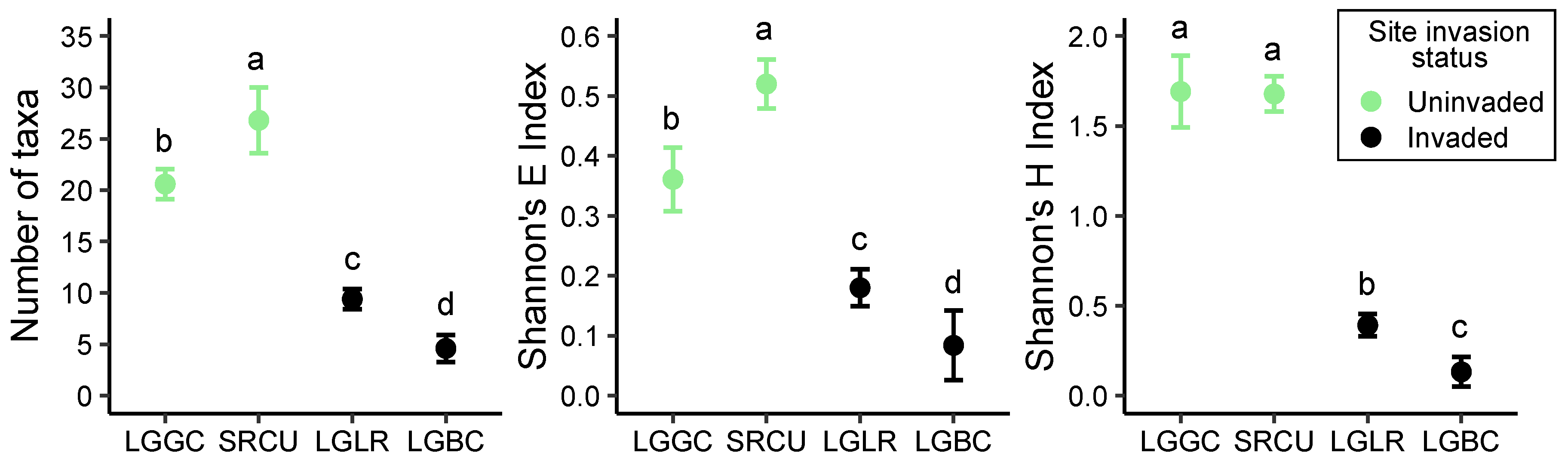
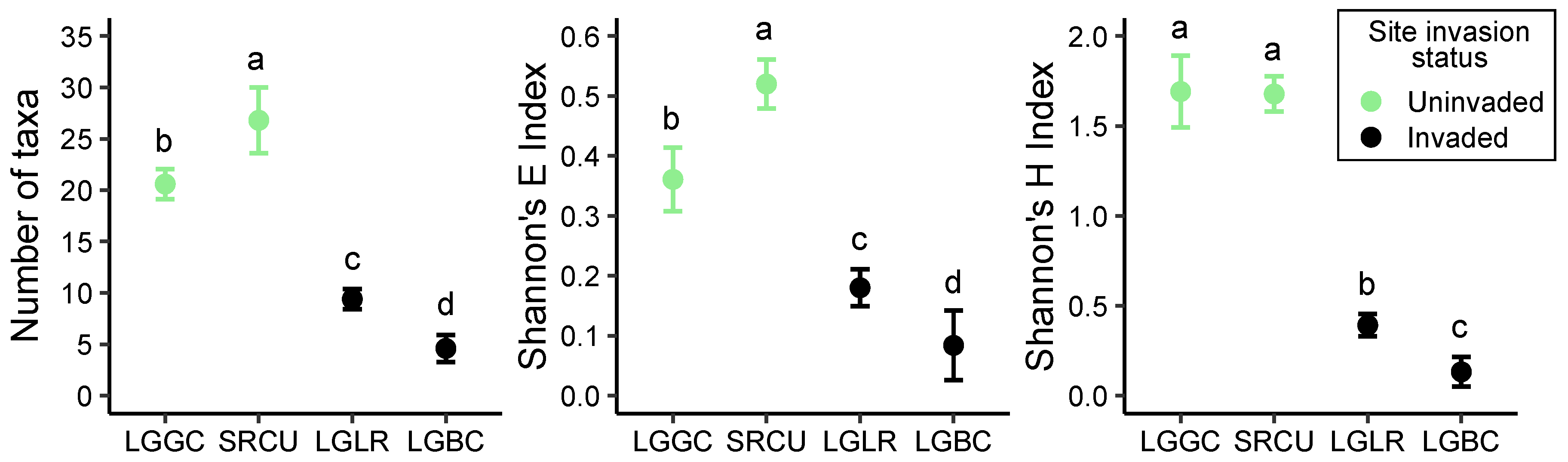
2.1. Invader Impact on Community Composition and Diversity
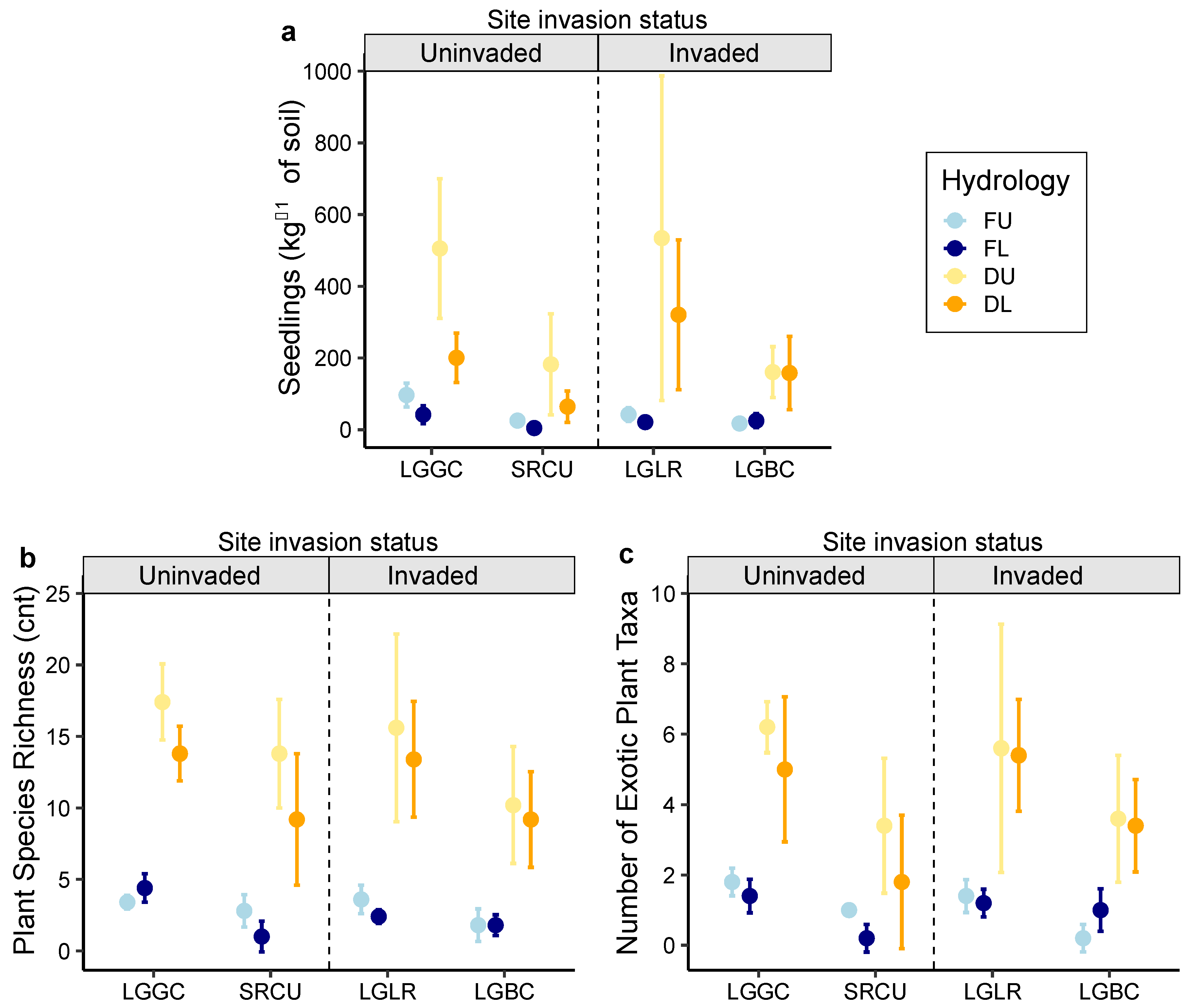
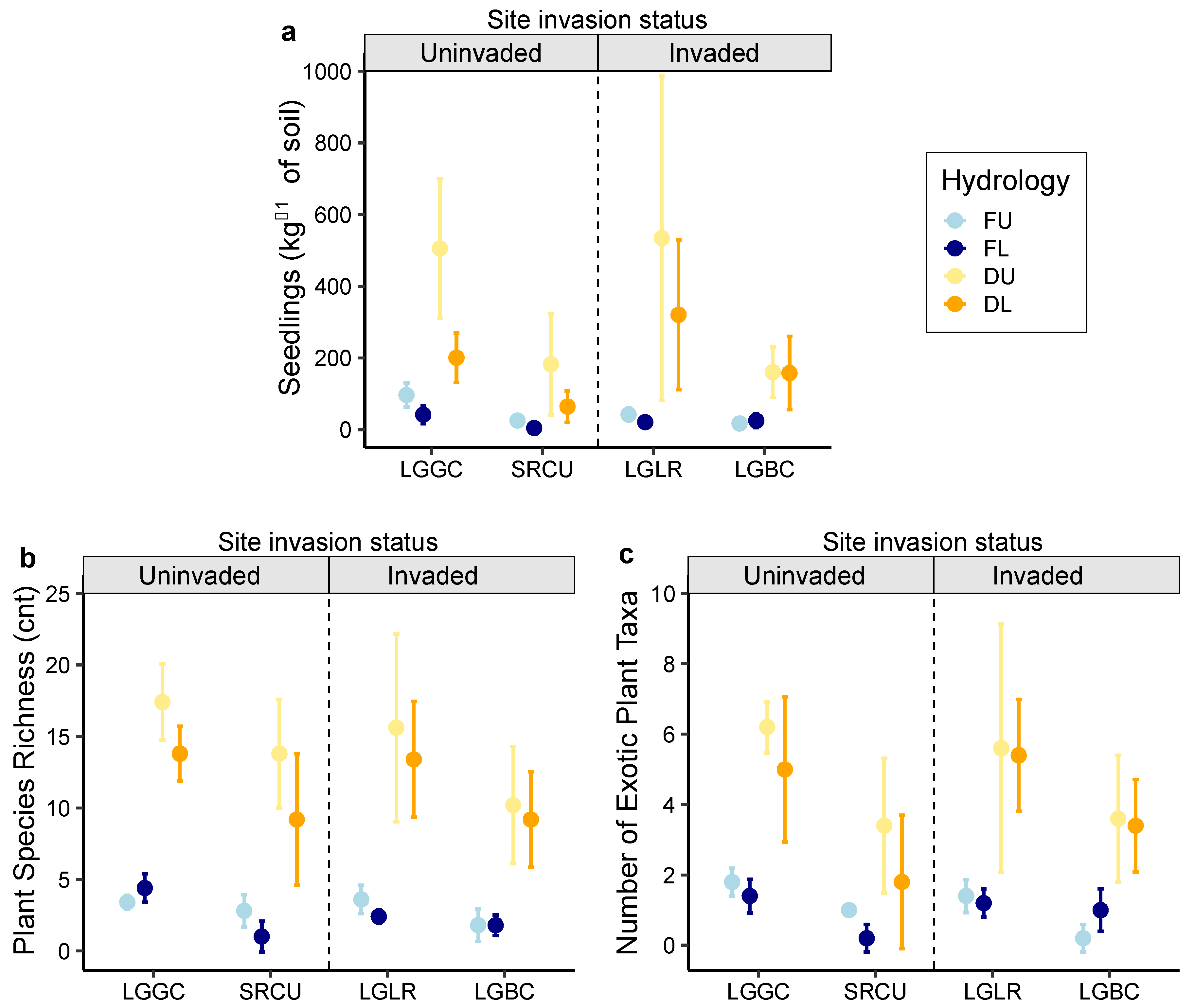
2.2. Seeding Emergence Assay
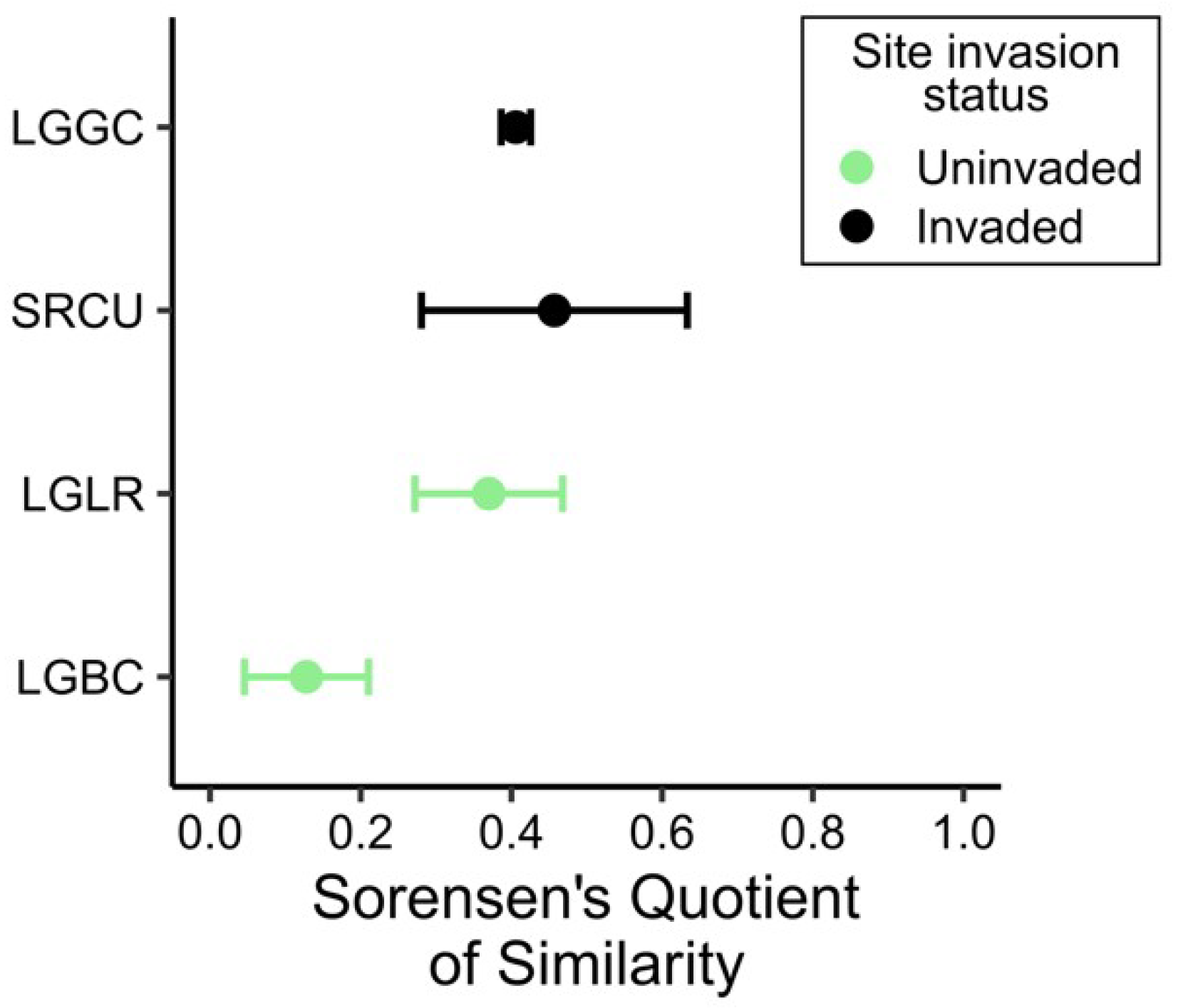
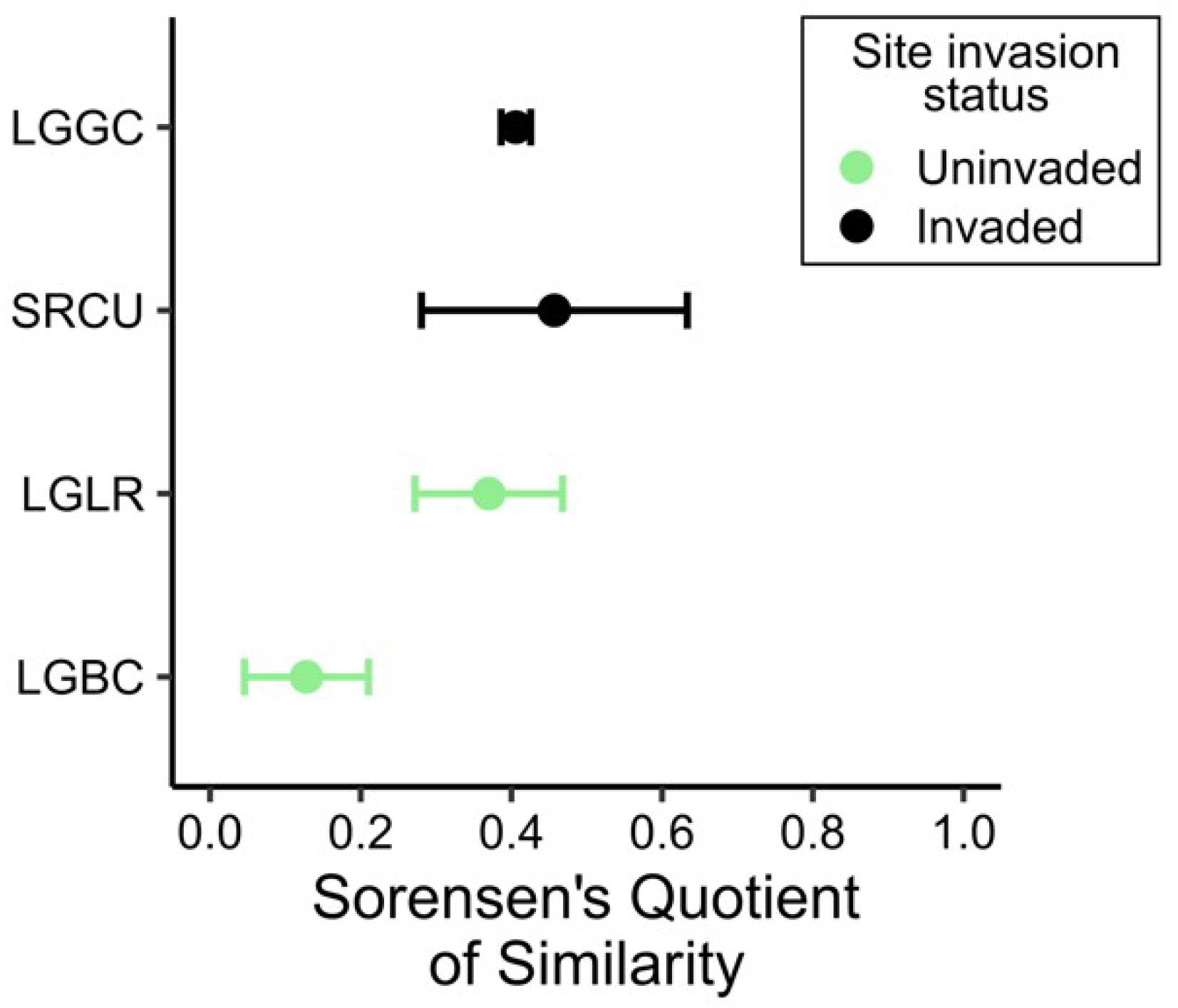
2.3. Comparison of the Soil Seed Bank and Extant Vegetation
3. Discussion
4. Materials and Methods
4.1. Study Area and Focal Invasive Species
4.2. Seed Bank Sampling and Seedling Emergence Assay
4.3. Standing Vegetation Assessment and Comparison to Seed Banks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vilá, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošık, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.J.W.; Grime, J.P. An experimental study of plant community invasibility. Ecology 1996, 77, 776–790. [Google Scholar] [CrossRef]
- Pausas, J.G.; Lloret, F.; Vilá, M. Simulating the effects of different disturbance regimes on Cortaderia selloana invasion. Biol. Cons. 2006, 128, 128–135. [Google Scholar] [CrossRef]
- Loo, S.E.; Mac Nally, R.; O’Dowd, D.J.; Lake, P.S. Secondary invasions: Implications of riparian restoration for in-stream invasion by an aquatic grass. Rest. Ecol. 2009, 17, 378–385. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Mooney, H.A. Community changes following shrub invasion of grassland. Oecologia 1986, 70, 508–513. [Google Scholar] [CrossRef]
- Bakker, J.; Poschlod, P.; Strykstra, R.; Bekker, R.; Thompson, K. Seed banks and seed dispersal: Important topics in restoration ecology. Acta Bot. Neerl. 1996, 45, 461–490. [Google Scholar] [CrossRef]
- Gioria, M.; Jarošík, V.; Pyšek, P. Impact of invasions by alien plants on soil seed bank communities: Emerging patterns. Perspect. Plant Ecol. 2014, 16, 132–142. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species by Means of Natural Selection, or Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Richardson, D.M.; Holmes, P.M.; Esler, K.J.; Galatowitsch, S.M.; Stromberg, J.C.; Kirkman, S.P.; Pysek, P.; Hobbs, R.J. Riparian vegetation: Degradation, alien plant invasions, and restoration prospects. Divers. Distrib. 2007, 13, 126–139. [Google Scholar] [CrossRef]
- Tererai, F.; Gaertner, M.; Jacobs, S.M.; Richardson, D.M. Resilience of invaded riparian landscapes: The potential role of soil-stored seed banks. Environ. Manag. 2015, 55, 86–99. [Google Scholar] [CrossRef]
- Planty-Tabacchi, A.M.; Tabacchi, E.; Naiman, R.J.; Deferrari, C.; Décamps, H. Invasibility of species-rich communities in riparian zones. Conserv. Biol. 1996, 10, 598–607. [Google Scholar] [CrossRef]
- Pysek, P.; Prach, K. Invasion dynamics of Impatiens glandulifera—A century of spreading reconstructed. Biol. Conserv. 1995, 74, 41–48. [Google Scholar] [CrossRef]
- Skaer Thomason, M.J.; McCort, C.D.; Netherland, M.D.; Grewell, B.J. Temporal and nonlinear dispersal patterns of Ludwigia hexapetala in a regulated river. Wetl. Ecol. Manag. 2018, 26, 751–762. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T.; Pedersen, L.J.; Jensen, L. Top-down control in freshwater lakes: The role of nutrient state, submerged macrophytes and water depth. Hydrobiologia 1997, 342, 151–164. [Google Scholar] [CrossRef]
- Zedler, J.B. Progress in wetland restoration ecology. Trends Ecol. Evol. 2000, 15, 402–407. [Google Scholar] [CrossRef]
- Klötzli, F.; Grootjans, A.P. Restoration of natural and semi-natural wetland systems in Central Europe: Progress and predictability of developments. Restor. Ecol. 2001, 9, 209–219. [Google Scholar] [CrossRef]
- Vilá, M.; Gimeno, I. Does invasion by an alien plant species affect the soil seedbank? J. Veg. Sci. 2007, 18, 423–430. [Google Scholar] [CrossRef]
- Gioria, M.; Osborne, B. Similarities in the impact of three large invasive plant species on soil seed bank communities. Biol. Inv. 2010, 12, 1671–1683. [Google Scholar] [CrossRef]
- Abella, S.R.; Chiquoine, L.P.; Backer, D.M. Soil, vegetation, and seed bank of a Sonoran Desert ecosystem along an exotic plant (Pennisetum ciliare) treatment gradient. Environ. Manag. 2013, 52, 946–957. [Google Scholar] [CrossRef]
- Gordon, D.R. Effects of invasive, non-indigenous plant species on ecosystem processes: Lessons from Florida. Ecol. Appl. 1998, 8, 975–989. [Google Scholar] [CrossRef]
- Manchester, S.J.; Bullock, J.M. The impacts of non-native species on UK biodiversity and the effectiveness of control. J. Appl. Ecol. 2000, 37, 845–864. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarosik, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Gioria, M.; Pyšek, P.; Moravcova, L. Soil seed banks in plant invasions: Promoting species invasiveness and long term impact on plant community dynamics. Preslia 2012, 84, 327–350. [Google Scholar]
- Espeland, E.K.; Perkins, L.B.; Leger, E.A. Comparison of seed bank estimation techniques using six weed species in two soil types. Rangel Ecol. Manag. 2010, 63, 243–247. [Google Scholar] [CrossRef]
- Schneider, H.E.; Allen, E.B. Effects of elevated nitrogen and exotic plant invasion on soil seed bank composition in Joshua Tree National Park. Plant Ecol. 2012, 213, 1277–1287. [Google Scholar] [CrossRef]
- Leck, M.A.; Simpson, R.L. Seed bank of a freshwater tidal wetland: Turnover and relationship to vegetation change. Am. J. Bot. 1987, 74, 360–370. [Google Scholar] [CrossRef]
- Grillas, P.; Garcia-Murillo, P.; Geertz-Hansen, O.; Marbá, N.; Montes, C.; Duarte, C.M.; Tan Ham, L.; Grossmann, A. Submerged macrophyte seed bank in a Mediterranean temporary marsh: Abundance and relationship with established vegetation. Oecologia 1993, 94, 1–6. [Google Scholar] [CrossRef]
- Smith, L.M.; Kadlec, J.A. Seed banks and their role during drawdown of a North American marsh. J. Appl. Ecol. 1983, 20, 673–684. [Google Scholar] [CrossRef]
- Wilson, S.D.; Moore, D.R.J.; Keddy, P.A. Relationships of marsh seed banks to vegetation patterns along environmental gradients. Freshw. Biol. 1993, 29, 361–370. [Google Scholar] [CrossRef]
- Combroux, I.C.S.; Bornette, G.; Amoros, C. Plant regenerative strategies after a major disturbance: The case of a riverine wetland restoration. Wetlands 2002, 22, 234–246. [Google Scholar] [CrossRef]
- Wetzel, P.R.; van der Valk, A.G.; Toth, L.A. Restoration of wetland vegetation on the Kissimmee River floodplain: Potential role of seed banks. Wetlands 2001, 21, 189–198. [Google Scholar] [CrossRef]
- Gillard, M.; Grewell, B.J.; Deleu, C.; Futrell, C.J.; Thiébaut, G. Germination and seedling growth of water primroses: A cross experiment between two invaded ranges with contrasting climates. Front. Plant Sci. 2017, 8, 1677. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.E.; Baldwin, A.H. Seedling emergence from seed banks of tidal freshwater wetlands: Response to inundation and sedimentation. Aquat. Bot. 2004, 78, 243–254. [Google Scholar] [CrossRef]
- Van der Valk, A.G.; Davis, C.B. Role of the seed bank in the vegetative dynamics of prairie glacial marshes. Ecology 1978, 59, 322–335. [Google Scholar] [CrossRef]
- Keddy, P.A.; Ellis, T.H. Seedling recruitment of 11 wetland plant species along a water level gradient: Shared or distinct responses? Can. J. Bot. 1985, 63, 1876–1879. [Google Scholar] [CrossRef]
- Casanova, M.T.; Brock, M.A. How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol. 2000, 147, 237–250. [Google Scholar] [CrossRef]
- Nicol, J.M.; Ganf, G.G.; Pelton, G.A. Seed banks of a southern Australian wetland: The influence of water regime on the final floristic composition. Plant Ecol. 2003, 168, 191–205. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: London, UK, 2001. [Google Scholar]
- Avernethy, V.J.; Willby, N.J. Changes along a disturbance gradient in the density and composition of propagule banks in floodplain aquatic habitats. Plant Ecol. 1999, 140, 177–190. [Google Scholar] [CrossRef]
- Jurik, T.W.; Wang, S.C.; van der Valk, A.G. Effects of sediment load on seedling emergence from wetland seed banks. Wetlands 1994, 14, 159–165. [Google Scholar] [CrossRef]
- Dittmar, L.A.; Neely, R.K. Wetland seed bank response to sedimentation varying in loading rate and texture. Wetlands 1999, 19, 341–351. [Google Scholar] [CrossRef]
- Brown, S.C. Remnant seed banks and vegetation as predictors of restored marsh vegetation. Can. J. Bot. 1998, 76, 620–629. [Google Scholar] [CrossRef]
- Mason, T.J.; French, K.; Russell, K.J. Moderate impacts of plant invasion and management in coastal hind dune seed banks. Biol. Conserv. 2006, 134, 428–439. [Google Scholar] [CrossRef]
- Liu, G.; Li, W.; Zhou, J.; Liu, W.Z.; Yang, D.; Davy, A.J. How does the propagule bank contribute to cyclic vegetation change in a lakeshore marsh with seasonal drawdown? Aquat. Bot. 2006, 84, 137–143. [Google Scholar] [CrossRef]
- Handlová, V.; Münzbergová, Z. Seed banks of managed and degraded grasslands in the Krkonoš Mts., Czech Republic. Folia Geobot. 2006, 41, 275–288. [Google Scholar] [CrossRef]
- Parker, I.M.; Simberloff, D.; Lonsdale, W.M.; Goodell, K.; Wonham, M.; Karieva, P.M.; Williamson, M.H.; Von Holle, B.; Moyle, P.B.; Byers, J.E.; et al. Impact: Toward a framework for understanding the ecological effects of invaders. Biol. Inv. 1999, 1, 3–19. [Google Scholar] [CrossRef]
- Grewell, B.J.; Netherland, M.D.; Skaer Thomason, M.J. Establishing Research and Management Priorities for Invasive Water Primrose (Ludwigia spp.); Technical Report ERDC/EL TR-16-2; U.S. Army Corps of Engineers, Engineer Research and Development Center/Environmental Laboratory: Vicksburg, MS, USA, 2016. [Google Scholar]
- Wagner, W.L.; Hoch, P.C.; Raven, P.H. Revised classification of the Onagraceae. Syst. Bot. Monographs 2007, 83, 1–222. [Google Scholar]
- Cui, N.; Wu, J.; Xiang, D.; Cheng, S.; Zhou, Q. A field study on seed bank and its potential applications in vegetation restoration of a polluted urban river in China. Ecol. Eng. 2013, 60, 37–44. [Google Scholar] [CrossRef]
- Esler, K.J.; Holmes, P.M.; Richardson, D.M.; Witkowski, E.T.F. Riparian vegetation management in landscapes invaded by alien plants: Insights from South Africa. S. Afr. J. Bot. 2008, 74, 397–400. [Google Scholar] [CrossRef]
- Gioria, M.; Dieterich, B.; Osborne, B. Battle of the giants: Primary and secondary invasions by large herbaceous species. Biol. Environ. 2011, 111, 1–17. [Google Scholar] [CrossRef]
- Bakker, J.D.; Wilson, S.D. Using ecological restoration to constrain biological invasion. J. Appl. Ecol. 2004, 41, 1058–1064. [Google Scholar] [CrossRef]
- Ruwanza, S.; Gaertner, M.; Esler, K.J.; Richardson, D.M. The effectiveness of active and passive restoration on recovery of indigenous vegetation in riparian zones in the Western Cape, South Africa. S. Afr. J. Bot. 2013, 88, 132–141. [Google Scholar] [CrossRef]
- Okada, M.; Grewell, B.J.; Jasieniuk, M. Clonal spread of invasive Ludwigia hexapetala and L. grandiflora in freshwater wetlands of California. Aquat. Bot. 2009, 91, 123–129. [Google Scholar] [CrossRef]
- Ruaux, B.; Greulich, S.; Haury, J.; Berton, J.P. Sexual reproduction of two alien invasive Ludwigia (Onagraceae) on the Middle Loire River, France. Aquat. Bot. 2009, 90, 143–148. [Google Scholar] [CrossRef]
- Gillard, M.; Grewell, B.J.; Deleu, C.; Thiébaut, G. Climate warming and water primroses: Germination responses of populations from two invaded ranges. Aquat. Bot. 2017, 136, 155–163. [Google Scholar] [CrossRef]
- Kellogg, C.H.; Bridgham, S.D. Colonization during early succession of restored freshwater marshes. Can. J. Bot. 2002, 80, 176–185. [Google Scholar] [CrossRef]
- Neff, K.P.; Rusello, K.; Baldwin, A.H. Rapid seed bank development in restored tidal freshwater wetlands. Rest. Ecol. 2009, 17, 539–548. [Google Scholar] [CrossRef]
- Wisheu, I.C.; Keddy, P.A. Seed banks of a rare wetland plant community, distribution patterns and effects of human-induced disturbance. J. Veget. Sci. 1991, 2, 181–188. [Google Scholar] [CrossRef]
- Hopfensperger, K.N. A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 2007, 116, 1438–1448. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M.; Chester, E.W. Long-term persistence of summer annuals in soil seed banks of seasonally dewatered mudflats. Plant Ecol. 2019, 220, 731–740. [Google Scholar] [CrossRef]
- Abella, S.R.; Chiquoine, L.P.; Vanier, C.H. Characterizing soil seed banks and relationships to plant communities. Plant Ecol. 2013, 214, 703–715. [Google Scholar] [CrossRef]
- Alexander, J.M.; D’Antonio, C.M. Seed bank dynamics of French broom in coastal California grasslands: Effects of stand age and prescribed burning on control and restoration. Rest. Ecol. 2003, 11, 185–197. [Google Scholar] [CrossRef]
- Beas, B.J.; Smith, L.M.; Hickman, K.R.; LaGrange, T.G.; Stutheit, R. Seed bank responses to wetland restoration: Do restored wetlands resemble reference conditions following sediment removal? Aquat. Bot. 2013, 108, 7–15. [Google Scholar] [CrossRef]
- Ooi, M.K.; Auld, T.D.; Denham, A.J. Climate change and bet-hedging: Interactions between increased soil temperature and seed bank persistence. Glob. Change Biol. 2009, 15, 2375–2386. [Google Scholar] [CrossRef]
- D’Antonio, C.M.; Jackson, N.E.; Horvitz, C.C.; Hedberg, R. Invasive plants in wildland ecosystems: Merging the study of invasion processes with management needs. Front. Ecol. Environ. 2004, 2, 513–521. [Google Scholar] [CrossRef]
- Ramsar (The Ramsar Convention on Wetlands). The List of Wetlands of International Importance. 9 September 2019. Available online: http://www.ramsar.org/sites/default/files/documents/library/sitelist (accessed on 16 September 2019).
- Baumgarten, S.; Grossinger, R.M.; Beller, E.E.; Trowbridge, W.; Askevold, R.A. Historical Ecology and Landscape Change in the Central Laguna de Santa Rosa; SFEI: Richmond, CA, USA, 2017. [Google Scholar]
- Cronk, C.B.; Fuller, J.L. Plant Invaders: The threat to Natural Ecosystems; Chapman and Hall: London, UK, 1995. [Google Scholar]
- Thouvenot, L.; Haury, J.; Thiebaut, G. A success story: Water primroses, aquatic plant pests. Aquat. Conserv. 2013, 23, 790–803. [Google Scholar] [CrossRef]
- Dandelot, S.; Verlaque, R.; Dutartre, A.; Cazaubon, A. Ecological, dynamic and taxonomic problems due to Ludwigia (Onagraceae) in France. Hydrobiologia 2005, 551, 131–136. [Google Scholar] [CrossRef]
- Rejmánková, E. Ecology of creeping macrophytes with special reference to Ludwigia peploides (H.B.K.) Raven. Aquat. Bot. 1992, 43, 283–299. [Google Scholar] [CrossRef]
- Poiani, K.A.; Johnson, W.C. Evaluation of the emergence method in estimating seed bank composition of prairie wetlands. Aquat. Bot. 1988, 32, 91–97. [Google Scholar] [CrossRef]
- Gross, K.L. A comparison of methods for estimating seed numbers in the soil. J. Ecol. 1990, 78, 1079–1093. [Google Scholar] [CrossRef]
- Flora of North America Editorial Committee. Flora of North America North of Mexico. Available online: http://floranorthamerica.org/cite (accessed on 30 September 2019).
- Baldwin, B.G.; Goldman, D.H.; Keil, D.J.; Patterson, R.; Rosatti, T.J.; Wilken, D.H. (Eds.) The Jepson Manual: Vascular Plants of California, second edition; University of California Press: Berkeley, CA, USA, 2012. [Google Scholar]
- Zardini, E.M.; Gu, H.; Raven, P.H. On the separation of two species within the Ludwigia uruguayensis complex (Onagraceae). Syst. Bot. 1991, 16, 242–244. [Google Scholar] [CrossRef]
- Zardini, E.M.; Peng, C.; Hoch, P.C. Chromosome numbers if Ludwigia sect. Oligospermum and sect. Oocarpon (Onagraceae). Taxon 1991, 40, 221–230. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Sørensen, T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Biologiske Skrifter 1948, 5, 1–34. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Num DF | Den DF | F-Value | p | |
|---|---|---|---|---|---|
| Total Seedlings | SITE | 3 | 61 | 13.63 | <0.0001 |
| HYD | 1 | 61 | 145.26 | <0.0001 | |
| DPT | 1 | 61 | 14.97 | 0.0003 | |
| HYD*DPT | 1 | 61 | 0 | 0.9769 | |
| SITE*HYD | 3 | 61 | 1.24 | 0.3022 | |
| SITE*DPT | 3 | 61 | 1.71 | 0.1743 | |
| SITE*HYD*DPT | 3 | 61 | 0.10 | 0.9600 | |
| Taxa Richness | SITE | 3 | 64 | 7.35 | 0.0003 |
| HYD | 1 | 64 | 198.95 | <0.0001 | |
| DPT | 1 | 64 | 4.93 | 0.0299 | |
| HYD*DPT | 1 | 64 | 0.09 | 0.7609 | |
| SITE*HYD | 3 | 64 | 0.90 | 0.4480 | |
| SITE*DPT | 3 | 64 | 1.79 | 0.1587 | |
| SITE*HYD*DPT | 3 | 64 | 1.17 | 0.3275 | |
| Exotic Seedlings | SITE | 3 | 64 | 15.76 | <0.0001 |
| HYD | 1 | 64 | 52.71 | <0.0001 | |
| DPT | 1 | 64 | 9.33 | 0.0033 | |
| HYD*DPT | 1 | 64 | 0.02 | 0.8807 | |
| SITE*HYD | 3 | 64 | 0.96 | 0.4189 | |
| SITE*DPT | 3 | 64 | 3.53 | 0.0196 | |
| SITE*HYD*DPT | 3 | 64 | 1.59 | 0.2009 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grewell, B.J.; Gillard, M.B.; Futrell, C.J.; Castillo, J.M. Seedling Emergence from Seed Banks in Ludwigia hexapetala-Invaded Wetlands: Implications for Restoration. Plants 2019, 8, 451. https://doi.org/10.3390/plants8110451
Grewell BJ, Gillard MB, Futrell CJ, Castillo JM. Seedling Emergence from Seed Banks in Ludwigia hexapetala-Invaded Wetlands: Implications for Restoration. Plants. 2019; 8(11):451. https://doi.org/10.3390/plants8110451
Chicago/Turabian StyleGrewell, Brenda J., Morgane B. Gillard, Caryn J. Futrell, and Jesús M. Castillo. 2019. "Seedling Emergence from Seed Banks in Ludwigia hexapetala-Invaded Wetlands: Implications for Restoration" Plants 8, no. 11: 451. https://doi.org/10.3390/plants8110451
APA StyleGrewell, B. J., Gillard, M. B., Futrell, C. J., & Castillo, J. M. (2019). Seedling Emergence from Seed Banks in Ludwigia hexapetala-Invaded Wetlands: Implications for Restoration. Plants, 8(11), 451. https://doi.org/10.3390/plants8110451