Advances in Molecular Genetics and Genomics of African Rice (Oryza glaberrima Steud)



Abstract
1. Background
2. Genetic Potential and Capacity for Climate Change Adaptation
3. Genetic and Molecular Basis of Important Traits
4. Genomic and Transcriptomic Resources
4.1. Genomic Sequences
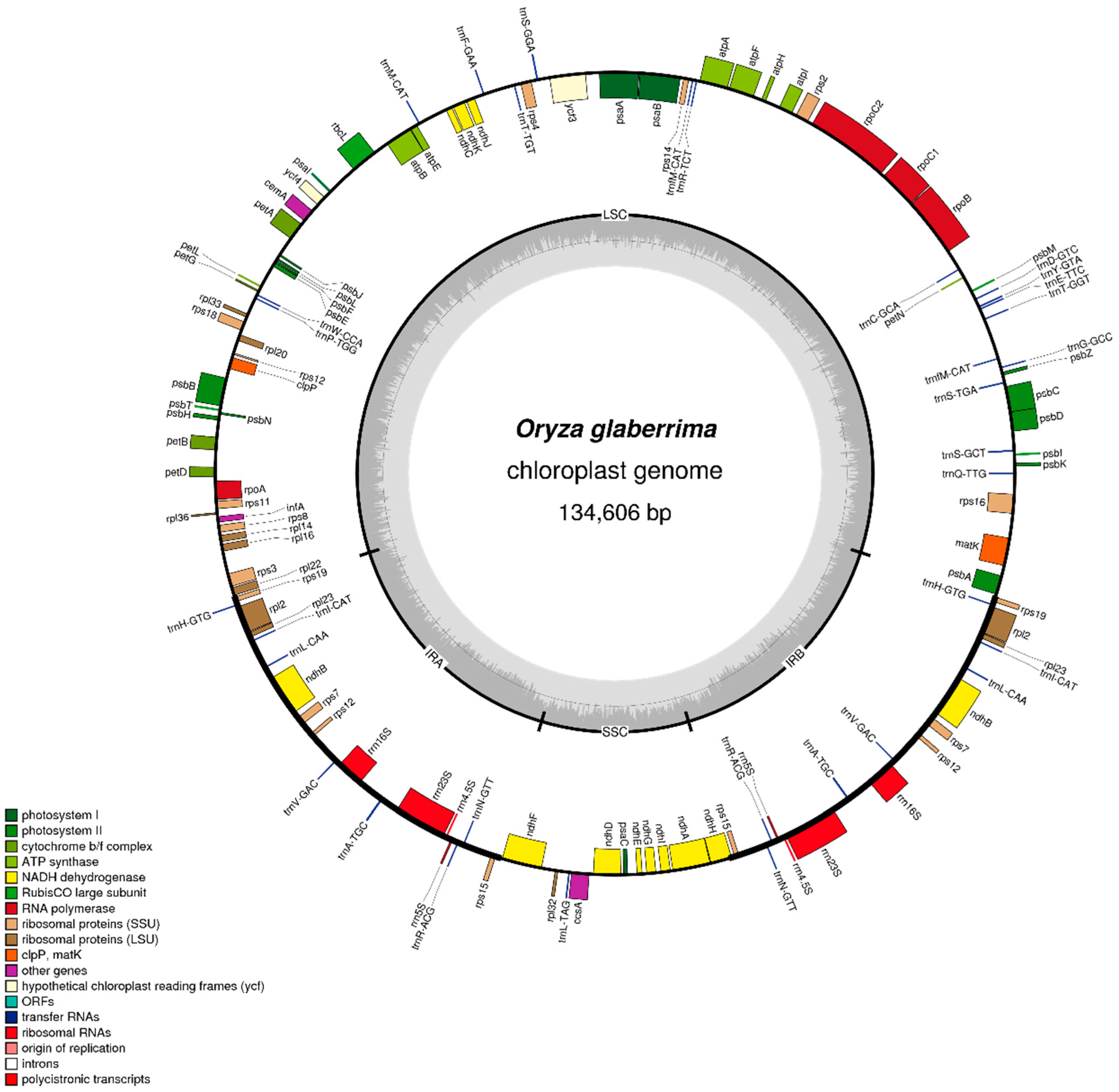
4.2. Chloroplast Genome Sequences
4.3. Transcriptomic Resources
5. Supporting the Conservation and Utilization of African Rice Germplasm Using Genomics
6. Grain Quality and Its Genetic Control
7. Challenges in Deploying African rice Genetic Diversity in Interspecific Breeding
8. Origin and Domestication of African Rice
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Seck, P.A.; Tollens, E.; Wopereis, M.C.S.; Diagne, A.; Bamba, I. Rising trends and variability of rice prices: Threats and opportunities for sub-Saharan Africa. Food Policy 2010, 35, 403–411. [Google Scholar] [CrossRef]
- van Ittersum, M.K.; van Bussel, L.G.J.; Wolf, J.; Grassini, P.; van Wart, J.; Guilpart, N.; Claessens, L.; de Groot, H.; Wiebe, K.; Mason-D’Croz, D.; et al. Can sub-Saharan Africa feed itself? Proc. Natl. Acad. Sci. USA 2016, 113, 14964–14969. [Google Scholar] [CrossRef] [PubMed]
- van Oort, P.A.J.; Zwart, S.J. Impacts of climate change on rice production in Africa and causes of simulated yield changes. Glob. Chang. Biol. 2018, 24, 1029–1045. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.H.; Yu, Y.; Haberer, G.; Marri, P.R.; Fan, C.Z.; Goicoechea, J.L.; Zuccolo, A.; Song, X.; Kudrna, D.; Ammiraju, J.S.S.; et al. The genome sequence of African rice (Oryza glaberrima) and evidence for independent domestication. Nat. Genet. 2014, 46, 982–988. [Google Scholar] [CrossRef] [PubMed]
- Jacquemin, J.; Bhatia, D.; Singh, K.; Wing, R.A. The International Oryza Map Alignment Project: Development of a genus-wide comparative genomics platform to help solve the 9 billion-people question. Curr. Opin. Plant Biol. 2013, 16, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.L.; Sanchez, P.L.; Yu, S.-B.; Lorieux, M.; Eizenga, G.C. Chromosome Segment Substitution Lines: A Powerful Tool for the Introgression of Valuable Genes from Oryza Wild Species into Cultivated Rice (O. sativa). Rice 2010, 3, 218–234. [Google Scholar] [CrossRef]
- Ammiraju, J.S.S.; Luo, M.; Goicoechea, J.L.; Wang, W.; Kudrna, D.; Mueller, C.; Talag, J.; Kim, H.; Sisneros, N.B.; Blackmon, B.; et al. The Oryza bacterial artificial chromosome library resource: Construction and analysis of 12 deep-coverage large-insert BAC libraries that represent the 10 genome types of the genus Oryza. Genome Res. 2006, 16, 140–147. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Meyer, R.S.; Choi, J.Y.; Sanches, M.; Plessis, A.; Flowers, J.M.; Amas, J.; Dorph, K.; Barretto, A.; Gross, B.; Fuller, D.Q.; et al. Domestication history and geographical adaptation inferred from a SNP map of African rice. Nat. Genet. 2016, 48, 1083. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-M.; Chao, D.-Y.; Wu, Y.; Huang, X.; Chen, K.; Cui, L.-G.; Su, L.; Ye, W.-W.; Chen, H.; Chen, H.-C.; et al. Natural alleles of a proteasome α2 subunit gene contribute to thermotolerance and adaptation of African rice. Nat. Genet. 2015, 47, 827. [Google Scholar] [CrossRef]
- Bimpong, I.K.; Serraj, R.; Chin, J.H.; Mendoza, E.M.T.; Hernandez, J.E.; Mendioro, M.S. Determination of genetic variability for physiological traits related to drought tolerance in African rice (Oryza glaberrima). J. Plant Breed. Crop Sci. 2011, 3, 60–67. [Google Scholar]
- Ndjiondjop, M.N.; Wambugu, P.W.; Rodrigues, S.J.; Karlin, G. The effects of drought on rice cultivation in subSaharan Africa and its mitigation: A review. Afr. J. Agric. Res. 2018, 13, 1257–1271. [Google Scholar] [CrossRef]
- Dingkuhn, M.; Audebert, A.Y.; Jones, M.P.; Etienne, K.; Sow, A. Control of stomatal conductance and leaf rolling in O. sativa and O. glaberrima upland rice. Field Crops Res. 1999, 61, 223–236. [Google Scholar] [CrossRef]
- Bimpong, I.K.; Serraj, R.; Chin, J.H.; Ramos, J.; Mendoza, E.M.T.; Hernandez, J.E.; Mendioro, M.S.; Brar, D.S. Identification of QTLs for Drought-Related Traits in Alien Introgression Lines Derived from Crosses of Rice (Oryza sativa cv. IR64) × O. glaberrima under Lowland Moisture Stress. J. Plant Biol. 2011, 54, 237–250. [Google Scholar] [CrossRef]
- Shaibu, A.A.; Uguru, M.I.; Sow, M.; Maji, A.T.; Ndjiondjop, M.N.; Venuprasad, R. Screening African Rice (Oryza glaberrima) for Tolerance to Abiotic Stresses: II. Lowland Drought. Crop Sci. 2018, 58, 133–142. [Google Scholar] [CrossRef]
- Jones, M.P.; Dingkuhn, M.; Aluko/snm, G.K.; Semon, M. Interspecific Oryza Sativa L. × O. glaberrima Steud. progenies in upland rice improvement. Euphytica 1997, 94, 237–246. [Google Scholar] [CrossRef]
- Moukoumbi, Y.D.; Sie, M.; Vodouhe, R.; Bonou, W.; Toulou, B.; Ahanchede, A. Screening of rice varieties for their weed competitiveness. Afr. J. Agric. Res. 2011, 6, 5446–5456. [Google Scholar]
- Fofana, B.; Rauber, R. Weed suppression ability of upland rice under low-input conditions in West Africa. Weed Res. 2000, 40, 271–280. [Google Scholar] [CrossRef]
- Johnson, D.E. The influence of rice plant type on the effect of weed competition on Oryza sativa and Oryza glaberrima. Weed Res. 1998, 38, 207–216. [Google Scholar] [CrossRef]
- Sano, Y.; Sano, R.; Morishima, H. Neighbour Effects Between two Co-Occurring Rice Species, Oryza sativa and O. glaberrima. J. Appl. Ecol. 1984, 21, 245–254. [Google Scholar] [CrossRef]
- Ndjiondjop, M.N.; Seck, P.A.; Lorieux, M.; Futakuchi, K.; Yao, K.N.; Djedatin, G.; Sow, M.E.; Bocco, R.; Cisse, F.; Fatondji, B. Effect of Drought on Oryza glaberrima Rice Accessions and Oryza glaberrima Derived-lines. Asian J. Agric. Res. 2012, 6, 144–157. [Google Scholar] [CrossRef]
- Plowright, R.A.; Coyne, D.L.; Nash, P.; Jones, M.P. Resistance to the rice nematodes Heterodera sacchari, Meloidogyne graminicola and M. incognita in Oryza glaberrima and O. glaberrima × O. sativa interspecific hybrids. Nematology 1999, 1, 745. [Google Scholar] [CrossRef]
- Reversat, G.; Destombes, D. Screening for resistance to Heterodera sacchari in the two cultivated rice species, Oryza sativa and O. glaberrima. Fundam. Appl. Nematol. 1998, 21, 307–317. [Google Scholar]
- Sikirou, M.; Shittu, A.; Konaté, K.A.; Maji, A.T.; Ngaujah, A.S.; Sanni, K.A.; Ogunbayo, S.A.; Akintayo, I.; Saito, K.; Dramé, K.N.; et al. Screening African rice (Oryza glaberrima) for tolerance to abiotic stresses: I. Fe toxicity. Field Crops Res. 2018, 220, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Sahrawat, K.L.; Sika, M. Comparative tolerance of Oryza sativa and Oryza glaberrima cultivars for iron toxicity in West Africa. Int. Rice Res. Notes 2002, 27, 30–31. [Google Scholar]
- Nwilene, F.E.; Adam, A.; Williams, C.T.; Ukwungwu, M.N.; Dakouo, D.; Nacro, S.; Hamadoun, A.; Kamara, S.I.; Okhidievbie, O.; Abamu, F.J. Reactions of differential rice genotypes to African rice gall midge in West Africa. Int. J. Pest Manag. 2002, 48, 195–201. [Google Scholar] [CrossRef]
- Thiémélé, D.; Boisnard, A.; Ndjiondjop, M.-N.; Chéron, S.; Séré, Y.; Aké, S.; Ghesquière, A.; Albar, L. Identification of a second major resistance gene to Rice yellow mottle virus, RYMV2, in the African cultivated rice species, O. glaberrima. Theor. Appl. Genet. 2010, 121, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Ndjiondjop, M.N.; Albar, L.; Fargette, D.; Fauquet, C.; Ghesquiere, A. The genetic basis of high resistance to rice yellow mottle virus (RYMV) in cultivars of two cultivated rice species. Plant Dis. 1999, 83, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Thottappilly, G.; Rossel, H.W. Evaluation of resistance to rice yellow mottle virus in Oryza species. Indian J. Virol. 1993, 9, 65–73. [Google Scholar]
- Albar, L.; Ndjiondjop, M.N.; Esshak, Z.; Berger, A.; Pinel, A.; Jones, M.; Fargette, D.; Ghesquiere, A. Fine genetic mapping of a gene required for Rice yellow mottle virus cell-to-cell movement. Theor. Appl. Genet. 2003, 107, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Djedatin, G.; Ndjiondjop, M.-N.; Mathieu, T.; Cruz, C.M.V.; Sanni, A.; Ghesquière, A.; Verdier, V. Evaluation of African cultivated rice Oryza glaberrima for resistance to bacterial blight. Plant Dis. 2011, 95, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Fujita, D.; Doi, K.; Yoshimura, A.; Yasui, H. A major QTL for resistance to green rice leafhopper (Nephotettix cincticeps Uhler) derived from African rice (Oryza glaberrima Steud.). Breed. Sci. 2010, 60, 336–341. [Google Scholar] [CrossRef][Green Version]
- Futakuchi, K.; Fofana, M.; Sie, M. Varietal Differences in Lodging Resistance of African Rice (Oryza glaberrima Steud.). Asian J. Plant Sci. 2008, 7, 569–573. [Google Scholar] [CrossRef]
- Platten, J.D.; Egdane, J.A.; Ismail, A.M. Salinity tolerance, Na+ exclusion and allele mining of HKT1; 5 in Oryza sativa and O. glaberrima: Many sources, many genes, one mechanism? BMC Plant Biol. 2013, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Linares, O.F. African rice (Oryza glaberrima): History and future potential. Proc. Natl. Acad. Sci. USA 2002, 99, 16360–16365. [Google Scholar] [CrossRef] [PubMed]
- Watarai, M.; Inouye, J. Internode elongation under different rising water conditions in African floating rice (Oryza glaberrima Steud.). J. Fac. of Agric. Kyushu Univ. 1998, 42, 7. [Google Scholar]
- Koide, Y.; Pariasca Tanaka, J.; Rose, T.; Fukuo, A.; Konisho, K.; Yanagihara, S.; Fukuta, Y.; Wissuwa, M.; Yano, M. QTLs for phosphorus deficiency tolerance detected in upland NERICA varieties. Plant Breed. 2013, 132, 259–265. [Google Scholar] [CrossRef]
- Dasgupta, S.; Hossain, M.M.; Huq, M.; Wheeler, D. Climate Change, Soil Salinity, and the Economics of High-Yield Rice Production in Coastal Bangladesh; Policy Research Working Paper; No. WPS 7140; World Bank Group: Washington, DC, USA, 2014. [Google Scholar]
- Kurukulasuriya, P.; Jain, S.; Mahamadou, A.; Mano, R.; Kabubo-Mariara, J.; El-Marsafawy, S.; Molua, E.; Ouda, S.; Ouedraogo, M.; Sene, I.; et al. Will African Agriculture Survive Climate Change? World Bank Econ. Rev. 2006, 20, 367–388. [Google Scholar] [CrossRef]
- Wambugu, P.; Furtado, A.; Waters, D.; Nyamongo, D.; Henry, R. Conservation and utilization of African Oryza genetic resources. Rice 2013, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Dufey, I.; Draye, X.; Lutts, S.; Lorieux, M.; Martinez, C.; Bertin, P. Novel QTLs in an interspecific backcross Oryza sativa × Oryza glaberrima for resistance to iron toxicity in rice. Euphytica 2015. [Google Scholar] [CrossRef]
- Ohmori, Y.; Sotta, N.; Fujiwara, T. Identification of introgression lines of Oryza glaberrima Steud. with high mineral content in grains. Soil Sci. Plant Nutr. 2016, 62, 456–464. [Google Scholar] [CrossRef]
- Petitot, A.-S.; Kyndt, T.; Haidar, R.; Dereeper, A.; Collin, M.; de Almeida Engler, J.; Gheysen, G.; Fernandez, D. Transcriptomic and histological responses of African rice (Oryza glaberrima) to Meloidogyne graminicola provide new insights into root-knot nematode resistance in monocots. Ann. Bot. 2017, 119, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Orjuela, J.; Deless, E.F.T.; Kolade, O.; Cheron, S.; Ghesquiere, A.; Albar, L. A Recessive Resistance to Rice yellow mottle virus Is Associated with a Rice Homolog of the CPR5 Gene, a Regulator of Active Defense Mechanisms. Mol. Plant-Microbe Interact. 2013, 26, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.G.; Carabali, S.J.; Giraldo, O.X.; Martinez, C.P.; Correa, F.; Prado, G.; Tohme, J.; Lorieux, M. Identification of a Rice stripe necrosis virus resistance locus and yield component QTLs using Oryza sativa × O. glaberrima introgression lines. BMC Plant Biol. 2010, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Komeda, N.; Asano, K.; Uehara, K.; Gamuyao, R.; Shim-Angeles, R.B.; Nagai, K.; Doi, K.; Wang, D.R.; Yasui, H.; et al. Convergent Loss of Awn in Two Cultivated Rice Species, (Oryza sativa and Oryza glaberrima) Is Caused by Mutations in Different Loci. Genes Genomes Genet. 2015. [Google Scholar] [CrossRef]
- Zhang, Q.-J.; Zhu, T.; Xia, E.-H.; Shi, C.; Liu, Y.-L.; Zhang, Y.; Liu, Y.; Jiang, W.-K.; Zhao, Y.-J.; Mao, S.-Y.; et al. Rapid diversification of five Oryza AA genomes associated with rice adaptation. Proc. Natl. Acad. Sci. USA 2014, 111, E4954–E4962. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.; Hamada, M.; Kanamori, H.; Namiki, N.; Wu, J.; Itoh, T.; Matsumoto, T.; Sasaki, T.; Ikawa, H.; Tanaka, T.; et al. Distinct evolutionary patterns of Oryza glaberrima deciphered by genome sequencing and comparative analysis. Plant J. 2011, 66, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Hurwitz, B.; Yu, Y.; Collura, K.; Gill, N.; SanMiguel, P.; Mullikin, J.C.; Maher, C.; Nelson, W.; Wissotski, M.; et al. Construction, alignment and analysis of twelve framework physical maps that represent the ten genome types of the genus Oryza. Genome Biol. 2008, 9, R45. [Google Scholar] [CrossRef] [PubMed]
- Wambugu, P.W. Genomic Characterization of African Wild and Cultivated Oryza Species. Ph.D. Thesis, University of Queenland, Brisbane, Australia, 2017. [Google Scholar]
- Monat, C.; Pera, B.; Ndjiondjop, M.-N.; Sow, M.; Tranchant-Dubreuil, C.; Bastianelli, L.; Ghesquière, A.; Sabot, F. De Novo Assemblies of Three Oryza glaberrima Accessions Provide First Insights about Pan-Genome of African Rices. Genome Biol. Evol. 2017, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pariasca-Tanaka, J.; Chin, J.H.; Dramé, K.N.; Dalid, C.; Heuer, S.; Wissuwa, M. A novel allele of the P-starvation tolerance gene OsPSTOL1 from African rice (Oryza glaberrima Steud) and its distribution in the genus Oryza. Theor. Appl. Genet. 2014, 127, 1387–1398. [Google Scholar] [CrossRef] [PubMed]
- Mondal, T.K.; Panda, A.K.; Rawal, H.C.; Sharma, T.R. Discovery of microRNA-target modules of African rice (Oryza glaberrima) under salinity stress. Sci. Rep. 2018, 8, 570. [Google Scholar] [CrossRef] [PubMed]
- Veltman, M.A.; Flowers, J.M.; van Andel, T.R.; Schranz, M.E. Origins and geographic diversification of African rice (Oryza glaberrima). PLoS ONE 2019, 14, e0203508. [Google Scholar] [CrossRef] [PubMed]
- IRGSP. The map-based sequence of the rice genome. Nature 2005, 436, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liang, C.; Chen, C.; Zhang, W.; Sun, S.; Liao, Y.; Zhang, X.; Yang, L.; Song, C.; Wang, M.; et al. Whole-genome sequencing of Oryza brachyantha reveals mechanisms underlying Oryza genome evolution. Nat. Commun. 2013, 4, 1595. [Google Scholar] [CrossRef] [PubMed]
- Stein, J.C.; Yu, Y.; Copetti, D.; Zwickl, D.J.; Zhang, L.; Zhang, C.; Chougule, K.; Gao, D.; Iwata, A.; Goicoechea, J.L.; et al. Genomes of 13 domesticated and wild rice relatives highlight genetic conservation, turnover and innovation across the genus Oryza. Nat. Genet. 2018, 50, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Gore, M.A.; Ross-Ibarra, J.; Ware, D.H.; Buckler, E.S.; Chia, J.-M.; Elshire, R.J.; Sun, Q.; Ersoz, E.S.; Hurwitz, B.L.; Peiffer, J.A.; et al. A first-generation haplotype map of maize. Science 2009, 326, 1115–1117. [Google Scholar] [CrossRef] [PubMed]
- Hansey, C.N.; Vaillancourt, B.; Sekhon, R.S.; de Leon, N.; Kaeppler, S.M.; Buell, C.R. Maize (Zea mays L.) genome diversity as revealed by RNA-sequencing. PLoS ONE 2012, 7, e33071. [Google Scholar] [CrossRef]
- Ndjiondjop, M.-N.; Wambugu, P.; Sangare, J.R.; Dro, T.; Kpeki, B.; Gnikoua, K. Oryza glaberrima Steud. In The Wild Oryza Genomes; Mondal, T.K., Henry, R.J., Eds.; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar]
- Pariasca-Tanaka, J.; Lorieux, M.; He, C.; McCouch, S.; Thomson, M.J.; Wissuwa, M. Development of a SNP genotyping panel for detecting polymorphisms in Oryza glaberrima/O. sativa interspecific crosses. Euphytica 2014. [Google Scholar] [CrossRef]
- Mariac, C.; Sabot, F.; Santoni, S.; Vigouroux, Y.; Couvreur, T.L.P.; Scarcelli, N.; Pouzadou, J.; Barnaud, A.; Billot, C.; Faye, A.; et al. Cost-effective enrichment hybridization capture of chloroplast genomes at deep multiplexing levels for population genetics and phylogeography studies. Mol. Ecol. Resour. 2014, 14, 1103–1113. [Google Scholar] [CrossRef]
- Wambugu, P.W.; Brozynska, M.; Furtado, A.; Waters, D.L.; Henry, R.J. Relationships of wild and domesticated rices (Oryza AA genome species) based upon whole chloroplast genome sequences. Sci. Rep. 2015, 5, 13957. [Google Scholar] [CrossRef]
- Win, K.T.; Yamagata, Y.; Doi, K.; Uyama, K.; Nagai, Y.; Toda, Y.; Kani, T.; Ashikari, M.; Yasui, H.; Yoshimura, A. A single base change explains the independent origin of and selection for the nonshattering gene in African rice domestication. New Phytol. 2017, 213, 1925–1935. [Google Scholar] [CrossRef]
- Nabholz, B.; Sarah, G.; Sabot, F.; Ruiz, M.; Adam, H.; Nidelet, S.; Ghesquière, A.; Santoni, S.; David, J.; Glémin, S. Transcriptome population genomics reveals severe bottleneck and domestication cost in the African rice (Oryza glaberrima). Mol. Ecol. 2014, 23, 2210–2227. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.M.; Zheng, X.M.; Ge, S. Genetic diversity and domestication history of African rice (Oryza glaberrima) as inferred from multiple gene sequences. Theor. Appl. Genet. 2011, 123, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Yıldırım, Y.; Tinnert, J.; Forsman, A. Contrasting patterns of neutral and functional genetic diversity in stable and disturbed environments. Ecol. Evol. 2018, 8, 12073–12089. [Google Scholar] [CrossRef] [PubMed]
- Ta, K.N.; Sabot, F.; Adam, H.; Vigouroux, Y.; De Mita, S.; Ghesquière, A.; Do, N.V.; Gantet, P.; Jouannic, S. miR2118-triggered phased siRNAs are differentially expressed during the panicle development of wild and domesticated African rice species. Rice 2016, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Ganie, S.A.; Debnath, A.B.; Gumi, A.M.; Mondal, T.K. Comprehensive survey and evolutionary analysis of genome-wide miRNA genes from ten diploid Oryza species. BMC Genom. 2017, 18, 711. [Google Scholar] [CrossRef] [PubMed]
- Baldrich, P.; Hsing, Y.-I.C.; San Segundo, B. Genome-Wide Analysis of Polycistronic MicroRNAs in Cultivated and Wild Rice. Genome Biol. Evol. 2016, 8, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Wambugu, P.W.; Ndjiondjop, M.-N.; Henry, R.J. Role of genomics in promoting the utilization of plant genetic resources in genebanks. Brief. Funct. Genom. 2018, 17, 198–206. [Google Scholar] [CrossRef] [PubMed]
- McCouch, S.R.; McNally, K.L.; Wang, W.; Sackville Hamilton, R. Genomics of gene banks: A case study in rice. Am. J. Bot. 2012, 99, 407–423. [Google Scholar] [CrossRef]
- FAO. Second Report on the World’s Plant Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2010; p. 299. [Google Scholar]
- Ndjiondjop, M.N.; Semagn, K.; Zhang, J.; Gouda, A.C.; Kpeki, S.B.; Goungoulou, A.; Wambugu, P.; Dramé, K.N.; Bimpong, I.K.; Zhao, D. Development of species diagnostic SNP markers for quality control genotyping in four rice (Oryza L.) species. Mol. Breed. 2018, 38, 131. [Google Scholar] [CrossRef]
- Orjuela, J.; Sabot, F.; Chéron, S.; Vigouroux, Y.; Adam, H.; Chrestin, H.; Sanni, K.; Lorieux, M.; Ghesquière, A. An extensive analysis of the African rice genetic diversity through a global genotyping. Theor. Appl. Genet. 2014, 127, 2211–2223. [Google Scholar] [CrossRef]
- Singh, N.; Wu, S.; Raupp, W.J.; Sehgal, S.; Arora, S.; Tiwari, V.; Vikram, P.; Singh, S.; Chhuneja, P.; Gill, B.S.; et al. Efficient curation of genebanks using next generation sequencing reveals substantial duplication of germplasm accessions. Sci. Rep. 2019, 650, 410779. [Google Scholar] [CrossRef] [PubMed]
- Wambugu, P.W.; Muthamia, Z.K. Second Report on the State of Plant Genetic Resources for Food and Agriculture in Kenya; FAO: Rome, Italy, 2009. [Google Scholar]
- Semon, M.; Nielsen, R.; Jones, M.P.; McCouch, S.R. The population structure of African cultivated rice Oryza glaberrima (Steud.): Evidence for elevated levels of linkage disequilibrium caused by admixture with O. sativa and ecological adaptation. Genetics 2005, 169, 1639–1647. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.J.; Lee, J.K.; Hong, S.W.; Park, Y.J.; McNally, K.L.; Kim, N.S. Genetic diversity and phylogenetic relationship in AA Oryza species as revealed by Rim2/Hipa CACTA transposon display. Genes Genet. Syst. 2006, 81, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Ndjiondjop, M.-N.; Semagn, K.; Gouda, A.C.; Kpeki, S.B.; Dro Tia, D.; Sow, M.; Goungoulou, A.; Sie, M.; Perrier, X.; Ghesquiere, A.; et al. Genetic Variation and Population Structure of Oryza glaberrima and Development of a Mini-Core Collection Using DArTseq. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Schlötterer, C.; Tobler, R.; Kofler, R.; Nolte, V. Sequencing pools of individuals-mining genome-wide polymorphism data without big funding. Nat. Rev. Genet. 2014, 15, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Wang, P.; Xu, Y. Bulked sample analysis in genetics, genomics and crop improvement. Plant Biotechnol. J. 2016, 14, 1941–1955. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Lv, P.; Hou, S.; Li, S.; Ji, G.; Ma, X.; Du, R.; Liu, G. Combining Next Generation Sequencing with Bulked Segregant Analysis to Fine Map a Stem Moisture Locus in Sorghum (Sorghum bicolor L. Moench): e0127065. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Ramirez-Gonzalez, R.H.; Segovia, V.; Bird, N.; Fenwick, P.; Holdgate, S.; Berry, S.; Jack, P.; Caccamo, M.; Uauy, C. RNA-Seq bulked segregant analysis enables the identification of high-resolution genetic markers for breeding in hexaploid wheat. Plant Biotechnol. J. 2015, 13, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Olasanmi, B.; Akoroda, M.O.; Okogbenin, E.; Egesi, C.; Kahya, S.S.; Onyegbule, O.; Ewa, F.; Guitierrez, J.; Ceballos, H.; Tohme, J.; et al. Bulked segregant analysis identifies molecular markers associated with early bulking in cassava (Manihot esculenta Crantz). Euphytica 2014, 195, 235–244. [Google Scholar] [CrossRef]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S.; et al. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Wambugu, P.; Ndjiondjop, M.-N.; Furtado, A.; Henry, R. Sequencing of bulks of segregants allows dissection of genetic control of amylose content in rice. Plant Biotechnol. J. 2018, 16, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Falade, K.O.; Semon, M.; Fadairo, O.S.; Oladunjoye, A.O.; Orou, K.K. Functional and physico-chemical properties of flours and starches of African rice cultivars. Food Hydrocoll. 2014, 39, 41–50. [Google Scholar] [CrossRef]
- Gayin, J.; Bertoft, E.; Manful, J.; Yada, R.Y.; Abdel-Aal, E.S.M. Molecular and thermal characterization of starches isolated from African rice (Oryza glaberrima). Starch-Stärke 2016, 68, 9–19. [Google Scholar] [CrossRef]
- Gayin, J.; Chandi, G.K.; Manful, J.; Seetharaman, K. Classification of Rice Based on Statistical Analysis of Pasting Properties and Apparent Amylose Content: The Case of Oryza glaberrima Accessions from Africa. Cereal Chem. 2015, 92, 22–28. [Google Scholar] [CrossRef]
- Gayin, J.; Abdel-Aal, E.-S.M.; Manful, J.; Bertoft, E. Unit and internal chain profile of African rice (Oryza glaberrima) amylopectin. Carbohydr. Polym. 2016, 137, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Gayin, J. Structural and Functional Characteristics of African Rice (Oryza glaberrima) Flour and Starch. Ph.D. Thesis, The University of Guelph, Guelph, ON, Canada, 2015. [Google Scholar]
- Wang, K.; Wambugu, P.W.; Zhang, B.; Wu, A.C.; Henry, R.J.; Gilbert, R.G. The biosynthesis, structure and gelatinization properties of starches from wild and cultivated African rice species (Oryza barthii and Oryza glaberrima). Carbohydr. Polym. 2015, 129, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Vargas, C.G.; Costa, T.M.H.; de Rios, A.O.; Flôres, S.H. Comparative study on the properties of films based on red rice (Oryza glaberrima) flour and starch. Food Hydrocoll. 2017, 65, 96–106. [Google Scholar] [CrossRef]
- Manful, J.T.; Graham-Acquaah, S. African Rice (Oryza glaberrima): A Brief History and Its Growing Importance in Current Rice Breeding Efforts. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Koide, Y.; Ogino, A.; Yoshikawa, T.; Kitashima, Y.; Saito, N.; Kanaoka, Y.; Onishi, K.; Yoshitake, Y.; Tsukiyama, T.; Saito, H.; et al. Lineage-specific gene acquisition or loss is involved in interspecific hybrid sterility in rice. Proc. Natl. Acad. Sci. USA 2018, 115, E1955–E1962. [Google Scholar] [CrossRef]
- Koide, Y.; Onishi, K.; Nishimoto, D.; Baruah, A.R.; Kanazawa, A.; Sano, Y. Sex-independent transmission ratio distortion system responsible for reproductive barriers between Asian and African rice species. New Phytol. 2008, 179, 888–900. [Google Scholar] [CrossRef]
- Xie, Y.; Xu, P.; Huang, J.; Ma, S.; Xie, X.; Tao, D.; Chen, L.; Liu, Y.G. Interspecific Hybrid Sterility in Rice Is Mediated by OgTPR1 at the S1 Locus Encoding a Peptidase-like Protein. Mol. Plant 2017, 10, 1137–1140. [Google Scholar] [CrossRef]
- Lorieux, M.; Garavito, A.; Bouniol, J.; Gutiérrez, A.; Ndjiondjop, M.N.; Guyot, R.; Pompilio Martinez, C.; Tohme, J.; Ghesquière, A. Unlocking the Oryza glaberrima Treasure for Rice Breeding in Africa; CABI: Wallingford, UK, 2013; pp. 130–143. [Google Scholar]
- Garavito, A.; Guyot, R.; Lozano, J.; Gavory, F.; Samain, S.; Panaud, O.; Tohme, J.; Ghesquière, A.; Lorieux, M. A genetic model for the female sterility barrier between Asian and African cultivated rice species. Genetics 2010, 185, 1425–1440. [Google Scholar] [CrossRef][Green Version]
- Nayar, N.M. Origin and Cytogenetics of Rice. In Advances in Genetics; Caspari, E.W., Ed.; Academic Press: Amsterdam, Netherlands, 1973; Volume 17, pp. 153–292. [Google Scholar]
- Nayar, N.M. Evolution of the African Rice: A Historical and Biological Perspective. Crop Sci. 2012, 52, 505–516. [Google Scholar] [CrossRef]
- Nayar, N.M. The history and genetic transformation of the African rice, Oryza glaberrima Steud. (Gramineae). Curr. Sci. 2010, 99, 1681–1689. [Google Scholar]
- Nayar, N.M. Chapter 5—The Origin of African Rice. In Origin and Phylogeny of Rices; Nayar, N.M., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 117–168. [Google Scholar]
- Choi, J.Y.; Zaidem, M.; Gutaker, R.; Dorph, K.; Singh, R.K.; Purugganan, M.D. The complex geography of domestication of the African rice Oryza glaberrima. PLoS Genet. 2019, 15, e1007414. [Google Scholar] [CrossRef] [PubMed]
- Cubry, P.; Tranchant-Dubreuil, C.; Thuillet, A.C.; Monat, C.; Ndjiondjop, M.N.; Labadie, K.; Cruaud, C.; Engelen, S.; Scarcelli, N.; Rhone, B.; et al. The Rise and Fall of African Rice Cultivation Revealed by Analysis of 246 New Genomes. Curr. Biol. 2018, 28, 2274–2282. [Google Scholar] [CrossRef] [PubMed]
- Porteres, R. Berceaux Agricoles Primaires Sur le Continent Africain. J. Afr. Hist. 1962, 3, 195–210. [Google Scholar] [CrossRef]
- Hu, M.; Lv, S.; Wu, W.; Fu, Y.; Liu, F.; Wang, B.; Li, W.; Gu, P.; Cai, H.; Sun, C.; et al. The domestication of plant architecture in African rice. Plant J. 2018, 94, 661–669. [Google Scholar] [CrossRef]
- Wambugu, P.W.; Nyamongo, D.; Ndjiondjop, M.-N.; Henry, R.J. Evolutionary Relationships Among the Oryza Species. In The Wild Oryza Genomes; Mondal, T.K., Henry, R.J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 41–54. [Google Scholar]
- Gross, B.L.; Steffen, F.T.; Olsen, K.M. The molecular basis of white pericarps in African domesticated rice: Novel mutations at the Rc gene. J. Evol. Biol. 2010, 23, 2747–2753. [Google Scholar] [CrossRef]
- Wu, W.; Liu, X.; Wang, M.; Meyer, R.S.; Luo, X.; Ndjiondjop, M.-N.; Tan, L.; Zhang, J.; Wu, J.; Cai, H.; et al. A single-nucleotide polymorphism causes smaller grain size and loss of seed shattering during African rice domestication. Nat. Plants 2017, 3, 17064. [Google Scholar] [CrossRef]
- Lv, S.; Wu, W.; Wang, M.; Meyer, R.S.; Ndjiondjop, M.N.; Tan, L.; Zhou, H.; Zhang, J.; Fu, Y.; Cai, H.; et al. Genetic control of seed shattering during African rice domestication. Nat. Plants 2018, 4, 331–337. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, S.; Li, X.; Zhang, B.; Jiang, L.; Tang, Y.; Zhao, J.; Ma, X.; Cai, H.; Sun, C.; et al. Deletions linked to PROG1 gene participate in plant architecture domestication in Asian and African rice. Nat. Commun. 2018, 9, 4157. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Trait | Reference |
|---|---|
| Weed competitiveness | [15,16,17,18] |
| Drought tolerance | [15,19,20] |
| Resistance to nematodes | [21,22] |
| Resistance to iron toxicity | [23,24] |
| Resistance to African gall midge | [25] |
| Resistance to Rice Yellow Mortal Virus | [26,27,28,29] |
| Resistance to bacterial leaf blight (BLB) | [30,31] |
| Tolerance to lodging | [15,32] |
| Resistance to green rice leafhopper (Nephotettix cincticeps Uhler) | [31] |
| Tolerance to salinity | [8,33,34] |
| Tolerance to soils acidity | [19] |
| Tolerance to submergences | [35] |
| Species | Feature | Reference | |
|---|---|---|---|
| Genome Size | Gene Count | ||
| O. glaberrima | 316 Mb | 33,164 | [4] |
| O. sativa | 370 Mb | 37,544 | [54] |
| O. brachyantha | 261 Mb | 32,038 | [55] |
| O. barthii | 308 Mb | 34,575 | [56] |
| O. meridionalis | 336 Mb | 29,308 | [56] |
| O. punctata | 394 Mb | 31,679 | [56] |
| O. glumaepatula | 373 Mb | 35,674 | [56] |
| Feature | CG14 (I-OMAP) | CG14 | TOG5681 | G22 |
|---|---|---|---|---|
| Assembly size | 316 Mb | 299 Mb | 292 Mb | 305 Mb |
| Gene count | 33,164 | 50,000 | 51,262 | 49,662 |
| Scaffold N50 | 217 kb | 10 kb | 13 kb | 14 kb |
| Sequencing platform | Roche/454 GS-FLX Titanium Sequencing and Sanger | Illumina | Illumina | Illumina |
| Assembly approach | Reads aligned to O. sativa refseq | De novo | De novo | De novo |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wambugu, P.W.; Ndjiondjop, M.-N.; Henry, R. Advances in Molecular Genetics and Genomics of African Rice (Oryza glaberrima Steud). Plants 2019, 8, 376. https://doi.org/10.3390/plants8100376
Wambugu PW, Ndjiondjop M-N, Henry R. Advances in Molecular Genetics and Genomics of African Rice (Oryza glaberrima Steud). Plants. 2019; 8(10):376. https://doi.org/10.3390/plants8100376
Chicago/Turabian StyleWambugu, Peterson W., Marie-Noelle Ndjiondjop, and Robert Henry. 2019. "Advances in Molecular Genetics and Genomics of African Rice (Oryza glaberrima Steud)" Plants 8, no. 10: 376. https://doi.org/10.3390/plants8100376
APA StyleWambugu, P. W., Ndjiondjop, M.-N., & Henry, R. (2019). Advances in Molecular Genetics and Genomics of African Rice (Oryza glaberrima Steud). Plants, 8(10), 376. https://doi.org/10.3390/plants8100376