A Novel Aspect of Essential Oils: Coating Seeds with Thyme Essential Oil induces Drought Resistance in Wheat

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Results
2.1. The Effects of Seed Coating with Thyme Oil on Seed Germination and the Root and Shoot Development of Wheat Seedlings
2.2. Stress Imposition
2.3. The Effects of Seed Coating with Thyme Oil on Root and Shoot Development, Normalized Difference Vegetation Index, Carbon and Nitrogen Content, and the Stable Isotopic Compositions 13C and 15N, under Well Watered and Water/Nutrient Stress Conditions
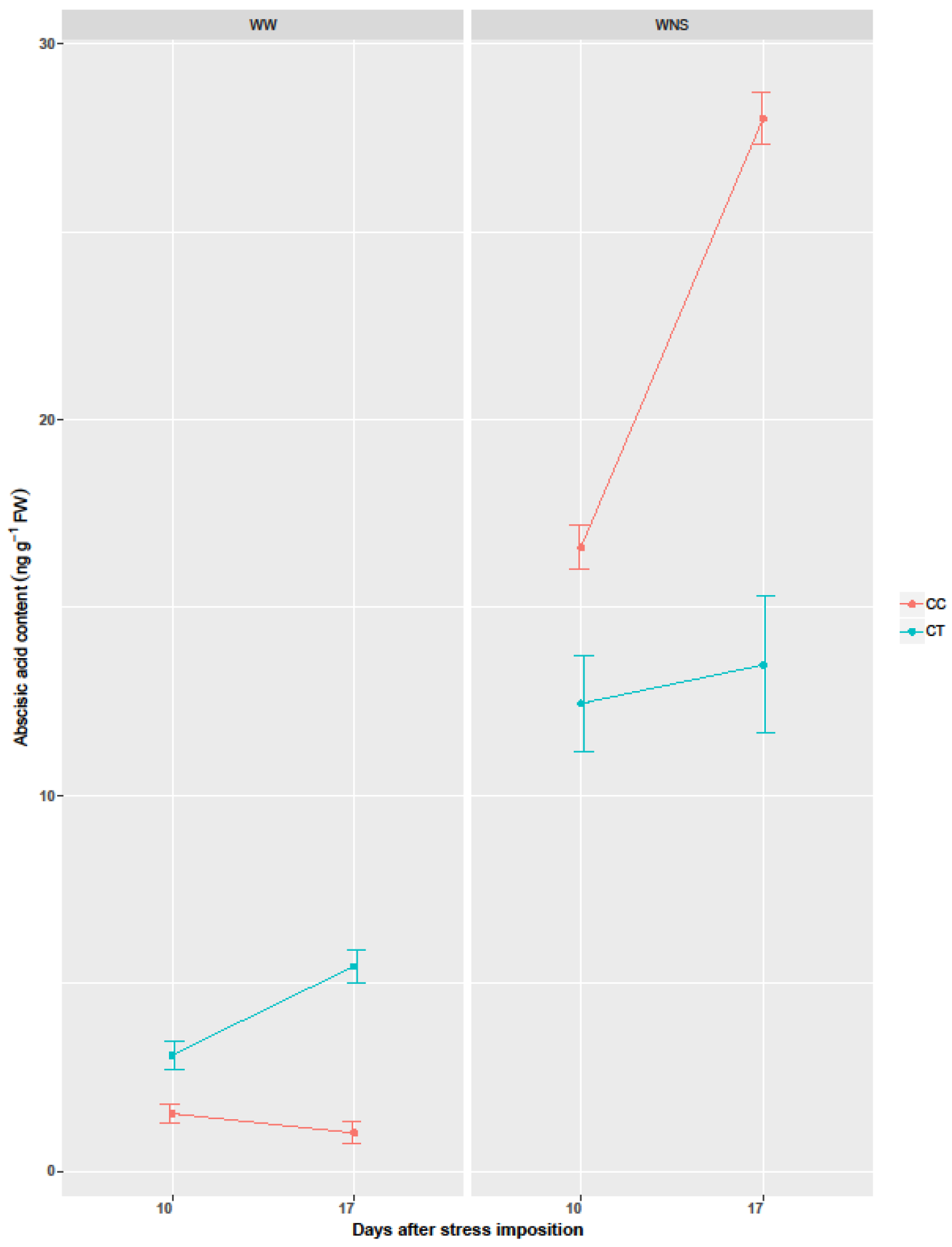
2.4. The Effect of Coating with Thyme Oil on ABA Content under Well-Watered and Water/Nutrient Stress Conditions
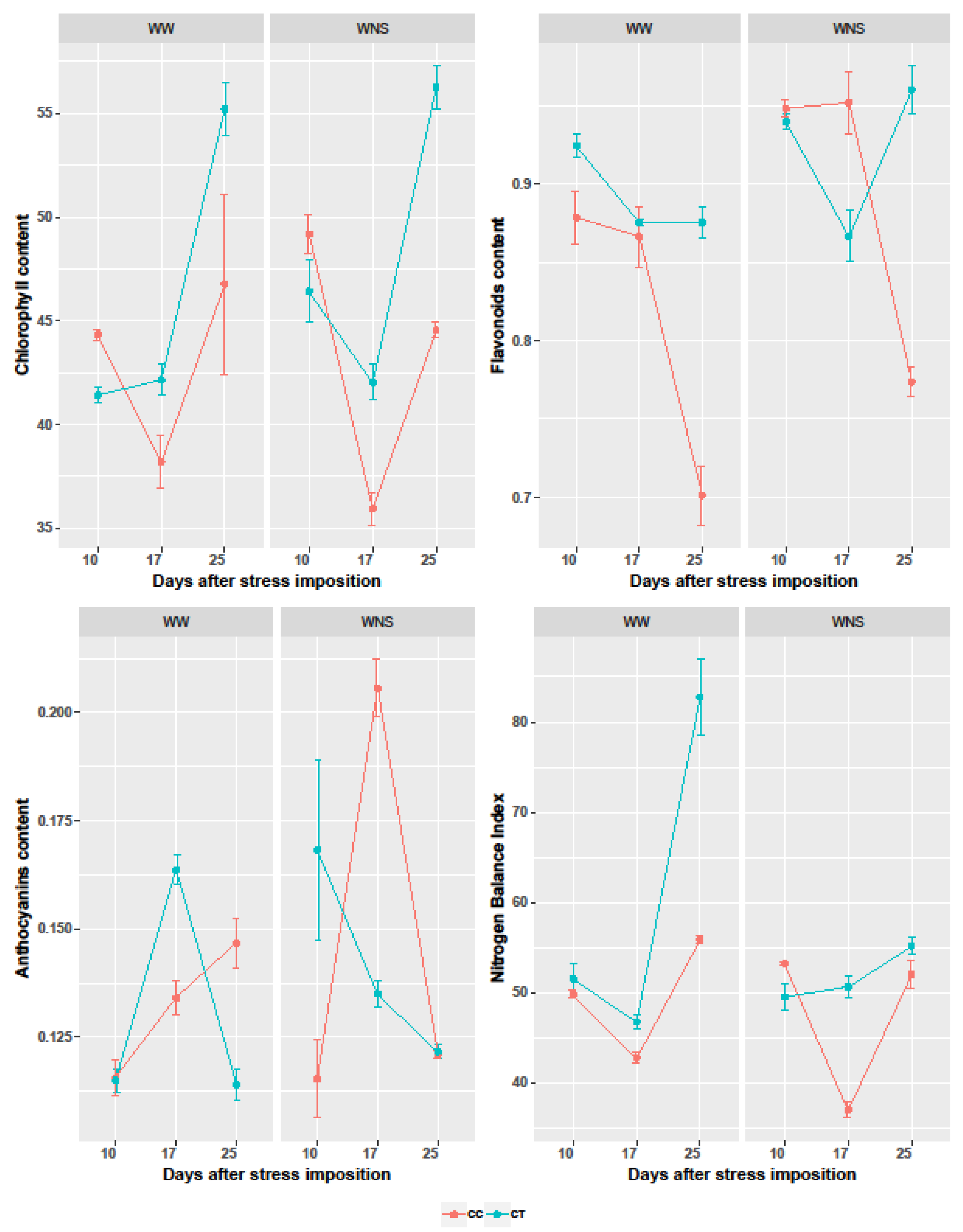
2.5. The Effect of Coating with Thyme Oil on Plant Response under Well-Watered and Water/Nutrient Stress Conditions Measured with Dualex
3. Discussion
3.1. The Effects of Thyme Oil on Seed Germination, Seedling Development, and Plant Physiology under Well-Watered Conditions
3.2. The Effect of Water/Nutrient Stress Imposition in Control Plants
3.3. The Effect of Thyme Oil on Water/Nutrient Stress Resistance
4. Material and Methods
4.1. Plant Material
4.2. Seed Coating Treatment
4.3. Seed Germination Assay
4.4. The Pot Experiment
4.4.1. Growth Conditions
4.4.2. Irrigation and Nutritive Supply Conditions
4.4.3. Sampling
4.4.4. Leaf Pigments
4.4.5. ABA Analysis
4.4.6. Shoot Total Carbon and Nitrogen, and Stable Carbon and Nitrogen Isotope Composition
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Curtis, B.C. Wheat in the World. In Bread Wheat Improvement and Production; Curtis, B.C., Rajaram, S., Macpherson, H.G., Eds.; Plant Production Protectio Series; FAO: Roma, Italy, 2002; Volume 30, pp. 1–18. [Google Scholar]
- Araus, J.L. The problem of sustainable water use in the Mediterranean and research requirements for agriculture. Ann. Appl. Biol. 2004, 144, 259–272. [Google Scholar] [CrossRef]
- Boguszewska, D.; Zagdanska, B. ROS as signalling molecules and enzymes of plant response to unfavorable environmental conditions, oxidative stress-molecular mechanisms and biological effects. In Oxidative Stress—Molecular Mechanisms and Biological Effects, 1st ed.; Lushchak, V., Semchyshyn, H.M., Eds.; InTech: Janeza Trdine, Croatia, 2012; pp. 341–362. [Google Scholar]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed]
- Parry, M.L.; Rosenzweig, C.; Iglesias, A.; Livermore, M.; Fischer, G. Effects of climate change on global food production under SRES emissions and socio–economic scenarios. Glob. Environ. Chang. 2004, 4, 53–67. [Google Scholar] [CrossRef]
- Tavares, L.C.; Rufino, C.A.; Brunes, A.P.; Friedrich, F.F.; Barros, A.C.S.A.; Villela, F.A. Physiological performance of wheat seeds coated with micronutrients. J. Seed Sci. 2013, 35, 28–345. [Google Scholar] [CrossRef]
- Lutts, S.; Benincasa, P.; Wojtyla, L.; Kubala, S.; Pace, R.; Lechowska, K.; Quinet, M.; Garnczarska, M. Seed priming: New comprehensive approaches for an old empirical technique. In New Challenges in Seed Biology-Basic and Translational Research Driving Seed Technology, 1st ed.; Araujo, S., Balestrazzi, A., Eds.; InTech Open: Rijeka, Croatia, 2016; pp. 1–47. [Google Scholar]
- Kaufman, G. Seed coating: A tool for stand establishment; A stimulus to seed quality. HortTechnology 1991, 1, 98–102. [Google Scholar] [CrossRef]
- Porter, F.E.; Scott, J.M. Plant Seed Coating. U.S. Patent 4,251,952, 24 February 1981. [Google Scholar]
- Ben Jabeur, M.; Ghabri, E.; Myriam, M.; Hamada, W. Thyme essential oil as a defense inducer of tomato against graymold and Fusarium wilt. Plant Phys. Biochem. 2015, 94, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Medina, S.; Vicente, R.; Amador, A.; Araus, J.L. Interactive Effects of Elevated [CO2] and Water Stress on Physiological Traits and Gene Expression during Vegetative Growth in Four Durum Wheat Genotypes. Front. Plant Sci. 2016, 7, 17–38. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Bachmann, K.A.; Bailer, A.J.; Bolger, P.M.; Borak, J.; Cai, L.; Cedergreen, N.; Cherian, M.G.; Chiueh, C.C.; Clarkson, T.W.; et al. Biological stress response terminology: Integrating the concepts of adaptive response and preconditioning stress within a hormetic dose–response framework. Toxicol. Appl. Pharmacol. 2007, 222, 122–128. [Google Scholar] [CrossRef]
- Rentzsch, S.; Podzimska, D.; Voegele, A.; Imbeck, M.; Müller, K.; Linkies, A.; Leubner-Metzger, G. Dose- and tissue-specific interaction of monoterpenes with the gibberellin-mediated release of potato tuber bud dormancy, sprout growth and induction of α-amylases and β-amylases. Planta 2011, 235, 137–151. [Google Scholar] [CrossRef]
- Dudai, N.; Ben-Ami, M.; Chaimovich, R.; Chaimovitsh, D. Essential oils as allelopathic agents: Bioconversion of monoterpenes by germinating wheat seeds. Acta Hort. 2004, 629, 505–508. [Google Scholar] [CrossRef]
- Mathieu, Y.; Lapous, D.; Thomine, S.; Laurière, C.; Guern, J. Cytoplasmic acidification as an early phosphorylation-dependent response of tobacco cells to elicitors. Planta 1996, 199, 416–424. [Google Scholar] [CrossRef]
- He, D.Y.; Yazaki, Y.; Nishizawa, Y.; Takai, R.; Yamada, K.; Sakano, K.; Shibuya, N.; Minami, E. Gene Activation by cytoplasmic acidification in suspension-cultured rice cells in response to the potent elicitor, N-Acetylchitoheptaose. Mol. Plant Micro Interact. 1998, 11, 1167–1174. [Google Scholar] [CrossRef]
- Blankenship, J.O.; Smith, D.R. Breaking seed dormancy in: Parry’s clover by acid treatment. J. Range Manag. 1967, 20, 50. [Google Scholar] [CrossRef]
- Benno, B.; Hubert, F.; Roger, W.P. Evidence that acid solutions induce plant cell elongation by acidifying the cytosol and stimulating the proton pump. Fed. Eur. Biochem. Soc. 1984, 174, 223–227. [Google Scholar] [CrossRef]
- Hamabata, A.; Garcia-Maya, M.; Romero, T.; Bernal-Lugo, I. Kinetics of the Acidification Capacity of Aleurone Layer and Its Effect upon Solubilization of Reserve Substances from Starchy Endosperm of Wheat. Plant Physiol. 1988, 86, 643–644. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, A.I.; Vidaver, W.; Quick, W.A. Acidification, growth promoter, and red light effects on germination of skotodormant lettuce seeds (Lactuca sativa). Canad. J. Bot. 1984, 6, 1108–1115. [Google Scholar] [CrossRef]
- Jung, J.K.H.; McCouch, S. Getting to the roots of it: Genetic and hormonal control of root architecture. Front. Plant Sci. 2013, 4, 186. [Google Scholar] [CrossRef]
- Zingaretti, S.M.; Inácio, M.C.; Pereira, L.D.M.; Paz, T.A.; França, S.D.C. Water stress and agriculture. In Responses of Organisms to Water Stress, 1st ed.; Akinci, S., Ed.; InTech: Lexington, KY, USA, 2013. [Google Scholar]
- Araus, J.L.; Slafer, G.A.; Reynolds, M.P.; Royo, C. Plant breeding and drought in C3 cereals: What should we breed for? Ann. Bot. 2002, 89, 925–940. [Google Scholar] [CrossRef]
- Araus, J.L.; Villegas, D.; Aparicio, N.; Del Moral, L.F.; El Hani, S.; Rharrabti, Y.; Ferrio, J.P.; Royo, C. Environmental factors determining carbon isotope discrimination and yield in durum wheat under Mediterranean conditions. Crop Sci. 2003, 43, 170–180. [Google Scholar] [CrossRef]
- Hubick, K.T.; Farquhar, G.D.; Shorter, R. Correlation between water-use efficiency and carbon isotope discrimination in diverse peanut (Arachis) germplasm. Funct. Plant Biol. 1986, 13, 803–808. [Google Scholar] [CrossRef]
- Sanchez-Bragado, R.; Serret, M.D.; Araus, J.L. The nitrogen contribution of different plant parts to wheat grains: Exploring genotype, water, and nitrogen effects. Front. Plant Sci. 2017, 7, 19–86. [Google Scholar] [CrossRef] [PubMed]
- Steyn, W.; Wand, S.; Holcroft, D.; Jacobs, G. Anthocyanins in vegetative tissues: A proposed unified function in photoprotection. New Phytol. 2002, 155, 349–361. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Goulas, Y.; Cerovic, Z.G.; Cartelat, A.; Moya, I. Dualex: A new instrument for field measurements of epidermal UV-absorbance by chlorophyll fluorescence. Appl. Optics 2004, 43, 4488–4496. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, S.; Sun, H.; Wang, Y.; Shao, L. Root size, distribution and soil water depletion as affected by cultivars and environmental factors. Field Crops Res. 2009, 114, 75–83. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Belford, R.K.; Tennant, D. Root: Shoot ratios of old and modern, tall and semi-dwarf wheats in a Mediterranean environment. Plant Soil 1990, 121, 89–98. [Google Scholar] [CrossRef]
- Song, L.; Li, F.M.; Fan, X.W.; Xiong, Y.C.; Wang, W.Q.; Wu, X.B.; Turner, N.C. Soil water availability and plant competition affect the yield of spring wheat. Eur. J. Agron. 2009, 31, 51–60. [Google Scholar] [CrossRef]
- Ghashghaie, J.; Tcherkez, G. Isotope Ratio Mass Spectrometry Technique to Follow Plant Metabolism: Principles and Applications of 12C/13C Isotopes. In Advances in Botanical Research: Metabolomics Coming of Age With Its Technological Diversity, 1st ed.; Rolin, D., Ed.; Academic Press, Elsevier: London, UK, 2013; Volume 67, pp. 377–405. [Google Scholar]
- Farquhar, G.D.; Wetselaar, R.; Weir, B. Gaseous nitrogen losses from plants. In Gaseous Loss of Nitrogen from Plant-Soil Systems; Springer: Dordrecht, The Netherlands, 1983; pp. 159–180. [Google Scholar]
- Takahama, U.; Oniki, T. Flavonoids and some other phenolics as substrates of peroxidase: Physiological significance of the redox reactions. J. Plant Res. 2000, 113, 301–309. [Google Scholar] [CrossRef]
- Marshall, J.D.; Brooks, J.R.; Lajtha, K. Sources of variation in the stable isotopic composition of plants. In Stable Isotopes in Ecology and Environmental Science, 2nd ed.; Michener, R., Lajtha, K., Eds.; Wiley-Blackwell: Malden, MA, USA, 2007; pp. 22–60. [Google Scholar]
- Vessey, J.K.; Henry, L.T.; Chaillou, S.; Raper, C.D., Jr. Root-zone acidity affects relative uptake of nitrate and ammonium from mixed nitrogen sources. J. Plant Nutr. 1990, 13, 95–116. [Google Scholar] [CrossRef] [PubMed]
- Fiche de Données de Sécurité: Agicote Rouge Cereals; Quick-FDS [18207-41570-12577-010076]; Aegilops Applications, Val de Reuil, France. 5 December 2017. Available online: http://www.utdf.coop/alfresco/service/api/node/content/workspace/SpacesStore/ec008e4e-1c48-4401-9a42-6df5cffcd4cb/101098.pdf?guest=true (accessed on 16 February 2018).
- Thameur, A.; Ferchichi, A.; López-Carbonell, M. Involvement of abscisic acid metabolites and the oxidative status of barley genotypes in response to drought. Can. J. Plant Sci. 2014, 94, 1481–1490. [Google Scholar] [CrossRef]
- Coplen, T.B. Explanatory Glossary of Terms Used in Expression of Relative Isotope Ratios and Gas Ratios. IUPAC Provisional Recommendations. Inorganic Chemistry Division. Comission on Isotopic Abundances and Atomic Weights 2008. Available online: http://old.iupac.org/reports/provisional/abstract08/coplen_prs.Pdf (accessed on 25 January 2019).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Germination (%) | Shoot Dry Weight (g) | Root Dry Weight (g) |
| CC | 76.16 b ± 1.25 | 0.76 b ± 0.016 | 0.16 b ± 0.011 |
| CT | 100.00 a ± 0.0 | 2.44 a ± 0.028 | 0.34 a ± 0.012 |
| ANOVA Coating | 152.4 *** | 3289 *** | 182.1 *** |
| Traits | Shoot Length (cm) | Root Length (cm) | Root/Shoot Ratio |
| CC | 34.33 b ± 0.57 | 19.16 b ± 0.28 | 0.21 a ± 0.01 |
| CT | 38.33 a ± 0.57 | 41.66 a ± 1.52 | 0.14 b ± 0.003 |
| ANOVA Coating | 54.75 *** | 323.3 *** | 63.38 *** |
| Water Content (%) | Nutritive Supply (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Days after water stress started | 6 | 10 | 12 | 14 | 17 | 25 | 6 | 10 | 12 | 14 | 17 | 25 |
| Pots containing seeds coated with water (control) | 81.0 | 60.9 | 41.3 | 32.5 | 31.0 | 30.6 | 40.5 | 30.4 | 20.6 | 16.2 | 15.5 | 15.3 |
| Pots containing seeds coated with thyme oil | 80.3 | 60.3 | 40.4 | 30.7 | 30.1 | 30.3 | 40.1 | 30.1 | 20.2 | 15.3 | 15.05 | 15.1 |
| Average decrease (%) | 19.3 | 39.4 | 59.1 | 68.4 | 69.4 | 69.6 | 9.7 | 19.7 | 29.6 | 34.2 | 34.7 | 34.8 |
| Traits | Shoot Dry Weight (mg) | Root Dry Weight (mg) | NDVI | Root to Shoot Ratio | ||||||||
| Coating | WW | WNS | RR (%) | WW | WNS | RR (%) | WW | WNS | RR (%) | WW | WNS | RR (%) |
| CC | 35.89 b ± 3.83 | 16.04 d ± 0.57 | 55.3% | 31.28 b ± 0.72 | 9.05 d ± 0.22 | 71% | 0.16 c ± 0.00 | 0.14 d ± 0.005 | 12.5% | 0.879 a ± 0.1 | 0.565 b ± 0.03 | 35.7% |
| CT | 44.94 a ± 0.38 | 23.40 c ± 1.42 | 47.9% | 44.32 a ± 0.68 | 21.51 c ± 0.94 | 51.4% | 0.21 a ± 0.005 | 0.19 b ± 0.005 | 9.52% | 0.986 a ± 0.01 | 0.921 a ± 0.06 | 6.59% |
| ANOVA | ||||||||||||
| Coating (C) | 47.008 *** | 1006.094 *** | 300 *** | 23.182 *** | ||||||||
| Irrigation + nutrient supply (I + N) | 298.811 *** | 3138.739 *** | 33.333 *** | 11.870 ** | ||||||||
| (T × (I + N)) | 0.498 n.s. | 0.509 n.s. | 1.333 n.s. | 6.521 * | ||||||||
| Traits | Shoot N content (DM, %) | Shoot δ15N (‰) | Shoot C content (DM, %) | Shoot δ13C (‰) | ||||||||
| Coating | WW | WNS | IR (%) | WW | WNS | IR/RR (%) | WW | WNS | IR/RR (%) | WW | WNS | IR (%) |
| CC | 3.54 b ± 0.08 | 3.74 b ± 0.11 | 5.34% | 5.06 b ± 0.26 | 4.32 c ± 0.07 | RR 14.6% | 47.38 a ± 2.1 | 40.88 b ± 0.58 | RR 13.7% | 30.77 c ± 0.05 | −29.80 a ± 0.08 | 3.15% |
| CT | 2.87 c ± 0.14 | 4.25 a ± 0.15 | 32.2% | 4.16 c ± 0.29 | 5.86 a ± 0.07 | IR 29% | 42.46 b ± 0.16 | 45.15 a ± 1.67 | IR 5.9% | 30.00 b ± 0.05 | −29.68 a ± 0.12 | 1.06% |
| ANOVA | ||||||||||||
| Coating (C) | 1.155 n.s. | 7.487 * | 0.168 n.s. | 86.58 *** | ||||||||
| Irrigation + nutrient supply (I + N) | 116.302 *** | 17.065 ** | 5.729 * | 187.56 *** | ||||||||
| (T × (I + N)) | 64.311 *** | 108.959 *** | 33.363 *** | 46.16 *** | ||||||||
| Variables | Coating (C) | Irrigation + Nutrient Supply (I + N) | Days Post Stress (dps) | C × (I + N) | C × dps | (I + N) × dps | C × (I + N) × dps |
|---|---|---|---|---|---|---|---|
| ABA | 77.49 *** | 1691.9 *** | 97.72 *** | 291.59 *** | 26.98 *** | 53.65 *** | 84.18 *** |
| Traits | Chlorophyll | Flavonoids | Anthocyanins | NBI |
|---|---|---|---|---|
| Coating (C) | 63.291 *** | 141.53 *** | 1.91 n.s. | 207.33 *** |
| Irrigation + Nutrient supply (I + N) | 4.185 n.s. | 138.03 *** | 27.22 *** | 101.57 *** |
| Time (dps) | 155.954 *** | 152.42 *** | 75.45 *** | 356.45 *** |
| Interaction (C × (I + N)) | 3.225 n.s. | 25.31 *** | 0.79 n.s. | 37.51 *** |
| Interaction (C × dps) | 53.42 *** | 209.13 *** | 35.58 *** | 77.50 *** |
| Interaction ((I + N) × dps) | 14.261 *** | 8.01 ** | 19.83 *** | 97.94 *** |
| Interaction (C × (I + N) × dps) | 0.791 n.s. | 11.86 *** | 93.97 *** | 82.85 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben-Jabeur, M.; Vicente, R.; López-Cristoffanini, C.; Alesami, N.; Djébali, N.; Gracia-Romero, A.; Serret, M.D.; López-Carbonell, M.; Araus, J.L.; Hamada, W. A Novel Aspect of Essential Oils: Coating Seeds with Thyme Essential Oil induces Drought Resistance in Wheat. Plants 2019, 8, 371. https://doi.org/10.3390/plants8100371
Ben-Jabeur M, Vicente R, López-Cristoffanini C, Alesami N, Djébali N, Gracia-Romero A, Serret MD, López-Carbonell M, Araus JL, Hamada W. A Novel Aspect of Essential Oils: Coating Seeds with Thyme Essential Oil induces Drought Resistance in Wheat. Plants. 2019; 8(10):371. https://doi.org/10.3390/plants8100371
Chicago/Turabian StyleBen-Jabeur, Maissa, Rubén Vicente, Camilo López-Cristoffanini, Noura Alesami, Naceur Djébali, Adrian Gracia-Romero, Maria Dolores Serret, Marta López-Carbonell, Jose Luis Araus, and Walid Hamada. 2019. "A Novel Aspect of Essential Oils: Coating Seeds with Thyme Essential Oil induces Drought Resistance in Wheat" Plants 8, no. 10: 371. https://doi.org/10.3390/plants8100371
APA StyleBen-Jabeur, M., Vicente, R., López-Cristoffanini, C., Alesami, N., Djébali, N., Gracia-Romero, A., Serret, M. D., López-Carbonell, M., Araus, J. L., & Hamada, W. (2019). A Novel Aspect of Essential Oils: Coating Seeds with Thyme Essential Oil induces Drought Resistance in Wheat. Plants, 8(10), 371. https://doi.org/10.3390/plants8100371