Inhibition of Lipid Peroxidation of Kiwicha (Amaranthus caudatus) Hydrolyzed Protein Using Zebrafish Larvae and Embryos




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Amaranth Protein Concentrate (APC) Preparation
2.3. In Vitro APC Gastrointestinal Hydrolysis
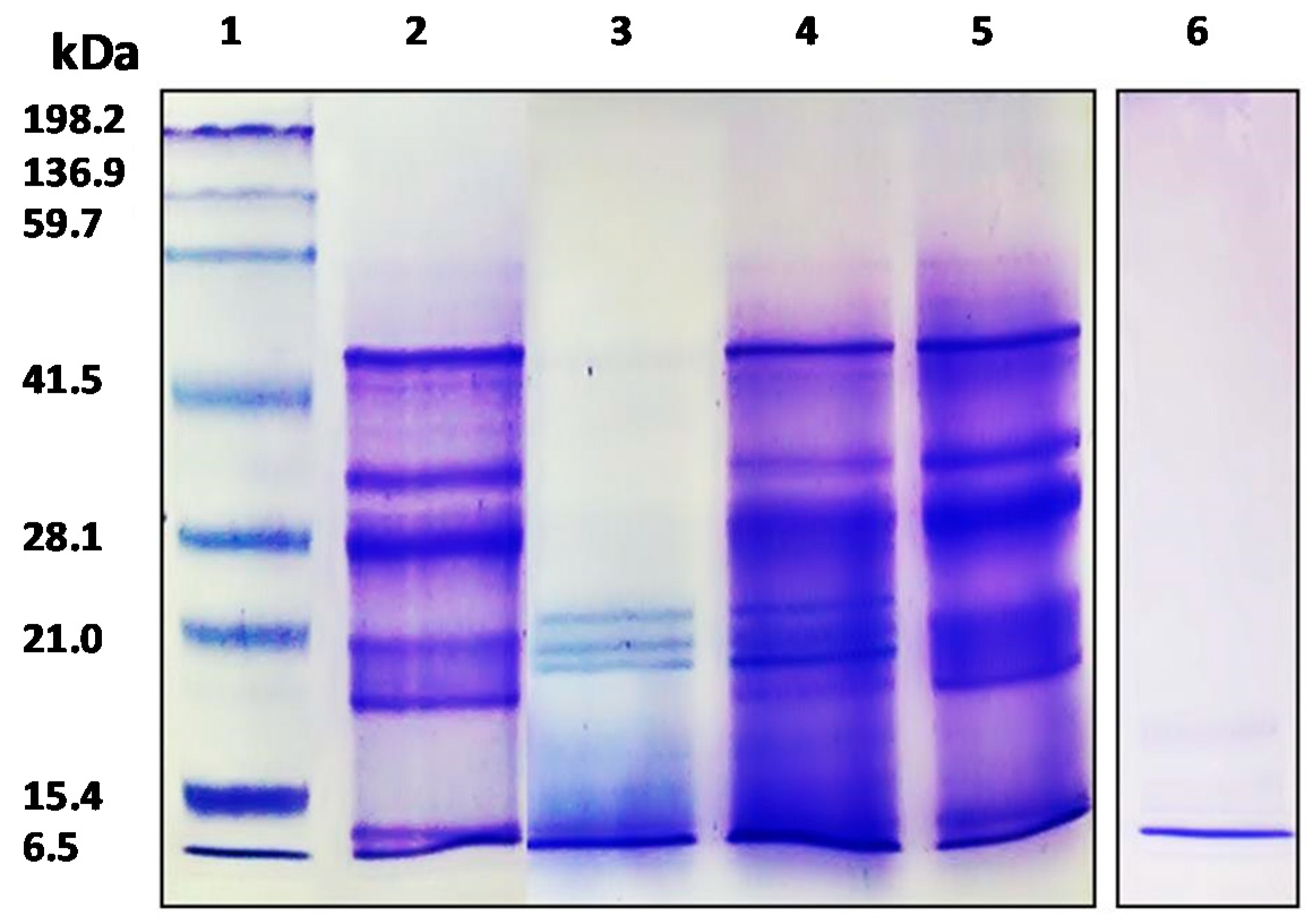
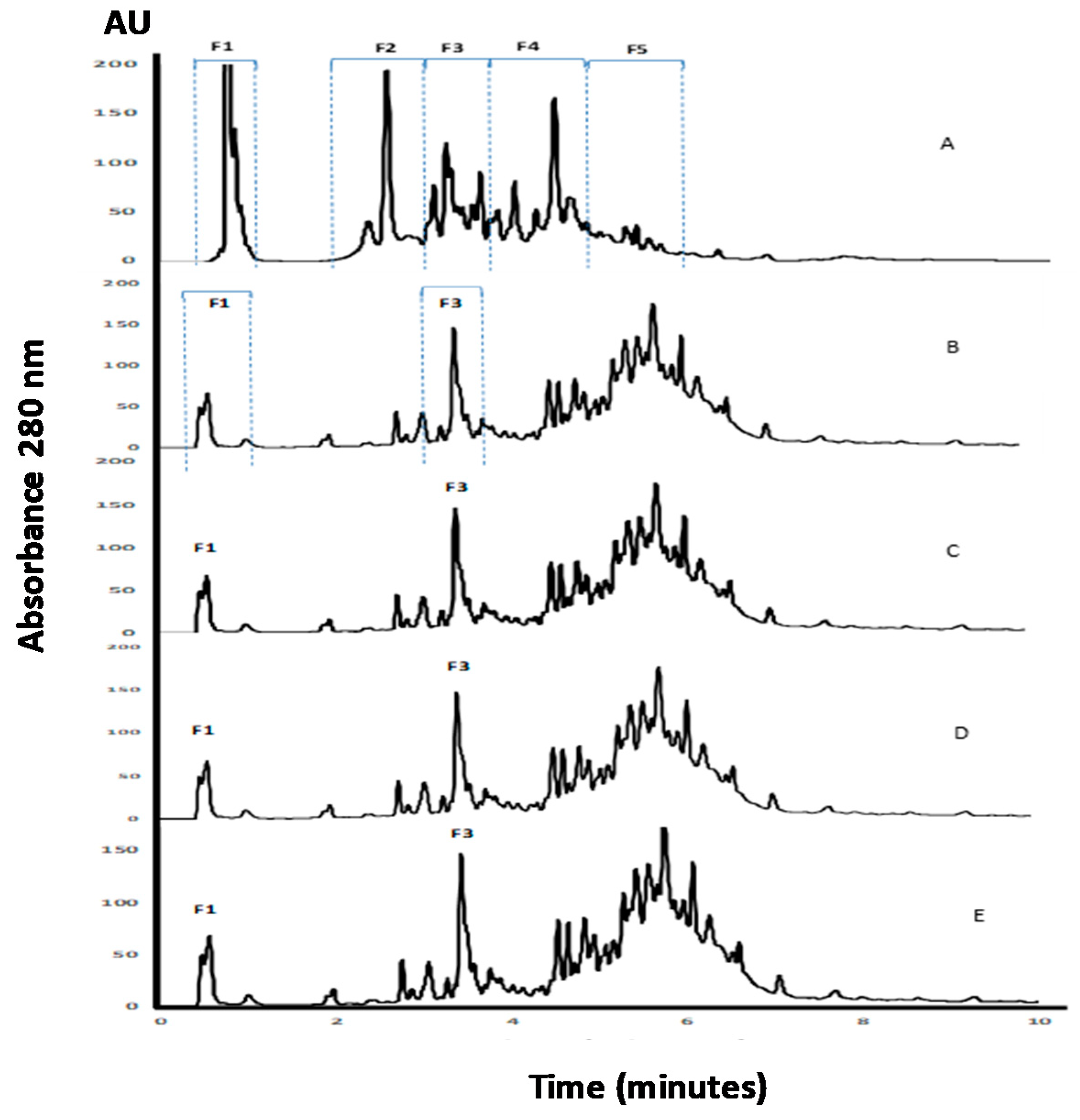
2.4. APC and Hydrolysates Analysis Using SDS-PAGE Electrophoresis and RP-UHPLC Chromatographic Methods
2.5. Evaluation of In Vitro Lipid Peroxidation of APC and Its Hydrolysates
2.6. Evaluation of In Vivo Lipid Peroxidation Inhibition in Zebrafish Larvae in the Presence of H2O2 and Ethanol of APC and Hydrolysates
2.7. Exposure of Embryos of Zebrafish to APC Hydrolysates and AAPH
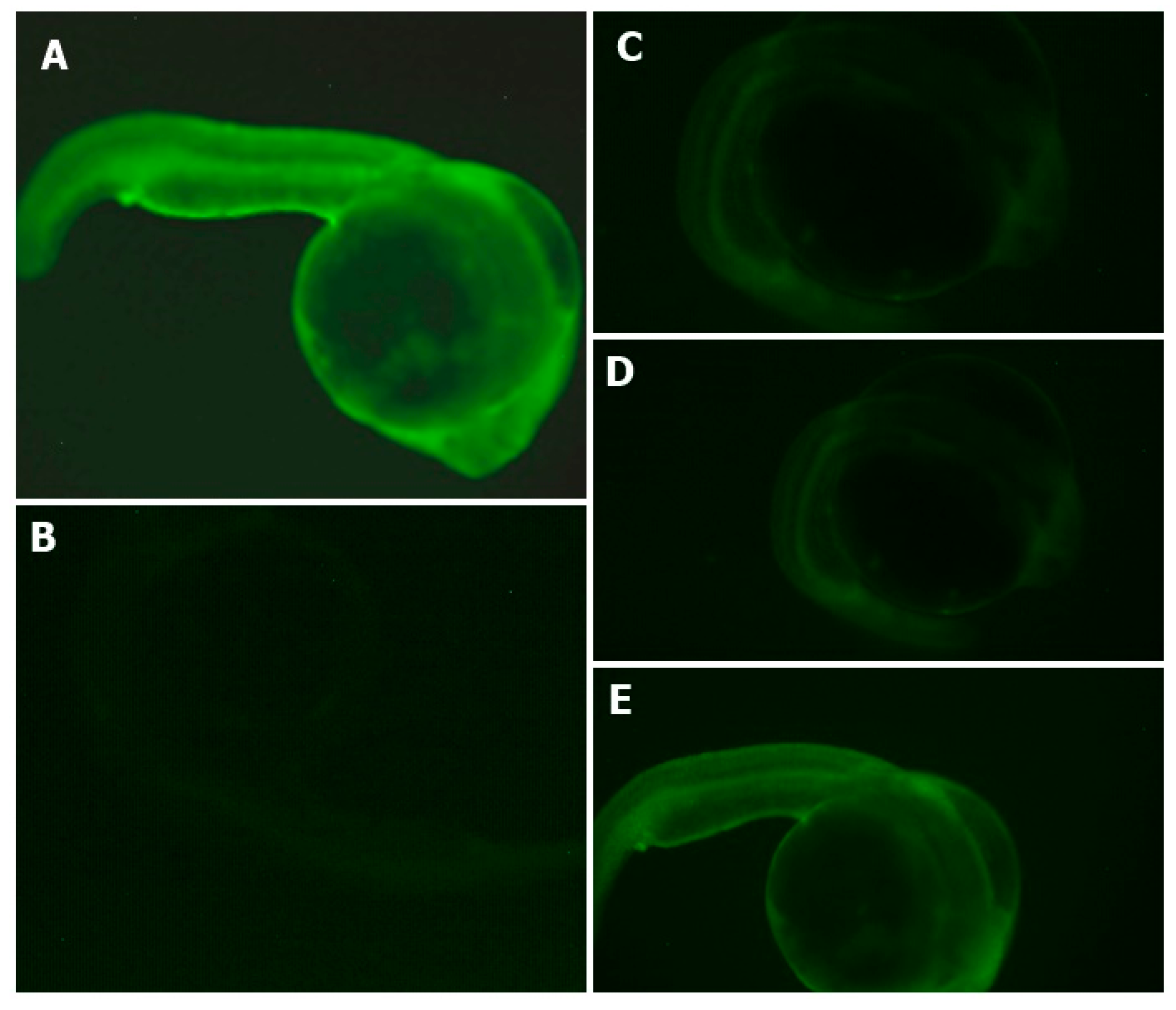
2.8. Estimation of Intracellular ROS Generation and Image Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. Analysis of APC by SDS-PAGE and RP-UHPLC Methods
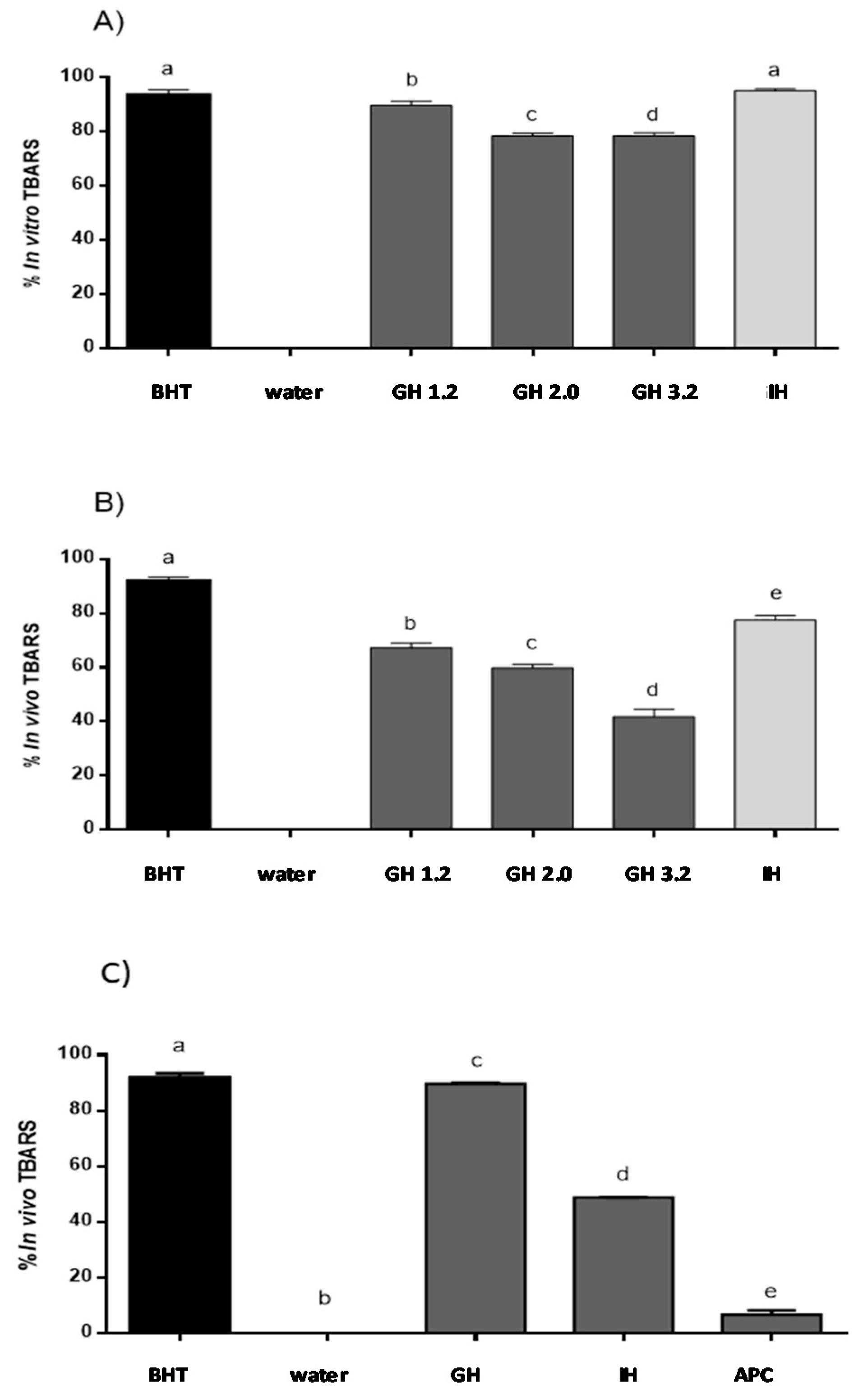
3.2. Evaluation of Reduction Lipid Peroxidation Using the In Vitro Model
3.3. Evaluation of Inhibition of Lipid Peroxidation Using the In Vivo Zebrafish Larvae Model
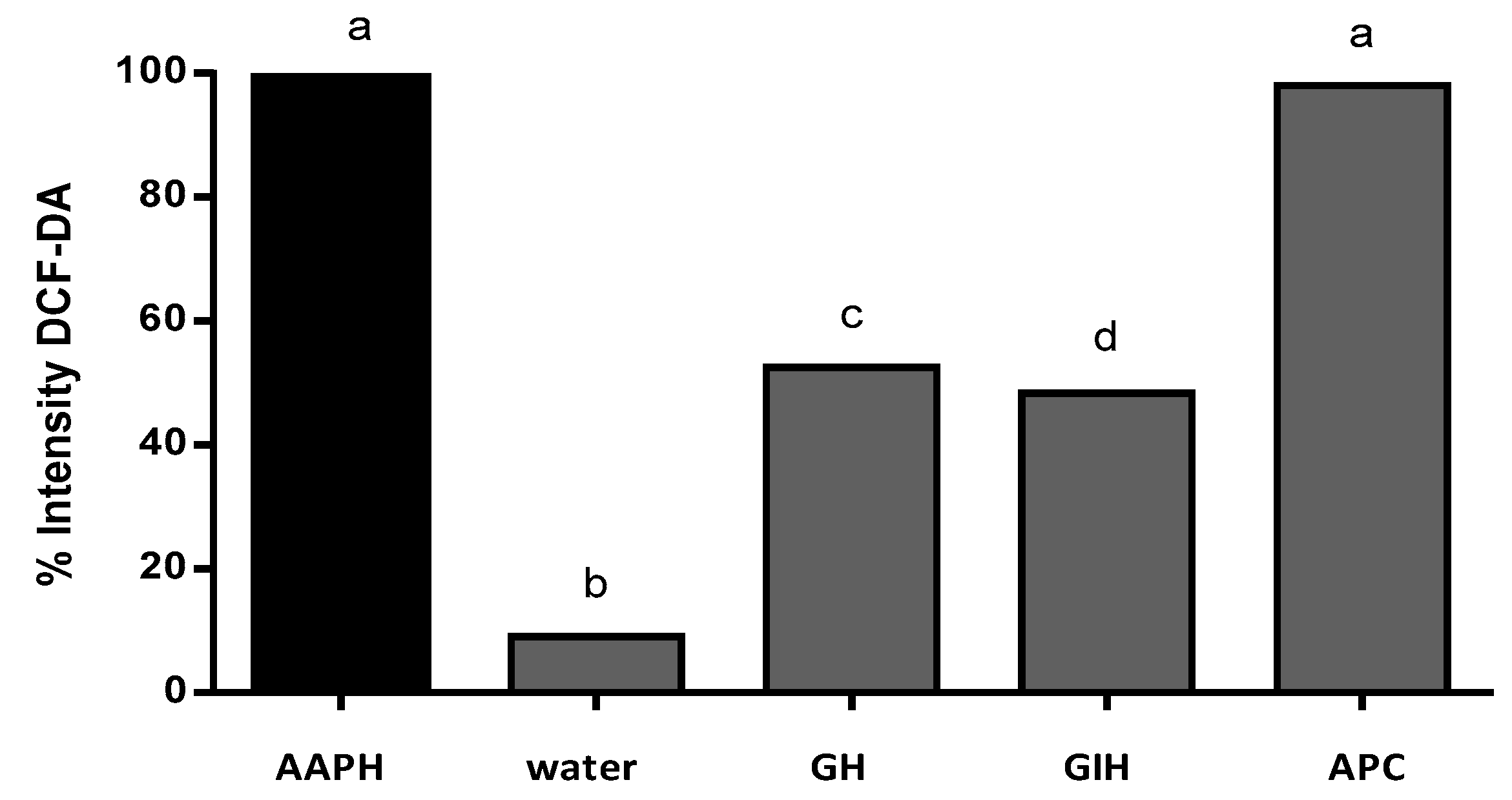
3.4. APC Hydrolysate Protective Effects of Oxidative Stress Produced with AAPH in a Zebrafish Embryo No-Chorion Model
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lado, M.B.; Burini, J.; Rinaldi, G.; Añón, M.C.; Tironi, V.A. Effects of the dietary addition of Amaranth (Amaranthus mantegazzianus) protein isolate on antioxidant status, lipid profiles and blood pressure of rats. Plant Foods Hum. Nutr. 2015, 70, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Fritz, M.; Vecchi, B.; Rinaldi, G.; Añón, M.C. Amaranth seed protein hydrolysates have in vivo and in vitro antihypertensive activity. Food Chem. 2011, 126, 878–884. [Google Scholar] [CrossRef]
- Montoya-Rodríguez, A.; González de Mejia, E. Pure peptides from amaranth (Amaranthus hypocondriacus) proteins inhibit LOX-1 receptor and cellular markers associated with atherosclerosis development in vitro. Food Res. Int. 2015, 77, 204–214. [Google Scholar] [CrossRef]
- Orsini Delgado, M.C.; Nardo, A.; Pavlovic, M.; Rogniaux, H.; Añón, M.C.; Tironi, V.A. Identification and characterization of antioxidant peptides obtained by gastrointestinal digestion of amaranth proteins. Food Chem. 2016, 197, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, W.F.; Marien, W.; Terras, F.R.; De Bolle, M.F.; Proost, P.; Van Damme, J.; Dillen, L.; Clayes, M.; Rees, S.B. Antimicrobial peptides from Amaranthus caudatus seeds with sequence homology to the cysteine/glycine-rich domain of chitin-binding proteins. Biochemistry 1992, 31, 4308–4314. [Google Scholar] [CrossRef] [PubMed]
- Vilcacundo, R.; Miralles, B.; Carrillo, W. Hernández-Ledesma B In vitro chemopreventive properties of peptides released from quinoa (Chenopodium quinoa Willd.) protein under simulated gastrointestinal digestion. Food Res. Int. 2018, 105, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal 2009, 11, 861–905. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Sakagami, H.; Fujisawa, S. Cytotoxicity and apoptosis induction by butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT). Anticancer Res. 2003, 23, 4693–4701. [Google Scholar] [PubMed]
- Nakagawa, Y.; Tayama, K.; Nakao, T.; Hiraga, K. On the mechanism of butylated hydroxytoluene-induced hepatic toxicity in rats. Biochem. Pharmacol. 1984, 33, 2669–2674. [Google Scholar] [CrossRef]
- Vaahtera, L.; Brosche, M.; Wrzaczek, M.; Kangasjarvi, J. Specificity in ROS signaling and transcript signatures. Antioxid. Redox Signal 2014, 21, 1422–1441. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Özyürek, M.; Güçluü, K.; Çapanoğlu, E. Antioxidant activity/capacity measurement. 2. Hydrogen atom transfer (HAT)-based, mixed-mode (electron transfer (ET)/HAT), and lipid peroxidation assays. J. Agric. Food Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef] [PubMed]
- Laguerre, M.; Lecomte, J.; Villeneuve, P. Evaluation of the ability of antioxidants to counteract lipid oxidation: Existing methods, new trends and challenges. Prog. Lipid Res. 2007, 46, 244–282. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Özyürek, M.; Güçluü, K.; Çapanoğlu, E. Antioxidant activity/capacity measurement. 3. Reactive oxygen and nitrogen species (ROS/RNS) scavenging assays, oxidative stress biomarkers, and chromatographic/chemometric assays. J. Agric. Food Chem. 2016, 64, 1046–1070. [Google Scholar] [CrossRef] [PubMed]
- Ghani, M.A.; Barril, C.; Bedgood, D.R., Jr.; Prenzler, P.D. Measurement of antioxidant activity with the thiobarbituric acid reactive substances assay. Food Chem. 2017, 230, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Espín, S.; Sánchez-Virosta, P.; García-Fernández, A.J.; Eeva, T. A microplate adaptation of the thiobarbituric acid reactive substances assay to determine lipid peroxidation fluorometrically in small sample volumes. Toxicol 2017, 34, 94–98. [Google Scholar]
- Alvarez-Jubete, L.; Wijngaard, H.; Arendt, E.K.; Gallagher, E. Polyphenol composition and in vitro antioxidant activity of amaranth, quinoa buckwheat and wheat as affected by sprouting and baking. Food Chem. 2010, 119, 770–778. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Peñas, E.; Dueñas, M.; Frias, J.; Martínez-Villaluenga, C. Optimizing germination conditions to enhance the accumulation of bioactive compounds and the antioxidant activity of kiwicha (Amaranthus caudatus) using response surface methodology. LWT-Food Sci. Technol. 2017, 76, 245–252. [Google Scholar] [CrossRef]
- González-Montoya, M.; Hernández-Ledesma, B.; Silván, J.M.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Peptides derived from in vitro gastrointestinal digestion of germinated soybean proteins inhibit human colon cancer cells proliferation and inflammation. Food Chem. 2018, 242, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.C.; Wrieden, W.L. Influence of milk proteins on lipid oxidation in aqueous emulsion. I. Casein, whey protein, and α-lactalbumin. J. Dairy Res. 1982, 49, 239–248. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Bioactive peptides from Atlantic salmon (Salmo salar) with angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory, and antioxidant activities. Food Chem. 2017, 218, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Jang, G.H.; Byun, C.H.; Jeun, M.; Searson, P.; Lee, K.H. Zebrafish models for functional and toxicological screening of nanoscale drug delivery systems: Promoting preclinical applications. Biosci. Rep. 2017, 37, BSR20170199. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.S. Zebrafish. A complete animal model to enumerate the nanoparticle toxicity. J. Nanobiotechnol. 2016, 14, 65. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.H.; Wen, Z.H.; Lin, C.S.; Chakraborty, C. The zebrafish model: Use in studying cellular mechanisms for a spectrum of clinical disease entities. Curr. Neurovasc. Res. 2007, 4, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, W.; Gómez-Ruiz, J.A.; Miralles, B.; Ramos, M.; Barrio, D.; Recio, I. Identification of antioxidant peptides of hen egg-white lysozyme and evaluation of inhibition of lipid peroxidation and cytotoxicity in the Zebrafish model. Eur. Food Res. Technol. 2016, 242, 1777–1785. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Barrio, D.; Carpio, C.; García-Ruiz, A.; Rúales, J.; Hernández-Ledesma, B.; Carrillo, W. Digestibility of quinoa (Chenopodium quinoa Willd.) protein concentrate and its potential to inhibit lipid peroxidation in the Zebrafish larvae model. Plant Foods Hum. Nutr. 2017, 72, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Acosta, C.; Carpio, C.; Vilcacundo, R.; Carrillo, W. Identification of proteins isolate from amaranth (Amaranthus caudatus) by sodium dodecyl sulfate-polyacrylamide gel electrophoresis with water and NaCl 0.1 m solvents. Asian J. Pharm. Clin. Res. 2016, 9, 331–333. [Google Scholar]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Hernández-Ledesma, B. Release of dipeptidyl peptidase IV, α-amylase and α-glucosidase inhibitory peptides from quinoa (Chenopodium quinoa Willd.) during in vitro simulated gastrointestinal digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef]
- Poveda, T.; Vilcacundo, R.; Carpio, C.; Carrillo, W. Analysis of sesame proteins isolate (Sesamum indicum L) with water and salt treatment. Asian J. Pharm. Clin. Res. 2016, 9, 404–407. [Google Scholar]
- Lara, D.; Vilcacundo, E.; Carrillo, C.; Silva, M.; Alvarez, M.; Carpio, C.; Carrillo, W. Obtention of protein concentrate and polyphenols from macadamia (Macadamia integrifolia) with aqueous extraction method. Asian J. Pharm. Clin. Res. 2017, 10, 138–142. [Google Scholar]
- Carrillo, W.; Guzmán, X.; Vilcacundo, E. Native and heated hydrolysates of milk proteins and their capacity to inhibit lipid peroxidation in the zebrafish larvae model. Foods 2017, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, W.; Tubón, J.; Vilcacundo, R. Isolation of hen egg White lysozyme by cation Exchange chromatography, analysis of its digestibility and evaluation of the inhibition lipid peroxidation in zebrafish model. Asian J. Pharm. Clin. Res. 2016, 9, 345–349. [Google Scholar]
- Bilotta, J.; Barnett, J.A.; Hancock, L.; Saszik, S. Ethanol exposure alters zebrafish development: A novel model of fetal alcohol syndrome. Neurotoxicol. Teratol. 2004, 26, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Cunliffe, V.T. Zebrafish: A Practical Approach. Edited by Nüsslein-Volhard C and Dahm R Oxford University Press. Genet. Res. Camb. 2003, 82, 79–83. [Google Scholar] [CrossRef]
- Rosenkranz, A.R.; Schmaldienst, S.; Stuhlmeier, K.M.; Chen, W.; Knapp, W.; Zlabinger, G.P. A microplate assay for the detection of oxidative product using 2%,7%-dichlorofluorescin-diacetate. J. Immunol. Met. 1992, 156, 39–45. [Google Scholar] [CrossRef]
- Martínez, N.E.; Añón, M.C. Composition and structural characterization of amaranth protein isolates. An electrophoretic and calorimetric study. J. Agric. Food Chem. 1996, 44, 2523–2530. [Google Scholar] [CrossRef]
- Thanapornpoonpong, S.N.; Vearasilp, S.; Pawelzik, E.; Gorinstein, S. Influence of various nitrogen applications on protein and amino acid profiles of amaranth and quinoa. J. Agric. Food Chem. 2008, 56, 11464–11470. [Google Scholar] [CrossRef] [PubMed]
- Orsini Delgado, M.C.; Tironi, V.A.; Añón, M.C. Antioxidant activity of amaranth protein or their hydrolysates under simulated gastrointestinal digestion. LWT-Food Sci. Technol. 2011, 44, 1752–1760. [Google Scholar] [CrossRef]
- Orsini Delgado, M.C.; Galleano, M.; Añón, M.C.; Tironi, V.A. Amaranth peptides from simulated gastrointestinal digestion: Antioxidant activity against reactive species. Plant Foods Hum. Nutr. 2015, 70, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Siddique, Y.H.; Ara, G.; Afzal, M. Estimation of lipid peroxidation induced by hydrogen peroxide in cultured human lymphocytes. Dose-Response 2012, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, S.P.; Zhao, K.J.; Ji, Z.N.; Song, Z.H.; Dong, T.T.; Lo, C.K.; Cheung, J.K.; Zhu, S.Q.; Tsim, K.W. A polysaccharide isolated from Cordyceps sinensis, a traditional Chinese medicine, protects PC12 cells against hydrogen peroxide-induced injury. Life Sci. 2003, 73, 2503–2513. [Google Scholar] [CrossRef]
- Salahudeen, A.K.; Clark, E.C.; Nath, K.A. Hydrogen peroxide-induced renal injury. A protective role for pyruvate in vitro and in vivo. J. Clin. Invest. 1991, 88, 1886–1893. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Pace, R.D.; Dessai, A.P.; Bovell-Benjamin, A.; Phillips, B. Modified extraction method for determining 2-Thiobarbituric acid values in meat with increased specificity and simplicity. J. Food Sci. 2002, 67, 2833–2836. [Google Scholar] [CrossRef]
- Ko, E.Y.; Cho, S.H.; Kwon, S.H.; Eom, C.Y.; Jeong, M.S.; Lee, W.; Jeon, Y.J. The roles of Nf-κb and Ros in regulation of pro-inflammatory mediators of inflammation induction in LPS-stimulated zebrafish embryos. Fish Shellfish Immunol. 2017, 68, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Segner, H. Zebrafish (Danio rerio) as a model organism for investigating endocrine disruption. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Rouach, H.; Park, M.K.; Orfanelli, M.T.; Janvier, B.; Nordmann, R. Ethanol-induced oxidative stress in the rat cerebellum. Alcohol Alcohol 1987, 1, 207–211. [Google Scholar]
- Montoliu, C.; Vallés, S.; Renau-Piqueras, J.; Guerri, C. Ethanol-induced oxygen radical formation and lipid peroxidation in rat brain: Effect of chronic alcohol consumption. J. Neurochem. 1994, 63, 1855–1862. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, R.; Ribière, C.; Rouach, H. Ethanol-induced lipid peroxidation and oxidative stress in extrahepatic tissues. Alcohol Alcohol 1990, 25, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.S.; So, K.F.; Chang, R.C.C. Anti-aging herbal medicine—How and why can they be used in aging-associated neurodegenerative diseases? Ageing Res. Rev. 2010, 9, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Peyrat-Maillard, M.N.; Cuvelier, M.E.; Berset, C. Antioxidant activity of phenolic compounds in 2,2′-azobis (2-amidinopropane) dihydrochloride (AAPH)-induced oxidation: Synergistic and antagonistic effects. J. Am. Oil Chem. Soc. 2003, 80, 1007–1012. [Google Scholar] [CrossRef]
- Joshi, G.; Perluigi, M.; Sultana, R.; Agrippino, R.; Calabrese, V.; Butterfield, D.A. In vivo protection of synaptosomes by ferulic acid ethyl ester (FAEE) from oxidative stress mediated by 2, 2-azobis (2-amidino-propane) dihydrochloride (AAPH) or Fe2+/H2O2: Insight into mechanisms of neuroprotection and relevance to oxidative stress-related neurodegenerative disorders. Neurochem. Int. 2006, 48, 318–327. [Google Scholar] [PubMed]
- Kanski, J.; Lauderback, C.; Butterfield, D.A. 5-Aminosalicylic acid protection against oxidative damage to synaptosomal membranes by alkoxyl radicals in vitro. Neurochem. Res. 2001, 26, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Kanski, J.; Aksenova, M.; Stoyanova, A.; Butterfield, D.A. Ferulic acid antioxidant protection against hydroxyl and peroxyl radical oxidation in synaptosomal and neuronal cell culture systems in vitro: Structure-activity studies. J. Nutr. Biochem. 2002, 13, 273–281. [Google Scholar] [CrossRef]
- Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Lopez-Burillo, S.; Reiter, R.J. Oxidative damage to catalase induced by peroxyl radicals: Functional protection by melatonin and other antioxidants. Free Rad. Res. 2003, 37, 543–553. [Google Scholar] [CrossRef]
- Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Rodriguez, C. Melatonin regulation of antioxidant enzyme gene expression. Cell. Mol. Life Sci. 2002, 59, 1706–1713. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Tan, D.X.; Reiter, R.J.; Karbownik, M.; Manchester, L.C.; Cuzzocrea, S.; Barberi, I. Individual and synergistic antioxidative actions of melatonin: Studies with vitamin E, vitamin C, glutathione and desferrrioxamine (desferoxamine) in rat liver homogenates. J. Pharm. Pharmacol. 2001, 53, 1393–1401. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.C.; Kim, S.Y.; Kim, Y.T.; Kim, E.A.; Lee, S.H.; Ko, S.C.; Jang, H.S. In vitro and in vivo antioxidant activities of polysaccharide purified from aloe vera (Aloe barbadensis) gel. Carbohydr. Polym. 2014, 99, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Ko, C.I.; Jee, Y.; Jeong, Y.; Kim, M.; Kim, J.S.; Jeon, Y.J. Anti-inflammatory effect of fucoidan extracted from Ecklonia cava in zebrafish model. Carbohydr. Polym. 2013, 92, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Goh, A.R.; Youn, G.S.; Yoo, K.Y.; Won, M.H.; Han, S.Z.; Lim, S.S.; Keun, W.W.L.; Soon, Y.C.; Park, J. Aronia melanocarpa concentrate ameliorates pro-inflammatory responses in HaCaT keratinocytes and 12-O-tetradecanoylphorbol-13-acetate-induced ear edema in mice. J. Med. Food. 2016, 19, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Jensen, G.S.; Attridge, V.L.; Beaman, J.L.; Guthrie, J.; Ehmann, A.; Benson, K.F. Antioxidant and anti-inflammatory properties of an aqueous cyanophyta extract derived from Arthrospira platensis: Contribution to bioactivities by the non-phycocyanin aqueous fraction. J. Med. Food. 2015, 18, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, W.; Barrio, D.; Welbaum, J.; Carpio, C.; Vilcacundo, R.; Morales, D.; Ortiz, J. Antimicrobial and antioxidant peptides obtained from food proteins. In Bioactive Peptides: Types, Roles and Research; Powell, K., Ed.; Nova Science Publisher: New York, NY, USA, 2017; pp. 37–58. ISBN 978-1-53610-994-8. [Google Scholar]
- Liégeois, C.; Lermusieau, G.; Collin, S. Measuring antioxidant efficiency of wort, malt, and hops against the 2,2‘-azobis (2-amidinopropane) dihydrochloride-induced oxidation of an aqueous dispersion of linoleic acid. J. Agric. Food Chem. 2000, 48, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.G.; Agar, N.S.; Jones, G.L. Oxidative insult to human red blood cells induced by free radical initiator AAPH and its inhibition by a commercial antioxidant mixture. Life Sci. 2001, 69, 75–86. [Google Scholar] [CrossRef]
- Liao, W.; Ning, Z.; Chen, L.; Wei, Q.; Yuan, E.; Yang, J.; Ren, J. Intracellular antioxidant detoxifying effects of diosmetin on 2, 2-azobis (2-amidinopropane) dihydrochloride (AAPH)-induced oxidative stress through inhibition of reactive oxygen species generation. J. Agric. Food Chem. 2014, 62, 8648–8654. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilcacundo, R.; Barrio, D.A.; Piñuel, L.; Boeri, P.; Tombari, A.; Pinto, A.; Welbaum, J.; Hernández-Ledesma, B.; Carrillo, W. Inhibition of Lipid Peroxidation of Kiwicha (Amaranthus caudatus) Hydrolyzed Protein Using Zebrafish Larvae and Embryos. Plants 2018, 7, 69. https://doi.org/10.3390/plants7030069
Vilcacundo R, Barrio DA, Piñuel L, Boeri P, Tombari A, Pinto A, Welbaum J, Hernández-Ledesma B, Carrillo W. Inhibition of Lipid Peroxidation of Kiwicha (Amaranthus caudatus) Hydrolyzed Protein Using Zebrafish Larvae and Embryos. Plants. 2018; 7(3):69. https://doi.org/10.3390/plants7030069
Chicago/Turabian StyleVilcacundo, Rubén, Daniel Alejandro Barrio, Lucrecia Piñuel, Patricia Boeri, Andrea Tombari, Adelita Pinto, James Welbaum, Blanca Hernández-Ledesma, and Wilman Carrillo. 2018. "Inhibition of Lipid Peroxidation of Kiwicha (Amaranthus caudatus) Hydrolyzed Protein Using Zebrafish Larvae and Embryos" Plants 7, no. 3: 69. https://doi.org/10.3390/plants7030069
APA StyleVilcacundo, R., Barrio, D. A., Piñuel, L., Boeri, P., Tombari, A., Pinto, A., Welbaum, J., Hernández-Ledesma, B., & Carrillo, W. (2018). Inhibition of Lipid Peroxidation of Kiwicha (Amaranthus caudatus) Hydrolyzed Protein Using Zebrafish Larvae and Embryos. Plants, 7(3), 69. https://doi.org/10.3390/plants7030069