Assembly of the Cutin Polyester: From Cells to Extracellular Cell Walls

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Plant Cuticle: Evolution of an Ancestral Lipopolysaccharide Membrane?
3. Reactive Chemical Groups of Cutin Monomers Determine Branching Level of Cutin
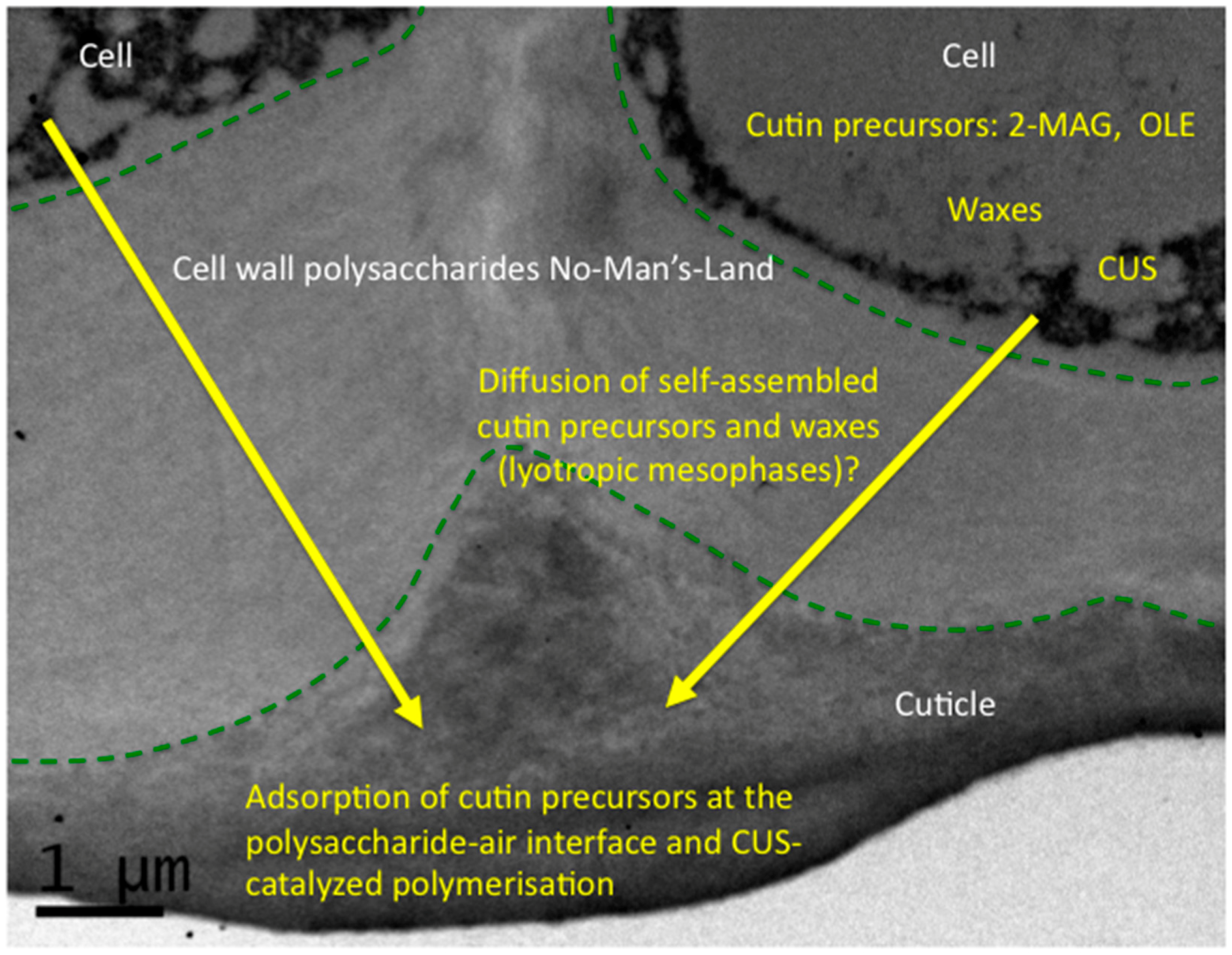
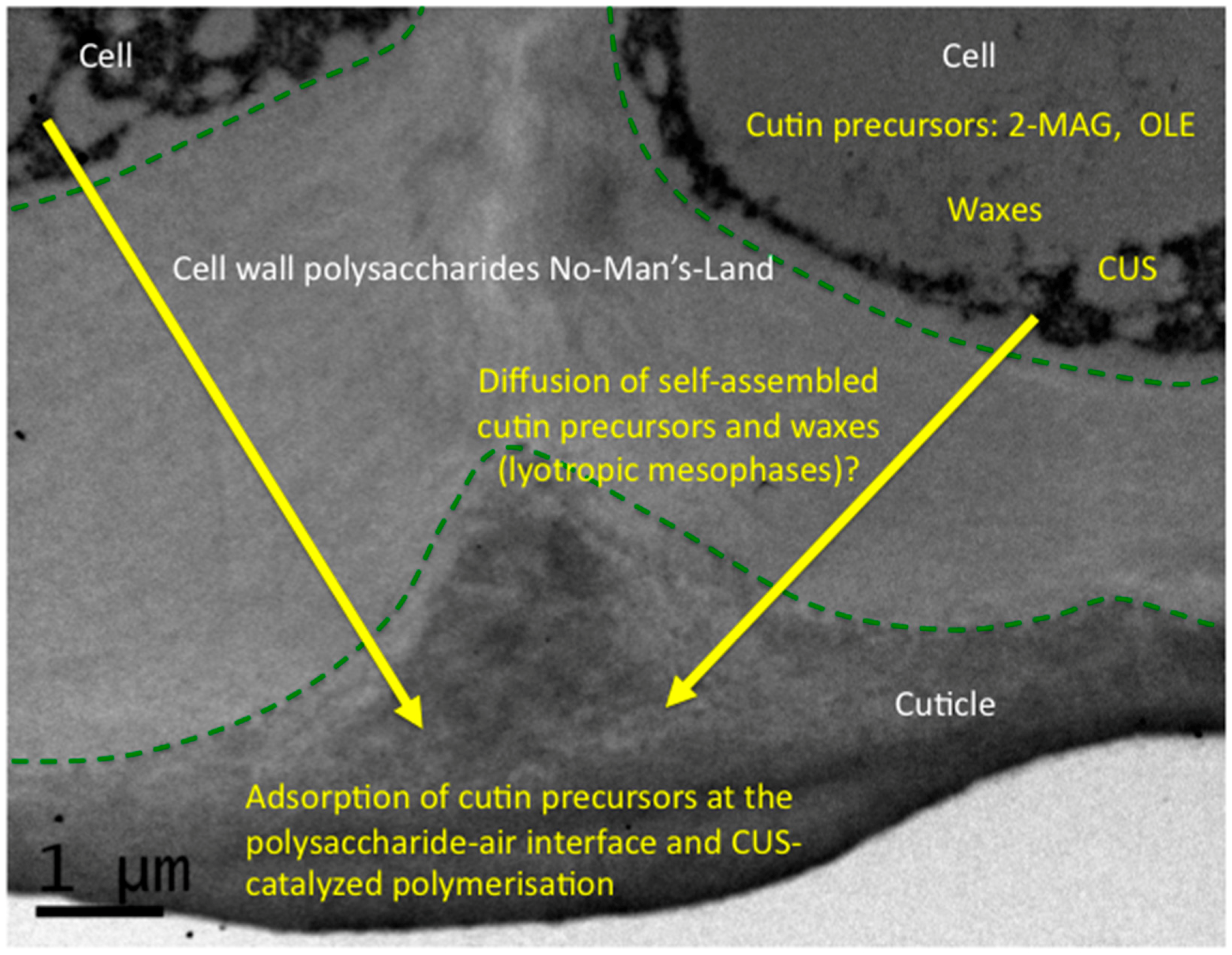
4. The Monomer-to-Polymer Transition: An Interfacial and Hydrophobic Activation of Cutin Polymerization?
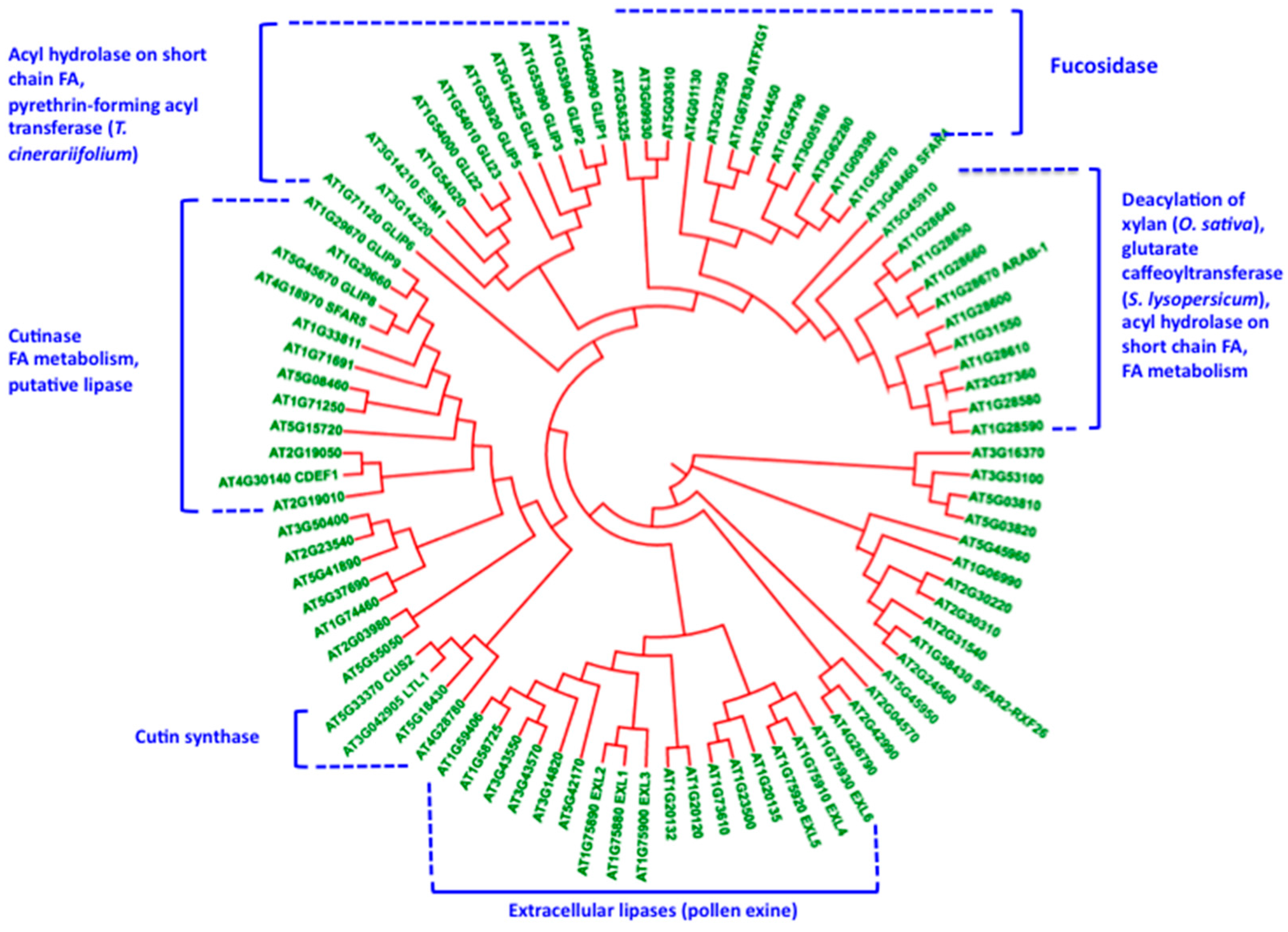
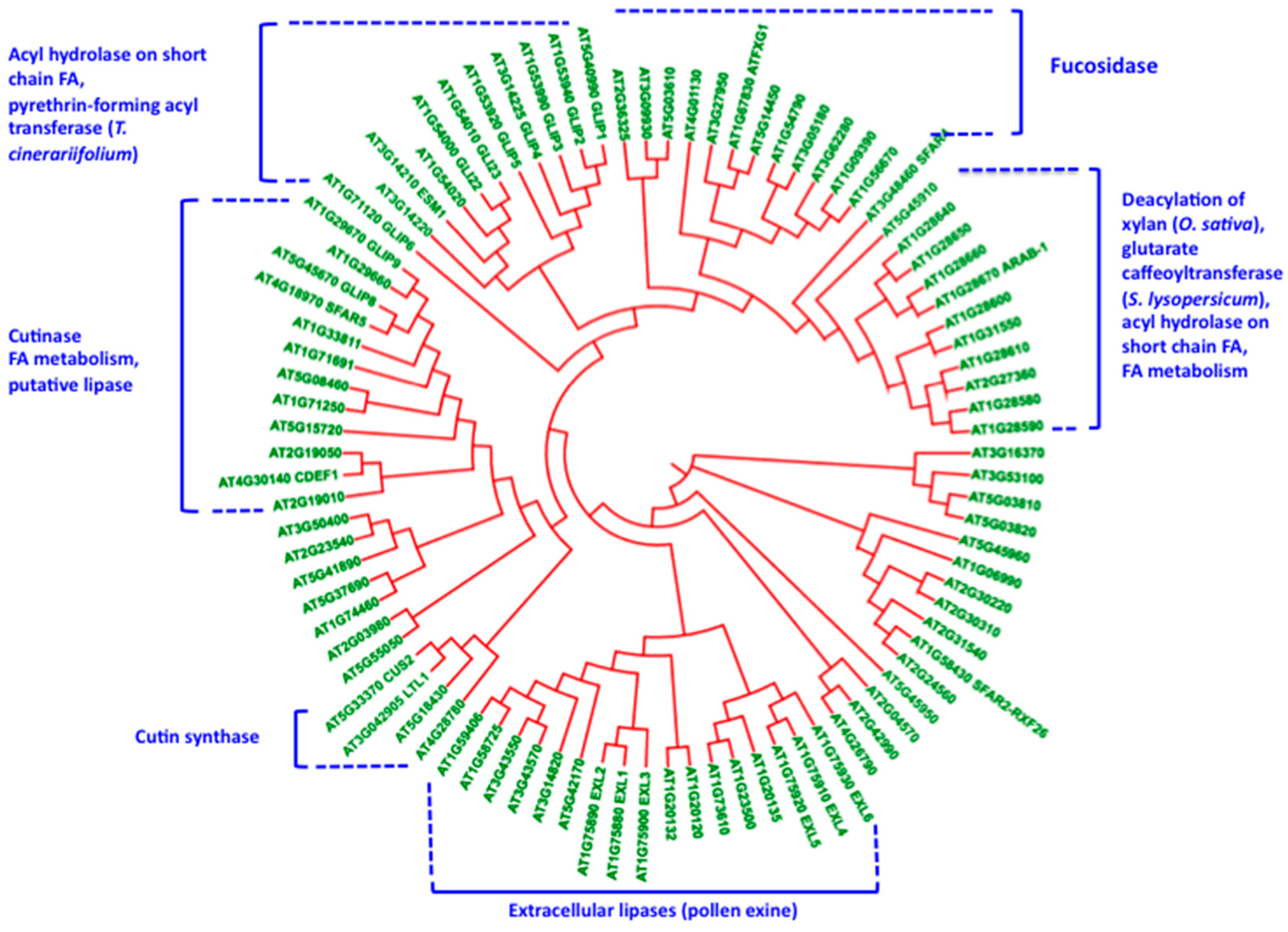
5. Cutin Synthase and the Polymorphism of the GDSL-Lipase Superfamily: Relationships with the Construction of the Lipopolysaccharide Cutin Structure?
6. Conclusions
Author Contributions
Conflicts of Interest
References
- Baker, E.A.; Martin, J.T. Cutin of plant cuticles. Nature 1963, 199, 1268–1270. [Google Scholar] [CrossRef]
- Kolattukudy, P.E. Biosynthesis of a lipid polymer, cutin, structural component of plant cuticle. Biochem. Biophys. Res. Commun. 1970, 41, 299–305. [Google Scholar] [CrossRef]
- Jeffree, C.E. The fine structure of the plant cuticle. In Annual Plant Reviews Volume 23: Biology of the Plant Cuticle; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2007; pp. 11–125. [Google Scholar]
- Trécul, M.A. Mémoire sur l'origine et le développment de la cuticule. C.R. Acad. Sci. 1856, 42, 579–583. [Google Scholar]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.; Bres, C.; Mauxion, P.; Bakan, B.; Rothan, C. Breeding for cuticle-associated traits in crop species: Traits, targets and strategies. J. Exp. Bot. 2017, 68, 5369–5387. [Google Scholar] [CrossRef] [PubMed]
- Alan, P.A.; Ofelia, R.S.; Patricia, T.; Maribel, R.S.R. Cereal bran and wholegrain as a source of dietary fibre: Technological and health aspects. Int. J. Food Sci. Nutr. 2012, 63, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Schieber, A. Side streams of plant food processing as a source of valuable compounds: Selected examples. Annu. Rev. Food Sci. Technol. 2017, 8, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Gandini, A.; Pascoal, C.; Silvestre, A.J.D. Suberin: A promising renewable resource for novel macromolecular materials. Prog. Polym. Sci. 2006, 31, 878–892. [Google Scholar] [CrossRef]
- Garcia, H.; Ferreira, R.; Martins, C.; Sousa, A.F.; Freire, C.S.R.; Silvestre, A.J.D.; Kunz, W.; Rebelo, L.P.N.; Pereira, C.S. Ex situ reconstitution of the plant biopolyester suberin as a film. Biomacromolecules 2014, 15, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Fameau, A.L.; Gaillard, C.; Marion, D.; Bakan, B. Interfacial properties of functionalized assemblies of hydroxy-fatty acids salts isolated from tomato peels. Green Chem. 2012, 15, 341–346. [Google Scholar] [CrossRef]
- Perez, L.; Mol, C.; Bakus, R.C.; Rogers, J.; Rodriguez, G. Plant Extract Compositions for Forming Protective Coatings. WO2017100636 A1, 15 June 2017. [Google Scholar]
- Renault, H.; Alber, A.; Horst, N.A.; Lopes, A.B.; Fich, E.A.; Kriegshauser, L.; Wiedemann, G.; Ullmann, P.; Herrgott, L.; Erhardt, M.; et al. A phenol-enriched cuticle is ancestral to lignin evolution in land plants. Nat. Commum. 2017, 8, 14713. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; Hori, K.; Sasaki-Sekimoto, Y.; Kobayashi, A.; Kato, T.; Yuno-Ohta, N.; Nobusawa, T.; Ohtaka, K.; Shimojima, M.; Ohta, H. Primitive extracellular lipid components on the surface of the charophytic alga Klebsormidium flaccidum and their possible biosynthetic pathways as deduced from the genome sequence. Front. Plant. Sci. 2016, 7, 952. [Google Scholar] [CrossRef] [PubMed]
- Buda, G.J.; Barnes, W.J.; Fich, E.A.; Park, S.; Yeats, T.H.; Zhao, L.X.; Domozych, D.S.; Rose, J.K.C. An ATP binding cassette transporter is required for cuticular wax deposition and desiccation tolerance in the moss Physcomitrella patens. Plant Cell 2013, 25, 4000–4013. [Google Scholar] [CrossRef] [PubMed]
- Caldicott, A.B.; Eglinton, G. Gas chromatographic mass spectrometric studies of long-chain hydroxy-acids. 7. Cutin acids from Bryophytes omega-1 hydroxy alkanoic acid in 2 liverwort species. Phytochemistry 1976, 15, 1139–1143. [Google Scholar] [CrossRef]
- Gemma, S.; Molteni, M.; Rossetti, C. Lipopolysaccharides in Cyanobacteria: A brief overview. Adv. Microbiol. 2016, 6, 391–397. [Google Scholar] [CrossRef]
- Konovalova, A.; Kahne, D.E.; Silhavy, T.J. Outer membrane biogenesis. Annu. Rev. Microbiol. 2017, 71, 539–556. [Google Scholar] [CrossRef] [PubMed]
- Chateigner-Boutin, A.L.; Bouchet, B.; Alvarado, C.; Bakan, B.; Guillon, F. The wheat grain contains pectic domains exhibiting specific spatial and development-associated distribution. PLoS ONE 2014, 9, e89620. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.X.; Malitsky, S.; De Oliveira, S.; Branigan, C.; Franke, R.B.; Schreiber, L.; Aharoni, A. SHINE transcription factors act redundantly to pattern the archetypal surface of Arabidopsis flower organs. PLoS Genet. 2011, 7, e1001388. [Google Scholar] [CrossRef] [PubMed]
- Mazurek, S.; Garroum, I.; Daraspe, J.; De Bellis, D.; Olsson, V.; Mucciolo, A.; Butenko, M.A.; Humbel, B.M.; Nawrath, C. Connecting the molecular structure of cutin to ultrastructure and physical properties of the cuticle in petals of Arabidopsis. Plant Physiol. 2017, 173, 1146–1163. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.H.; Qiu, F.; Yan, B.; Wang, H.; Mort, A.J.; Stark, R.E. NMR studies of molecular structure in fruit cuticle polyesters. Phytochemistry 2001, 57, 1035–1042. [Google Scholar] [CrossRef]
- Tian, S.Y.; Fang, X.H.; Wang, W.M.; Yu, B.W.; Cheng, X.F.; Qiu, F.; Mort, A.J.; Stark, R.E. Isolation and Identification of oligomers from partial degradation of lime fruit cutin. J. Agric. Food Chem. 2008, 56, 10318–10325. [Google Scholar] [CrossRef] [PubMed]
- Pollard, M.; Beisson, F.; Li, Y.H.; Ohlrogge, J.B. Building lipid barriers: Biosynthesis of cutin and suberin. Trends Plant Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Philippe, G.; Gaillard, C.; Petit, J.; Geneix, N.; Dalgalarrondo, M.; Bres, C.; Mauxion, J.P.; Franke, R.; Rothan, C.; Schreiber, L.; et al. Ester cross-link profiling of the cutin polymer of wild-type and cutin synthase tomato mutants highlights different mechanisms of polymerization. Plant Physiol. 2016, 170, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Matzke, K.; Riederer, M. The composition of the cutin of the caryopses and leaves of Triticum aestivum L. Planta 1990, 182, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Gerard, H.C.; Osman, S.F.; Fett, W.F.; Moreau, R.A. Separation, identification and quantification of monomers from cutin polymers by high-performance liquid chromatography and evaporative light-scattering detection. Phytochem. Anal. 1992, 3, 139–144. [Google Scholar] [CrossRef]
- Franke, R.; Briesen, I.; Wojciechowski, T.; Faust, A.; Yephremov, A.; Nawrath, C.; Schreiber, L. Apoplastic polyesters in Arabidopsis surface tissues—A typical suberin and a particular cutin. Phytochemistry 2005, 66, 2643–2658. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, G.; Beisson, F.; Ohlrogge, J.; Pollard, M. Analysis of the aliphatic monomer composition of polyesters associated with Arabidopsis epidermis: Occurrence of octadeca-cis-6, cis-9-diene-1,18-dioate as the major component. Plant J. 2004, 40, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Molina, I.; Ohlrogge, J.B.; Pollard, M. Deposition and localization of lipid polyester in developing seeds of Brassica napus and Arabidopsis thaliana. Plant J. 2008, 53, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Beisson, F.; Li, Y.H.; Bonaventure, G.; Pollard, M.; Ohlrogge, J.B. The acyltransferase GPAT5 is required for the synthesis of suberin in seed coat and root of Arabidopsis. Plant Cell 2007, 19, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Pollard, M.; Li-Beisson, Y.; Ohlrogge, J. Quantitative analysis of glycerol in dicarboxylic acid-rich cutins provides insights into Arabidopsis cutin structure. Phytochemistry 2016, 130, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Graca, J.; Pereira, H. Methanolysis of bark suberins: Analysis of glycerol and acid monomers. Phytochem. Anal. 2000, 11, 45–51. [Google Scholar] [CrossRef]
- Graca, J.; Schreiber, L.; Rodrigues, J.; Pereira, H. Glycerol and glyceryl esters of omega-hydroxyacids in cutins. Phytochemistry 2002, 61, 205–215. [Google Scholar] [CrossRef]
- Fich, E.A.; Segerson, N.A.; Rose, J.K.C. The plant polyester cutin: Biosynthesis, structure, and biological roles. Annu. Rev. Plant Biol. 2016, 67, 207–233. [Google Scholar] [CrossRef] [PubMed]
- Yeats, T.H.; Martin, L.B.B.; Viart, H.M.F.; Isaacson, T.; He, Y.H.; Zhao, L.X.; Matas, A.J.; Buda, G.J.; Domozych, D.S.; Clausen, M.H.; et al. The identification of cutin synthase: Formation of the plant polyester cutin. Nat. Chem. Biol. 2012, 8, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Girard, A.L.; Mounet, F.; Lemaire-Chamley, M.; Gaillard, C.; Elmorjani, K.; Vivancos, J.; Runavot, J.L.; Quemener, B.; Petit, J.; Germain, V.; et al. Tomato GDSL1 Is required for cutin deposition in the fruit cuticle. Plant Cell 2012, 24, 3119–3134. [Google Scholar] [CrossRef] [PubMed]
- Benitez, J.J.; Heredia-Guerrero, J.A.; de Vargas-Parody, M.I.; Cruz-Carrillo, M.A.; Morales-Florez, V.; de la Rosa-Fox, N.; Heredia, A. Biodegradable polyester films from renewable aleuritic acid: Surface modifications induced by melt-polycondensation in air. J. Phys. D Appl. Phys. 2016, 49, 175601. [Google Scholar] [CrossRef]
- Olshansky, Y.; Polubesova, T.; Chefetz, B. Reconstitution of cutin monomers on smectite surfaces: Adsorption and esterification. Geoderma 2014, 232, 406–413. [Google Scholar] [CrossRef]
- Heredia-Guerrero, J.A.; San-Miguel, M.A.; Sansom, M.S.P.; Heredia, A.; Benitez, J.J. Chemical reactions in 2D: Self-assembly and self-esterification of 9(10),16-dihydroxypalmitic acid on mica surface. Langmuir 2009, 25, 6869–6874. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.; Bres, C.; Just, D.; Garcia, V.; Mauxion, J.P.; Marion, D.; Bakan, B.; Joubes, J.; Domergue, F.; Rothan, C. Analyses of tomato fruit brightness mutants uncover both cutin-deficient and cutin-abundant mutants and a new hypomorphic allele of GDSL-lipase. Plant Physiol. 2014, 164, 888–906. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.; Bres, C.; Mauxion, J.P.; Tai, F.W.J.; Martin, L.B.B.; Fich, E.A.; Joubes, J.; Rose, J.K.C.; Domergue, F.; Rothan, C. The glycerol-3-phosphate acyltransferase GPAT6 from tomato plays a central role in fruit cutin biosynthesis. Plant Physiol. 2016, 171, 894–913. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.L.; Pollard, M.; Li-Beisson, Y.; Beisson, F.; Feig, M.; Ohlrogge, J. A distinct type of glycerol-3-phosphate acyltransferase with sn-2 preference and phosphatase activity producing 2-monoacylglycerol. Proc. Natl. Acad. Sci. USA 2010, 107, 12040–12045. [Google Scholar] [CrossRef] [PubMed]
- Sterk, P.; Booij, H.; Schellekens, G.A.; Vankammen, A.; Devries, S.C. Cell-specific expression of the carrot EP2 lipid transfer protein gene. Plant Cell 1991, 3, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Blein, J.P.; Coutos-Thevenot, P.; Marion, D.; Ponchet, M. From elicitins to lipid-transfer proteins: A new insight in cell signalling involved in plant defence mechanisms. Trends Plant Sci. 2002, 7, 293–296. [Google Scholar] [CrossRef]
- Kachroo, A.; Robin, G.P. Systemic signaling during plant defense. Curr. Opin. Plant Biol. 2013, 16, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Le, L.Q.; Lorenz, Y.; Scheurer, S.; Fotisch, K.; Enrique, E.; Bartra, J.; Biemelt, S.; Vieths, S.; Sonnewald, U. Design of tomato fruits with reduced allergenicity by dsRNAi-mediated inhibition of ns-LTP (Lyc e 3) expression. Plant Biotechnol. J. 2006, 4, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Vollhardt, D.; Brezesinski, G. Phase characteristics of 1-monopalmitoyl-rac-glycerol monolayers at the air/water interface. Langmuir 2016, 32, 7316–7325. [Google Scholar] [CrossRef] [PubMed]
- Larsson, K. Lyotropic liquid crystals and their dispersions relevant in foods. Curr. Opin. Colloid Interface Sci. 2009, 14, 16–20. [Google Scholar] [CrossRef]
- Mezzenga, R.; Grigorov, M.; Zhang, Z.D.; Servais, C.; Sagalowicz, L.; Romoscanu, A.I.; Khanna, V.; Meyer, C. Polysaccharide-induced order-to-order transitions in lyotropic liquid crystals. Langmuir 2005, 21, 6165–6169. [Google Scholar] [CrossRef] [PubMed]
- Alfutimie, A.; Curtis, R.; Tiddy, G.J.T. The phase behaviour of mixed saturated and unsaturated monoglycerides in hydrophobic system. Colloids Surf. Physicochem. Eng. Asp. 2015, 482, 329–337. [Google Scholar] [CrossRef]
- Guzman-Puyol, S.; Benitez, J.J.; Dominguez, E.; Bayer, I.S.; Cingolani, R.; Athanassiou, A.; Heredia, A.; Heredia-Guerrero, J.A. Pectin-lipid self-assembly: Influence on the formation of polyhydroxy fatty acids nanoparticles. PLoS ONE 2015, 10, e0124639. [Google Scholar] [CrossRef] [PubMed]
- Manrich, A.; Moreira, F.K.V.; Otoni, C.G.; Lorevice, M.V.; Martins, M.A.; Mattoso, L.H.C. Hydrophobic edible films made up of tomato cutin and pectin. Carbohydr. Polym. 2017, 164, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Panaiotov, I.; Ivanova, M.; Verger, R. Interfacial and temporal organization of enzymatic lipolysis. Curr. Opin. Colloid Interface Sci. 1997, 2, 517–525. [Google Scholar] [CrossRef]
- Reis, P.; Holmberg, K.; Miller, R.; Kragel, J.; Grigoriev, D.O.; Leser, M.E.; Watzke, H.J. Competition between lipases and monoglycerides at interfaces. Langmuir 2008, 24, 7400–7407. [Google Scholar] [CrossRef] [PubMed]
- Wilde, P.J.; Clark, D.C.; Marion, D. Influence of competitive adsorption of a lysopalmitoylphosphatidylcholine on the functional properties of puroindoline, a lipid-binding protein isolated from wheat flour. J. Agric. Food Chem. 1993, 41, 1570–1576. [Google Scholar] [CrossRef]
- Kobayashi, S. Lipase-catalyzed polyester synthesis—A green polymer chemistry. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 338–365. [Google Scholar] [CrossRef] [PubMed]
- Akoh, C.C.; Lee, G.C.; Liaw, Y.C.; Huang, T.H.; Shaw, J.F. GDSL family of serine esterases/lipases. Prog. Lipid Res. 2004, 43, 534–552. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.P.; Huang, L.M.; Chen, L.F.O.; Chan, M.T.; Shaw, J.F. Genome-wide analysis of GDSL-type esterases/lipases in Arabidopsis. Plant Mol. Biol. 2017, 95, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Chepyshko, H.; Lai, C.P.; Huang, M.; Liu, J.H.; Shaw, J.F. Multifunctionality and diversity of GDSL esterase/lipase gene family in rice (Oryza sativa L. japonica) genome: New insights from bioinformatics analysis. BMC Genom. 2012, 13, 309. [Google Scholar] [CrossRef] [PubMed]
- Volokita, M.; Rosilio-Brami, T.; Rivkin, N.; Zik, M. Combining comparative sequence and genomic data to ascertain phylogenetic relationships and explore the evolution of the large GDSL-lipase family in land plants. Mol. Biol. Evol. 2011, 28, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, B. Crystal structure of a full-length autotransporter. J. Mol. Biol. 2010, 396, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Kikuta, Y.; Yamada, G.; Mitsumori, T.; Takeuchi, T.; Nakayama, K.; Katsuda, Y.; Hatanaka, A.; Matsuda, K. Requirement of catalytic-triad and related amino acids for the acyltransferase activity of Tanacetum cinerariifolium GDSL Lipase/Esterase TcGLIP for ester-bond formation in pyrethrin biosynthesis. Biosci. Biotechnol. Biochem. 2013, 77, 1822–1825. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Angkawidjaja, C.; Koga, Y.; Kanaya, S. Calcium-independent opening of lid1 of a family I.3 lipase by a single Asp to Arg mutation at the calcium-binding site. Protein Eng. Des. Sel. 2014, 27, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Fiser, A.; Simon, I. Predicting redox state of cysteines in proteins. Methods Enzymol. 2002, 353, 10–21. [Google Scholar] [PubMed]
- Ferre, F.; Clote, P. DiANNA 1.1: An extension of the DiANNA web server for ternary cysteine classification. Nucleic Acids Res. 2006, 34, W182–W185. [Google Scholar] [CrossRef] [PubMed]
- Ceroni, A.; Passerini, A.; Vullo, A.; Frasconi, P. DISULFIND: A disulfide bonding state and cysteine connectivity prediction server. Nucleic Acids Res. 2006, 34, W177–W181. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Shimada, T.; Kondo, M.; Tamai, A.; Mori, M.; Nishimura, M.; Hara-Nishimura, I. Ectopic expression of an esterase, which is a candidate for the unidentified plant cutinase, causes cuticular defects in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Oh, I.S.; Park, A.R.; Bae, M.S.; Kwon, S.J.; Kim, Y.S.; Lee, J.E.; Kang, N.Y.; Lee, S.M.; Cheong, H.; Park, O.K. Secretome analysis reveals an Arabidopsis lipase involved in defense against Alternaria brassicicola. Plant Cell 2005, 17, 2832–2847. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, F.; Sampedro, J.; Zarra, I.; Revilla, G. AtFXG1, an Arabidopsis gene encoding alpha-L-fucosidase active against fucosylated xyloglucan oligosaccharides. Plant Physiol. 2002, 128, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.C.; Zhang, L.J.; Li, F.; Zhang, D.M.; Liu, X.L.; Wang, H.; Xu, Z.P.; Chu, C.C.; Zhou, Y.H. Control of secondary cell wall patterning involves xylan deacetylation. Nat. Plants 2017, 3, 17017. [Google Scholar] [CrossRef] [PubMed]
- Teutschbein, J.; Gross, W.; Nimtz, M.; Milkowski, C.; Hause, B.; Strack, D. Identification and localization of a lipase-like acyltransferase in phenylpropanoid metabolism of tomato (Solanum lycopersicum). J. Biol. Chem. 2010, 285, 38374–38381. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakan, B.; Marion, D. Assembly of the Cutin Polyester: From Cells to Extracellular Cell Walls. Plants 2017, 6, 57. https://doi.org/10.3390/plants6040057
Bakan B, Marion D. Assembly of the Cutin Polyester: From Cells to Extracellular Cell Walls. Plants. 2017; 6(4):57. https://doi.org/10.3390/plants6040057
Chicago/Turabian StyleBakan, Bénédicte, and Didier Marion. 2017. "Assembly of the Cutin Polyester: From Cells to Extracellular Cell Walls" Plants 6, no. 4: 57. https://doi.org/10.3390/plants6040057
APA StyleBakan, B., & Marion, D. (2017). Assembly of the Cutin Polyester: From Cells to Extracellular Cell Walls. Plants, 6(4), 57. https://doi.org/10.3390/plants6040057