
Light Mitigates Bismuth Toxicity While Sustaining Iron Homeostasis in Lepidium sativum Seedlings

 , ,
, ,  ,
,  ,
,  and
and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
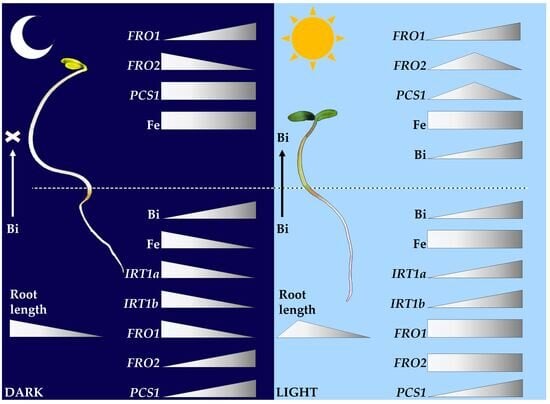
2.1. Root Growth Responses to Bismuth Are Modulated by Light/Dark Exposure in Garden Cress Seedlings
2.2. Identification of LsCDC27B as a Reliable Housekeeping Gene in Garden Cress
2.3. LsFRO1, LsFRO2, and LsIRT1 Expression Is Oppositely Regulated in Dark- and Light-Grown Seedlings Exposed to Bismuth
2.4. Bismuth Is Absorbed by Roots and Impairs Iron Homeostasis in Dark-Grown but Not in Light-Grown Seedlings
3. Materials and Methods
3.1. Germination Test and Seedling Growth Evaluation on Filter Paper
3.2. Total RNA Extraction and qRT-PCR Analysis
3.3. Microscopic Observation of Starch Granules
3.4. Micro X Ray Fluorescence (µ-XRF) Spectroscopy
3.5. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zacchini, M. Bismuth Interaction with Plants: Uptake and Transport, Toxic Effects, Tolerance Mechanisms—A Review. Chemosphere 2024, 360, 142414. [Google Scholar] [CrossRef] [PubMed]
- Passatore, L.; Pietrini, F.; Carloni, S.; Massimi, L.; Giusto, C.; Zacchini, M.; Iannilli, V. Morpho-Physiological and Molecular Responses of Lepidium sativum L. Seeds Induced by Bismuth Exposure. Sci. Total Environ. 2022, 831, 154896. [Google Scholar] [CrossRef] [PubMed]
- Pietrini, F.; Passatore, L.; Carloni, S.; Massimi, L.; Astolfi, M.L.; Giusto, C.; Zacchini, M. Bismuth Exposure Affects Morpho-Physiological Performances and the Ionomic Profile in Garden Cress (Lepidium sativum L.) Plants. Front. Environ. Sci. 2023, 11, 1221573. [Google Scholar] [CrossRef]
- Nagata, T. Growth Inhibition and IRT1 Induction of Arabidopsis thaliana in Response to Bismuth. J. Plant Biol. 2015, 58, 311–317. [Google Scholar] [CrossRef]
- Nishimura, M.; Nagata, T. Phytotoxicity and Iron Homeostasis Disturbance of Bismuth on IRT1 Promoter Knockout Arabidopsis thaliana. Plant 2021, 9, 48. [Google Scholar] [CrossRef]
- Liang, G. Iron Uptake, Signaling, and Sensing in Plants. Plant Commun. 2022, 3, 100349. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, I.; Campbell, N.H.; Ash, J.S.; Connolly, E.L. Expression Profiling of the Arabidopsis Ferric Chelate Reductase (FRO) Gene Family Reveals Differential Regulation by Iron and Copper. Planta 2006, 223, 1178–1190. [Google Scholar] [CrossRef] [PubMed]
- Grill, E.; Loffler, S.; Winnackert, E.-L.; Zenk, M.H. Phytochelatins, the Heavy-Metal-Binding Peptides of Plants, Are Synthesized from Glutathione by a Specific y-Glutamylcysteine Dipeptidyl Transpeptidase (Phytochelatin Synthase). Proc. Natl. Acad. Sci. USA 1989, 86, 6838–6842. [Google Scholar] [CrossRef] [PubMed]
- Trofimov, K.; Mankotia, S.; Ngigi, M.; Baby, D.; Satbhai, S.B.; Bauer, P. Shedding Light on Iron Nutrition: Exploring Intersections of Transcription Factor Cascades in Light and Iron Deficiency Signaling. J. Exp. Bot. 2025, 76, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Hernández, L.E.; Sobrino-Plata, J.; Montero-Palmero, M.B.; Carrasco-Gil, S.; Flores-Cáceres, M.L.; Ortega-Villasante, C.; Escobar, C. Contribution of Glutathione to the Control of Cellular Redox Homeostasis under Toxic Metal and Metalloid Stress. J. Exp. Bot. 2015, 66, 2901–2911. [Google Scholar] [CrossRef] [PubMed]
- Capobianco, G.; Bonifazi, G.; Serranti, S.; Marabottini, R.; Antenozio, M.L.; Cardarelli, M.; Brunetti, P.; Stazi, S.R. A Green Approach Based on Micro-X-Ray Fluorescence for Arsenic, Micro- and Macronutrients Detection in Pteris vittata. Water 2022, 14, 2202. [Google Scholar] [CrossRef]
- APAT. “Proposal of a Technical Guidance on Analysis Methods for Soil and Contaminated Sites”. In Use of Ecotoxicological and Biological Indicators (Proposta Di Guida Tecnica Su Metodi Di Analisi per Il e i Siti Contaminati. Utilizzo Di Indicatori Biologici Ed Ecotossicologici); APAT, Agenzia per La Protezione Dell’Ambiente e per i Servizi Tecnici: Rome, Italy, 2004. [Google Scholar]
- Wang, H.; Wang, J.; Jiang, J.; Chen, S.; Guan, Z.; Liao, Y.; Chen, F. Reference Genes for Normalizing Transcription in Diploid and Tetraploid Arabidopsis. Sci. Rep. 2014, 4, 6781. [Google Scholar] [CrossRef] [PubMed]
- Graeber, K.; Linkies, A.; Wood, A.T.A.; Leubner-Metzger, G. A Guideline to Family-Wide Comparative State-of-the-Art Quantitative RT-PCR Analysis Exemplified with a Brassicaceae Cross-Species Seed Germination Case Study. Plant Cell 2011, 23, 2045–2063. [Google Scholar] [CrossRef] [PubMed]
- Blilou, I.; Frugier, F.; Folmer, S.; Serralbo, O.; Willemsen, V.; Wolkenfelt, H.; Eloy, N.B.; Ferreira, P.C.G.; Weisbeek, P.; Scheres, B. The Arabidopsis HOBBIT Gene Encodes a CDC27 Homolog That Links the Plant Cell Cycle to Progression of Cell Differentiation. Genes Dev. 2002, 16, 2566–2575. [Google Scholar] [CrossRef] [PubMed]
- Vert, G.A.; Briat, J.F.; Curie, C. Dual regulation of the Arabidopsis high-affinity root iron uptake system by local and long-distance signals. Plant Physiol. 2003, 132, 796–804, Erratum in Plant Physiol. 2024, 194, 2772–2774. https://doi.org/10.1093/plphys/kiad677. [Google Scholar] [CrossRef] [PubMed]
- Lanquar, V.; Lelièvre, F.; Bolte, S.; Hamès, C.; Alcon, C.; Neumann, D.; Vansuyt, G.; Curie, C.; Schröder, A.; Krämer, U.; et al. Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron. EMBO J. 2005, 24, 4041–4051. [Google Scholar] [CrossRef] [PubMed]
- Le¡ková, A.; Giehl, R.F.H.; Hartmann, A.; Farga¡ová, A.; von Wirén, N. Heavy Metals Induce Iron Deficiency Responses at Different Hierarchic and Regulatory Levels. Plant Physiol. 2017, 174, 1648–1668. [Google Scholar] [CrossRef] [PubMed]
- Talebi, M.; Tabatabaei, B.E.S.; Akbarzadeh, H. Hyperaccumulation of Cu, Zn, Ni, and Cd in Azolla Species Inducing Expression of Methallothionein and Phytochelatin Synthase Genes. Chemosphere 2019, 230, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.F.; Curie, C. IRT1, an Arabidopsis Transporter Essential for Iron Uptake from the Soil and for Plant Growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Dhankher, O.P.; Carreira, L.; Lee, D.; Chen, A.; Schroeder, J.I.; Balish, R.S.; Meagher, R.B. Overexpression of Phytochelatin Synthase in Arabidopsis Leads to Enhanced Arsenic Tolerance and Cadmium Hypersensitivity. Plant Cell Physiol. 2004, 45, 1787–1797. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Moon, J.S.; Ko, T.-S.; Petros, D.; Goldsbrough, P.B.; Korban, S.S. Overexpression of Arabidopsis Phytochelatin Synthase Paradoxically Leads to Hypersensitivity to Cadmium Stress. Plant Physiol. 2003, 131, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Ueda, Y.; Mukai, A.; Ochiai, K.; Matoh, T. Rice Phytochelatin Synthases OsPCS1 and OsPCS2 Make Different Contributions to Cadmium and Arsenic Tolerance. Plant Direct 2018, 2, e00034. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, X.; Liang, L.; Chen, C.; Wei, S.; Zhou, Q. Phytochelatin and Oxidative Stress Under Heavy Metal Stress Tolerance in Plants. In Reactive Oxygen Species and Oxidative Damage in Plants Under Stress; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 191–217. [Google Scholar]
- Dayer, S.; Herrera, J.C.; Dai, Z.; Burlett, R.; Lamarque, L.J.; Delzon, S.; Bortolami, G.; Cochard, H.; Gambetta, G.A. Nighttime Transpiration Represents a Negligible Part of Water Loss and Does Not Increase the Risk of Water Stress in Grapevine. Plant Cell Environ. 2021, 44, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Marzi, D.; Brunetti, P.; Mele, G.; Napoli, N.; Calò, L.; Spaziani, E.; Matsui, M.; De Panfilis, S.; Costantino, P.; Serino, G.; et al. Light Controls Stamen Elongation via Cryptochromes, Phytochromes and COP1 through HY5 and HYH. Plant J. 2020, 103, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Patat, A.S.; Sen, F.; Erdogdu, B.S.; Uncu, A.T.; Uncu, A.O. Construction and Characterization of a de Novo Draft Genome of Garden Cress (Lepidium sativum L.). Funct. Integr. Genom. 2022, 22, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.H.; Chu, H.; Zhang, C.; Ghosh, D.; Gong, X.; Xu, J. A Quantitative Analysis of Stem Cell Homeostasis in the Arabidopsis Columella Root Cap. Front. Plant Sci. 2015, 6, 206. [Google Scholar] [CrossRef]
- Antenozio, M.L.; Capobianco, G.; Costantino, P.; Vamerali, T.; Bonifazi, G.; Serranti, S.; Brunetti, P.; Cardarelli, M. Arsenic Accumulation in Pteris vittata: Time Course, Distribution, and Arsenic-Related Gene Expression in Fronds and Whole Plantlets. Environ. Pollut. 2022, 309, 119773. [Google Scholar] [CrossRef] [PubMed]
- Sherman, J. The Theoretical Derivation of Fluorescent X-Ray Intensities from Mixtures. Spectrochim. Acta 1955, 7, 283–306. [Google Scholar] [CrossRef]
- de Winter, N.J.; Sinnesael, M.; Makarona, C.; Vansteenberge, S.; Claeys, P. Trace Element Analyses of Carbonates Using Portable and Micro-X-Ray Fluorescence: Performance and Optimization of Measurement Parameters and Strategies. J. Anal. At. Spectrom. 2017, 32, 1211–1223. [Google Scholar] [CrossRef]
- Capobianco, G.; Brunetti, P.; Bonifazi, G.; Costantino, P.; Cardarelli, M.; Serranti, S. The use of micro-energy dispersive X-ray fluorescence spectrometry combined with a multivariate approach to determine element variation and distribution in tobacco seedlings exposed to arsenate. Spectrochim. Acta Part B At. Spectrosc. 2018, 147, 132–140. [Google Scholar] [CrossRef]
- Marzi, D.; Mazzurco-Miritana, V.; Passatore, L.; Zacchini, M.; Pietrini, F.; Carloni, S.; Peruzzi, E.; Louro, M.; Sousa Silva, M.; Cordeiro, C.; et al. Assessment of Phyto-Compatibility after Soil Remediation: Insights from Plant Growth, Physiological, and Metabolomic Analyses. J. Environ. Manag. 2025, 393, 127239. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Caissutti, C.; Marzi, D.; Capobianco, G.; Serranti, S.; Licursi, V.; Zacchini, M.; Brunetti, P. Light Mitigates Bismuth Toxicity While Sustaining Iron Homeostasis in Lepidium sativum Seedlings. Plants 2026, 15, 1898. https://doi.org/10.3390/plants15121898
Caissutti C, Marzi D, Capobianco G, Serranti S, Licursi V, Zacchini M, Brunetti P. Light Mitigates Bismuth Toxicity While Sustaining Iron Homeostasis in Lepidium sativum Seedlings. Plants. 2026; 15(12):1898. https://doi.org/10.3390/plants15121898
Chicago/Turabian StyleCaissutti, Cristina, Davide Marzi, Giuseppe Capobianco, Silvia Serranti, Valerio Licursi, Massimo Zacchini, and Patrizia Brunetti. 2026. "Light Mitigates Bismuth Toxicity While Sustaining Iron Homeostasis in Lepidium sativum Seedlings" Plants 15, no. 12: 1898. https://doi.org/10.3390/plants15121898
APA StyleCaissutti, C., Marzi, D., Capobianco, G., Serranti, S., Licursi, V., Zacchini, M., & Brunetti, P. (2026). Light Mitigates Bismuth Toxicity While Sustaining Iron Homeostasis in Lepidium sativum Seedlings. Plants, 15(12), 1898. https://doi.org/10.3390/plants15121898