Abstract
The interspecific relations that have been previously observed within the Stipa dasyphylla group are intricate and require further clarification. The aim of this study was to determine whether the specimens from Serbia deserve a separate taxonomic status. Various “dasyphyllous” Stipa species (those with hairy adaxial leaves) from the Balkan Peninsula were collected and analysed using morphological methods and microscopic observations of the macro- and micro-ornamentation of their leaves and lemmas. Based on analyses of 160 individuals from 17 populations belonging to three “dasyphyllous” Stipa species (S. pontica, S. ucrainica, and S. dasyphylla) which were collected in the central and eastern Balkan Peninsula (Serbia, North Macedonia, Bulgaria, Greece) and in Central Europe (Czech Republic, Slovakia, Hungary, Romania), Stipa stevanoviciorum is described as a new species from Serbia. This taxon includes the subspecies stevanoviciorum, which grows on ultramafic substrates, and the subspecies pseudodasyphylla, which is found on carbonate bedrock. These taxa differ from the closely related S. dasyphylla primarily in the length of their upper cauline leaves and the distance from the end of the dorsal line of the hairs to the top of the anthecium. The ecological and biogeographical characteristics of the taxa and a morphological comparison with similar species are given together with the key to the species’ identification. Images of key morphological characters are included, along with information on their distribution, habitat, and conservation implications.
1. Introduction
Stipa L. is one of the largest genera in the family Poaceae and the subfamily Pooideae [1]. Due to its very complicated taxonomy and unclear delimitation, as well as its uncertain number of species, the genus has always been considered obscure [2]. According to the current narrow approach, the genus encompasses approximately 150 species distributed throughout Asia, Europe, and North Africa, with the greatest diversity being found in the open grasslands and steppes of the warm temperate zones of the Old World [3,4,5,6,7,8,9,10,11,12]. To date, there are many unresolved taxonomic issues within the different taxonomic levels of this genera, yet new species continue to be described, mainly species from Asia but also species from Africa and Europe [11,12,13,14,15,16,17,18,19,20,21,22,23,24,25].
The nominal section of the genus Stipa L., which is widely regarded as one of the most taxonomically problematic genera, comprises up to 55 species that depend on various concepts [5,7,8,12,21,26,27,28,29,30,31,32,33]. Furthermore, one of its supposedly taxonomically sound and well-differentiated groups—the series Dasyphyllae Martinovský (= “dasyphyllous” Stipa species)—has turned out to be very intricate, encompassing Central- and Eastern-European species, as well as those distributed in the Caucasus, Siberia and Asia. The species of this series represent a natural, clearly defined group of feather grasses within the section Stipa, which is primarily characterized by its leaf ornamentation, i.e., the leaf surface of these species is never bare. The indumentum elements appear in different forms, such as hairs, bristles, conical papillae, or transitional structures in between these. These indumentum structures tend to vary, amongst the species of this group, in length, frequency, and the size of the angle they form with the leaf surface. They actually represent homologous and evolutionarily ancient structures of epidermal origin that have an important taxonomic value, and indicate the natural specificity of this group [33]. Despite all of these characteristics, the interspecific relations within this group are intricate and require further clarification (for details see [34]).
Based on the key and the descriptions given in Flora Europea [7], the S. dasyphylla group comprises four species: S. dasyphylla (Lindem.) Czern. ex Trautv., S. zaleskii Wilensky, S. pontica P. A. Smirn., and S. ucrainica P. A. Smirn, although there are different interpretations regarding the representatives of this group [30,34,35]. The nominal species of the series, S. dasyphylla, has been known in Europe for the longest time. It was the first representative of the complex to appear in the botanical literature [34]. The type specimen was collected in 1853 by Chernyaev in Ukraine, Kharkov, on knolls and hills of Rogan [32]. The ornamentation of the leaf blade (both on the vegetative and cauline leaves) is a constant feature that clearly distinguishes S. dasyphylla from other species of the group. S. dasyphylla has leaves whose abaxial surface is covered with abundant long soft hairs over their entire length, while the adaxial surface also bares sparse or abundant long, more or less soft hairs throughout its whole length [32]. The presence of these soft hairs over the whole leaf length makes S. dasyphylla one of the most easily recognizable species within the S. dasyphylla group. The other species of the S. dasyphylla group, S. ucrainica and S. zalesskii (including S. pontica), are known to grow in the Balkan Peninsula [7,36,37]. On the other hand, the distribution of S. dasyphylla in the Balkans was not previously clear. This was mainly due to the fact that Martinovský [34], in his earlier work, considered S. dasyphylla as an aggregate species comprising three taxonomically closely related microspecies that occur in the relic central- and southeastern- European steppes, and considered that misidentifications were likely to happen. This species is generally distributed from Central Asia (north to southern part of western Siberia), across the European part of Russia, to Central Europe (west to Switzerland and Germany) [32,38]. In the Balkan Peninsula, it has so far been recorded at one site in Greece [32] and most recently in Serbia [39]. Valdés et al. [38] have also reported its observation in Bulgaria, but on the basis of personal communication with H. Scholz from 2003.
In this study, we focused on populations of S. dasyphylla from Serbia and other geographically close populations of “dasyphyllous” Stipa species in the central and eastern Balkan Peninsula and Central Europe. We performed classical morphometric analyses to gain insight into the diversity of the populations studied. The main aim of this study was to determine whether the populations of S. dasyphylla from Serbia are morphologically outstanding or not.
2. Results
In our research on the distribution and diversity of the genus Stipa in the Balkan Peninsula, we have found only a few populations that belong to the “dasyphyllous” Stipa species. By comparing and examining plants during fieldwork, as well as the available herbarium specimens, we found that the most morphologically striking and diverse populations are plants from Serbia that were previously identified as S. dasyphylla.
2.1. Morphometric Analyses
The PCA shows that, with the exception of the populations from Serbia, the other populations show only a very low degree of separation. Along the first two principal component axes, the populations of S. dasyphylla from Serbia are completely separated, while the other populations of S. dasyphylla, from Central Europe, are associated with S. ucrainica and S. pontica, which largely overlap. Along the first and third discriminant principal component axes, populations of S. dasyphylla from Central Europe overlap with populations of S. dasyphylla from Serbia. At the same time, this project completely separates the populations of S. dasyphylla from the largely overlapping S. ucrainica and S. pontica populations (Figure 1). The characters that contribute most to the observed structure of variability are the plant height (PH), panicle length (LP), number of flowers in the panicle (NF), length of vegetative shoots (LV), basal leaf-blade diameter (BASD), length of the anthecium (AL), distance from the end of the dorsal line of the hairs to the top of the anthecium (DDL), and distance from the end of the ventral line of the hairs to the top of the anthecium (DVL).
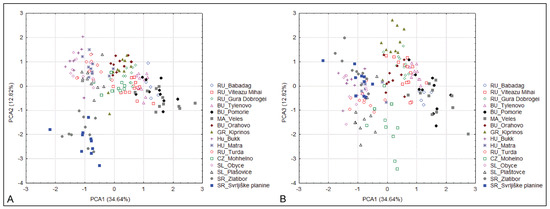
Figure 1.
Principal component analysis (PCA) of 160 individuals from 17 populations belonging to three species of “dasyphyllous“ Stipa from the Balkan Peninsula based on 20 morphometric characters. (A) the two first PCA axes, (B) the first and third PCA axes. The acronyms in the figure represent the country code and site toponym: BU—Bulgaria, GR—Greece, HU—Hungary, RU—Romania, CZ—Czech, SL—Slovenia, SR—Serbia.
The CDA that was performed on six groups of individuals identified a priori as potential OTUs based on their taxonomic and biogeographic affiliations not only confirmed the results of the PCA, i.e., the complete morphological segregation of S. dasyphylla from Serbia, but also the segregation between Central European S. dasyphylla, S. ucrainica and S. pontica along the first three discriminant axes (Figure 2). In addition, the CDA revealed a significant morphological diversification between the western and eastern Serbian populations of S. dasyphylla. On the third discriminant axis, the Illyrian populations from the ultramafic rocks of western Serbia (S. dasyphylla-SR-W) and the Moesian populations from the limestones of eastern Serbia (S. dasyphylla-SR-E) are almost completely separated from each other. At the same time, the CDA showed extensive overlap between Danubian populations of S. ucrainica from Bulgaria and Romania (S. ucrainica-BU-RU) and Macedonian populations of S. ucrainica from North Macedonia (S. ucrainica-MA) (Figure 2).
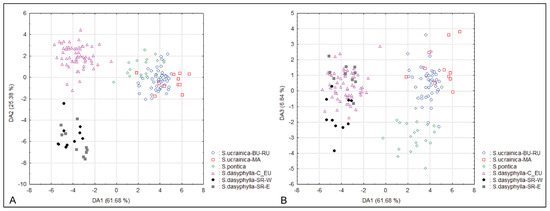
Figure 2.
Canonical discriminant analyses of 160 individuals from 17 populations belonging to three species of “dasyphyllous” Stipa from the Balkan Peninsula, based on 20 morphometric characters. All individuals are included in six groups identified a priori as potential OTUs based on their taxonomic and biogeographic affiliations. (A) the two first discriminant axes, (B) the first and the third discriminant axes. The acronyms in the figure represent the combination of the species name and biogeografic affiliation of the group: BU—Bulgaria, C_EU—central Europe, MA—North Macedonia, RU—Romania, SR-E—eastern Serbia, SR-W—western Serbia.
The UPGMA cluster analysis revealed that the degree of morphological diversification between the eastern and western populations of S. dasyphylla in Serbia is extremely high. In contrast, the morphological differentiation between the Macedonian and Bulgarian-Romanian populations of S. ucrainica in the Black Sea hinterland is minimal. At the same time, S. pontica and the Central European S. dasyphylla show a moderate degree of segregation (Figure 3).
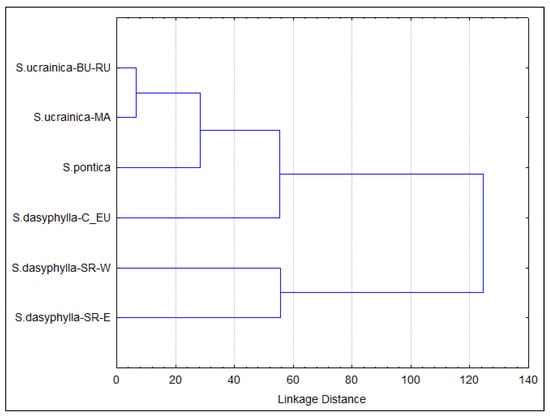
Figure 3.
Overall similarities among morphological groups of “dasyphyllous” Stipa from the Balkan Peninsula based on the use of the agglomerative UPGMA clustering method on morphometric data. All individuals are included in six groups identified a priori as potential OTUs based on their taxonomic and biogeographic affiliations. The acronyms in the figure represent the combination of the species name and biogeografic affiliation of the group: BU—Bulgaria, C_EU—central Europe, MA—North Macedonia, RU—Romania, SR-E—eastern Serbia, SR-W—western Serbia.
The classification function based on morphological characters revealed that, with the exception of the Macedonian (S. ucrainica-MA) and the Bulgarian-Romanian populations of S. ucrainica (S. ucrainica-BU-RU), all other individuals were assigned to their respective a priori-defined groups. The Macedonian individuals (S. ucrainica-MA) were correctly classified in 72.73% of cases, while 93.88% of the individuals of the Bulgarian-Romanian populations of S. ucrainica were correctly classified (Table 1).

Table 1.
The classification function based on morphological characters for six a priori-identified groups of “dasyphyllous” Stipa from the Balkan Peninsula.
The discriminant function analysis (DFA) showed that the most significant factors contributing to this divergence were the following characters: the length of the upper cauline leaves (LCL), ligule uppermost culm leaf length (LUL), dorsal row length (DL), distance from the end of the ventral line of the hairs to the top of the anthecium (DVL), and basal leaf-blade diameter (BASD) (Table 2, Figure 4).
Table 2.
Factor loadings of the first three principal component axes and discriminant function analysis (DFA) based on morphological characters for six a priori-identified groups of “dasyphyllous” Stipa from the Balkan Peninsula.
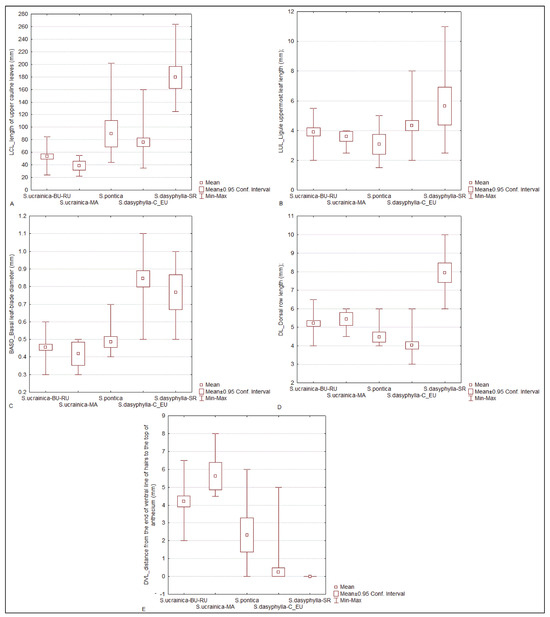
Figure 4.
Box-and-whisker plots showing variation in four most significant factors contributing to the divergence among morphological groups of “dasyphyllous” Stipa from the Balkan Peninsula revealed obtained by discriminant function analysis (DFA). (A) length of upper cauline leaves (LCL), (B) ligule uppermost culm leaf length (LUL), (C) dorsal row length (DL), (D) basal leaf-blade diameter (BASD), (E) distance from the end of ventral line of hairs to the top of anthecium (DVL). The whiskers represent extreme values, the boxes include 0.95% of confidence interval, and small squares indicate the mean values.
2.2. Microscope Observations
The most striking qualitative differences that were observed relate to the ornamentation of the leaf indumentum, i.e., the quality and appearance of the hairs on the abaxial leaf surface, and the leaf diameter. Indeed, the collected specimens marked as S. dasyphylla have a larger leaf diameter and are equipped with long soft hairs that make them soft to the touch (Figure 5D–F). The densest, longest, and most abundant leaf hairs were observed in specimens from Serbia, especially those from Mount Zlatibor (Figure 5A), followed by specimens from Mount Svrljiške (Figure 5E).

Figure 5.
Abaxial surface of the vegetative leaf blades of: (A) S. ucrainica—Bulgaria, (B) S. ucrainica—North Macedonia, (C) S. pontica—Greece, (D) S. dasyphylla—Slovakia, (E) S. stevanoviciorum subsp. pseudodasyphylla—Serbia: Svrljiške, (F) S. stevanoviciorum subsp. stevanoviciorum—Serbia: Zlatibor.
In contrast, the specimens of S. ucrainica and S. pontica had thinner leaves with more or less appressed, shorter hairs that were less abundant and not soft to the touch (Figure 5A–C).
2.3. Taxonomic Treatment
Taking all of our results into account, we provide a detailed description and map the distribution of a new species, S. stevanoviciorum, with two subspecies: S. stevanoviciorum subsp. stevanoviciorum and S. stevanoviciorum subsp. pseudodasyphylla subsp. nov. In the morphological descriptions, the value ranges correspond to the mean ± standard deviation, with the minimum and maximum values given in brackets.
Stipa stevanoviciorum Kabaš & D. Lakušić, sp. nova
Holotype: Serbia, Zlatibor Mt., Stublo, near Dubrava Monastery, 43.5806821 N, 19.6596977 E, 859 m, rocky grassland, serpentinite, 1 June 2021, Coll. E. Kabaš, P. Lazarević (BEOU 17763) (Figure 6).

Figure 6.
Holotype of S. stevanoviciorum subsp. stevanoviciorum.
Diagnosis: Stipa stevanoviciorum is most similar to S. dasyphylla, but differs in the following key characters: the culm ornamentation (pubescent vs. scabrous, rarely pubescent), the panicle basal internode surface (pubescent vs. scabrous, rarely pubescent), the upper sheaths ornamentation (glabrous to papilose vs. scabrous to pubescent), the basal leaf ligule margin (glabrous vs. ciliate to ciliolate, rarely glabrous), the length of the upper cauline leaves (125–264 mm vs. 35–160 mm), the length of the dorsal row of lemma hairs (6–10 mm vs. 3–6 mm); anthecium (22–25 mm vs. 19–22 mm), and several other morphological traits (Table 3).
Table 3.
Main differences between Stipa dasyphylla and subspecies of S. stevanoviciorum. The most significant diagnostic characters are given in boldface (* diagnostic between S. dasyphylla and S. stevanoviciorum, ** diagnostic between subspecies of S. stevanoviciorum). Quantitative continuous characters and ratios are reported as mean ± standard deviation, with the minimum and the maximum values in brackets.
Description: The plant is perennial, tufted, with a few culms and numerous vegetative shoots (Figure 7A,B), (22–)32–52(–59) cm tall, mostly four-noded, glabrous at nodes, and scabrous (the lower internodes) to more or less densely pubescent below them (the upper internodes). The leaves of the vegetative shoots: the sheaths of the external and internal leaves are mostly glabrous, the ligules are membranous and acute or slightly obtuse, the glabrous internal blades are convolute, the external blades are convolute to flat, and the leaves are green to pale green, are (39–)42–59(–70) cm long, are 1 mm in diameter, have an abaxial surface that is densely and softly hairy, and an adaxial surface pubescent or sparsely pilose. Cauline leaves: the sheaths are smooth to slightly papilose (usually in upper part of sheath), the ligules of the upper cauline leaves are (2.5-)2.9–8.3(-11) mm long, acute at the apex with very short cilia, and hairy on the back and margins, the blades are flat and are green or pale green, the uppermost culm is (12–)14–21(–27) cm long, and the abaxial surface is hairy and scabrous. The panicle is (7–)7–14(–18) cm long, exerted to partially enclosed, with (4–)5–10(–12) spikelets, branches with patent long and short hairs. The glumes are subequal and narrowly lanceolate, and the lower glume is (61–)66–92(–120) mm long while the upper glume is (54–)57–83(–110) mm long. The anthecium is (21–)22.1–24.3(–25.5) mm long, (1–)1.3–1.8(–2) wide, and callus with hairs, and the foot of the callus is curved, the peripheral ring is flattened (1–)1–1.3–1.5 × 0.5 mm, the lemma is straw-coloured, with seven lines of hairs, the dorsal and subdorsal lines are slightly fused at the base, the ventral line reaches the top of lemma, the dorsal line terminates (8–)9.3–11.6(–13) mm below the top of lemma length, the awn is (273–)295–364(–435) mm long, bigeniculate, the column is smooth and glabrous, twisted, brown or yellow, and (81–)88–101(–108) mm long, and the seta is (186–)204–265(–327) mm long and pilose, with (5–)5.3–6.8(–7.5) mm long hairs that gradually decrease in length towards the apex.

Figure 7.
Stipa stevanoviciorum subsp. stevanoviciorum on classical site on Zlatibor Mt. (A) habitus, (B) anthecium with callus, (C) basal leaves, (D) habitat (photograph by Predrag Lazarević.).
Etymology: the new species is named in honour of our teachers Prof. dr Vladimir Stevanović and Prof. dr Branka Stevanović, Serbian botanists known for their contributions to the study of Balkan plants, especially the Pontic flora and the vegetation of the Pannonian and Central Balkan rocky steppes.
Distribution and habitat: Stipa stevanoviciorum is an endemic taxon, known so far only from Mount Zlatibor in western Serbia and from Mount Svrljiške in eastern Serbia (Figure 8). It grows on a xerophilous rocky grassland in the mountain rocky steppe vegetation of the class Festuco-Brometea Br.-Bl. et Tx. ex Soó 1947. The plants from Zlatibor Mt. in western Serbia inhabit the ultramafic substrate and contribute to the vegetation composition of the rocky steppe vegetation of the order Halacsyetalia sendtneri Ritter-Studnička 1974 (Figure 7D), while the plants from Svrljiške Mt. in eastern Serbia grow on a carbonate rocky grassland that occurrs in the mountain steppe vegetation of the order Festucetalia valesiacae Soó 1947.
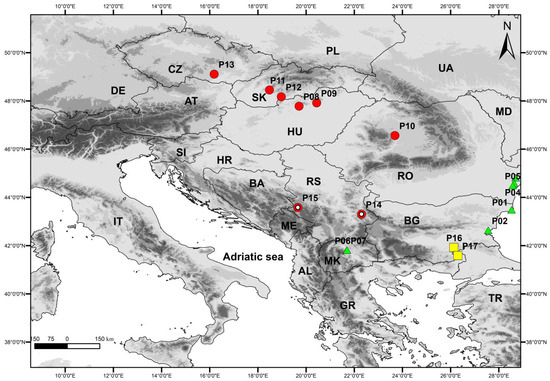
Figure 8.
Studied populations of Stipa ucrainica (green triangle), S. dasyphylla (red circles), S. stevanoviciorum (red circles with white dots), and S. pontica (yellow sqares). Data on populations can be found in Table 1.
Phenology: flowering and fruiting period: May–August.
Conservation status: At present, only about 100 flowering individuals have been recorded on Zlatibor Mt. in western Serbia, and approximately the same number of individuals have been recorded on Svrljiške Mt. in eastern Serbia, occupying a very small area of dry rocky grasslands. Due to its small population size and limited distribution, S. stevanoviciorum should be considered critically endangered (CR B2aci,ii,iii). The proximity of the road and the lack of grazing in that area, together with the closure of vegetation cover, may pose a threat to the survival of the very small populations of S. stevanoviciorum. The survival of these populations depends on the conservation of their habitats through national legislation, active in situ conservation (such as the preservation of open grasslandsand ex situ conservation measures, including seed banks and in vitro propagation.
Infraspecific Variability
Stipa stevanoviciorum subsp. stevanoviciorum
Description: The plant is perennial and tufted, with a few culms and numerous vegetative shoots (Figure 7), and is (35.3–)40.7–56.4(–59.5) cm tall, its blades are convolute to flat, silvery pale green, (417–)427–584(–698) cm long, and 1 mm in diameter, with an abaxial surface that is densely and softly hairy and an adaxial surface that is pubescent or sparsely pilose. Its panicle is (8.5–)9.2–16.9(–18.5) cm long and exerted, with (7–)7.2–10.8(–12) spikelets. Its glumes are subequal and narrowly lanceolate. Its anthecium is (21–)21.9–24.6(–25.5) mm long and callus with hairs, with a straw-coloured lemma that has seven lines of hairs, and its dorsal and subdorsal lines are slightly fused at the base, with the ventral line reaching the top of the lemma, the dorsal line terminating (9–)9.9–11.9(–13) mm below the top of lemma length, the awn being (273–)294.4–348.6(–357) mm long and bigeniculate, the column being smooth, glabrous, twisted, brown, and (81–)84.2–97.2(–102) mm long, and the seta being (186–)207–254(–268) mm long and pilose, with its hairs gradually decreasing in length towards the apex.
Distribution and habitat: Stipa stevanoviciorum subsp. stevanoviciorum is an endemic taxon, hitherto known only from Zlatibor Mt. in western Serbia and occurring on the ultramafic rocky grassland in the mountain rocky steppe vegetation of the order Halacsyetalia sendtneri at about 860 m. asl. (Figure 7C). The species occurs in a small area near the dirt road. The following species dominate the community at the classic site: Stipa pulcherrima C. Koch, Festuca valesiaca Schleicher ex Gaudin, Danthonia alpina Vest, Artemisia alba Turra, and Asperula purpurea (L.) Ehrend., while the following serpentinophyte species co-existing with them are the characteristic elements of the vegetation of the order Halacsyetalia sendtneri: Euphorbia glabriflora Vis., Halacsya sendtneri (Boiss.) Dörfl., Iris reichenbachii var. bosniaca Beck, Paragymnopteris marantae (L.) K.H. Shing, Potentilla visianii Pančić, Silene armeria L., Silene paradoxa L., Stachys recta subsp. baldaccii (K.Malý) Hayek, etc.
Phenology: flowering and fruiting period: May–August.
Conservation status: So far, just over 100 flowering individuals have been counted in a very small dry grassland area on Zlatibor Mt.; hence, the new subspecies should be considered as a critically endangered taxon (CR).
Stipa stevanoviciorum subsp. pseudodasyphylla Kabaš & D. Lakušić, subsp. nova
Holotype: Serbia, Svrljiške mountain, Gornji Rinj, 43.30267 N, 22.30041 E, 862 m, carbonate, 2 June 2023, Coll. E. Kabaš, V. Stanković, V. Đorđević (BEOU 71076) (Figure 9).

Figure 9.
Holotype of Stipa stevanoviciorum subsp. pseudodasyphylla.
Diagnosis: Similar to Stipa stevanoviciorum subsp. Stevanoviciorum, from which the subspecies differs mostly in the following characters: plant height (223–460 mm vs. 353–595 mm); number of flowers in a panicle (4–12 vs. 7–12); length of the upper cauline leaves (125–190 mm vs. 159–264 mm), length of the ligule of the uppermost culm leaf (6–11 mm vs. 2.5–4 mm; upper sheath ornamentation—glabrous to papilose vs. papilose; basal ligule length (2–4 mm vs. 1–2 mm); basal ligule tip—(cilate vs. ciliolate); type of basal leaves in sward—in some individuals only flat or only convolute leaves are present vs. both flat and convolute leaves are always present; colour of the column—yellow vs. brown; and somewhat denser, longer, and softer hairs on the abaxial side of the leaf blades (Table 3).
Description: The plant is perennial and tufted, with a few culms and numerous vegetative shoots (Figure 9), is (22.3–)28–42.4(–46) cm tall, has blades that are convolute to flat, green, (394–)412–604(–699) mm long, and 1 mm in diameter, has an abaxial surface that is densely and softly hairy, and has an adaxial surface that is pubescent or sparsely pilose. The panicle is (6.6–)7–11.8(–15.5) cm long and exerted, with (4–)4–8.6(–12) spikelets. The glumes are subequal and narrowly lanceolate. The anthecium is (21.5–)22.3–24(–24) mm long and callus with hairs, with a lemma that is straw-coloured, has seven lines of hairs, has dorsal and subdorsal lines that are slightly fused at the base, has a ventral line reaching the top of the lemma, has a dorsal line terminating (8–)8.9–11.3(–12) mm below the top of the lemma length, has an awn that (287–)296.8–378.4(–435) mm long, has a bigeniculate column that is smooth, glabrous, twisted, yellow, and (93–)93.2–103.4(–108) mm long, and has seta that are (193–)203–276(–327) mm long and pilose, with the hairs gradually decreasing in length towards the apex.
Etymology: The name of the subspecies agrees with the name of the closely related and very similar Stipa dasyphylla, to which group it undoubtedly belongs. However, the similar habitus together with some significant differences in certain morphological features led us to add the prefix pseudo- to the species attribute dasyphylla, as we consider it appropriate for the name of the new taxon.
Distribution and habitat: Stipa stevanoviciorum subsp. pseudodasyphylla is an endemic taxon that is only known from Svrljiške Mt. in eastern Serbia. The subspecies occurs in a small area of dry rocky grassland on carbonate rocks at several closely spaced microlocalities at an altitude of about 850 m. Approximately 100 individuals (cc. 30 flowering) occur within the provisory ass. Stipo-Festucetum rubrae.
Phenology: flowering and fruiting period: May–August.
Conservation status: Since only about 30 flowering individuals have been recorded so far in the Svrljiške Mt. in eastern Serbia, in a very small dry grassland area, the new subspecies should be considered as a critically endangered taxon (CR). However, considering the larger number of potential habitats of this plant in a wider area, the actual number of individuals is likely to be higher.
2.4. Herbarium Specimens Examined and Field Observations
Stipa stevanoviciorum subsp. stevanoviciorum
Serbia, Mt. Zlatibor, Stublo, near Dubrava Monastery, 43.580373 N, 19.65938 E, 770 m, rocky grassland, ultramafite, 13 June 2022, E. Kabaš, S. Vukojičić, P. Lazarević, det.: E. Kabaš, sub. S.dasyphylla (Lindem.) Trautv. (BEOU-KEGB 71001).
Serbia, Mt. Zlatibor, Stublo, near Dubrava Monastery, 43.581646 N, 19.660864 E, 790 m, rocky grassland, serpentinite, 27 September 2021, E. Kabaš, P. Lazarević, det.: E. Kabaš, sub. S.dasyphylla (Lindem.) Trautv.
Serbia, Mt. Zlatibor, Stublo, near Dubrava Monastery, 43.5806821 N, 19.6596977 E, 756 m, rocky grassland, serpentinite, 27 September 2021, E. Kabaš, P. Lazarević, det.: E. Kabaš, sub. S.dasyphylla (Lindem.) Trautv.
Stipa stevanoviciorum subsp. pseudodasyphylla
Serbia, agrest. Niša [surroundings of Niš], 1879, S. Petrović, MW0761191, det. Smirnov sub. S.dasyphylla Czern.
Serbia, Niševačka N 34, Stipa graffiana Pleš?, 1879, S. Petrović, 2039, det. Smirnov sub. S. dasyphylla Czern.
3. Discussion
The Stipa species belonging to the series Dasyphyllae represent a natural, clearly defined group within the section Stipa, whose adaxial leaves are hairy, making it easy to distinguish them from the other groups in the section. However, some doubts have been expressed regarding determining the interspecific relations between these taxa [34]. In [34], seven taxa are even listed within the so-called Stipa rubens complex, emphasising the need for a thorough revision of the localities where S. dasyphylla has previously been reported and pointing out that the presence of S. dasyphylla cannot be excluded from the area of south-eastern Europe. This statement proved to be correct in the light of the findings of our study. Moreover, Martinovský [34] considered the taxa of the S. dasyphylla group as phylogenetically young and morphologically characterized by minor but erratic, seamless distinctive features. This also seems to be the case with our new taxa from Serbia. S. stevanoviciorum is probably another of these phylogenetically young taxa that have diversified within the small, isolated relic islands that present the remnants of the true steppes in the study area. On the other hand, more recent research [32] has narrowed the group down to just two species, S. dasyphylla being one and S. zalesskii the other (including S. pontica and S. ucrainica on the subspecies level). Either way, it is an uncontroversial opinion that S. dasyphylla has always been considered a good taxon distinguished from the related taxa by its unique characteristics. These very properties, such as hairy silvery leaves that are soft to the touch, lead us to think that populations from Serbia fall within the variety of S. dasyphylla [40]. However, a closer examination of the herbarium material and especially of the plants in the field, which include S. dasyphylla and related taxa in the wider area of Serbia, North Macedonia, Greece, Bulgaria, Romania, Hungary, Slovakia, and the Czech Republic, led us to the conclusion that the plants from Serbia are clearly different from what was considered typical for S. dasyphylla, as well as from other representatives of the series Dasyphyllae. This was later confirmed by the results of our morphometrica analyses. PCA showed a high degree of separation of Serbian populations of S. dasyphylla, while “typical” populations from Central Europe were associated with the remaining populations of S. ucrainica and S. pontica from North Macedonia, Greece, Bulgaria, and Romania. Moreover, DCA analysis and UPGMA cluster analysis not only confirmed the results of the PCA, but also showed significant morphological differentiation between two Serbian populations. The extent and quality of the morphological differences, together with a very pronounced disjunct distribution, provided us with sufficient arguments to split the populations from Serbia into a new species, S. stevanoviciorum, with two subspecies. The most significant differences that are recognisable with the naked eye and under the microscope concern the ornamentation of the leaf blade and the habit. The specimens collected in Serbia which correspond to S. stevanoviciorum are more robust, have a larger leaf diameter, and are equipped with dense, long, and tender hairs, which makes them soft to the touch. Nevertheless, at first glance, S. stevanoviciorum and its subspecies can still easily be confused with S. dasyphylla, to which it is the most closely related. However it differs from the latter by a number of qualitative characters: by having pubescent culms, a pubescent basal internode, a pubescent panicle basal internode, glabrous to papillose upper sheaths, mostly both flat and convolute leaves present in the tuft, as well as the length of the upper cauline leaves (125–264 mm vs. 35–160 mm), the length of the dorsal row of lemma hairs (6–10 mm vs. 3–6 m), the distance from the end of the ventral line of hairs to the top of the anthecium (0–0 vs. 0–5), the length of the hairs on the seta (5–7.5) mm vs. (3.5–6 mm), and the length ratios of different characters (Table 3).
Although the cluster analysis revealed considerable morphological divergence between the two newly proposed subspecies, we have applied the concept of subspecies here for the following reasons. First, the results of the PCA and CDA show that the overlaps on the first two principal component and discriminant axes are very pronounced, as well as that a slight separation occurs only on the third CDA axis, which describes only 6.84% of the total discrimination. In addition, most of the quantitative characters overlap, and only a few characters show a significant tendency towards separation. At the same time, these two groups of populations (i.e., the newly proposed subspecies) show a very strong ecological and geographical differentiation. The fact that, in addition to subtle morphological differences, the populations from western and eastern Serbia also differ geologically (carbonate vs. ultramafic substrate), climatically (humid temperate vs. dry subcontinental climate), and historically–biogeographically (Illyrian vs. Moesian phytoprovince) is, in our opinion, a strong argument in favour of giving them each a separate taxonomic status at the subspecies level.
Key to the taxa:
1a. Leaves always convolute, 0.5–0.9 mm in diameter, abaxial surface aculeolate and with sparse, subappressed hairs.........................................….……………….………...………2
1b. Leaves flat or convolute, when convolute diameter c. 1 mm, with dense, long hairs, soft to the touch..................................................................................................……….…3
2a. Ventral line of hairs on lemma ending 2–6 mm below the insertion of the awn, sheaths mostly scabrid, leaf diameter c. 0.5 mm.....................................................S. ucrainica
2b. Ventral lines of hair on lemma ± reaching the insertion of the awn, sheaths glabrous, leaf diameter 0.5–0.9 mm..................………………….……….……................S. pontica
3a. Anthecium 19–24 mm, length of the dorsal row of lemma hairs 6–10 mm, length of upper cauline leaves 35–160 mm.........................................................................S. dasyphylla
3b. Anthecium 22–25 mm, length of the dorsal row of lemma hairs 3–6 mm, length of upper cauline leaves 125–264 mm................................................................................................4
4a. Plant height 40–57 cm, number of flowers in panicle 7–10, length of the culm leaf ligule 2.5–4 mm, ligule tip ciliolate, column brown......................................S. stevanoviciorum subsp. stevanoviciorum
4b. Plant height 28–42 cm, number of flowers in panicle 4–8, length of the uppermost culm leaf ligule 6–8 mm, ligule tip ciliate, column yellow........................S. stevanoviciorum subsp. pseudodasyphylla
4. Materials and Methods
Plant Material and Morphometric Analyses
Morphometric analyses were performed on the material collected in the field as well as on herbarium specimens deposited in BEOU, BEO, and MW (herbarium abbreviations follow [40]). In addition, various virtual herbaria and GBIFs were reviewed (see Appendix A for links).
The morphometric study included 160 individuals from 17 populations belonging to three “dasyphyllous” Stipa species (S. pontica, S.ucrainica, and S.dasyphylla) collected in the central and eastern Balkan Peninsula and Central Europe (Table 4). In this study, we included taxa previously classified in the S. dasyphylla group, as outlined in Flora Europaea [7], which are characterised by a densely hairy adaxial leaf surface and an abaxial surface with conical papillae that are often accompanied by 0.5–1 mm-long hairs, and which occur in Europe.
Table 4.
Studied populations of Stipa ucrainica, S. dasyphylla, and S. pontica.
From each population, from 6 to 15 plant samples were used for morphometric analyses. The voucher specimens were deposited in the Herbarium of the Institute of Botany and Botanical Garden, of the Faculty of Biology, University of Belgrade (BEOU). The morphological characteristics were observed and measured using a stereo microscope Leica MZ 75 and photographed using Leica DFC 295 camera with Leica Application Suite (LAS) Version 4.12.0.
A total of 37 morphological characters were measured (24 quantitative and 13 qualitative), which are listed in Table 5. Descriptive statistics (minimum and maximum values, mean, standard deviation, and coefficient of variation) were calculated for each quantitative morphometric characteristic measured. Prior to all multivariate analyses, the morphometric data were transformed using Box-Cox transformation in PAST 4.17 [41]. In order to eliminate a single characteristic from highly correlated pairs, pairwise Spearman correlation analyses were performed, and four characters with a Spearman correlation coefficient greater than 0.80 were excluded from the multivariate analysis. Different multivariate approaches were used to compare all characters and groups assessed, including principal component analysis (PCA), canonical discriminant analysis (CDA), and cluster analysis using Statistica v.8.0 (StatSoft and Inc 2007). Principal component analysis (PCA) was performed to reveal the overall morphological variation and relationships between individuals in all populations. Canonical discriminant analysis (CDA) was used to test the hypothesis of morphological segregation of six groups of individuals identified a priori as potential OTUs based on their taxonomic and biogeographic affiliations. S. ucrainica is divided into two groups: S. ucrainica-BU-RU (Bulgaria and Romania) from the Danube phytoprovince and S. ucrainica-MA (North Macedonia) from the Macedonian phytoprovince. S. dasyphylla is divided into three groups: S. dasyphylla-C-EU (Central Europe) from the Pannonian phytoprovince, S. dasyphylla-SR-W (Serbia, western) from the Illyrian phytoprovince, and S. dasyphylla-SR-E (Serbia, eastern) from the Balkan (Moesian) phytoprovince. Finally, S. pontica from the Thracian phytoprovince is defined as a separate group (Table 4). The biogeographical affiliation follows the phytogeographical map of Europe [42], in which each phytoprovince represents a unit defined by historical, ecological, and geographical barriers, each with different floristic and ecosystem characteristics (Table 4). Finally, the UPGMA (unweighted pair group method with arithmetic mean) cluster analysis based on Mahalanobis distances between all analysed groups was computed, the classification function was used to determine the percentage of correctly classified individuals in each group, and discriminant function analysis (DFA) was used to estimate the contribution of individual characters to the overall differentiation. The threatened status of S. stevanoviciorum in Serbia was assessed using the IUCN criteria [43].
Table 5.
Morphological characters measured in this study. Characters with asterisk (*) are included in the multivariate analysis; highly correlated characters excluded from the multivariate analysis are marked with two asterisks (**).
Author Contributions
Conceptualization, E.K. and D.L.; methodology, E.K., D.L. and J.N.; validation, E.K., D.L. and S.V.; formal analysis, E.K. and D.L.; investigation, E.K., P.L., V.S. and S.V.; data curation, E.K., V.S., S.V. and P.L.; writing—original draft preparation, E.K. and D.L.; writing—review and editing, E.K., D.L., S.V., V.S., P.L. and J.N.; visualization, E.K., D.L., P.L. and J.N.; funding acquisition, E.K., D.L., S.V., P.L. and J.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Science Fund of the Republic of Serbia, grant number 7750112—Balkan biodiversity across spatial and temporal scales—patterns and mechanisms driving vascular plant diversity (BalkBioDrivers), by the Serbian Ministry of Education, Science and Technological Development (project no. 451-03-68/2022-14/200178), and the State Enterprise for Management Forest “Srbijašume”, Forest Estate “Užice”—Protected Plants Monitoring Project.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
We thank Dragana Rabrenović for the help with the arranging and scanning of the type material and Vladan Djordjević for the help on the field and for the useful suggestions during the manuscript preparation and submission. We are also grateful to the Reviewers who had kindly reviewed the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| BEOU | University of Belgrade |
| BEO | Natural History Museum |
| MW | Moscow State University |
| GBIF | Global Biodiversity Information Facility |
| IUCN | International Union for Conservation of Nature |
Appendix A
Table A1.
The links to virtual herbaria consulted for the purposes of this study.
Table A1.
The links to virtual herbaria consulted for the purposes of this study.
References
- Soreng, R.J.; Peterson, P.M.; Romschenko, K.; Davidse, G.; Zuloaga, F.O.; Judziewicz, E.J.; Filgueiras, T.S.; Davis, J.I.; Morrone, O. A worldwide phylogenetic classification of the Poaceae (Gramineae). J. Syst. Evol. 2015, 53, 117–137. [Google Scholar] [CrossRef]
- Jacobs, S.; Bayer, R.; Everett, J.; Arriaga, M.; Barkworth, M.; Sabin-Badereau, A.; Torres, A.; Vázquez, F.; Bagnall, N. Systematics of the tribe Stipeae (Gramineae) using molecular data. Aliso 2007, 23, 349–361. [Google Scholar] [CrossRef][Green Version]
- Roshevitz, R.Y. Stipa. In Flora URSS 2; Komarov, V.L., Ed.; Academiae Scientiarum URSS: Sankt Petersburg, Russia, 1934; pp. 66–93. [Google Scholar]
- Tzvelev, N.N. Zlaki (Gramineae). In Rastieniya Centralnoi Azii. Po Materialam Botanicheskogo Instituta im. Komarova VL (Plantae Asiae Centralis, Secus Materies Instituti Botanici Nomine Komarovii VL); Grubov, V.I., Ed.; Nauka: Leningrad, Russia, 1968; Volume 4, pp. 1–243. [Google Scholar]
- Tzvelev, N.N. Zlaki SSSR; Nauka: Leningrad, Russia, 1976. [Google Scholar]
- Bor, N.L. Stipa L. In Flora des Iranischen Hochlandes und der Umrahmenden Gebirge: Persien, Afghanistan, Teile Von West-Pakistan, Nord-Iraq, Azerbaidjan, Turkmenistan 70; Rechinger, K.H., Ed.; Akademische Druck: Graz, Austria, 1970; pp. 395–410. [Google Scholar]
- Martinovský, J.O. Stipa. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Chater, A.O., Richardson, I.B.K., Eds.; Cambridge University Press: Cambridge, UK, 1982; Volume 5, pp. 247–252. [Google Scholar]
- Freitag, H. The genus Stipa (Gramineae) in southwest and south Asia. Notes R. Bot. Gard. Edinb. 1985, 42, 355–489. [Google Scholar]
- Wu, Z.L.; Phillips, S.M. Tribe Stipeae. In Flora of China; Wu, Z.Y., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2006; Volume 22, pp. 188–212. [Google Scholar]
- Barkworth, M.E.; Arriaga, M.O.; Smith, J.F.; Jacobs, S.W.; Valdés-Reyna, J.; Bushman, B.S. Molecules and morphology in South American stipeae (Poaceae). Syst. Bot. 2008, 33, 719–731. [Google Scholar] [CrossRef]
- Nobis, M. Taxonomic revision of the Stipa lipskyi group (Poaceae: Stipa section Smirnovia) in the Pamir Alai and Tian Shan Mountains. Pl. Syst. Evol. 2013, 299, 1307–1354. [Google Scholar] [CrossRef]
- Nobis, M.; Gudkova, P.D.; Nowak, A.; Sawicki, J.; Nobis, A. A Synopsis of the Genus Stipa (Poaceae) in Middle Asia, Including a Key to Species Identification, an Annotated Checklist, and Phytogeographic Analyses1. Ann. Mo. Bot. Gard. 2020, 105, 1–63. [Google Scholar] [CrossRef]
- Kotukhov, Y.A. New species of grasses (Poaceae) from south Altai, Saur and Tarbagatai. Turczaninowia 1998, 1, 7–21. [Google Scholar]
- Kotukhov, Y.A. New species of the genus Stipa L. (Poaceae) from western Kazakhstan. Turczaninowia 1998, 1, 9–15. [Google Scholar]
- Noltie, H.J. Notes relating to the flora of Bhutan: XXXVIII. Gramineae I, tribe Stipeae. Edinb. J. Bot. 1999, 56, 285–291. [Google Scholar] [CrossRef]
- Vázquez Pardo, F.M.; Ramos, S. Two new taxa and a new combination for Stipa (Gramineae: Stipeae) in Tunisia. Bot. J. Linn. Soc. 2007, 153, 439–444. [Google Scholar] [CrossRef]
- Vázquez Pardo, F.M.; Perez-Chiscano, J.L.; Gutiérrez, M.; Ramos, S. A new species of Stipa sect. Leiostipa (Poaceae) from SW Spain. Willdenowia 2009, 39, 261–264. [Google Scholar] [CrossRef][Green Version]
- Nobis, M.; Nowak, A.; Nobis, A. Stipa zeravshanica sp. nov. (Poaceae), an endemic species from rocky walls of the western Pamir Alai Mountains (middle Asia). Nord. J. Bot. 2013, 31, 666–675. [Google Scholar] [CrossRef]
- Nobis, M.; Ebel, A.L.; Nowak, A.; Turginov, O.T.; Kupriyanov, A.N.; Nobis, A.; Olonova, M.V.; Paszko, B.; Piwowarczyk, R.; Chen, W.-L.; et al. Contribution to the flora of Asian and European countries: New national and regional vascular plant records, 2. Acta Bot. Gall. 2014, 161, 209–221. [Google Scholar] [CrossRef]
- Nobis, M.; Nobis, A.; Klichowska, E.; Nowak, A.; Nowak, S.; Gudkova, P.D. Stipa dickorei sp. nov. (Poaceae), three new records and a checklist of feather grasses of China. Phytotaxa 2016, 267, 29–39. [Google Scholar] [CrossRef][Green Version]
- Vázquez Pardo, F.M.; Gutiérrez Esteban, M. Calssification of species of Stipa with awns having plumose distal segments. Telopea 2011, 13, 155–176. [Google Scholar] [CrossRef][Green Version]
- Zhao, L.Q.; Guo, K. Stipa albasiensis (Poaceae), a new species from Inner Mongolia, China. Ann. Bot. Fenn. 2011, 48, 522–524. [Google Scholar] [CrossRef]
- Cataldo, D.; Giardina, S.A.; Moraldo, B.; Raimondo, F.M. Stipa valdemonensis (Poaceae), a new species from Sicily. Plant Biosyst. 2012, 146, 658–663. [Google Scholar] [CrossRef]
- Tzvelev, N.N. Notes on the tribe Stipeae Dumort. (Poaceae). Nov. Sist. Vyss. Rastenii 2012, 43, 20–29. [Google Scholar] [CrossRef]
- Tzvelev, N.N. On some hybridogenous taxa in the genus Stipa L. (Poaceae). Nov. Sist. Vyss. Rastenii 2014, 45, 5–8. [Google Scholar] [CrossRef]
- Smirnov, P.A. Die neuen russischen Stipa-Pennata-Arten. Repert. Nov. Specierum Regni Veg. 1925, 21, 231–235. [Google Scholar]
- Martinovský, J.O. Kavyly serie Pulcherrimae na Slovensku. Biológia 1965, 7, 498–510. [Google Scholar]
- Martinovský, J.O. Über drei neue Stipa-Sippen aus dem Verwandtschaftskreis Stipa joannis s. l. XXII. Beitrag zur Kenntnis der Stipa-Sippen. Oesterreichische Bot. Z. 1970, 118, 171–181. [Google Scholar]
- Martinovský, J.O. Clavis analytica nec non descriptiones breves taxorum generis Stipa in Europa centrali provenientium. Preslia 1977, 49, 97–113. [Google Scholar]
- Klokov, M.; Osychnyuk, V. Stipae Ucrainicae. Novosti Sist. Vyss. Nizsh. Rast. 1976, 1975, 7–91. [Google Scholar]
- Tzvelev, N.N. On the feather-grasse (Stipa L., Gramineae) in the Ukraine. Byuletin Mosk. Obs. Ispyt. Prirody. Otd. Biol. 1986, 91, 116–124. [Google Scholar]
- Gonzalo, R.; Aedo, C.; García, M.A. Taxonomic revision of Stipa subsection Pulcherrimae (Poaceae). In Taxonomía y Sistemática de Stipa Subsections Stipa L. (Poaceae) y Grupos Afines; Gonzalo, R., Ed.; Universidad Complutense de Madrid: Madrid, Spain, 2013; pp. 136–216. [Google Scholar] [CrossRef]
- Nobis, M.; Klichowska, E.; Nowak, A.; Gudkova, P.D.; Rola, K. Multivariate morphometric analysis of the Stipa turkestanica group (Poaceae: Stipa sect. Stipa). Plant Syst. Evol. 2016, 302, 137–153. [Google Scholar] [CrossRef]
- Martinovský, J.O. Taxonomische Studie über die Stipa-Serie Dasyphyllae. XXV Beitrag zur Kenntnis der Gattung Stipa. Preslia 1975, 47, 249–261. [Google Scholar]
- Gudkova, P.D.; Yu Korolyuk, A.; Ryzhakova, D.D.; Kriuchkova, E.A.; Nobis, M. The genus Stipa (Poaceae) in the Altai Krai: Taxonomy and distribution. Turczaninowia 2020, 23, 32–49. [Google Scholar] [CrossRef]
- Apostolova, I.; Petrova, A.S.; Meshinev, T.; Danihelka, J. Stipa ucrainica (Poaceae): A recently recognized native species of the Bulgarian flora. Phytol. Balcan. 2008, 14, 257–262. [Google Scholar]
- Kabaš, E.; Niketić, M.; Ćušterevska, R.; Tomović, G.; Vukojičić, S.; Lakušić, D. Rare and threatened Stipa ucrainica (Poaceae) found in Macedonia—A new Pontic disjunction in the Balkans. Phyton Ann. Rei. Bot. 2017, 57, 37–45. [Google Scholar] [CrossRef]
- Valdés, B.; Scholz, H.; Raab-Straube, E.V.; Parolly, G. Poaceae (Pro Parte Majore). Euro+ Med Plantbase-The Information Resource for Euro-Mediterranean Plant Diversity. 2009. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 1 August 2021).
- Kabaš, E.; Vukojičić, S.; Aćić, S.; Lakušić, D. Phytosociology of Stipa-dominated steppe-like vegetation on ultramafics of Central Balkans. Bot. Serb. 2022, 46, 17–27. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. 2021. Available online: https://sweetgum.nybg.org/ih/ (accessed on 1 August 2021).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Meusel, H.; Jäger, J.E. Vergleichende Chorologie der Zentraleuropäischen Flora; Karten. Bd. 3.; Gustav Fischer: Stuttgart, Germany, 1992; pp. 422–688. [Google Scholar]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; IUCN Species Survival Commission: Cambridge, UK, 2012. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).